Introduction
Local ischemia and hypoxia following myocardial
infarction results in the loss of cardiomyocytes and tissue damage
(1). Stem cell therapy for
myocardial regeneration is a promising treatment and the
development of resident cardiac stem cell (CSC) therapy provides a
novel option for treating myocardial infarction (2). Sca-1 (stem cells antigen-1) is a
mouse protein that is commonly used to identify CSCs and these CSCs
are capable of electrical coupling with local cardiomyocytes by
differentiation into functional cardiomyocytes (3,4).
Following myocardial infarction, CSCs migrate into the
peri-ischemic and ischemic areas of the heart via cytokine-induced
chemotaxis and participate in myocardial regeneration by
differentiating into new cardiomyocytes, endothelial cells or
smooth muscle cells (5–7).
The migratory ability of CSCs is important for
successful implantation and subsequent amelioration of function
during myocardial regeneration (8). Multiple signaling pathways
participate in CSC migration and recruitment. The
phosphatidylinositol 3-kinase/protein kinase B (PI3K/Akt) signaling
pathway has been demonstrated to be essential for stem cell
functions, including cell migration, cell survival and apoptosis
(9). Stromal cell-derived factor 1
(SDF-1) is also a key factor in cardiac stem cell recruitment: By
binding to a specific G protein coupled receptor on the cell
membrane, C-X-C chemokine receptor type 4 (CXCR-4), SDF-1 induces
chemotaxis and homing of several progenitor cells including CSCs,
endothelial progenitor cells and mesenchymal stem cells (10). Elevated SDF-1 expression in the
heart following infarction is correlated with increased CSC
recruitment and improved myocardial regeneration (11). Successful migration of CSCs to the
site of infarction or ischemic myocardium is important for cardiac
regeneration, with poor migration of CSCs limiting the potential of
cell therapy (12). Exogenous
delivery of cell migration-enhancing cytokines may overcome this
limitation and improve the efficacy of stem cell therapy for
myocardial regeneration (13).
Basic fibroblast growth factor (bFGF) is an
important molecule involved in heart remodeling and regeneration
following myocardial infarction (14). In the myocardial regeneration
process, bFGF stimulates the proliferation and migration of
fibroblasts and participates in the formation of collagen to
maintain heart function following loss of cardiomyocytes (15). bFGF may improve ventricular
remodeling by regulating the relative proportions of collagen I and
III (16). bFGF also stimulates
the migration and proliferation of vascular endothelial cells and
smooth muscle cells, and is viewed as a multipotent angiogenic
stimulus that is important for tissue regeneration (17). Following myocardial infarction, the
expression of bFGF in cardiomyocytes, vascular endothelial cells
and smooth muscle cells in the ischemic and peri-ischemic area is
elevated, improving cell migration, increasing the local blood
supply and subsequently ameliorating cardiac function (18,19).
The adult human heart is recognized as a
self-renewing organ as it contains its own native stem cells, CSCs
(20). These CSCs are able to
renew themselves, differentiating into new cardiomyocytes,
endothelial cells and smooth muscle cells to replace injured or
dead cells, under the stimulation of cytokines such as vascular
endothelial growth factor (VEGF), bFGF and hypoxia-inducible
factors (HIF) (4,5). Following myocardial infarction,
activated CSCs migrate from their in situ residence and
gather in ischemic and peri-ischemic areas to participate in tissue
regeneration (8). Cell therapy
based on CSCs provides a novel treatment for myocardial infarction
and subsequent heart failure (21,22).
There are three main subgroups of CSCs in the heart, of which the
Sca-1+ group is the most numerous. It has previously
been demonstrated that the number of Sca-1+ cells is
~100 fold greater than the other two subgroups. Sca-1+
CSCs display three central characteristics: Multipotency,
self-renewal and clone formation ability (23).
There is previous evidence that in a murine
myocardial infarction model, intra-myocardium injection of
Sca-1+ CSCs reduced infarction size, attenuated left
ventricular remodeling and ameliorated cardiac function (24). However, CSC therapy is limited by
insufficient numbers of transplanted cells to replace the numerous
lost cardiomyocytes (25,26). In order to solve this problem, it
is important to induce resident CSC migration into the
peri-ischemic and ischemic areas.
The present study aimed to investigate the effect of
exogenously delivered bFGF on the migratory ability of CSCs in
vitro and in vivo, which is important for CSC-based
therapy for myocardial infarction.
Materials and methods
Ethics statement
Animal studies were approved by Soochow University
Scientific and Animal Ethics Committee (approval no. 20120055) and
were in compliance with Chinese national regulations on the use of
experimental animals. Procedures for animal studies were performed
in accordance with the Guide for the Care and Use of Laboratory
Animals (revised in 1996) by the US National Institutes of Health
(Bethesda, MD, USA).
Isolation and culture of
Sca-1+CSCs
The present study used 6-week old, male C57/BL6 mice
(weight, 15–20 g; n=20). All animals were purchased from the
Laboratory Animal Center of Soochow University (Suzhou, China).
They were maintained on standard diet and water with a 12-h
light/dark cycle, room temperature and 50% humidity at the animal
center of The First Affiliated Hospital of Soochow University.
CSCs were prepared following euthanasia with carbon
dioxide. C57/BL6 mouse hearts were minced and digested with 0.1%
collagenase B (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and
0.2% trypsin (Invitrogen; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) at 3°C for 2 h. Cells were selected using a magnetic
selection system (Miltenyi Biotec, Inc., Auburn, CA, USA). The
purity of isolated cells was verified by flow cytometry. Isolated
CSC were cultured in Dulbecco's modified Eagle's medium/nutrient
mixture F12 (DMEM/F12) (Invitrogen; Thermo Fisher Scientific, Inc.)
with 10% fetal bovine serum (Gibco; Thermo Fisher Scientific,
Inc.), 10 ng/ml leukemia inhibitory factor (LIF; PeproTech, Inc.,
Rocky Hill, NJ, USA), 10 ng/ml cardiotroponin (PeproTech, Inc.), 10
ng/ml epidermal growth factor (EGF; PeproTech, Inc.) and a 1%
solution of penicillin and streptomycin. The medium was replaced
every three days.
Cell migration assay in vitro
A Transwell migration assay was used to measure cell
migration in vitro. CSC were pre-washed with serum-free
DMEM/F12 medium and 5×104 cells in 200 µl serum-free
DMEM/F12 medium were seeded into the upper chamber of a 24 well
Transwell plate (Merck KGaA). DMEM/F12 (600 µl) complete medium
with different concentrations of bFGF, or bFGF plus the Akt
inhibitor deguelin, was added to the lower chamber. Following
incubation for 24 h, the upper surface of the membrane was scraped
with a cotton swab to remove un-migrated cells. Cells migrating to
the lower surface of the membrane were stained at room temperature
for 20 min with 0.1% hexamethylpararosaniline. Each assay was
performed in triplicate wells and repeated three times. The number
of cells migrating to the lower surface of the membrane was counted
in 10 random ×200 fields using an inverted microscope and the mean
number of cells per field was used for statistical analysis. The
Migration Index was calculated as Migration cell number
(treated)/Migration cell number (untreated).
Myocardial infarction construction and
cell transplantation
Myocardial infarction was simulated through surgical
ligation of the left anterior descending (LAD) coronary artery with
a 7/0 prolene suture (Ethicon, Inc., Somerville, NJ, USA) and
successful ligation was verified through observation of a color
change with the naked eye from red to white in the infarct area. 30
wild-type specific pathogen free C57/BL6 mice (8–10 weeks) were
randomly assigned to three groups: CSC, bFGF-treated CSC (45 ng/ml)
and bFGF-treated CSC + Akt inhibitor (5.0 mg/kg oral Akt inhibitor
for 3 days prior to transplantation). For cell transplantation,
animals received intra-myocardium injections of 1×106
cell solution immediately following LAD ligation. All animals were
sacrificed one week following cell transplantation. A further 6
mice underwent LAD ligation without cell transplantation. bFGF
protein expression and Sca-1+ cell aggregation was
measured in these mice one week following simulated myocardial
infarction.
Cell migration and maintenance
assessment in vivo
Transplanted cells were labeled with the cell
tracking dye CM-Dil (Invitrogen; Thermo Fisher Scientific, Inc.).
Prior to cell transplantation, 5 µl CM-Dil was added to
1×106 cells suspended in 20 µl PBS and incubated at room
temperature for 20 min. Successful labeling was confirmed by flow
cytometry (BD FACSCalibur; BD Biosciences, Franklin Lakes, San
Jose, CA, USA) and immunofluorescence microscopy. One week
following transplantation, frozen tissue was obtained and sectioned
into three continuous 2 µm sections. Cells maintained in the
peri-infarct area and migrated to the infarct area were visualized
as green and counted in 10 random ×200 fields using
immunofluorescence microscopy for 10 sections. The mean number of
cells per field was obtained for statistical analysis.
Western blot analysis
The harvested cells or minced tissues were lysed in
a lysis buffer (Roche Diagnostics, Basel, Switzerland) at 37°C for
2 h. Protein concentrations were quantified using a bichoninic acid
protein assay kit (Pierce; Thermo Fisher Scientific, Inc). 30 µg
proteins were separated using 10% SDS-polyacrylamide gel
electrophoresis (SDS-PAGE) and transferred onto a polyvinylidene
difluoride (PVDF) membrane (EMD Millipore, Billerica, MA, USA).
Following blocking the membrane with Tris-buffered saline + 0.1%
Tween 20 (TBST) and 5% nonfat dried milk for 2 h at room
temperature, the membrane was washed twice with TBST, then
incubated with the primary antibody (Akt catalog no. ab 64148;
1:1,000; p-Akt catalog no. ab 38449; 1:1,000; bFGF catalog no. ab
168328; 1:1,500; CXCR-4 catalog no. ab1670; 1:1,000; SDF-1 catalog
no. ab18919; 1:1,000; GAPDH catalog no. ab9485; 1:2,000; Abcam,
Cambridge, UK) overnight at 4°C. Next, the membrane was washed with
TBST, then incubated with the peroxidase-conjugated secondary
antibody (catalog no. A21010 1:500; Wuhan Amyjet Scientific Co.,
Ltd., Wuhan, China). Bands were detected using a chemiluminescence
western blot detection system (Pierce; Thermo Fisher Scientific,
Inc.). The protein expression and phosphorylation levels were
normalized to baseline expression and glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) levels, respectively.
Immunohistochemistry
Hearts were fixed with 10% buffered formalin
solution overnight, dehydrated in graded alcohols and embedded in
paraffin. The tissue was subsequently cut into 2 µm thick sections,
after blocking with Tris buffered saline Tween-20 (TBST) and 5%
nonfat dried milk for 2 h, tissue sections were incubated at room
temperature for 2 h with antibodies against bFGF (catalog no.
ab168328; 1:500; Abcam). The Image-Pro Plus 6.0 (Media Cybernetics,
Inc., Rockville, MD, USA) was used to evaluate the positive
expression ratio of bFGF.
Capillary density assessment
10% Formalin-fixed and paraffin-embedded tissue
sections were sliced transversally into 2 µm sections and incubated
with antibodies against von Willebrand factor (vWF) at room
temperature for 2 h (catalog no. ab9378; 1:3,000; Abcam). Tissue
was then washed twice with TBST and incubated with
peroxidase-conjugated secondary antibodies (goat anti mouse;
catalog no. A996702; 1:1,000; Wuhan Amyjet Scientific Co., Ltd.).
Positive vessels were stained brown. The mean number of capillaries
per field in the infarct myocardium was obtained by counting in 10
randomly selected ×200 fields for statistical analysis for 10
sections.
Statistical analysis
Data were expressed as the mean ± standard deviation
and analyzed with SPSS 17.0 statistical software (SPSS, Inc.,
Chicago, IL, USA). Multiple comparisons were performed through
one-way analysis of variance tests and Scheffe's post hoc tests
when data were normally distributed, and through Kruskal-Wallis
tests and Dunn's post hoc tests when data were not normally
distributed. P<0.05 was considered to indicate a statistically
significant difference.
Results
bFGF expression increases and
Sca-1+ CSCs aggregate following myocardial
infarction
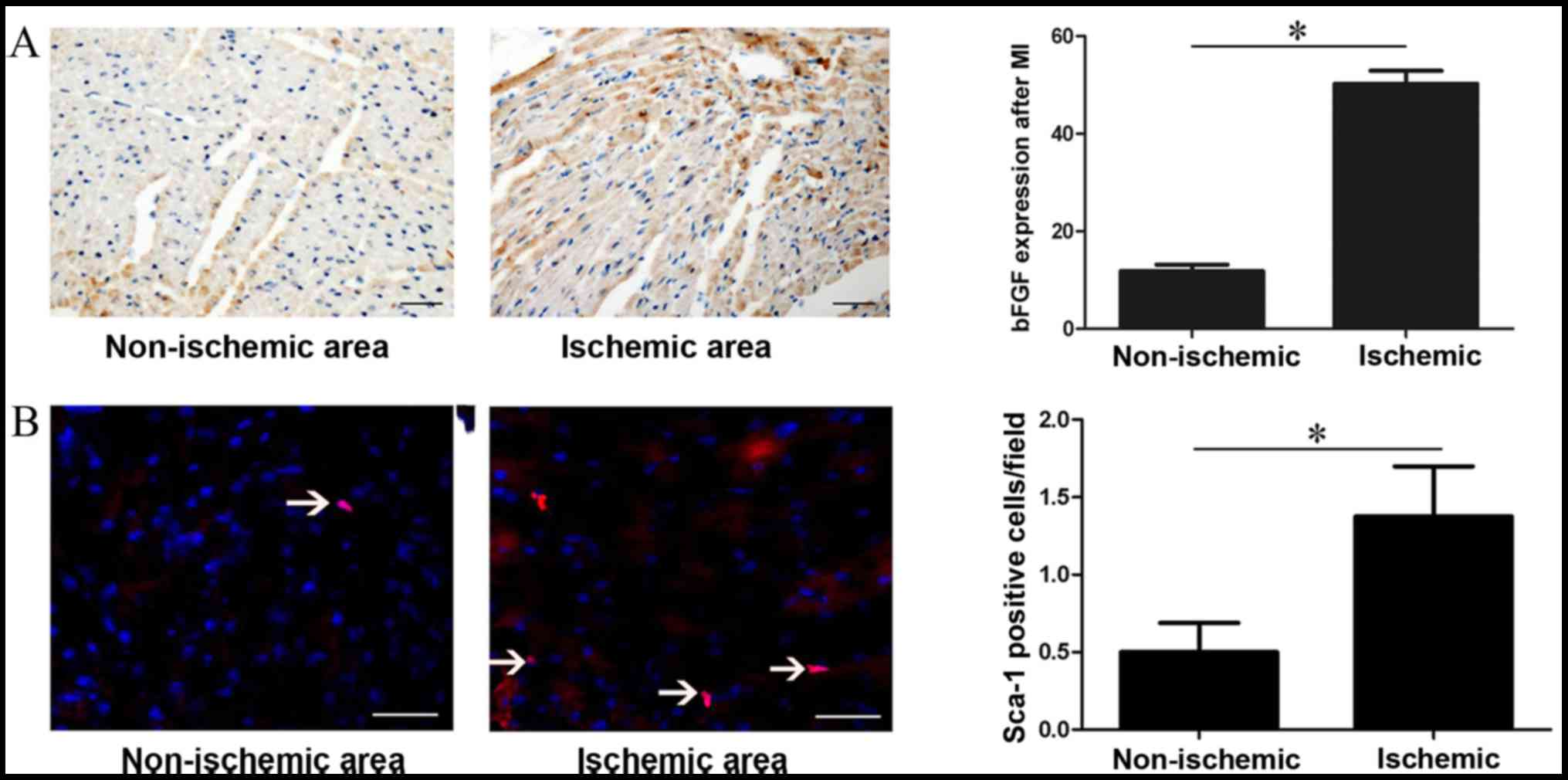
The expression of bFGF was analysed 1 week after
myocardial infarction simulation. Immunohistochemical analysis
revealed that bFGF expression was 50.3±2.61 in the ischemic area
and 11.83±1.38 in the non-ischemic area (P=0.001; Fig. 1A). Immunofluorescence analysis
demonstrated that regional ischemia following myocardial infarction
resulted in increased recruitment of Sca-1+ CSCs in the
ischemic area compared with the non-ischemic area (0.5±0.53 vs.
1.38±0.92, P=0.001; Fig. 1B).
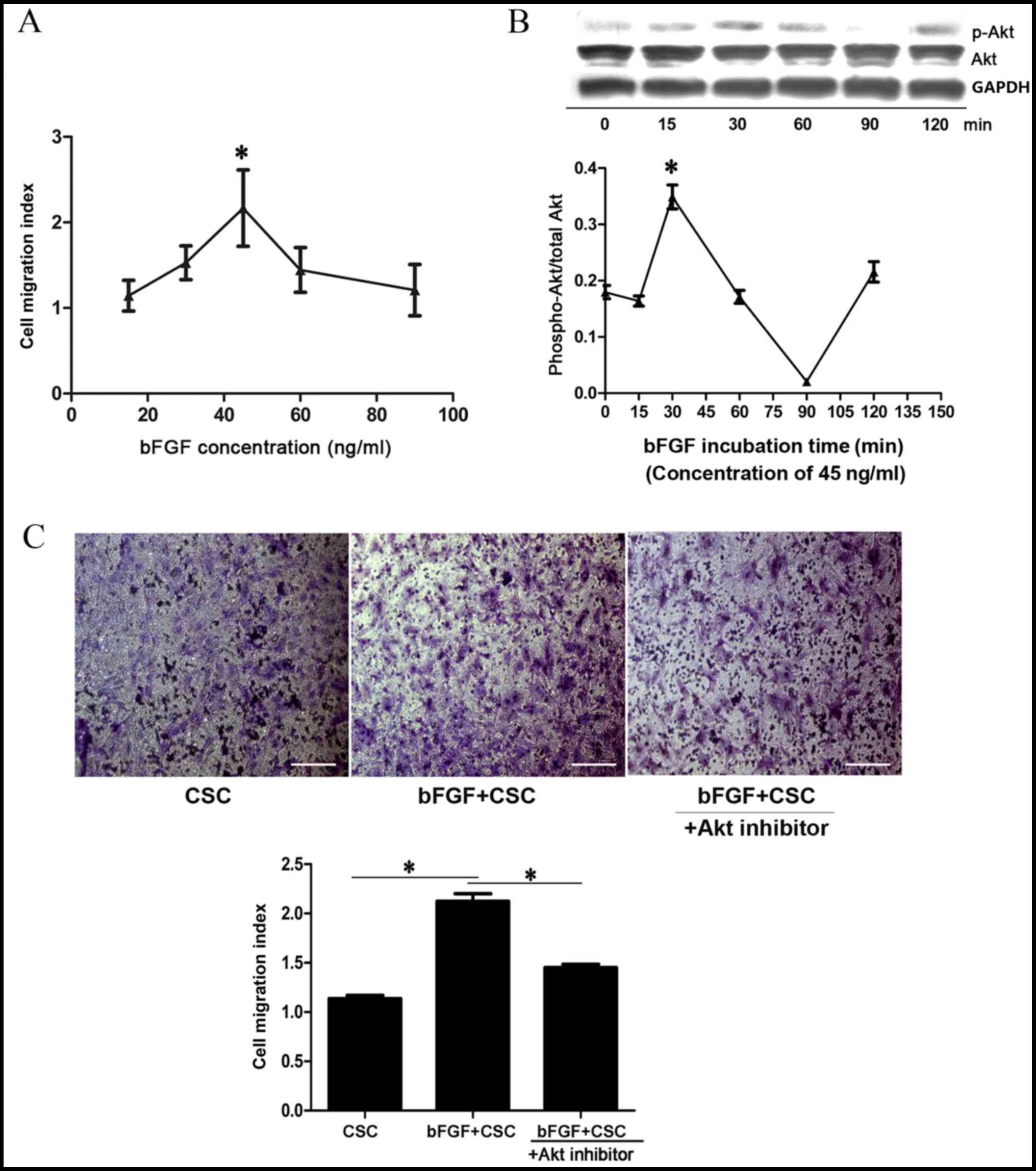
bFGF promotes Sca-1+ CSC
migration in vitro in an Akt-dependent manner
A Transwell assay was used to assess the effects of
bFGF on Sca-1+ CSC migration in vitro. A
concentration gradient of bFGF (15, 30, 45, 60 and 90 ng/ml) was
used. The migration index of Sca-1+ CSCs increased with
bFGF concentration and reached the highest level at 45 ng/ml
(2.16±0.44; P=0.001 vs. 0 ng/ml; Fig.
2A), and decreased to baseline level at 90 ng/ml (1.21±0.30;
Fig. 2A). To determine whether the
PI3K/Akt pathway participates in bFGF-induced Sca-1+ CSC
migration, cell samples treated with 45 ng/ml bFGF were collected
at 0, 15, 30, 60, 90 and 120 min and the level of Akt activation
was analysed by western blotting. The p-Akt/Akt ratio increased
with time and reached the highest level at 30 min, suggesting a
transient activation of p-Akt (0.34±0.02, P=0.001 vs. 0 min;
Fig. 2B). In addition, an Akt
inhibitor, deguelin, was used to determine whether bFGF dependent
Sca-1+ CSC migration could be attenuated. The
Sca-1+ CSC migration index was revealed to be 1.14±0.06
for CSCs, 2.12±0.13 for bFGF-treated CSCs (P=0.001 vs. CSC;
Fig. 2C) and 1.45±0.06 for
bFGF-treated CSCs + deguelin (P=0.004 vs. bFGF-treated CSCs;
Fig. 2C). This suggests that the
PI3K/Akt pathway is involved in CSC migration.
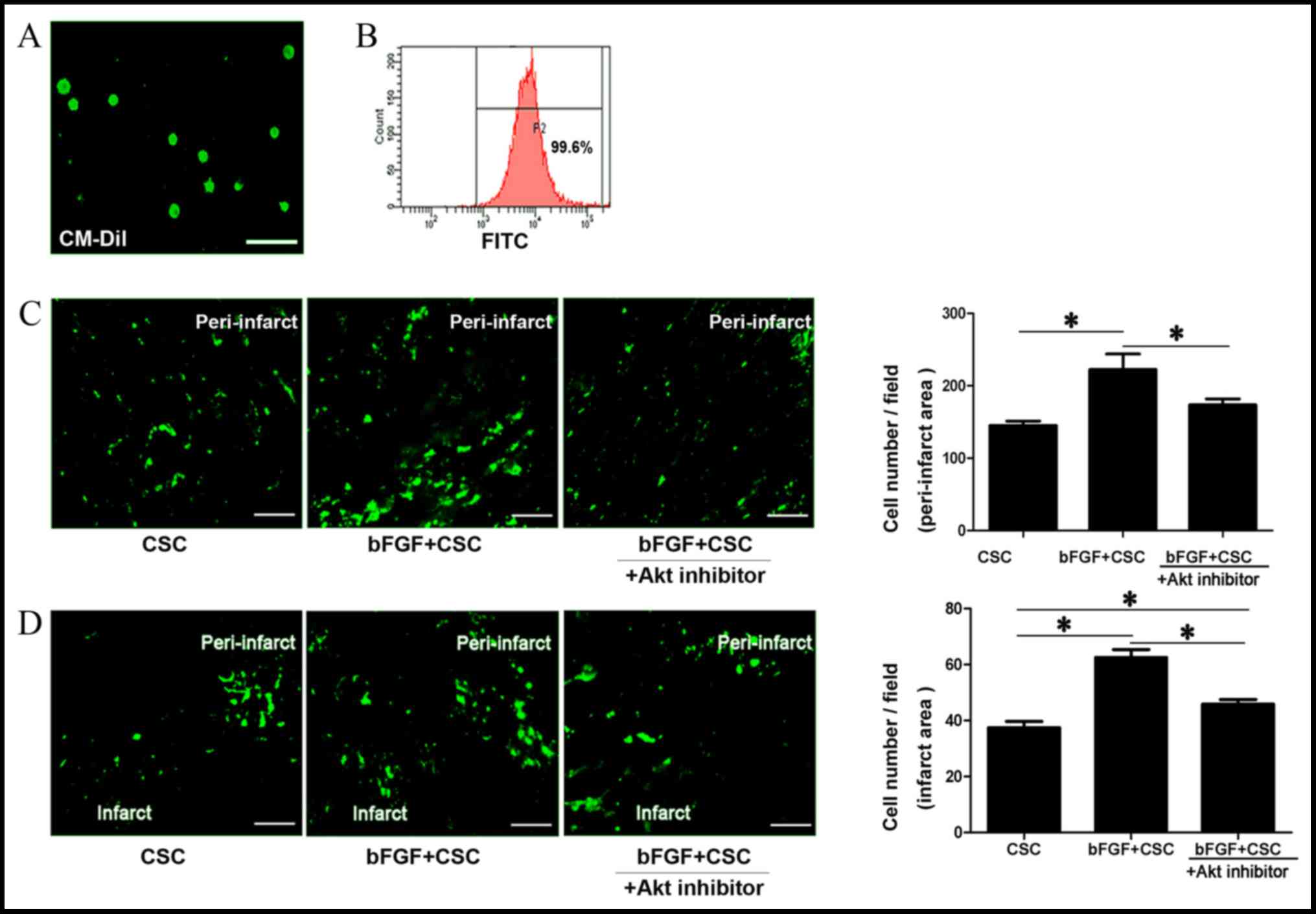
bFGF promoted cell maintenance and
migration in vivo
Sca-1+ CSCs were labeled with CM-Dil cell
tracking dye with a positive labeling rate of 99.6%, confirmed by
flow cytometry and fluorescent microscopy (Fig. 3A and B). One week after cell
transplantation, the viability of transplanted cells in the
peri-infarct area and migrated cells in the infarct area were
assessed via fluorescence microscopy. Sca-1+ CSCs
demonstrated higher cell maintenance rates in the peri-infarct area
when treated with bFGF (control CSCs, 166.4±20.45 cells/field;
bFGF-treated CSCs, 222.4±47.95 cells/field; P=0.001; Fig. 3C) and the Akt inhibitor deguelin
reduced this effect (bFGF-treated CSCs + deguelin, 173.6±18.39
cells/field; P=0.001 vs. bFGF-treated CSC; Fig. 3C). Cell number in the infarct area,
reflecting the migratory ability of the transplanted cells, was
also examined. Sca-1+ CSCs demonstrated improved
migratory ability when treated with bFGF, with increased numbers of
cells migrating into the infarct myocardium (control CSCs,
37.43±5.95 cells/field; bFGF-treated CSCs, 62.58±7.34 cells/field;
P=0.001; Fig. 3D). This effect was
partially blocked by the Akt inhibitor deguelin (bFGF-treated CSC +
deguelin 45.85±4.33 cells/field; P=0.001 vs. control CSCs, P=0.001
vs bFGF-treated CSCs; Fig. 3D).
This suggests that bFGF delivery promotes CSC maintenance in the
peri-infarct area and cell migration to the infarct area, and these
effects were blocked by an Akt inhibitor.
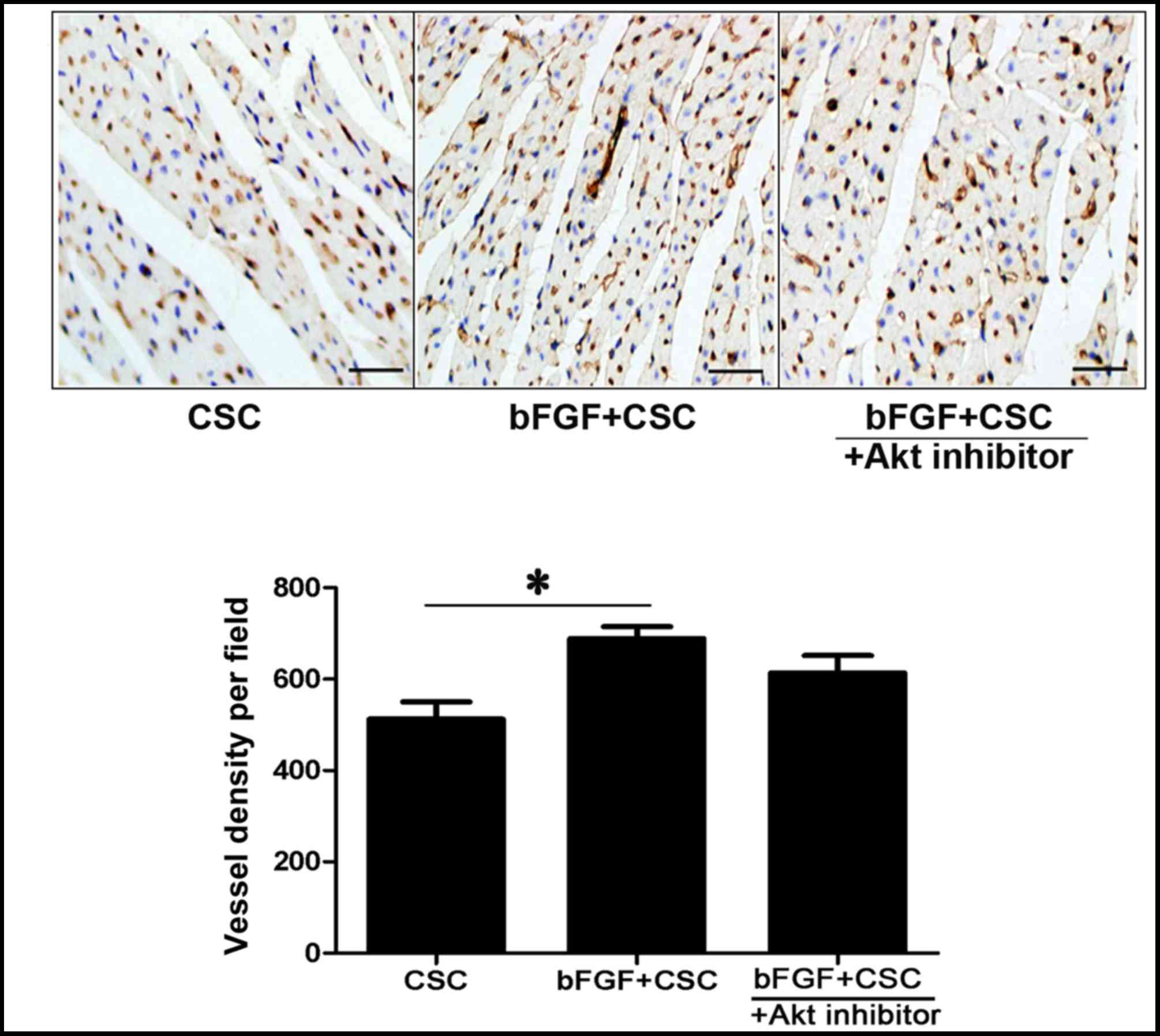
bFGF-treated CSC transplantation
increases capillary density in the infarcted myocardium
vWF immunohistochemistry was used to measure
capillary density in the infarcted myocardium. The mean number of
micro-vessels per field in the peri-infarct myocardium was
increased in the bFGF-treated CSC group (control CSC 512.5±75.76
vessels/field; bFGF-treated CSC 689.25±51.73 vessels/field;
P=0.001; Fig. 4). This effect was
partially attenuated by the Akt inhibitor deguelin (bFGF-treated
CSC + deguelin 613.5±76.44 vessels/field; Fig. 4) but no significant difference was
observed between the bFGF-treated CSC group and the bFGF-treated
CSC + deguelin group.
Upregulation of bFGF, CXCR-4 and SDF-1
is Akt dependent
Western blot analysis was performed to evaluate
protein expression of bFGF, CXCR-4 and SDF-1. An up-regulation of
bFGF (1.72±0.14 fold; P=0.001; Fig.
5A), CXCR-4 (1.94±0.1 fold; P=0.001; Fig. 5B) and SDF-1 (1.51±0.12 fold;
P=0.001; Fig. 5C) was observed in
the bFGF-treated CSC group compared with the control CSC group
following cell transplantation. This upregulation of bFGF and
CXCR-4 was partially attenuated by exogenous Akt inhibitor, with
smaller increases in protein expression of bFGF (1.58±0.07 fold;
P=0.001; Fig. 5A), CXCR-4
(1.79±0.09 fold; P=0.001 vs. CSC and P=0.001 vs. bFGF-treated CSC;
Fig. 5B) in the bFGF-treated CSC +
deguelin group compared with the control CSC group.
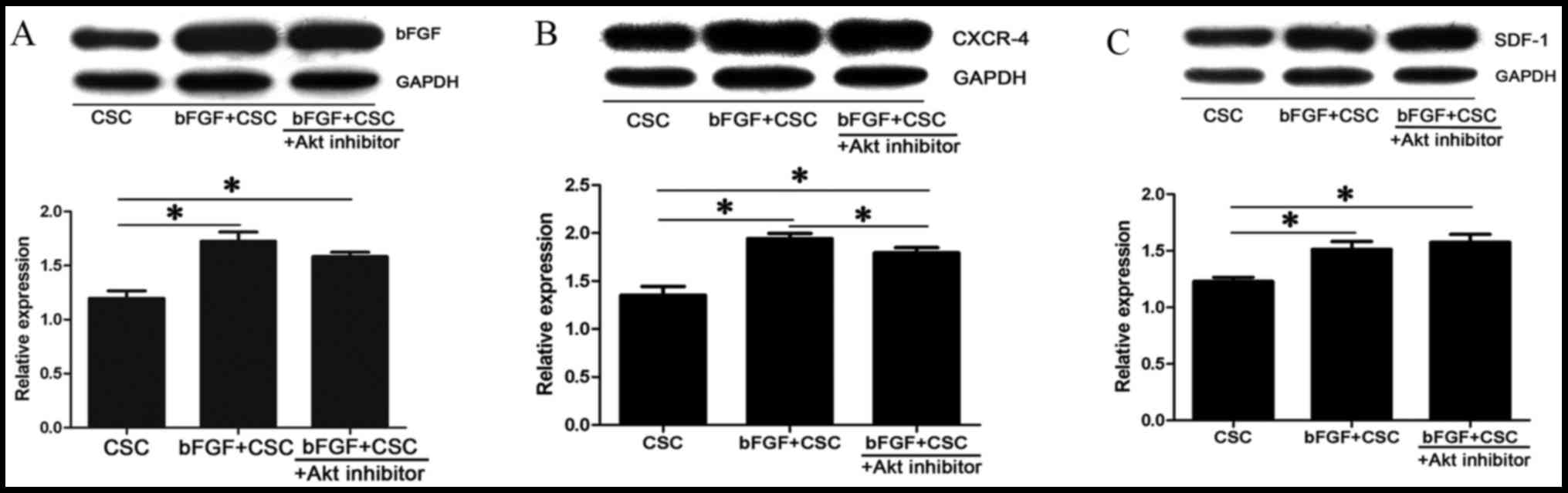 | Figure 5.Activation of CXCR-4 and SDF-1
following bFGF treatment. Protein expression levels, with
quantitation relative to GAPDH, of (A) bFGF, (B) CXCR-4 and (C)
SDF-1 in the peri-infarct myocardium one week following cell
transplantation, measured by western blot analysis. Each assay was
repeated three times. *P<0.05, with comparisons indicated by
lines. CXCR-4, C-X-C chemokine receptor type 4; SDF-1, stromal cell
derived factor 1; bFGF, basic fibroblast growth factor; CSC,
cardiac stem cells; Akt, protein kinase B; GADPH, glyceraldehyde
3-phosphate dehydrogenase. |
Discussion
The present study demonstrated that bFGF treatment
promoted the migratory ability of cultured CSCs. This effect was
observed in vitro and in vivo, with increased CSC
migration into the infarcted myocardium observed in the
bFGF-treated group. It was hypothesized that this effect may have
been mediated through the PI3K/Akt pathway, with specific blocking
of this pathway attenuating the effect of bFGF on CSC
migration.
The present study demonstrated that expression of
bFGF was elevated following myocardial infarction in both the
ischemic and peri-ischemic area. Immunofluorescence staining
revealed increased numbers of Sca-1+ CSCs in the
peri-ischemic area, suggesting links between Sca-1+ CSC
recruitment and bFGF concentration. Ischemia and hypoxia can
stimulate the release of bFGF from the intracellular and
extra-cellular matrices, and it binds to specific fibroblast growth
factor receptors on the cell membrane (17). bFGF is known to have potent
angiogenic abilities, and is able to protect cardiomyocytes from
ischemic/reperfusion injury by promoting capillary formation and
increasing micro-vessel density, which ameliorates the local blood
supply (27,28). In a rabbit myocardial infarction
model, intra-myocardial injection of bFGF promoted vascular
endothelium recovery and the maintenance of cardiac function
(29). The present study revealed
that bFGF-treated CSC transplantation led to improved angiogenesis
and increased vessel density. It was hypothesized that angiogenesis
increased the efficiency of stem cell transplantation by
facilitating cell survival and migration due to an increased supply
of blood and oxygen. This was in accordance with the observation
that there was increased stem cell maintenance in the implanted
area (30).
The migration of stem cells primarily occurs through
chemotaxis. Many signaling pathways, such as PI3K/Akt and the
SDF-1/CXCR-4 axis are involved in the mobilization, migration,
homing, and implantation of stem cells (11–13).
Myocardial infarction is an inflammatory response resulting from
ischemia and hypoxia, and is accompanied with augmented release of
cytokines such as VEGF, bFGF and HIF in the local area. These are
all important chemotaxis factors involved in cell migration
(31–33). Under the control of these
cytokines, SDF-1 recruits bone marrow and peripheral stem cells
into the ischemic and peri-ischemic area, where the concentration
is greatest (34). The present
study observed the up-regulation of SDF-1 and CXCR-4 in the
bFGF-treated group, coinciding with the improved migratory ability
in this group. When the PI3K/Akt pathway was blocked by an
inhibitor, the migration of CSCs was inhibited and the expression
of SDF-1 and CXCR-4 also decreased. This suggests that the PI3K/Akt
pathway may partially modulate the SDF-1/CXCR-4 axis in the process
of cell migration.
To conclude, the present study demonstrated that
bFGF promoted the migration of CSCs, both in vitro and in
vivo. This effect was partially mediated through the PI3K/Akt
pathway. Transplantation of bFGF-treated CSCs in a murine
myocardial infarction model increased CSC maintenance and
migration, as well as local angiogenesis. This pro-migratory effect
of bFGF demonstrates that its importance to myocardial regeneration
extends beyond its traditional angiogeneic ability. This
understanding, if incorporated into CSC therapy, may improve its
efficiency when treating myocardial infarct. The limitation of this
study was that the CSCs were transplanted through myocardium
injection. A more convenient method could have been intravenous
injection but the best method for cell transplantation requires
further study.
Acknowledgements
The present study was supported by The Youth Science
and Technology of Suzhou Science and Education Project (grant no.
KJXW2013004), The Youth Science Foundation of Jiangsu Province,
China (grant no. BK20140296) and The Science Foundation for Youth
Teacher of Soochow University (grant no. SDY2013A29).
References
|
1
|
Anderson KM: Discharge clinical
characteristics and 60-day readmission in patients hospitalized
with heart failure. J Cardiovasc Nurs. 29:232–241. 2014.PubMed/NCBI
|
|
2
|
Noseda M, Abreu-Paiva M and Schneider MD:
The Quest for the adult cardiac stem cell. Circ J. 79:1422–1430.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mosna F, Annunziato F, Pizzolo G and
Krampera M: Cell therapy for cardiac regeneration after myocardial
infarct: Which cell is the best? Cardiovasc Hematol Agents Med
Chem. 8:227–243. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nadal-Ginard B, Ellison GM and Torella D:
The cardiac stem cell compartment is indispensable for myocardial
cell homeostasis, repair and regeneration in the adult. Stem Cell
Res. 13:615–630. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Barile L, Messina E, Giacomello A and
Marbán E: Endogenous cardiac stem cells. Prog Cardiovasc Dis.
50:31–48. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bergmann O, Bhardwaj RD, Bernard S, Zdunek
S, Barnabé-Heider F, Walsh S, Zupicich J, Alkass K, Buchholz BA,
Druid H, et al: Evidence for cardiomyocyte renewal in humans.
Science. 324:98–102. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bearzi C, Rota M, Hosoda T, Tillmanns J,
Nascimbene A, De Angelis A, Yasuzawa-Amano S, Trofimova I, Siggins
RW, Lecapitaine N, et al: Human cardiac stem cells. Proc Natl Acad
Sci USA. 104:pp. 14068–14073. 2007; View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Forte G, Minieri M, Cossa P, Antenucci D,
Sala M, Gnocchi V, Fiaccavento R, Carotenuto F, De Vito P, Baldini
PM, et al: Hepatocyte growth factor effects on mesenchymal stem
cells: Proliferation, migration, and differentiation. Stem Cells.
24:23–33. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tang J, Wang J, Kong X, Yang J, Guo L,
Zheng F, Zhang L, Huang Y and Wan Y: Vascular endothelial growth
factor promotes cardiac stem cell migration via the PI3K/Akt
pathway. Exp Cell Res. 315:3521–3531. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tang JM, Wang JN, Zhang L, Zheng F, Yang
JY, Kong X, Guo LY, Chen L, Huang YZ, Wan Y and Chen SY: VEGF/SDF-1
promotes cardiac stem cell mobilization and myocardial repair in
the infarcted heart. Cardiovasc Res. 91:402–411. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Unzek S, Zhang M, Mal N, Mills WR, Laurita
KR and Penn MS: SDF-1 recruits cardiac stem cell-like cells that
depolarize in vivo. Cell Transplant. 16:879–886. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Madonna R, Rokosh G, De Caterina R and
Bolli R: Hepatocyte growth factor/Met gene transfer in cardiac stem
cells-potential for cardiac repair. Basic Res Cardiol. 105:443–452.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Urbanek K, Rota M, Cascapera S, Bearzi C,
Nascimbene A, De Angelis A, Hosoda T, Chimenti S, Baker M, Limana
F, et al: Cardiac stem cells possess growth factor-receptor systems
that after activation regenerate the infarcted myocardium,
improving ventricular function and long-term survival. Circ Res.
97:663–673. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Takehara N, Tsutsumi Y, Tateishi K, Ogata
T, Tanaka H, Ueyama T, Takahashi T, Takamatsu T, Fukushima M,
Komeda M, et al: Controlled delivery of basic fibroblast growth
factor promotes human cardiosphere-derived cell engraftment to
enhance cardiac repair for chronic myocardial infarction. J Am Coll
Cardiol. 52:1858–1865. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Virag JA, Rolle ML, Reece J, Hardouin S,
Feigl EO and Murry CE: Fibroblast growth factor-2 regulates
myocardial infarct repair: Effects on cell proliferation, scar
contraction, and ventricular function. Am J Pathol. 171:1431–1440.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tabata Y and Ikada Y: Vascularization
effect of basic fibroblast growth factor released from gelatin
hydrogels with different biodegradabilities. Biomaterials.
20:2169–2175. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Boodhwani M, Voisine P, Ruel M, Sodha NR,
Feng J, Xu SH, Bianchi C and Sellke FW: Comparison of vascular
endothelial growth factor and fibroblast growth factor-2 in a swine
model of endothelial dysfunction. Eur J Cardiothorac Surg.
33:645–650. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cuevas P, Barrios V, Giménez-Gallego G,
Martinez-Coso V, Cuevas B, Benavides J, Garcia-Segovia J and
Asin-Cardiel E: Serum levels of basic fibroblast growth factor in
acute myocardial infarction. Eur J Med Res. 2:282–284.
1997.PubMed/NCBI
|
|
19
|
Fujita M, Ikemoto M, Kishishita M, Otani
H, Nohara R, Tanaka T, Tamaki S, Yamazato A and Sasayama S:
Elevated basic fibroblast growth factor in pericardial fluid of
patients with unstable angina. Circulation. 94:610–613. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Anversa P, Rota M, Urbanek K, Hosoda T,
Sonnenblick EH, Leri A, Kajstura J and Bolli R: Myocardial aging-a
stem cell problem. Basic Res Cardiol. 100:482–493. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Linke A, Müller P, Nurzynska D, Casarsa C,
Torella D, Nascimbene A, Castaldo C, Cascapera S, Böhm M, Quaini F,
et al: Stem cells in the dog heart are self-renewing, clonogenic,
and multipotent and regenerate infarcted myocardium, improving
cardiac function. Proc Natl Acad Sci USA. 102:pp. 8966–8971. 2005;
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Urbanek K, Torella D, Sheikh F, De Angelis
A, Nurzynska D, Silvestri F, Beltrami CA, Bussani R, Beltrami AP,
Quaini F, et al: Myocardial regeneration by activation of
multipotent cardiac stem cells in ischemic heart failure. Proc Natl
Acad Sci USA. 102:pp. 8692–8697. 2005; View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Smith RR, Barile L, Messina E and Marbán
E: Stem cells in the heart: What's the buzz all about?––Part 1:
Preclinical considerations. Heart Rhythm. 5:749–757. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang X, Hu Q, Nakamura Y, Lee J, Zhang G,
From AH and Zhang J: The role of the sca-1+/CD31- cardiac
progenitor cell population in postinfarction left ventricular
remodeling. Stem Cells. 24:1779–1788. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Müller-Ehmsen J, Krausgrill B, Burst V,
Schenk K, Neisen UC, Fries JW, Fleischmann BK, Hescheler J and
Schwinger RHG: Effective engraftment but poor mid-term persistence
of mononuclear and mesenchymal bone marrow cells in acute and
chronic rat myocardial infarction. J Mol Cell Cardiol. 41:876–884.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang M, Methot D, Poppa V, Fujio Y, Walsh
K and Murry CE: Cardiomyocyte grafting for cardiac repair: Graft
cell death and anti-death strategies. J Mol Cell Cardiol.
33:907–921. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Iwakura A, Fujita M, Kataoka K, Tambara K,
Sakakibara Y, Komeda M and Tabata Y: Intramyocardial sustained
delivery of basic fibroblast growth factor improves angiogenesis
and ventricular function in a rat infarct model. Heart Vessels.
18:93–99. 2000. View Article : Google Scholar
|
|
28
|
Kawasuji M, Nagamine H, Ikeda M,
Sakakibara N, Takemura H, Fujii S and Watanabe Y: Therapeutic
angiogenesis with intramyocardial administration of basic
fibroblast growth factor. Ann Thorac Surg. 69:1155–1161. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bougioukas I, Didilis V, Ypsilantis P,
Giatromanolaki A, Sivridis E, Lialiaris T, Mikroulis D, Simopoulos
C and Bougioukas G: Intramyocardial injection of low-dose basic
fibroblast growth factor or vascular endothelial growth factor
induces angiogenesis in the infarcted rabbit myocardium. Cardiovasc
Pathol. 16:63–68. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fujio Y, Nguyen T, Wencker D, Kitsis RN
and Walsh K: Akt promotes survival of cardiomyocytes in vitro and
protects against ischemia-reperfusion injury in mouse heart.
Circulation. 101:660–667. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nervi B, Link DC and DiPersio JF:
Cytokines and hematopoietic stem cell mobilization. J Cell Biochem.
99:690–705. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Asahara T, Takahashi T, Masuda H, Kalka C,
Chen D, Iwaguro H, Inai Y, Silver M and Isner JM: VEGF contributes
to postnatal neovascularization by mobilizing bone marrow-derived
endothelial progenitor cells. EMBO J. 18:3964–3972. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ceradini DJ, Kulkarni AR, Callaghan MJ,
Tepper OM, Bastidas N, Kleinman ME, Capla JM, Galiano RD, Levine JP
and Gurtner GC: Progenitor cell trafficking is regulated by hypoxic
gradients through HIF-1 induction of SDF-1. Nat Med. 10:858–864.
2004. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kucia M, Jankowski K, Reca R, Wysoczynski
M, Bandura L, Allendorf DJ, Zhang J, Ratajczak J and Ratajczak MZ:
CXCR4-SDF-1 signalling, locomotion, chemotaxis and adhesion. J Mol
Histol. 35:233–245. 2004. View Article : Google Scholar : PubMed/NCBI
|