Introduction
Osteoarthritis (OA) is a chronic joint disease,
characterized by degeneration of articular cartilage and secondary
bone hyperplasia, ultimately resulting in stiffness, swelling,
pain, and functional incapacitation of joints (1). As one of the most common types of
arthritis, OA usually occurs in weight-bearing joints of the lower
extremities, such as the hip joints and knee joints (2). As recorded, the risk of disability
caused by OA is approximately 40% in males and 47% in females,
which would be higher in obese people (3). With the rapid aging of population,
more and more middle-aged and elderly people are bothered by OA,
seriously affecting the quality of life of patients (4,5).
Despite the etiology of OA is very complicated and has not been
clearly clarified, a variety of factors can lead to the development
of OA, like joint trauma, abnormal mechanical burden, poor
nutrition and genetic predisposition (6,7). At
present, most of the medicine used to treat OA can temporarily
relieve the pain of joints, but cannot improve the destruction of
inflammatory joints (8).
Therefore, it is important to find the molecular targets for the
development and progression of OA, and to create new therapeutic
strategies for improving the treatment and prognosis of OA
patients.
Receptor-interacting protein kinase 4 (RIPK4) is a
family member of the receptor-interacting kinase proteins, which
have served as crucial sensors of intracellular and extracellular
stresses (9). Located on
chromosome 21q22.3 and consisted of 8 exons and 7 introns, RIPK4
gene can encode 82 kd proteins (10). RIPK4, as the important apoptosis
modulator, contains an N-terminal RIP-like kinase domain and a
C-terminal region, which is characterized by the presence of 11
ankyrin repeats (11,12). To our knowledge, RIPK4 is widely
expressed in embryonic and mature tissues, such as myocardium,
liver, skeletal muscle and lung (13), as well as involved in a variety of
signaling pathways to play a vital part in regulating cells
apoptosis, proliferation, differentiation and inflammatory response
(14). So far, mounting evidence
have shown that RIPK4 is up-regulated in various tumor tissues,
which can promote the occurrence and progression of many malignant
tumors, such as ovarian cancer (15) and cervical cancer (16). Besides, RIPK4 is also required for
the formation of keratinocytes, the differentiation of epithelial
cells, the skin inflammation, the healing of wound surface, as well
as the tumor differentiation (11,17,18).
Although the occurrence of OA is closely related to chondrocyte
apoptosis as suggested by the previous study (19), there is no article reported whether
RIPK4 plays a regulatory role in the chondrocytes of OA until
now.
Therefore, this study is to investigate the
expression of RIPK4 and its effect on the apoptosis and
proliferation of chondrocytes during the OA, whereby providing a
reliable experimental basis for the clinical use of gene therapy to
treat OA.
Materials and methods
Ethics statement
This study was in accordance with the Helsinki
Declaration (20), all patients
were informed the purpose of the study and signed an informed
consent before participation. The experiments in the study were
approved by the Ethics Committee of Jingzhou Central Hospital
(Jingzhou, China).
Study subjects
From December 2015 to December 2016, we collected 28
specimens of degenerated cartilage tissues from 10 male and 18
female patients (mean age: 56.8±8.7 years) who underwent
arthroscopic debridement or total knee arthroplasty due to knee OA.
Patients were diagnosed according to the OA diagnostic criteria
issued by the American College Rheumatology (ACR) in 1995 (21). At the same time, 20 specimens of
normal joint tissues were obtained as healthy controls from 8 males
and 12 females (mean age: 57.4±10.1 years) who received amputation
due to severe trauma to the lower extremities. There was no
significant difference in the sex ratio and age between the
patients with OA and the normal controls (all P>0.05).
Chondrocyte isolation and culture
Articular cartilage was cut into pieces under
sterile condition before adding 0.2% type-II collagenase (Sigma,
USA) of 10 times volume for 3~4 h of stirring and digestion at
37°C. Next, 5,200 mesh sieve was used to filter the cell
suspension, which was centrifuged for 10 min at the rate of 1,200 ×
g to remove the supernatant. Then, cells were washed for 2 times
with 10% serum culture medium, and the cells obtained were cultured
in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal
bovine serum and placed in 37°C, 5% CO2 incubator with
saturated humidity.
Cell transfection and grouping
Cells under good state were spread on 6-well plate.
When the cell density reaches 80%, transfection experiment was
conducted with transfection mixture prepared according to the
instructions on Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) kit. The medium solution
was discarded, prepared transfection mixture was added into the
plate, and cells were cultured at 37°C. Cells were classified into
six groups: i) Normal group (normal articular chondrocytes); ii) OA
group (OA chondrocytes); iii) NC group (OA chondrocytes transfected
with negative control); iv) si-RIPK4 group (OA chondrocytes
transfected with si-RIPK4); v) Wnt3a group [OA chondrocytes treated
with a Wnt signaling agonist Wnt-3a (100 ng/ml)]; and vi)
si-RIPK4+Wnt3a group (OA chondrocytes treated with si-RIPK4 and the
Wnt signaling agonist Wnt-3a). The siRNA target sequence used to
effectively inhibit the expression of RIPK4 and negative control
(NC) sequence were purchased from Shanghai Genechem Co., Ltd.
(Shanghai, China).
Quantitative PCR detects the mRNA
expression of RIPK4
The total RNA extracted from joint tissues or cells
by Trizol reagent (Invitrogen; Thermo Fisher Scientific, Inc.) was
determined for RNA concentration and purity. Sample RNA was
reversed to cDNA in accordance with instructions on the reverse
transcription kit (cat. no. DRR047S; Takara Biotechnology Co.,
Ltd., Dalian, China) and the total system volume was 10 µl. Then 65
µl of DEPC-treated water was added into the cDNA for dilution and
fully mixing. qPCR reaction system contains 5 µl SsoFast EvaGreen
Supermix (cat. no. 1708882; Bio-Rad Laboratories, Inc., Hercules,
CA, USA), 0.5 µl forward primer (10 µM), 0.5 µl reverse primer (10
µM), and 4 µl cDNA. PCR amplification conditions included 40 cycles
of pre-degeneration at 95°C for 5 min, degeneration at 94°C for 60
sec, annealing at 55°C for 60 sec, and extending at 72°C for 120
sec. Primers were synthesized by Beijing Genomics Institute (BGI,
Beijing, China). The primers of RIPK4 were
5′-GGATGCCCACTACCACGTCA-3′ and 5′-TGCCAAACAGGCCATCCA-3′. GAPDH was
the internal reference gene and its primers were
5′-CTACCCACGGCAAGTTCAAT-3′ and 5′-GGATGCAGGGATGATGTTCT-3′. Each
gene of every sample would have three repeats. The reliability of
PCR results was evaluated by melting curve. Cq value (inflection
point of amplification curve), ∆Cq (=Cqtarget
gene-Cqinternal reference gene), ∆∆Cq
(=∆Cqcase group-∆Cqcontrol group), and
2−∆∆Cq was used to calculate the relative expression of
target genes (22). The experiment
was repeated for three times.
The protein expression detected by
western blotting
The total protein extracted from joint tissues or
cells was detected for the protein concentration by BCA kit
(Beyotime Biotechnology Co., Ltd., Shanghai, China). Then, 5X SDS
loading buffer was added for 5 min of denaturation at 95°C. Next,
SDS-PAGE electrophoresis was conducted before transferring membrane
and adding 5% skim milk for closure overnight at 4°C. Then, after
membrane washing with Tris-Buffered Saline Tween-20 (TBST), the
following primary antibodies (all in 1:1,000 dilution) were added
respectively, including RIPK4 (ab84365; Abcam, Cambridge, UK),
GSK-3β (ab68476; Abcam), β-catenin (ab6302; Abcam), Wnt3a (ab28472;
Abcam), and β-actin (ab8226; Abcam). After incubation overnight at
4°C, membranes were washed with TBST and secondary antibody
horseradish peroxidase (HRP) was added to incubate for 1 h at 37°C.
At last, the membrane was washed with TBST and horseradish
peroxidase ECL was added for development, scanning and recording.
Image J software (NIH Image; National Institutes of Health,
Bethesda, MD, USA) was used to analyze the gray value of target
bands. The experiment was repeated three times independently.
Cell proliferation detected by MTT
assay
The chondrocytes being transfected were diluted to a
certain concentration and spread on 96-well plates with
5×103 cells in each well. The plates were placed in
37°C, CO2 incubator for 24, 48, 72, and 96 h,
respectively. Next, 20 µl MTT reagent (Promega Corporation,
Madison, WI, USA) was added into each well for another 1–4 h of
incubation. Then, the absorbance value was determined by the
multifunctional microplate under the wavelength of 490 nm.
Cell apoptosis rate detected by flow
cytometry
Cells transfected for 24 h were digested by trypsin,
centrifuged, and collected. The cells obtained were washed with
cold PBS and made into 1×106 cells/ml single cell
suspension with 500 µl binding buffer solution (calcium containing
PBS). At room temperature, 100 ul of cell suspension was added into
the tube, followed by 5 µl Annexin V-FITC (BD Biosciences, Franklin
Lakes, NJ, USA) and 5 µl propidium iodide (PI) (BD Biosciences).
After 30 min of incubation at 4°C, 400 µl binding buffer was added
immediately before detection by the flow cytometry (BD
Biosciences), 104 cells each time. The data was analyzed
using Cell Quest (BD Biosciences) to calculate the rate of cell
apoptosis.
Statistical analysis
The data were analyzed with the statistical software
SPSS 21.0 (SPSS, Inc, Chicago, IL, USA). Measurement data were
presented by mean ± standard deviation. Difference between two
groups of measurement data that obey normal distribution was
analyzed by Student's t-test, while comparison among multiple
groups was conducted by one-way ANOVA. P<0.05 was considered to
indicate a statistically significant difference.
Results
Expression of RIPK4 in cartilage
tissues and chondrocytes
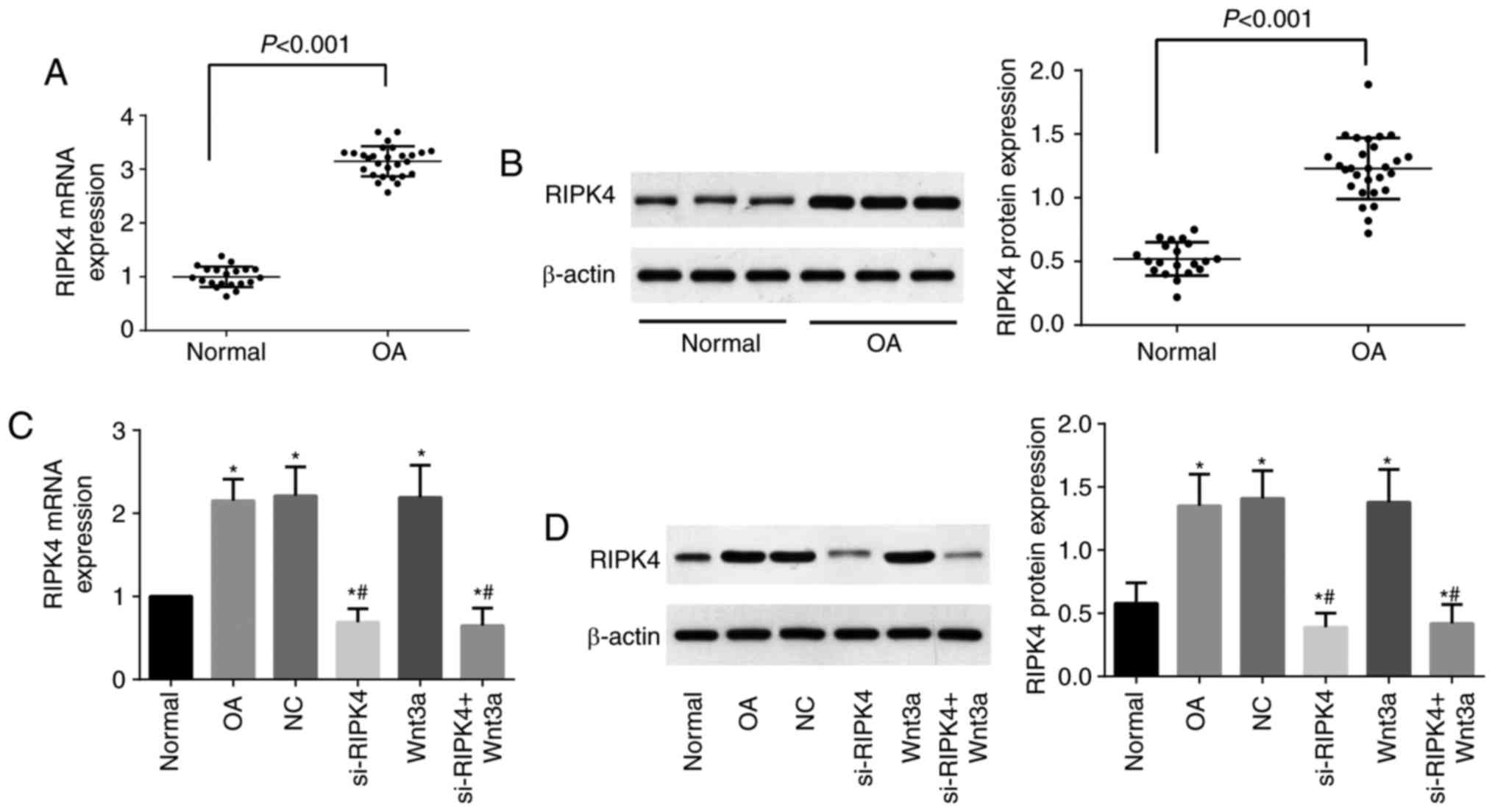
As shown in Fig. 1,
the mRNA and protein expression of RIPK4 in OA cartilage tissues
was significantly upregulated as compared with normal cartilage
tissues (P<0.05). In addition, we also examined the expression
of RIPK4 in chondrocytes after transfection, and by comparison with
the Normal group, the expression of RIPK4 was elevated dramatically
in the OA group, NC group, and Wnt3a group (all P<0.05), but
apparently reduced in the si-RIPK4 group and si-RIPK4+Wnt3a group
(P<0.05). Moreover, RIPK4 expression in the si-RIPK4 and
si-RIPK4+Wnt3a groups was remarkably lower than that in the OA
group and the NC group (all P<0.05), although there was no
significant alterations in Wnt3a group (P>0.05). Chondrocytes
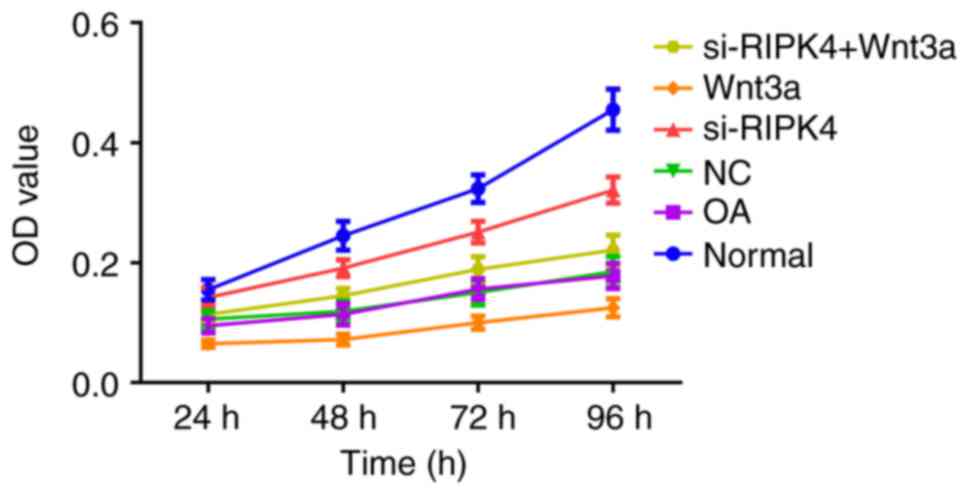
proliferation in each transfected group.
The proliferation of chondrocytes in each
transfected group was detected by MTT. Compared with the Normal
group, the proliferation ability of chondrocytes in the OA group
and NC group was obviously reduced (all P<0.05), but the OA
group and the NC group did not exhibit any significant difference
in cell proliferation (P>0.05), which suggested a reduced
proliferative activity of OA chondrocytes. Besides, the
proliferation ability of chondrocytes was appreciably enhanced in
si-RIPK4 group but obviously decreased in Wnt3a group when compared
with the OA group and NC group (P<0.05). Moreover, the
chondrocyte proliferation ability in the si-RIPK4+Wnt3a group was
lower than in the si-RIPK4 group (P<0.05), indicating that the
inhibition of RIPK4 expression could promote the growth of
chondrocytes, and Wnt3a could reverse the effect of si-RIPK4 on
chondrocyte proliferation (Fig.
2).
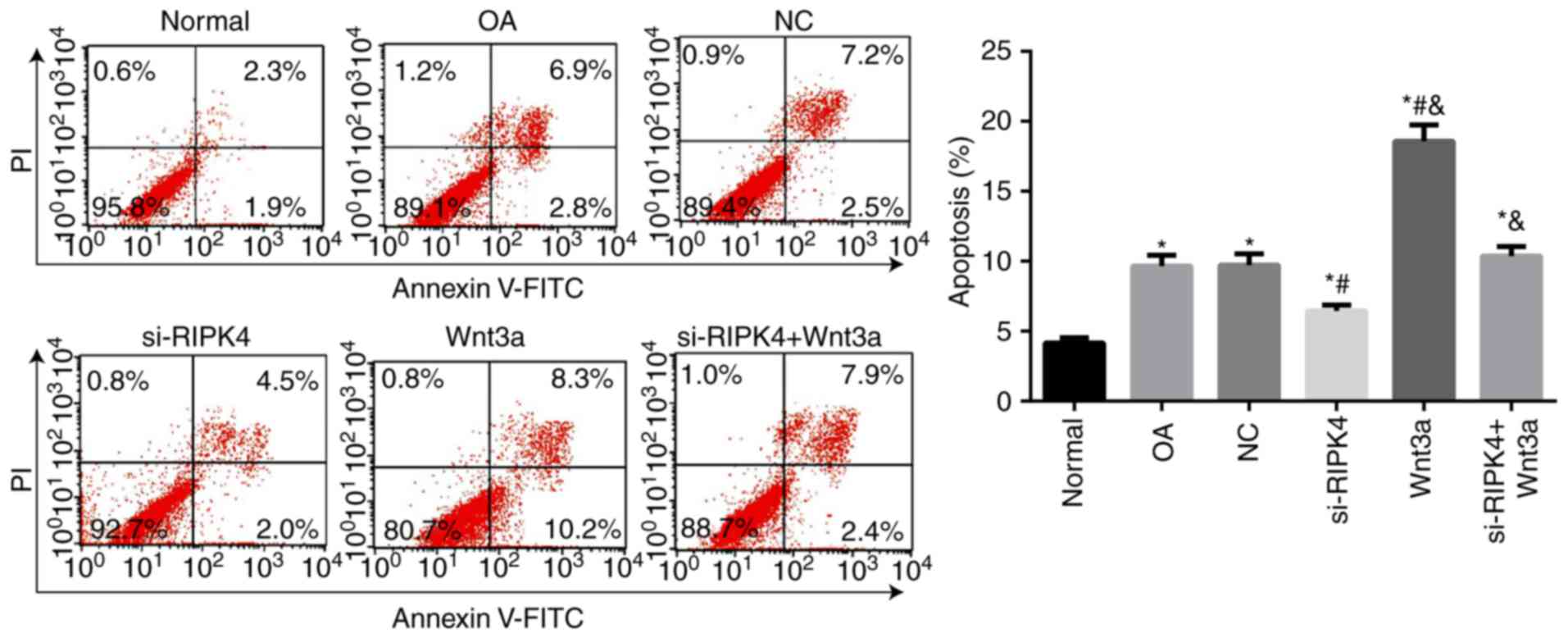
Chondrocytes apoptosis in each
transfected group
The apoptosis of chondrocytes in each transfected
group was detected by flow cytometry (Fig. 3). When compared with the Normal
group, the OA group and NC group increased significantly in the
rate of chondrocytes apoptosis (all P<0.05), while there was no
significant difference in the rate of apoptosis between the OA
group and NC group (P>0.05), which indicated an elevation of the
apoptosis of OA chondrocytes. Meanwhile, the lower cell apoptosis
in si-RIPK4 group but higher cell apoptosis in Wnt3a group were
observed as compared to OA group and NC group (P<0.05).
Furthermore, the apoptosis rate of si-RIPK4+Wnt3a was significantly
elevated when compared with the si-RIPK4 group, suggesting that
silencing RIPK4 can effectively delay the apoptosis of
chondrocytes, and Wnt3a could reverse the effect of si-RIPK4 on
chondrocyte apoptosis.
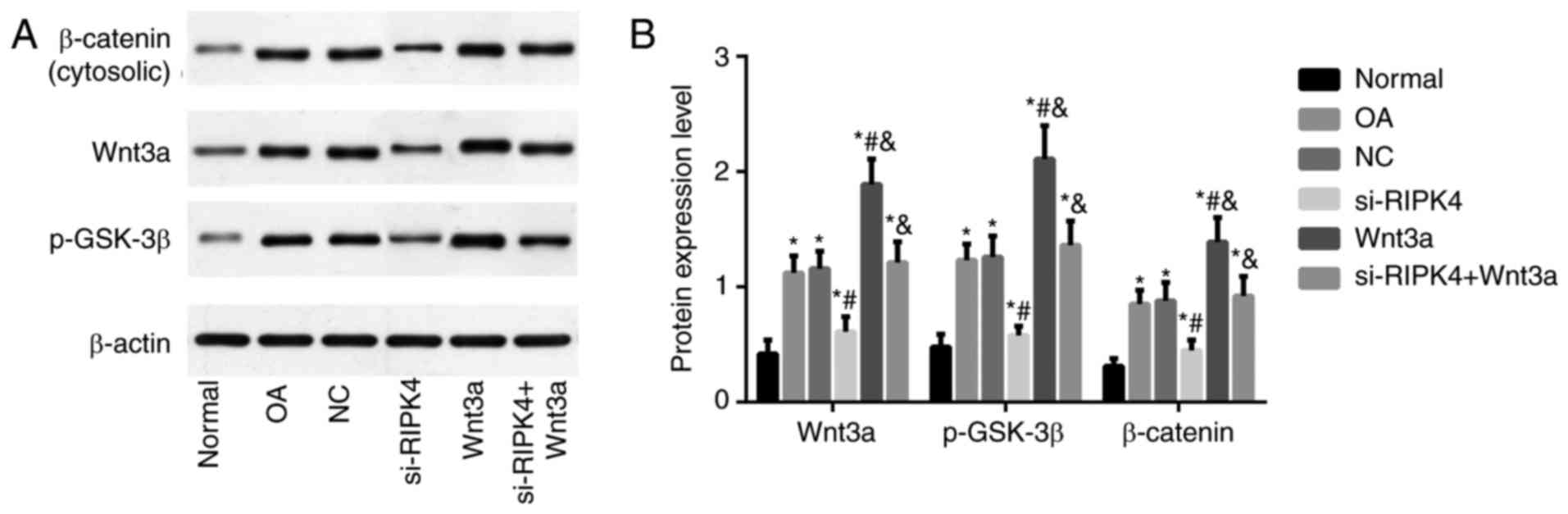
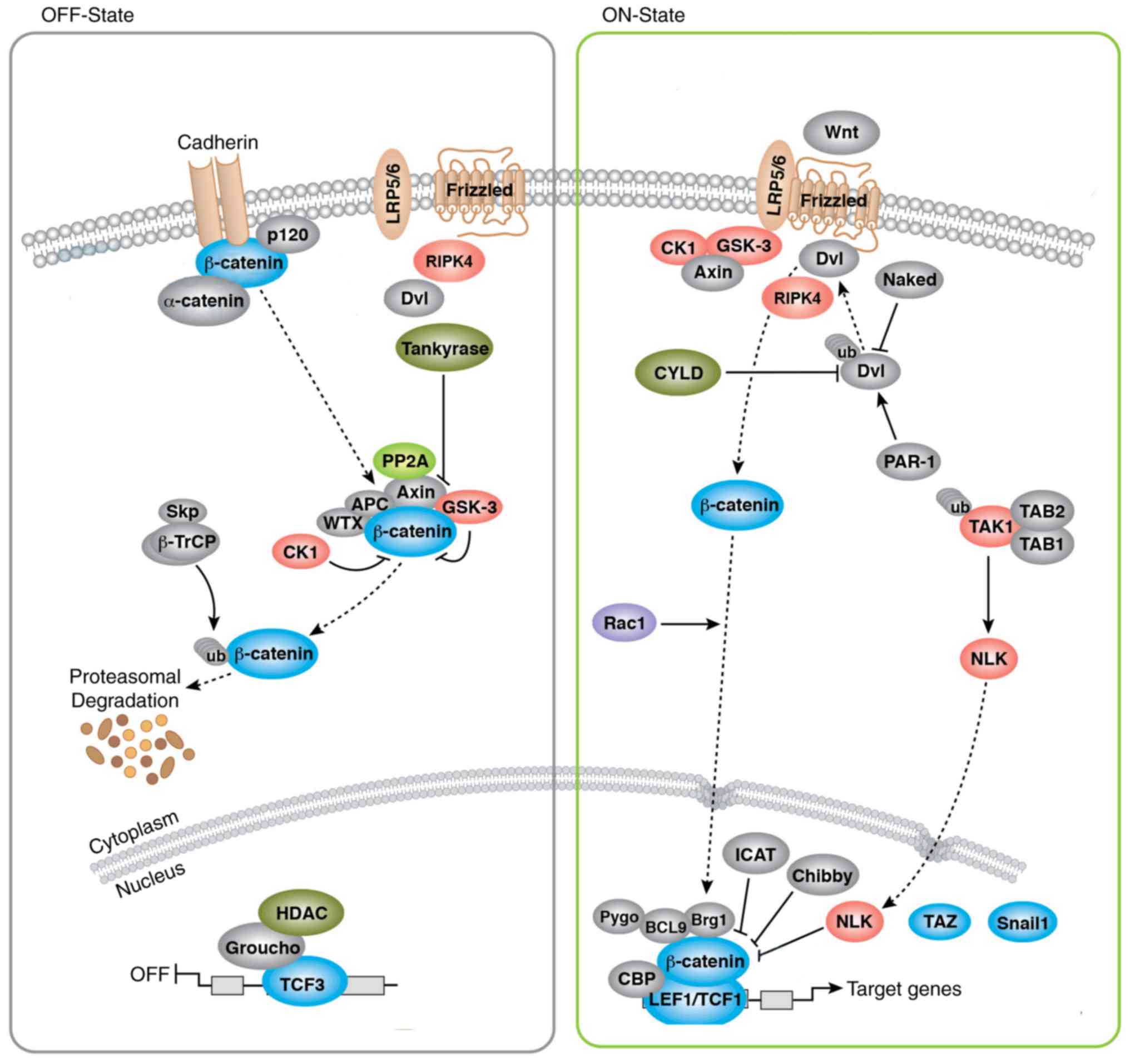
Expression of Wnt/β-catenin pathway
related-proteins in each transfected group
Western blot was used to detect the expressions of
Wnt/β-catenin pathway related-proteins. As shown in Fig. 4, the OA group and NC group showed a
remarkable increase in Wnt3a, β-catenin, and p-GSK-3β proteins, as
compared with the Normal group (all P<0.05), but no significant
difference was found in the expression of Wnt/β-catenin pathway
related-proteins between OA group and NC group (both P>0.05). By
comparison with OA group, the expressions of Wnt3a, β-catenin and
p-GSK-3β were statistically lowered in si-RIPK4 group, but were
highly upregulated in Wnt3a group. Besides, the elevated
expressions of Wnt/β-catenin pathway related-proteins were
displayed in si-RIPK4+Wnt3a, as compared to si-RIPK4 group (all
P<0.05).
Discussion
OA is generally accepted as the most prevalent
articular pathology, as well as the most frequent cause of
disability, becoming a major public health concern (23). Thus, we conducted this study to
better understand the molecular mechanisms implicated in the
apoptosis and proliferation of chondrocytes during OA
pathogenesis.
RIPK4, a key enzyme in the body, plays an important
role in keratinocyte differentiation and wound healing to be
involved in the process of proliferation, differentiation, and
repair of the epithelium (17,24).
A previous study found that RIPK4 was over-expressed in oral
keratinocytes, which could improve the levels of pro-inflammatory
cytokines by inducing the upregulation of CCL5 and CXCL11 (25). Besides, RIPK4 was also upregulated
in cervical cancer tissues and its high expression was closely
related to FIGO stage and lymph node metastasis of cervical cancer
(26). In addition, RIPK4 mRNA and
β-catenin mRNA was reported to be remarkably higher in ovarian
adenocarcinoma cells than in adjacent tissues (15). Consistent with the results of
previous studies, our study also leads to the following findings
that both RIPK4 mRNA and protein expressions were significantly
upregulated in the articular cartilage tissues of OA patients, as
well as chondrocytes, the only cellular component in cartilage,
showing RIPK4 participated in the development and progression of
OA. In the meantime, by isolating chondrocytes from OA and normal
cartilage tissues, we found that the proliferation of OA
chondrocytes was decreased while the apoptotic rate was increased
when compared with normal chondrocytes. During the OA development,
enhanced apoptosis of chondrocytes was credited as a sign of
progressive cartilage joint degeneration as indicated by Huang
et al (27), and there was
an explanation that OA chondrocytes still had some metabolic
function although the proliferative activity of OA chondrocytes was
relatively lower than normal ones, and, thus, a large number of
intracellular proteins and RNA exhibit to be accumulated, which
together with no cell division, can lead to cell necrosis or
apoptosis, eventually enhancing apoptosis rate (28,29).
More importantly, we performed cell transfection with specific
si-RNA of RIPK4 in vitro and we noticed that downregulated
RIPK4 promoted the proliferation and inhibited the apoptosis of OA
chondrocytes, suggesting that RIPK4 might be a potential target in
OA.
Furthermore, aberrant expression of RIPK4 has been
demonstrated to have impacts on several diseases, which is a key
regulatory protein of Wnt/β-catenin signaling (16). As a classical pathway,
Wnt/β-catenin signaling is very essential for the growth,
development, and death of cells (30,31).
It was worthy to point out that Wnt/β-catenin signaling had a deep
relationship with cartilage function, including the development and
differentiation of cartilage (32,33).
In general, the key molecules in the Wnt pathway including
β-catenin, GSK-3β and Wnt3a, in which β-catenin is the core
component, essential for the upstream molecules to exert functions
(34–36). There was a previous study stating
that the expression of β-catenin was increased in degenerated
articular chondrocytes and the loss of cartilage was linked with
the activation of Wnt signaling pathways (37). As for GSK-3β, a crucial part of the
degradation complex formed by β-catenin and Axin-APC-GSK-3β in
cytoplasm, and the inhibition of the activity of GSK-3β can promote
the accumulation of β-catenin in cells (38–40).
In our study, the protein expression of Wnt3a, p-β-catenin and
GSK-3β was significantly increased in OA chondrocytes, indicating
the activation of the Wnt/β-catenin pathway in OA. Interestingly,
the activation of Wnt/β-catenin signaling pathway by transfecting
Wnt3a inhibited chondrocyte proliferation but promoted cell
apoptosis in our study, and more importantly, Wnt3a could reverse
the effect of silencing RIPK4 on chondrocyte proliferation and
apoptosis. Of note, RIPK4 can bind with LRP6 to stabilize the
accumulation of intracellular β-catenin, which was transferred into
the nucleus to enhance classical Wnt3a pathway, and thereby
promoting tumor cell growth as shown by Huang et al
(15). Besides, Zhou et al
reported that Tetrandrine can reduce the levels of MMPs and
inflammatory factors in OA chondrocytes by inhibiting the
upregulation of Wnt/β-catenin pathway, and thus, exerting a
protective effect on chondrocytes (41). In addition, Chen et al also
reported that the over-expression of EZH2 in OA chondrocytes can
elevate the H3K27me3 level on SFRP1 promoter to activate
Wnt/β-catenin pathway, while inhibiting EZH2 can effectively reduce
the activity of Wnt/β-catenin pathway and delay the progression of
OA (42). Similarly, our study
also observed that silencing RIPK4 appreciably reduced Wnt3a,
β-catenin, and p-GSK-3β expressions in OA chondrocytes, suggesting
that downregulating RIPK4 effectively inhibited Wnt/β-catenin
pathway and promoted the proliferation and inhibit the apoptosis of
OA chondrocytes by regulating the transcription of downstream
target genes related to cell proliferation and apoptosis, as
demonstrated in Fig. 5. Notably,
previous evidence also pointed out that specifically activation of
Wnt/β-catenin pathway in chondrocytes in adult mice resulted in the
chondrocyte differentiation and the development of an OA-like
phenotype via upregulation of chondrocyte marker genes, suggesting
that the Wnt/β-catenin pathway may serve as a potential therapeutic
target in vivo (43). While
RIPK4 could mediate Wnt/β-catenin pathway in our study, thus we
made the hypotheses that RIPK4 might to be a potential new target
for the treatment of OA, which we need to be investigated in depth
in the future studies. In summary, our study found the increased
expression of RIPK4 in OA. Besides, silencing RIPK4 expression in
OA chondrocytes can suppress the activation of Wnt/β-catenin
pathway, hence promoting the proliferation and inhibiting the
apoptosis of chondrocytes, to exert protective roles in OA.
Acknowledgements
The authors appreciate the reviewers for their
suggestive and useful comments in this study.
References
|
1
|
Collison J: Osteoarthritis: Removing old
chondrocytes to combat disease. Nat Rev Rheumatol. 73:3882017.
View Article : Google Scholar
|
|
2
|
Johnson VL and Hunter DJ: The epidemiology
of osteoarthritis. Best Pract Res Clin Rheumatol. 28:5–15. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lawrence RC, Felson DT, Helmick CG, Arnold
LM, Choi H, Deyo RA, Gabriel S, Hirsch R, Hochberg MC, Hunder GG,
et al: Estimates of the prevalence of arthritis and other rheumatic
conditions in the United States. Part II. Arthritis Rheum.
58:26–35. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gomes-Neto M, Araujo AD, Junqueira ID,
Oliveira D, Brasileiro A and Arcanjo FL: Comparative study of
functional capacity and quality of life among obese and non-obese
elderly people with knee osteoarthritis. Rev Bras Reumatol Engl Ed.
56:126–130. 2016.(In English, Portuguese). PubMed/NCBI
|
|
5
|
Iannone F and Lapadula G: The
pathophysiology of osteoarthritis. Aging Clin Exp Res. 15:364–372.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Neogi T and Zhang Y: Epidemiology of
osteoarthritis. Rheum Dis Clin North Am. 39:1–19. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Michael JW, Schluter-Brust KU and Eysel P:
The epidemiology, etiology, diagnosis, and treatment of
osteoarthritis of the knee. Dtsch Arztebl Int. 107:152–162.
2010.PubMed/NCBI
|
|
8
|
Alcaraz MJ, Megias J, Garcia-Arnandis I,
Clérigues V and Guillén MI: New molecular targets for the treatment
of osteoarthritis. Biochem Pharmacol. 80:13–21. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang D, Lin J and Han J:
Receptor-interacting protein (RIP) kinase family. Cell Mol Immunol.
7:243–249. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bhr C, Rohwer A, Stempka L, Rincke G,
Marks F and Gschwendt M: DIK, a novel protein kinase that interacts
with protein kinase Cdelta. Cloning, characterization, and gene
analysis. J Biol Chem. 275:36350–36357. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen L, Haider K, Ponda M, Cariappa A,
Rowitch D and Pillai S: Protein kinase C-associated kinase (PKK), a
novel membrane-associated, ankyrin repeat-containing protein
kinase. J Biol Chem. 276:21737–21744. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Meylan E, Martinon F, Thome M, Gschwendt M
and Tschopp J: RIP4 (DIK/PKK), a novel member of the RIP kinase
family, activates NF-kappa B and is processed during apoptosis.
EMBO Rep. 3:1201–1208. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Meylan E and Tschopp J: The RIP kinases:
Crucial integrators of cellular stress. Trends Biochem Sci.
30:151–159. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Adams S and Munz B: RIP4 is a target of
multiple signal transduction pathways in keratinocytes:
Implications for epidermal differentiation and cutaneous wound
repair. Exp Cell Res. 316:126–137. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang X, McGann JC, Liu BY, Hannoush RN,
Lill JR, Pham V, Newton K, Kakunda M, Liu J, Yu C, et al:
Phosphorylation of Dishevelled by protein kinase RIPK4 regulates
Wnt signaling. Science. 339:1441–1445. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu DQ, Li FF, Zhang JB, Zhou TJ, Xue WQ,
Zheng XH, Chen YB, Liao XY, Zhang L, Zhang SD, et al: Increased
RIPK4 expression is associated with progression and poor prognosis
in cervical squamous cell carcinoma patients. Sci Rep. 5:119552015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rountree RB, Willis CR, Dinh H, Blumberg
H, Bailey K, Dean C Jr, Peschon JJ and Holland PM: RIP4 regulates
epidermal differentiation and cutaneous inflammation. J Invest
Dermatol. 130:102–112. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Adams S, Pankow S, Werner S and Munz B:
Regulation of NF-kappaB activity and keratinocyte differentiation
by the RIP4 protein: Implications for cutaneous wound repair. J
Invest Dermatol. 127:538–544. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rose J, Söder S, Skhirtladze C, Schmitz N,
Gebhard PM, Sesselmann S and Aigner T: DNA damage, discoordinated
gene expression and cellular senescence in osteoarthritic
chondrocytes. Osteoarthritis Cartilage. 20:1020–1028. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
The helsinki declaration of the world
medical association (WMA), . Ethical principles of medical research
involving human subjects. Pol Merkur Lekarski. 36:298–301. 2014.(In
Polish). PubMed/NCBI
|
|
21
|
Dias RC, Dias JM and Ramos LR: Impact of
an exercise and walking protocol on quality of life for elderly
people with OA of the knee. Physiother Res Int. 8:121–130. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tuo YL, Li XM and Luo J: Long noncoding
RNA UCA1 modulates breast cancer cell growth and apoptosis through
decreasing tumor suppressive miR-143. Eur Rev Med Pharmacol Sci.
19:3403–3411. 2015.PubMed/NCBI
|
|
23
|
Ebrahimzadeh MH, Makhmalbaf H,
Birjandinejad A and Soltani-Moghaddas SH: Cross-cultural adaptation
and validation of the persian version of the oxford knee score in
patients with knee osteoarthritis. Iran J Med Sci. 39:529–535.
2014.PubMed/NCBI
|
|
24
|
Holland P, Willis C, Kanaly S, Glaccum M,
Warren A, Charrier K, Murison J, Derry J, Virca G, Bird T and
Peschon J: RIP4 is an ankyrin repeat-containing kinase essential
for keratinocyte differentiation. Curr Biol. 12:1424–1428. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kwa MQ, Scholz GM and Reynolds EC: RIPK4
activates an IRF6-mediated proinflammatory cytokine response in
keratinocytes. Cytokine. 83:19–26. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Azizmohammadi S, Azizmohammadi S, Safari
A, Kaghazian M, Sadrkhanlo M, Behnod V and Seifoleslami M:
High-level expression of RIPK4 and EZH2 contributes to lymph node
metastasis and predicts favorable prognosis in patients with
cervical cancer. Oncol Res. 25:495–501. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang Z, Li J, Du S, Chen G, Qi Y, Huang
L, Xiao L and Tong P: Effects of UCP4 on the proliferation and
apoptosis of chondrocytes: Its possible involvement and regulation
in osteoarthritis. PLoS One. 11:e01506842016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Schroeppel JP, Crist JD, Anderson HC and
Wang J: Molecular regulation of articular chondrocyte function and
its significance in osteoarthritis. Histol Histopathol. 26:377–394.
2011.PubMed/NCBI
|
|
29
|
Zhong JH, Li J, Liu CF, Liu N, Bian RX,
Zhao SM, Yan SY and Zhang YB: Effects of microRNA-146a on the
proliferation and apoptosis of human osteoarthritis chondrocytes by
targeting TRAF6 through the NF-κB signalling pathway. Biosci Rep.
37:pii: BSR201605782017. View Article : Google Scholar
|
|
30
|
Wang Y, Li YP, Paulson C, Shao JZ, Zhang
X, Wu M and Chen W: Wnt and the Wnt signaling pathway in bone
development and disease. Front Biosci (Landmark Ed). 19:379–407.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lad EM, Cheshier SH and Kalani MY:
Wnt-signaling in retinal development and disease. Stem Cells Dev.
18:7–16. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Corr M: Wnt-beta-catenin signaling in the
pathogenesis of osteoarthritis. Nat Clin Pract Rheumatol.
4:550–556. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Johnson ML and Kamel MA: The Wnt signaling
pathway and bone metabolism. Curr Opin Rheumatol. 19:376–382. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Reinhold MI, Kapadia RM, Liao Z and Naski
MC: The Wnt-inducible transcription factor Twist1 inhibits
chondrogenesis. J Biol Chem. 281:1381–1388. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nalesso G, Sherwood J, Bertrand J, Pap T,
Ramachandran M, De Bari C, Pitzalis C and Dell'accio F: WNT-3A
modulates articular chondrocyte phenotype by activating both
canonical and noncanonical pathways. J Cell Biol. 193:551–564.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Funck-Brentano T, Bouaziz W, Marty C,
Geoffroy V, Hay E and Cohen-Solal M: Dkk-1-mediated inhibition of
Wnt signaling in bone ameliorates osteoarthritis in mice. Arthritis
Rheumatol. 66:3028–3039. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rockel JS, Yu C, Whetstone H, Craft AM,
Reilly K, Ma H, Tsushima H, Puviindran V, Al-Jazrawe M, Keller GM
and Alman BA: Hedgehog inhibits β-catenin activity in synovial
joint development and osteoarthritis. J Clin Invest. 126:1649–1663.
2016. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Delgado E, Bahal R, Yang J, Lee JM, Ly DH
and Monga SP: β-Catenin knockdown in liver tumor cells by a cell
permeable gamma guanidine-based peptide nucleic acid. Curr Cancer
Drug Targets. 13:867–878. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gosens R, Meurs H and Schmidt M: The
GSK-3/beta-catenin-signalling axis in smooth muscle and its
relationship with remodelling. Naunyn Schmiedebergs Arch Pharmacol.
378:185–191. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cui XP, Xing Y, Chen JM, Dong SW, Ying DJ
and Yew DT: Wnt/beta-catenin is involved in the proliferation of
hippocampal neural stem cells induced by hypoxia. Ir J Med Sci.
180:387–393. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhou X, Li W, Jiang L, Bao J, Tao L, Li J
and Wu L: Tetrandrine Inhibits the Wnt/β-catenin signalling pathway
and alleviates osteoarthritis: An in vitro and in vivo study. Evid
Based Complement Alternat Med. 2013:8095792013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen L, Wu Y, Wu Y, Wang Y, Sun L and Li
F: The inhibition of EZH2 ameliorates osteoarthritis development
through the Wnt/β-catenin pathway. Sci Rep. 6:291762016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhu M, Tang D, Wu Q, Hao S, Chen M, Xie C,
Rosier RN, O'Keefe RJ, Zuscik M and Chen D: Activation of
beta-catenin signaling in articular chondrocytes leads to
osteoarthritis-like phenotype in adult beta-catenin conditional
activation mice. J Bone Miner Res. 24:12–21. 2009. View Article : Google Scholar : PubMed/NCBI
|