Introduction
Salivary gland tumors are a rare group of tumors
that comprise almost 5% of head and neck cancers and ~0.5% of all
malignancies (1) and they differ
in their responsiveness to anticancer therapies because of their
heterogeneities in their phenotypic, biological and clinical
behavior (2). Mucoepidermoid
carcinoma (MEC) is the most common malignant salivary gland tumor
in adults and children and usually affects the parotid and minor
salivary glands. Surgery is the most common treatment for MEC, but
often results in diminishing the quality of life by devastating
functional and cosmetic consequences (3). Thus, it is necessary to develop
promising chemotherapeutic agents to kill residual tumor cells and
prevent the recurrence of MEC.
For centuries, medicinal natural products have been
regarded as precious sources of chemotherapeutic agents, and still
lots of medicines are originated from plant-derived natural
products (4,5). Over the past, plant-derived natural
products or their single components have been currently employed
for the treatments of various diseases (6,7).
They have anti-neoplastic potentials as suitable candidates for
extending the range of therapeutic options for several types of
cancers (8). Recently, our
research demonstrated that natural products induce apoptotic cell
death in human oral cancer cell lines. For examples, methanol
extracts of Dianthus chinensis and Acalypha australis
L. target specificity protein 1 (Sp1) to induce apoptosis (9) and Impatiens balsamina L.
promotes apoptosis by downregulation of Akt pathway in human oral
cancer cells (10). Several papers
also reviewed the beneficial of natural products for the treatment
of oral cancer, for which current treatment approaches have not
succeeded in improving long-term clinical outcome (11,12).
These suggest that natural products may be promising
chemotherapeutic drug candidates for treatment of oral cancer.
Potentilla discolor of the rosaceae commonly founded in the
north temperate, is a noticed traditional Chinese medicine
(13,14). It has been also reported to improve
the complications of diabetes mellitus in both experimental animals
and clinical trials (15).
However, the anticancer role of Potentilla discolor in human
mucoepidermoid carcinoma (MEC) cell lines still remains to be
determined. Our main objectives were to assess antitumor effects of
methanol extract of Potentilla discolor (MEPD) and decipher
the molecular mechanism involved in its antitumor activity in human
MEC cells.
Materials and methods
Cell culture and chemical
treatment
Salivary gland mucoepidermoid carcinoma MC3 cells
which were kindly provided by Professor Wu Junzheng in the Forth
Military Medical University (Xi'an, China), were isolated by
repeated in situ-transplants of MEC-1 human mucoepidermoid
carcinoma cell line derived from palatal salivary gland as
described in the study by Wen et al (16). YD15 cells were obtained from
Professor Jin Kim in Yonsei University (Seoul, Korea), which was
derived from salivary gland mucoepidermoid carcinoma of tongue
origin (17). MC3 cells were grown
in DMEM and YD15 cells were grown in RPMI-1640; both types of media
were supplemented with 10% fetal bovine serum (FBS) and 100 U/ml
each of penicillin and streptomycin in a humidified atmosphere
containing 5% CO2 at 37°C. An equal number of cells were
seeded and allowed to attach. All experiments were performed in
cells cultured at 50–60% confluence. Methanol extract of
Potentilla discolor (MEPD) was supplied from Korea Plant
Extract Bank at the Korea Research Institute of Bioscience and
Biotechnology (Cheongju, Korea), and cryptotanshinone was purchased
from Sigma-Aldrich (St. Louis, MO, USA). Each chemical was
dissolved in dimethyl sulfoxide (DMSO), aliquoted, and stored at
−20°C.
Trypan blue exclusion assay
The cytotoxicity of MEPD or cryptotanshinone was
determined with trypan blue solution (Gibco, Paisley, UK).
Harvested cells were stained with trypan blue (0.4%) and then
viable cells were counted using a hemocytometer.
Western blot analysis
Whole-cell lysates were prepared with lysis buffer
and protein concentration in each sample was measured using a DC
Protein Assay Kit (Bio-Rad Laboratories, Madison, WI, USA). After
normalization, equal amounts of protein were separated by SDS-PAGE
and then transferred to Immuno-Blot PVDF membranes. The membranes
were blocked with 5% skim milk in TBST at room temperature (RT) for
2 h, and incubated with primary antibodies and corresponding
HRP-conjugated secondary antibodies (1:5,000; catalog no. SC-2004
for anti-rabbit and SC-2020 for anti-goat). Rabbit anti-human
polyclonal antibodies against cleaved PARP (1:3,000; catalog no.
9541), p-STAT3 (1:1,000; catalog no. 9145), total STAT3 (1:3,000;
catalog no. 4904) and PUMA (1:1,000; catalog no. 4976) were
purchased from Cell Signaling Technology, Inc. (Charlottesville,
VA, USA). p53 antibody (1:2,000; catalog no. OP43) was obtained by
Calbiochem (San Diego, CA, USA). Goat anti-human actin polyclonal
antibody (1:5,000; catalog no. SC-1615) was obtained from Santa
Cruz Biotechnology, Inc., (Santa Cruz, CA, USA). The immunoreactive
bands were visualized by ImageQuant LAS 500 (GE Healthcare Life
Sciences, Piscataway, NJ, USA).
4′-6-Diamidino-2-phenylindole (DAPI)
staining
To detect nuclear morphological changes of apoptotic
cells, cells were stained with DAPI solution (Sigma-Aldrich).
Briefly, cells were fixed in 100% methanol at RT for 10 min,
deposited on slides, and stained with DAPI solution (2 µg/ml). The
morphological changes of apoptotic cells were observed under a
fluorescence microscopy.
Live/dead assay
The cytotoxicity of MEPD or cryptotanshinone were
determined using Live/Dead Viability/Cytotoxicity assay (Life
Technologies, Grand Island, NY, USA). The polyanionic dye
calcein-AM is retained within live cells, producing an intense
green fluorescence through intracellular esterase activity.
Ethidium homodimer-1 enters cells with damaged membranes and binds
to nucleic acids, producing a bright red fluorescence in dead
cells. Briefly, cells were stained with 2 µM calcein-AM and 4 µM
ethidium homodimer-1, and then incubated for 30 min at RT. Cells
were analyzed under a fluorescence microscopy.
Immunofluorescence staining
MC3 and YD15 cells were seeded on 4-well culture
plate and treated with DMSO or MEPD. After 24 h, cells were fixed
and permeabilized using the cytofix/cytoperm solution for 1 h at
4°C. Cells were then blocked with 1% bovine serum albumin (BSA) in
PBS for 1 h at RT and incubated overnight at 4°C with antibodies
against p-STAT3 or PUMA. Subsequently, the cells were exposed to
the FITC-conjugated secondary antibody for 1 h at RT and were
visualized using a fluorescence microscope equipped with the
appropriate filters for DAPI and FITC dyes.
Construction of STAT3 overexpression
vector and transient transfection
The open reading frame of human STAT3 (NM_139276)
was amplified from cDNA that was synthesized in HSC3 cells using
the specific primers of the gene (primer sequence; STAT3 sense,
5′-GATATCATGGCCCAATGGAATCAG-3′, with an included EcoRV site, STAT3
antisense, 5′-GATATCTCACATGGGGGAGGTAGC-3′, with an included EcoRV
site), and then cloned into pGEM® T-easy vector
(Promega, Madison, WI, USA). The STAT3 was confirmed by sequence
analysis. Finally, the gene was cloned into the multi cloning site
of pcDNA3.1 (+) vector (Invitrogen, San Diego, CA, USA). MC3 and
YD15 cells were transfected by two kinds of vector constructs
(pcDNA3.1; pcDNA3.1-STAT3) by Lipofectamine 3000 transfection
reagent (Life Technologies, Carlsbad, CA, USA) according to the
manufacturer's instructions, respectively.
Statistical analysis
Student's t-test or one-way ANOVA followed by
Tukey's post hoc test were used to determine the significance of
differences between the control and treatment groups; P-values of
<0.05, 0.01 or 0.001 are considered significant.
Results
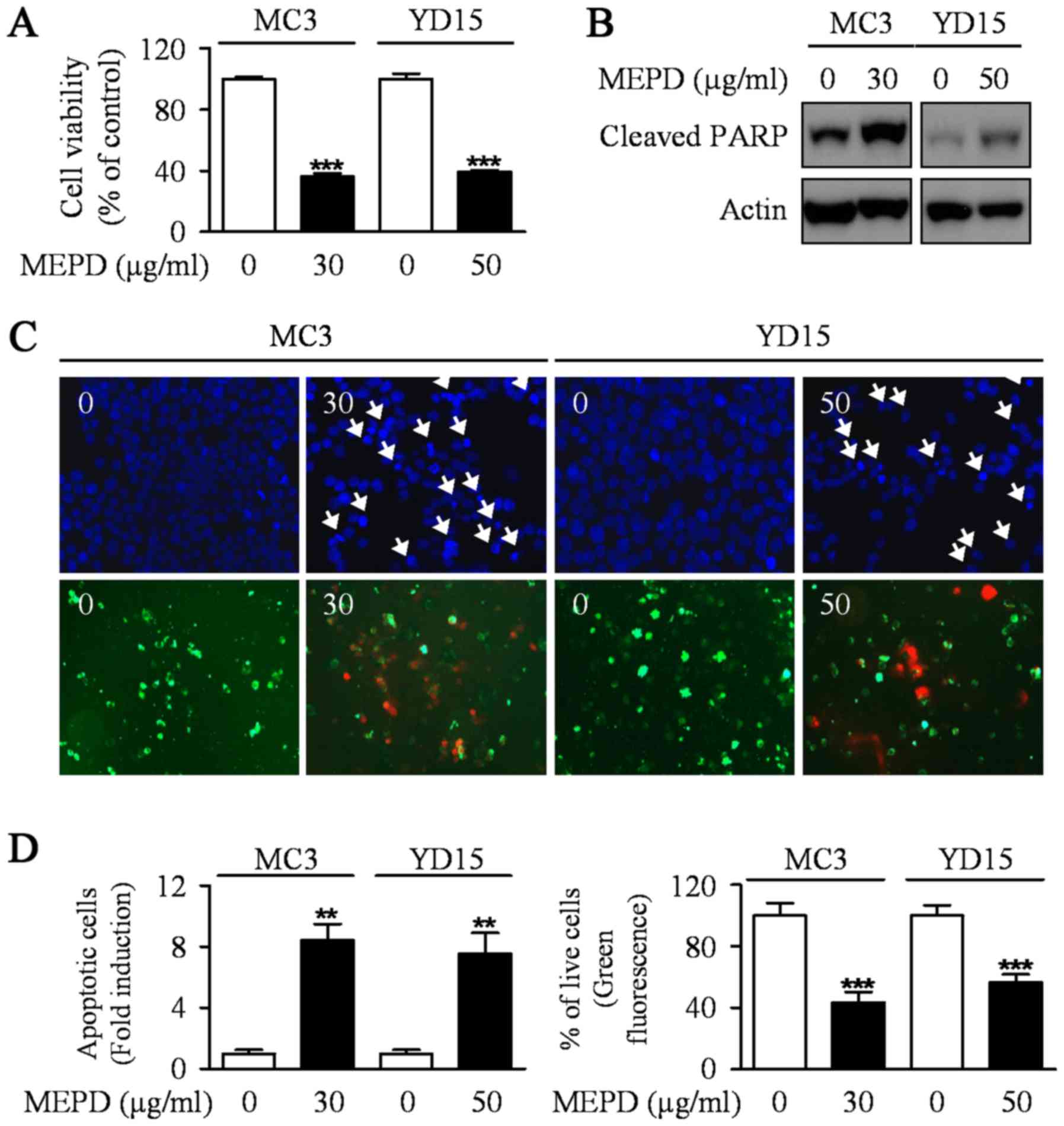
MEPD suppresses the growth of human
mucoepidermoid carcinoma cells of salivary gland by inducing
apoptotic cell death
To explore the potential antitumor activities of
MEPD in MC3 and YD15 human mucoepidermoid carcinoma (MEC) cell
lines, we evaluated the growth-inhibitory effects of MEPD using
trypan blue exclusion assay after treatment of MEPD (30 µg/ml for
MC3 cells and 50 µg/ml for YD15 cells) for 48 h. MEPD treatment
significantly reduced cell viability in both cells relative to the
controls (Fig. 1A). In order to
demonstrate the effect of MEPD on apoptosis of MEC cells, we
performed western to detect the protein expression level of PARP in
both MC3 and YD15 cells. As presented in Fig. 1B, PARP cleaved in both cell lines
treated with MEPD. Moreover, qualitative evaluation of apoptotic
cell death in MEPD-treated cells was made by DAPI staining
(Fig. 1C, upper; Fig. 1D, left) and Live/dead assay
(Fig. 1C, down; Fig. 1D, right). The results showed that
DNA-fragmented or EthD-1-stained (red fluorescence) cells were
observed in MEPD-treated group indicating that apoptosis may be
involved in MEPD-mediated growth suppression in MEC cells of
salivary gland.
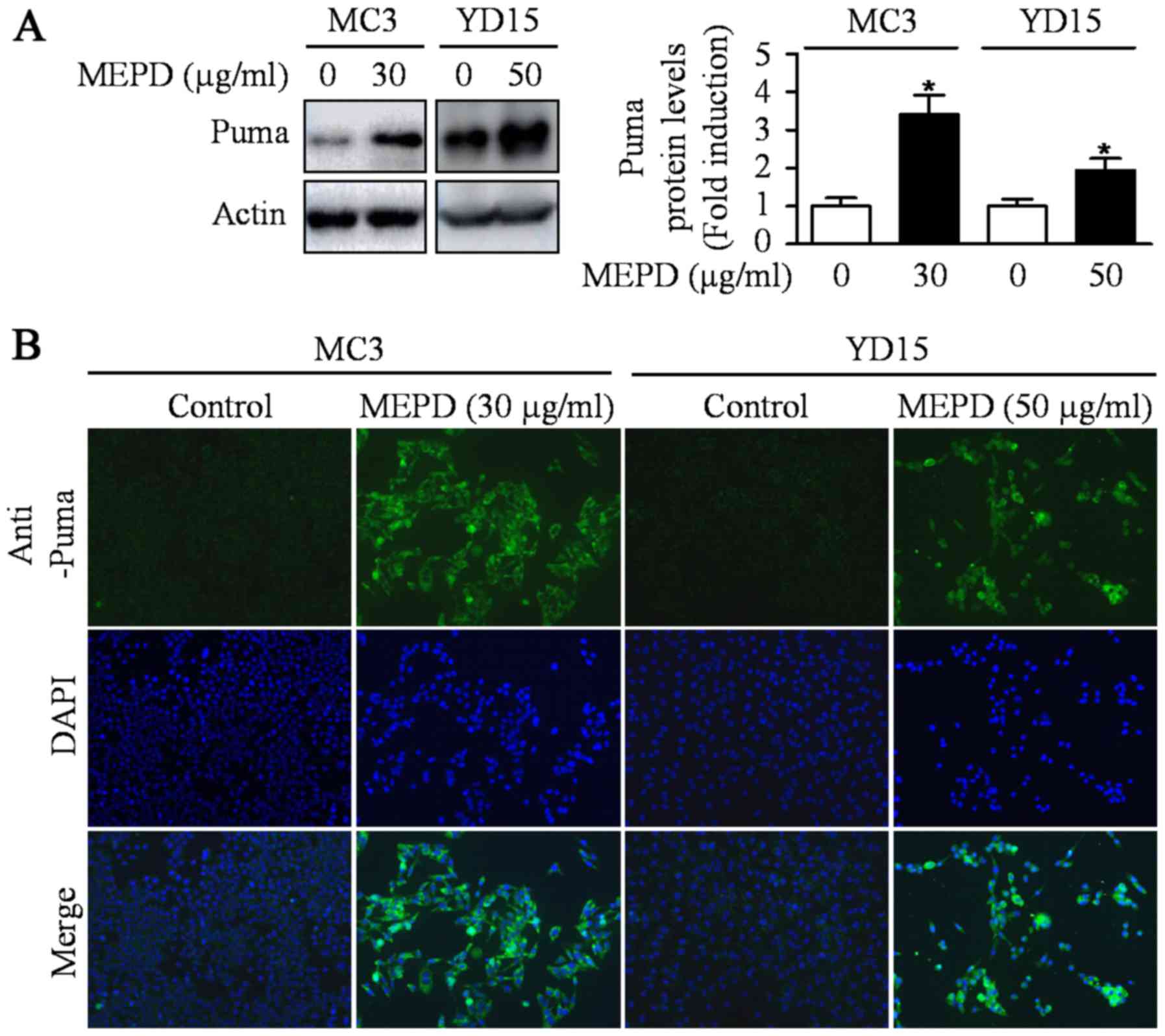
MEPD induces PUMA, but inactivates
STAT3
PUMA induced by natural compound is known as one of
the main mediators for apoptosis of cancer cells (18). A previous study also reported that
exogenous expression of PUMA resulted in suppression of tumor
growth by inducing apoptosis in an orthotopic mouse model of oral
cancer (19). To investigate
whether PUMA is related to MEPD-induced apoptosis, we detected
expression level of PUMA protein in MEPD-treated both MEC cell
lines. As shown in Fig. 2A, MEPD
markedly induced PUMA protein expression. We also visually observed
the increase in PUMA expression in MEPD-treated cells using
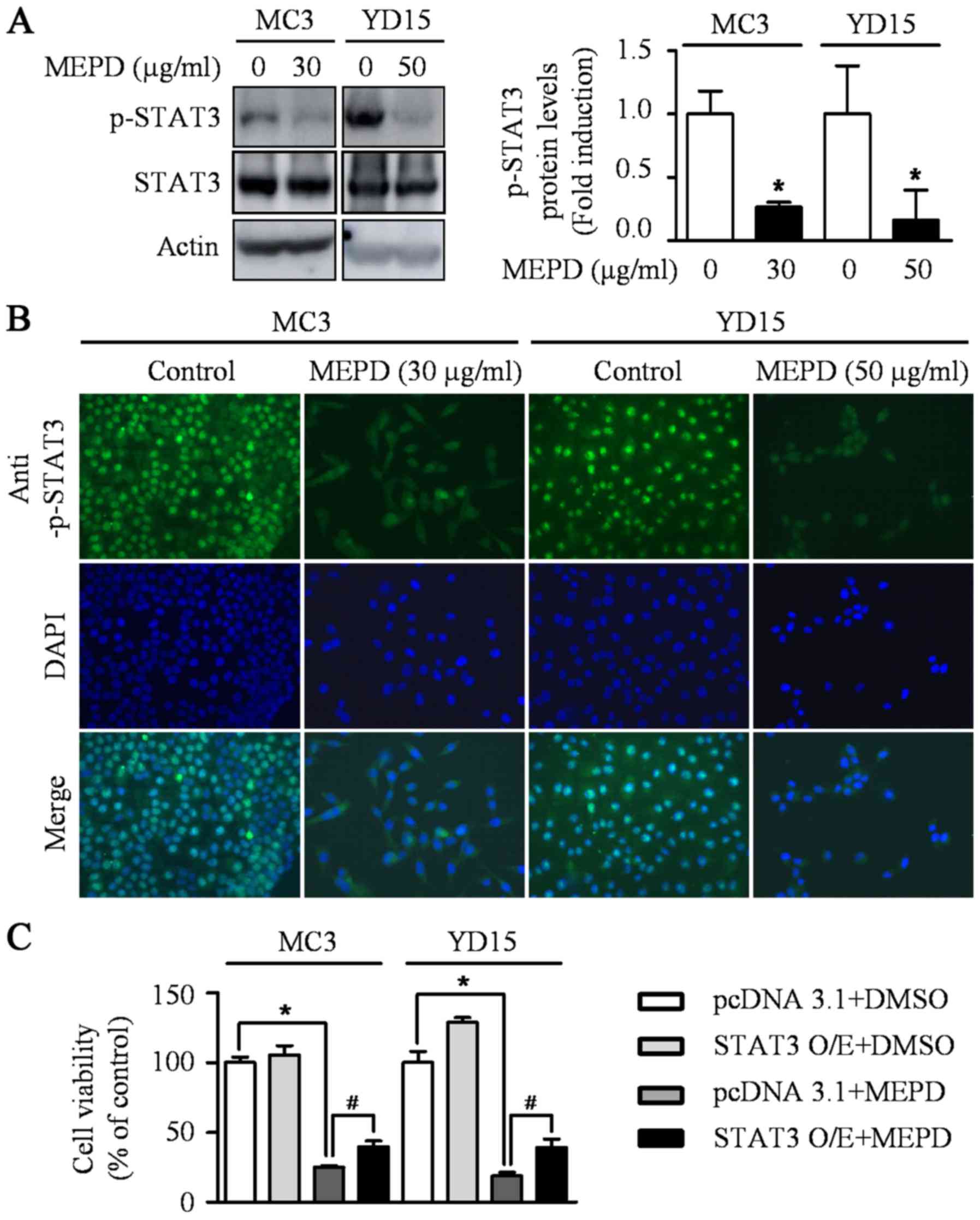
immunofluorescence staining compared to the controls (Fig. 2B). Several recent reports
demonstrated that STAT3 represents a target for therapeutic
intervention in oral cancer (20–22).
Thus, STAT3 signaling was examined after the treatment of MEPD in
MEC cell lines. The results demonstrated that MEPD significantly
inhibited the expression of phospho-STAT3 evidenced by western
blotting and immunofluorescence analysis (Fig. 3A and B). To further clarify the
functional role of STAT3 on cell viability in MEC cell lines, we
transfected STAT3 overexpression vector into both MEC cell lines.
The results showed that STAT3 overexpression slightly brought back
MEPD-mediated growth inhibition of MC3 and YD15 cells, but it was
statistically significant (Fig.
3C). These results suggest that MEPD-caused growth inhibition
may be due to both PUMA and STAT3 signaling in MEC cell lines.
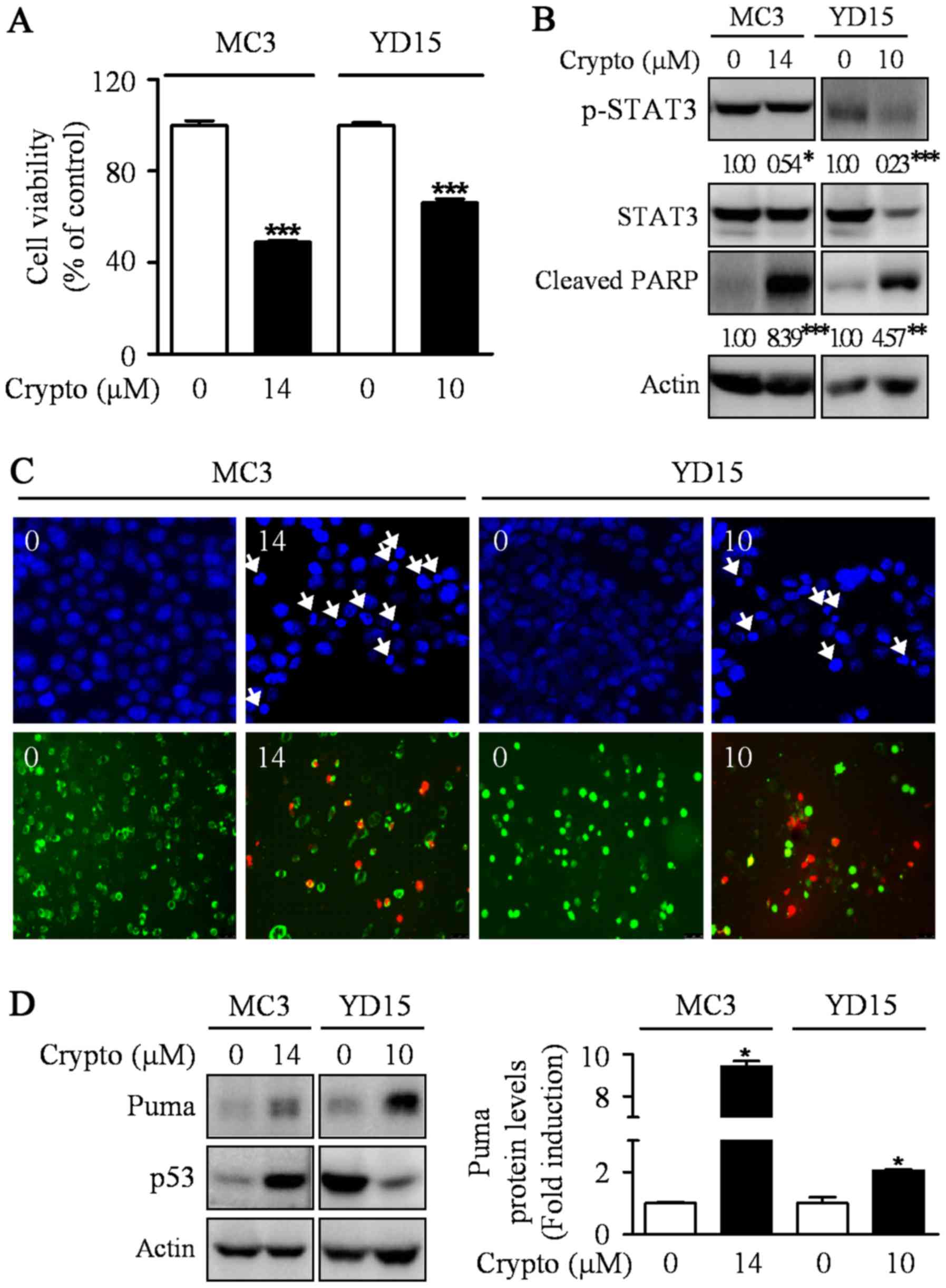
Inactivating STAT3 by cryptotanshinone
leads to apoptotic cell death by inducing PUMA expression
Previously, the abrogation of STAT3 was sufficient
to induce p53-mediated apoptosis and PUMA is a key mediator in
p53-mediated apoptosis (23,24).
Hence, we hypothesized that STAT3 may be involved in the induction
of PUMA protein during MEPD-mediated apoptosis in MEC cell lines.
To prove our hypothesis, we used a potent STAT3 inhibitor,
cryptotanshinone. The results showed the dephosphorylation of STAT3
by cryptotanshinone significantly inhibited cell growth and
increased apoptotic cell death in both MEC cell lines by enhancing
the expression level of PUMA protein (Fig. 4). We also investigated whether
cryptotanshinone affects p53 protein as an upstream molecule of
Puma in MC3 and YD15 cells. However, p53 was not consistently
affected by cryptotanshinone in both cell lines. In addition, we
checked protein levels of Puma in STAT3 overexpression cells to
clarify the relationship between STAT3 and Puma. The result showed
that STAT3 overexpression slightly reduced MEPD-mediated Puma
expression in both cell lines (data not shown). These results
suggest that blocking STAT3 signaling pathway may contribute to
apoptosis via induction of PUMA in both MC3 and YD15 cells.
Discussion
In order to kill the remaining cancer cells and
prevent the recurrence, chemotherapy is essential following
surgery. More than 50% of all available drugs on the market derived
from natural sources including plants, microbes and marine life, of
which over 70% of anticancer drugs have their origin from natural
resources (25). Naturally
Occurring Plant-based Anticancer Compound-Activity-Target database
(NPACT) provide a thousand of natural resources exhibiting
antitumorigenic activities which are experimentally validated
(in vitro or in vivo) in various cancers (26). Thus, the discovery of
pharmacologically active natural resources with antitumorigenic
activity is very important for developing a new cancer medicine. A
number of plant-derived natural compounds also has potential drug
carriers for the treatment of oral cancer. For example,
Impatiens balsamina L., a traditional herb medicine for the
treatment of rheumatism, swelling and fingernail inflammation,
regulates either AMPK or Akt signaling to promote apoptotic
response in human oral cancer cell lines (10,27).
Our prior study has reported that Smilax china L. can induce
apoptosis via the ERK signaling pathway to treat human MEC of
salivary gland (28) and it gives
notice of importance that natural compounds can be promising
resources of new drugs for MEC in the future. In this study, we
showed that one of natural resources, Potentilla discolor,
caused blocked cell growth and induced apoptotic cell death in both
MC3 and YD15 MEC cell lines. Recently, several Potentilla
discolor-derived active components induced apoptosis in human
HepG-2 hepatocellular carcinoma, MCF-7 breast adenocarcinoma and
T-84 colon carcinoma cell lines (29). This is consistent with our findings
that Potentilla discolor can have an ability to induce
apoptosis in various cancers including MEC.
Natural products with anticancer properties may have
various targets inside human cancer cells, but they often regulate
the members of Bcl-2 family which play a pivotal role in a
programmed cell death process (30). Our group recently found that
parthenolide increased Bim protein (28) and pycnogenol increased Bak protein
by increasing its protein stability (31), resulting in the induction of
apoptosis in human oral cancer supporting that natural
products-derived compounds can influence Bcl-2 family members to
exert their apoptotic activities in oral cancer. PUMA (p53
upregulated modulator of apoptosis) is one of BH3-only subgroup of
the Bcl-2 family. A considerable amount of literature published on
PUMA mentioned the involvement of PUMA in stress-induced apoptosis
(32–35). Our results also here suggest that
upregulation of PUMA by MEPD mediates apoptosis in human MEC cell
lines. In addition, we screened the expression levels of other
Bcl-2 family in MEPD-treated both cell lines; yet no Bcl-2 family
proteins except PUMA were commonly regulated by MEPD during
apoptotic condition (data not shown). PUMA was originally known as
a downstream target of p53 protein. However, our western blot data
showed no significant difference of p53 protein level after
treatment of MEPD in human MEC cell lines (data not shown). It
means p53-independent regulation of MEPD on PUMA protein during
MEPD-mediated apoptosis in human MEC cell lines.
Signal transducers and activators of transcription 3
(STAT3) is one of a family of transcription factors that regulate
apoptosis as well as cell proliferation, differentiation, and
angiogenesis and a target for inducing apoptosis in various cancers
(36,37). Several studies have revealed the
critical role of STAT3 in oral cancer development (22,38,39).
As previously stated from our laboratory, the inhibition of STAT3
by cryptotanshinone or sorafenib clearly induced apoptosis in human
MEC cell lines or tumor xenograft (40,41).
In the present study, MEPD clearly inhibited STAT3 to induce
apoptosis in human MEC cell lines and this observation may support
other previous research demonstrating that STAT3 as a promising
target for chemotherapy. In addition, inhibition of STAT3 by a
STAT3 inhibitor (cryptotanshinone) in both cell lines significantly
increased the expression of PUMA. Notably, prior study reported
that STAT5 directly controls the expression of pro-apoptotic
protein PUMA suggesting the possibility of the involvement of STATs
in the regulation of PUMA protein. Although more research is
required to determine direct interaction between STAT3 and PUMA,
this is the first time that STAT3 regulates PUMA to induce
apoptosis in human MEC cell lines.
In conclusion, the present study revealed that MEPD
caused a marked increase in salivary MEC cellular apoptosis in
vitro. The molecular targets of MEPD-induced apoptosis were
PUMA and STAT3. All these results should be useful in the search
for naturally occurring plant-based new potential antitumor agents
for the treatment of MEC.
Acknowledgements
This study was supported by the Basic Science
Research Program through the National Research Foundation of Korea
(NRF) funded by the Ministry of Science ICT & Future Planning
[2017R1A2B2003491].
References
|
1
|
Ettl T, Schwarz-Furlan S, Gosau M and
Reichert TE: Salivary gland carcinomas. Oral Maxillofac Surg.
16:267–283. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Goyal G, Mehdi SA and Ganti AK: Salivary
gland cancers: Biology and systemic therapy. Oncology. 29:773–780.
2015.PubMed/NCBI
|
|
3
|
Zhang L, Li L, Wang Y, Liu Y and Li C: MC3
Mucoepidermoid carcinoma cell line enriched cancer stem-like cells
following chemotherapy. Oncol Lett. 7:1569–1575. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kingston DG: Modern natural products drug
discovery and its relevance to biodiversity conservation. J Nat
Prod. 74:496–511. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Newman DJ and Cragg GM: Natural products
as sources of new drugs over the 30 years from 1981 to 2010. J Nat
Prod. 75:311–335. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hung HY, Qian K, Morris-Natschke SL, Hsu
CS and Lee KH: Recent discovery of plant-derived anti-diabetic
natural products. Nat Prod Rep. 29:580–606. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tarkang PA, Appiah-Opong R, Ofori MF,
Ayong LS and Nyarko AK: Application of multi-target
phytotherapeutic concept in malaria drug discovery: A systems
biology approach in biomarker identification. Biomark Res.
4:252016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gupta SC, Kim JH, Prasad S and Aggarwal
BB: Regulation of survival, proliferation, invasion, angiogenesis,
and metastasis of tumor cells through modulation of inflammatory
pathways by nutraceuticals. Cancer Metastasis Rev. 29:405–434.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shin JA, Kim JJ, Choi ES, Shim JH, Ryu MH,
Kwon KH, Park HM, Seo JY, Lee SY, Lim DW, et al: In vitro apoptotic
effects of methanol extracts of Dianthus chinensis and Acalypha
australis L. targeting specificity protein 1 in human oral cancer
cells. Head Neck. 35:992–998. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shin JA, Ryu MH, Kwon KH, Choi B and Cho
SD: Down-regulation of Akt by methanol extracts of Impatiens
balsamina L. promotes apoptosis in human oral squamous cell
carcinoma cell lines. J Oral Pathol Med. 44:420–428. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zlotogorski A, Dayan A, Dayan D, Chaushu
G, Salo T and Vered M: Nutraceuticals as new treatment approaches
for oral cancer: II. Green tea extracts and resveratrol. Oral
Oncol. 49:502–506. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ramshankar V and Krishnamurthy A:
Chemoprevention of oral cancer: Green tea experience. J Nat Sci
Biol Med. 5:3–7. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang J, Chen H, Zhang L, Wang Q and Lai
MX: Anti-diabetic effect of standardized extract of Potentilla
discolor Bunge and identification of its active components. Drug
Dev Res. 71:127–132. 2010.
|
|
14
|
Zhang L, Yang J, Chen XQ, Zan K, Wen XD,
Chen H, Wang Q and Lai MX: Antidiabetic and antioxidant effects of
extracts from Potentilla discolor Bunge on diabetic rats induced by
high fat diet and streptozotocin. J Ethnopharmacol. 132:518–524.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Song C, Huang L, Rong L, Zhou Z, Peng X,
Yu S and Fang N: Anti-hyperglycemic effect of Potentilla discolor
decoction on obese-diabetic (Ob-db) mice and its chemical
composition. Fitoterapia. 83:1474–1483. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wen DS, Zhu XL, Guan SM, Wu YM, Yu LL and
Wu JZ: Silencing of CXCR4 inhibits the proliferation, adhesion,
chemotaxis and invasion of salivary gland mucoepidermoid carcinoma
Mc3 cells in vitro. Oral Oncol. 44:545–554. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee EJ, Kim J, Lee SA, et al:
Characterization of newly established oral cancer cell lines
derived from six squamous cell carcinoma and two mucoepidermoid
carcinoma cells. Exp Mol Med. 37:379–390. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shankar S, Siddiqui I and Srivastava RK:
Molecular mechanisms of resveratrol
(3,4,5-trihydroxy-trans-stilbene) and its interaction with
TNF-related apoptosis inducing ligand (TRAIL) in
androgen-insensitive prostate cancer cells. Mol Cell Biochem.
304:273–285. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yeh CC, Hsieh HL, Lee J, Jan YH, Lai TC,
Hong CY, Hsiao M and Kuo MY: Polyethylenimine-mediated PUMA gene
delivery to orthotopic oral cancer: Suppression of tumor growth
through apoptosis induction in situ and prolonged survival. Head
Neck. 33:878–885. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang JG, Lu R, Ye XJ, Zhang J, Tan YQ and
Zhou G: Icaritin reduces oral squamous cell carcinoma progression
via the inhibition of STAT3 signaling. Int J Mol Sci. 18(pii):
E1322017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Peng HY, Cheng YC, Hsu YM, Wu GH, Kuo CC,
Liou JP, Chang JY, Jin SL and Shiah SG: MPT0B098, a microtubule
inhibitor, suppresses JAK2/STAT3 signaling pathway through
modulation of SOCS3 stability in oral squamous cell carcinoma. PLoS
One. 11:e01584402016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Brown ME, Bear MD, Rosol TJ, Premanandan
C, Kisseberth WC and London CA: Characterization of STAT3
expression, signaling and inhibition in feline oral squamous cell
carcinoma. BMC Vet Res. 11:2062015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Niu G, Wright KL, Ma Y, Wright GM, Huang
M, Irby R, Briggs J, Karras J, Cress WD, Pardoll D, et al: Role of
Stat3 in regulating p53 expression and function. Mol Cell Biol.
25:7432–7440. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hikisz P and Kiliańska ZM: PUMA, a
critical mediator of cell death-one decade on from its discovery.
Cell Mol Biol Lett. 17:646–669. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chinembiri TN, du Plessis LH, Gerber M,
Hamman JH and du Plessis J: Review of natural compounds for
potential skin cancer treatment. Molecules. 19:11679–11721. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mangal M, Sagar P, Singh H, Raghava GP and
Agarwal SM: NPACT: Naturally occurring plant-based anti-cancer
compound-activity-target database. Nucleic Acids Res.
41:D1124–D1129. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shin JA, Kwon KH and Cho SD:
AMPK-activated protein kinase activation by Impatiens balsamina L.
is related to apoptosis in HSC-2 human oral cancer cells.
Pharmacogn Mag. 11:136–142. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yu HJ, Shin JA, Lee SO, Kwon KH and Cho
SD: Extracellular signal-regulated kinase inhibition is required
for methanol extract of Smilax china L.-induced apoptosis through
death receptor 5 in human oral mucoepidermoid carcinoma cells. Mol
Med Rep. 9:663–668. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang J, Liu C, Huang RZ, Chen HF, Liao
ZX, Sun JY, Xia XK and Wang FX: Three new
C-27-carboxylated-lupane-triterpenoid derivatives from Potentilla
discolor Bunge and their in vitro antitumor activities. PLoS One.
12:e01755022017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Christodoulou MI, Kontos CK, Halabalaki M,
Skaltsounis AL and Scorilas A: Nature promises new anticancer
agents: Interplay with the apoptosis-related BCL2 gene family.
Anticancer Agents Med Chem. 14:375–399. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang IH, Shin JA, Kim LH, Kwon KH and Cho
SD: The caspase 3-dependent apoptotic effect of pycnogenol in human
oral squamous cell carcinoma HSC-3 cells. J Clin Biochem Nutr.
58:40–47. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang S, Zhu Z, Zhang X, Zhang N and Yao Z:
Idelalisib induces PUMA-dependent apoptosis in colon cancer cells.
Oncotarget. 8:6102–6113. 2017.PubMed/NCBI
|
|
33
|
Chatwichien J, Basu S, Budina-Kolomets A,
Murphy ME and Winkler JD: PUMA-dependent apoptosis in NSCLC cancer
cells by a dimeric β-carboline. Bioorg Med Chem Lett. 26:4884–4887.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhu HJ, Liu L, Fan L, Zhang LN, Fang C,
Zou ZJ, Li JY and Xu W: The BH3-only protein Puma plays an
essential role in p53-mediated apoptosis of chronic lymphocytic
leukemia cells. Leuk Lymphoma. 54:2712–2719. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang Y, Xing D and Liu L: PUMA promotes
Bax translocation by both directly interacting with Bax and by
competitive binding to Bcl-XL during UV-induced
apoptosis. Mol Biol Cell. 20:3077–3087. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xiong A, Yang Z, Shen Y, Zhou J and Shen
Q: Transcription factor STAT3 as a novel molecular target for
cancer prevention. Cancers. 6:926–957. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Al Zaid Siddiquee K and Turkson J: STAT3
as a target for inducing apoptosis in solid and hematological
tumors. Cell Res. 18:254–267. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mali SB: Review of STAT3 (Signal
transducers and activators of transcription) in head and neck
cancer. Oral Oncol. 51:565–569. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Leeman-Neill RJ, Seethala RR, Singh SV,
Freilino ML, Bednash JS, Thomas SM, Panahandeh MC, Gooding WE,
Joyce SC, Lingen MW, et al: Inhibition of EGFR-STAT3 signaling with
erlotinib prevents carcinogenesis in a chemically-induced mouse
model of oral squamous cell carcinoma. Cancer Prev Res. 4:230–237.
2011. View Article : Google Scholar
|
|
40
|
Yu HJ, Park C, Kim SJ, Cho NP and Cho SD:
Signal transducer and activators of transcription 3 regulates
cryptotanshinone-induced apoptosis in human mucoepidermoid
carcinoma cells. Pharmacogn Mag. 10 Suppl 3:S622–S629. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yu HJ, Shin JA, Jung JY, Nam JS, Hong IS,
Cho NP and Cho SD: Inhibition of myeloid cell leukemia-1:
Association with sorafenib-induced apoptosis in human
mucoepidermoid carcinoma cells and tumor xenograft. Head Neck.
37:1326–1335. 2015. View Article : Google Scholar : PubMed/NCBI
|