Introduction
Neuropathic pain is a chronic pain state associated
with multiple etiologies, including trauma, diabetes, cancer,
infection and autoimmune pathology (1). The incidence of neuropathic pain is
approximately 7–10% worldwide (2,3).
Patients with neuropathic pain induced by nerve injury exhibit
varying degrees of mechanical and heat hyperalgesia. Additionally,
sleep and anxiety disorders are more prevalent in individuals with
neuropathic pain, and quality of life is more seriously impaired,
compared with individuals with chronic non-neuropathic pain
(4,5). Chronic neuropathic pain also results
in considerable social and economic burden, leading to high medical
costs and reduced productivity (6,7).
The mechanism underlying neuropathic pain remains
unclear and it is still difficult to treat, despite the development
of various novel therapies (8).
The rapid development of gene chip technology has become a
technical basis for biological research; thousands of genes are
able to be detected simultaneously with gene chip analysis. At
present, large amounts of data have not been effectively explored,
despite the availability of large public gene chip databases.
Bioinformatics has revealed a considerable number of complex
biological interactions through the comprehensive utilization of
biology, computer science and statistics (9).
Vega-Avelaira et al (10) used a microarray expression profile
to identify differentially expressed genes (DEGs) associated with
neuropathic pain. However, the quality of one gene chip studied was
not verified. Therefore, for quality control, it should be removed
for reanalysis of the normalized unscaled standard errors (NUSE)
and RNA degradation curves in the microarray expression profile, as
performed by the affy package. The present study reanalyzed the
gene expression profile from the Vega-Avelaira et al
(10) study to further identify
the DEGs associated with neuropathic pain. Function and pathway
enrichment analyses of the DEGs were performed, followed by
protein-protein interaction (PPI) network construction.
Furthermore, reverse transcription-quantitative polymerase chain
reaction was performed in a mouse model of neuropathic pain to
verify the results obtained from the bioinformatics analysis.
Materials and methods
Microarray data
The gene expression profile GSE15041 was downloaded
from the Gene Expression Omnibus (GEO) of the National Center for
Biotechnology Information (http://www.ncbi.nlm.nih.gov/gds/) on the Affymetrix
Rat Genome 230 2.0 Array (Thermo Fisher Scientific, Inc., Waltham,
MA, USA). A total of 9 adult rat samples were analyzed in the
present study, including 3 samples from the ipsilateral spinal cord
of the sham group (Sham group), 3 samples from the contralateral
spinal cord of the surgery group (Contra group) and 3 samples from
the ipsilateral spinal cord of the surgery group (Ipsi group).
Additionally, the probe annotation package (rat2302.db) of GSE15041
was downloaded from Bioconductor to convert the probe names to gene
names (11).
Data preprocessing
Quality control of the gene chip was performed
through weighting and residual plot, relative log expression (RLE)
box plot, NUSE box plot, principal components plot (PCA) and RNA
degradation curve to remove unqualified samples (12,13).
The Robust Multi-array Average integrative algorithm was chosen for
preprocessing the microarray data, including background correction,
quartile data normalization and probe summarization. Following
this, the boxplot of the raw and normalized data was drawn
(14). The annotate package in R
(15) was subsequently used to
convert probe names to gene names. For each sample, a maximum
expression value was used for the genes with multiple probe
names.
Identification of DEGs
The limma package in R (16) was used to screen out the DEGs
(17). Genes with adjusted P value
of P<0.05 and |fold change| >2 were selected as DEGs. Then,
cluster analysis and heat map construction was performed using the
Pheatmap package (https://cran.r-project.org/package=pheatmap) in R to
further analyze the DEGs.
Enrichment analysis of DEGs
Gene function annotation is usually performed by
Gene ontology (GO) analysis (18).
The GO database consists of three categories, including biological
process, molecular function and cellular component. The Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathways database is a
comprehensive and recognized database encompassing numerous
biochemical pathways. In order to gain further insight into the
function and pathways of the identified DEGs, the Database for
Annotation, Visualization and Integrated Discovery (DAVID; version
6.8) tool (19) was used to
perform the GO and KEGG pathway enrichment analysis. The threshold
was set as P<0.05 and enrichment counts at >2.
PPI network analysis of DEGs
The Search Tool for the Retrieval of Interacting
Genes (STRING; version 10.0; http://www.string-db.org/) provides comprehensive and
easily accessible interaction information derived from known and
predicted protein data (20).
Interactions with a combined score of >0.4 were selected and a
network was built using the STRING online tool. Networks were
visualized using Cytoscape software (21) to identify hub proteins and key
genes associated with neuropathic pain. Nodes with higher degrees
(connected nodes) were considered hub proteins in the PPI network
and may have an essential role in the network.
Animals
Male 8-week-old C57BL/6J (weight, 20–23 g; n=10)
purchased from Model Animal Research Center of Nanjing University
(Nanjing, China) were used in the present study and were randomly
divided into the sham group and surgery group. All animals were
housed in a climate-controlled room (temperature: 22±2°C; humidity:
50–70%) with a 12 h light/dark cycle for at least 2 weeks, with
ad libitum access to food and water. All animal experiments
were approved by the Experimental Animals Welfare and Ethical
Inspection of Nanjing Drum Tower Hospital (Nanjing, China).
Neuropathic pain model
The most widely used model of neuropathic pain is
the unilateral sciatic nerve chronic constriction injury (CCI),
developed by Bennet et al (22). Briefly, animals were anesthetized
with an intraperitoneal injection of 80 mg/kg sodium pentobarbital
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). The skin of the
right hind limb was prepared at the mid-thigh level in a lateral
position prior to surgery. The sciatic nerve was carefully isolated
following skin incision and blunt dissection, two ligatures (5-0
chromic gut suture) were knotted loosely around the sciatic nerve.
The distance between ligatures was approximately 1 mm. The surgery
was the same in the sham group, however the nerve was only exposed
without ligation.
RNA extraction
Mice were deeply anesthetized with sodium
pentobarbital and were rapidly decapitated 7 days post-surgery. The
lumbar (L4-L5) segments of spinal cords were transversely sectioned
and hemi-dissected along the midline. The ipsilateral (CCI-Ipsi)
and contralateral (CCI-Contra) lumbar spinal cord of the surgery
group and the ipsilateral (Sham-Ipsi) lumbar spinal cord of the
sham group was collected. Total RNA was extracted using an RNA
extraction kit (Bioteke Corporation, Beijing, China) according to
the manufacturer's protocol. RNA concentration was determined using
NanoDrop ND-1000 Spectrometer (NanoDrop Technologies; Thermo Fisher
Scientific, Inc., Waltham, MA, USA).
RT-qPCR
RNA was reverse transcribed into cDNA using Takara
PrimeScript RT master mix (RR036A; Takara Biotechnology Co., Ltd.,
Dalian China). RT-qPCR was performed using an ABI StepOne Plus
Real-Time PCR system (Thermo Fisher Scientific, Inc.) and SYBR
Premix Ex Taq II master mix (Takara Biotechnology Co., Ltd.)
according to the manufacturer's protocol. The reaction system (10
µl) consisted of: cDNA (1 µl), forward primers (10 µM; 0.2 µl),
reverse primers (10 µM; 0.2 µl), ROX reference dye (0.2 µl),
RNase-free water (3.4 µl) and SYBR-Green mixture (5 µl). The
thermocycling conditions were as follows: Initial denaturation,
95°C for 30 sec, followed by 40 cycles of 95°C for 5 sec and 60°C
for 30 sec. GAPDH was used as a housekeeping gene. The relative
expression of genes was calculated using the 2−∆∆Ct
method (23). The primer sequences
used for RT-qPCR were as follows: GAPDH forward,
5′-TGTGTCCGTCGTGGATCTGA-3′ and reverse,
5′-TTGCTGTTGAAGTCGCAGGAG-3′; interleukin (IL)-6 forward,
5′-AGACAAAGCCAGAGTCCTTCA-3′ and reverse,
5′-GTGACTCCAGCTTATCTCTTGGT-3′; transcription factor AP-1 (c-Jun)
forward, 5′-CCTTCTACGACGATGCCCTCAA-3′ and reverse,
5′-GGGGTCGGTGTAGTGGTGATGT-3′; urikinase-type plasminogen activator
(Plau) forward, 5′-AGTGTGGCCAGAAGGCTCTA-3′ and reverse,
5′-GCTGCTCCACCTCAAACTTC-3′.
Statistical analysis
Data are presented as the mean ± standard deviation.
One-way analysis of variance followed by the Bonferroni post hoc
test was used to determine the statistical significance, with SPSS
software, version 19.0 (IBM Corp., Armonk, NY, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Data preprocessing
One disqualified gene chip sample (GSM375686) was
removed following quality control of the microarray expression
profile. A total of eight samples based on the GPL1355 platform
were included for research; this consisted of 2 samples from the
Sham group, 3 samples from the Contra group and 3 samples from the
Ipsi group. As a result, 13,686 genes were obtained from GSE15041.
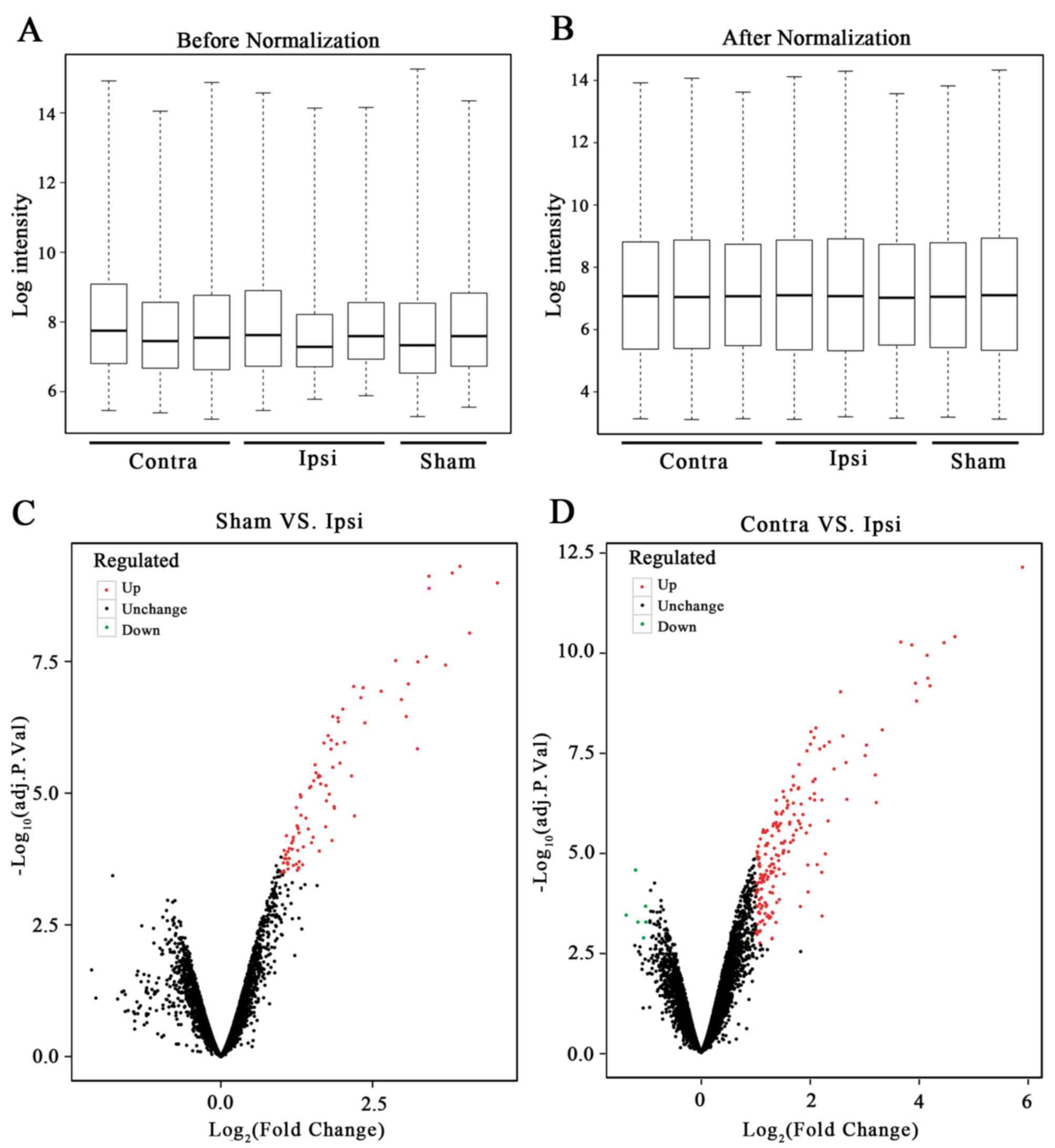
The box plot of the log expression values for all genes in each
sample before and after normalization were drawn (Fig. 1A and B). The median values of each
sample were extremely similar, indicating that the data should be
further analyzed.
Identification of the DEGs
The limma package in R was used to perform the DEG
analysis. No DEGs were identified between the Sham and Contra
group, 90 DEGs were identified between the Sham and Ipsi group and
191 DEGs were identified between the Contra and Ipsi group. The
volcanic plot of the Sham vs. Ipsi groups (Fig. 1C) and the Contra vs. Ipsi groups
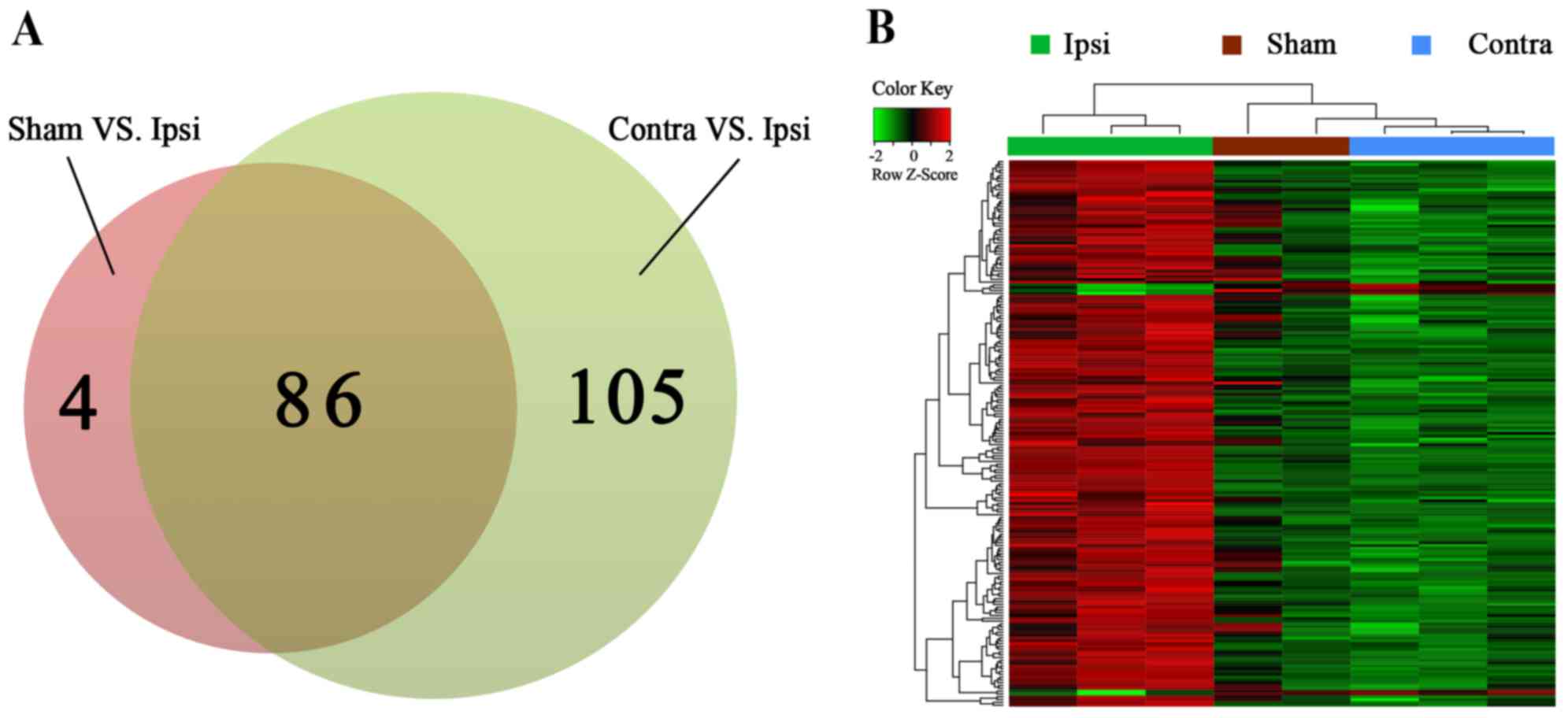
were drawn (Fig. 1D). In total, 86
common DEGs were selected by Venn diagram (Fig. 2A). A heat map of the identified
DEGs of the Sham, Contra and Ipsi groups was subsequently
constructed to perform cluster analysis (Fig. 2B). The DEG clustering was
predominantly separated into two clusters; one cluster was the Ipsi
group and the other was the Sham and Contra group.
Function and pathway enrichment
analysis
A total of 86 common DEGs were significantly
enriched in 77 GO terms (Table I)
and 17 KEGG pathways (Table II).
Notably, function enrichment analysis revealed that in cellular
component terms, DEGs were significantly enriched in ‘extracellular
space’; in biological processes, DEGs were enriched in response to
lipopolysaccharides and cytokines and ‘wound healing’; and in
molecular function terms, DEGs were enriched in neuropeptide
hormone activity. In KEGG pathway enrichment analysis, DEGs were
significantly enriched in tuberculosis, rheumatoid arthritis, human
T-cell lymphotropic virus type 1 (HTLV-1) and the mitogen-activated
protein kinase (MAPK) signaling pathway.
 | Table I.Functional enrichment analysis for
the common differentially expressed genes. |
Table I.
Functional enrichment analysis for
the common differentially expressed genes.
| GO category | GO number | GO term | Number of
differentially expressed genes | P-value |
|---|
| CC | GO:0005615 | Extracellular
space | 23 |
1.77×10‒8 |
| BP | GO:0071222 | Cellular response
to lipopolysaccharide | 8 |
1.02×10‒5 |
| MF | GO:0005184 | Neuropeptide
hormone activity | 5 |
1.15×10‒5 |
| BP | GO:0034097 | Response to
cytokine | 6 |
8.39×10‒5 |
| BP | GO:0043066 | Negative regulation
of apoptotic process | 11 |
1.06×10‒4 |
| BP | GO:0007565 | Female
pregnancy | 6 |
1.44×10‒4 |
| BP | GO:0042060 | Wound healing | 6 |
2.61×10‒4 |
| BP | GO:0032496 | Response to
lipopolysaccharide | 8 |
2.67×10‒4 |
| BP | GO:0032355 | Response to
estradiol | 7 |
2.92×10‒4 |
| BP | GO:0006954 | Inflammatory
response | 8 |
3.81×10‒4 |
| Table II.KEGG pathway enrichment analysis for
the common differentially expressed genes. |
Table II.
KEGG pathway enrichment analysis for
the common differentially expressed genes.
| KEGG number | KEGG term | Number of
differentially expressed genes | P-value |
|---|
| rno05152 | Tuberculosis | 8 |
2.79×10‒5 |
| rno05323 | Rheumatoid
arthritis | 5 |
1.01×10‒3 |
| rno04145 | Phagosome | 6 |
2.77×10‒3 |
| rno05166 | Human T-cell
lymphotropic virus type 1 infection | 7 |
2.99×10‒3 |
| rno05321 | Inflammatory bowel
disease | 4 |
4.10×10‒3 |
| rno05202 | Transcriptional
misregulation in cancer | 5 |
8.45×10‒3 |
| rno04010 | Mitogen-activated
protein kinase signaling pathway | 6 |
8.98×10‒3 |
| rno04514 | Cell adhesion
molecules | 5 |
1.04×10‒2 |
| rno04612 | Antigen processing
and presentation | 4 |
1.24×10‒2 |
| rno05146 | Amoebiasis | 4 |
1.82×10‒2 |
Protein-protein interaction network
analysis of DEGs
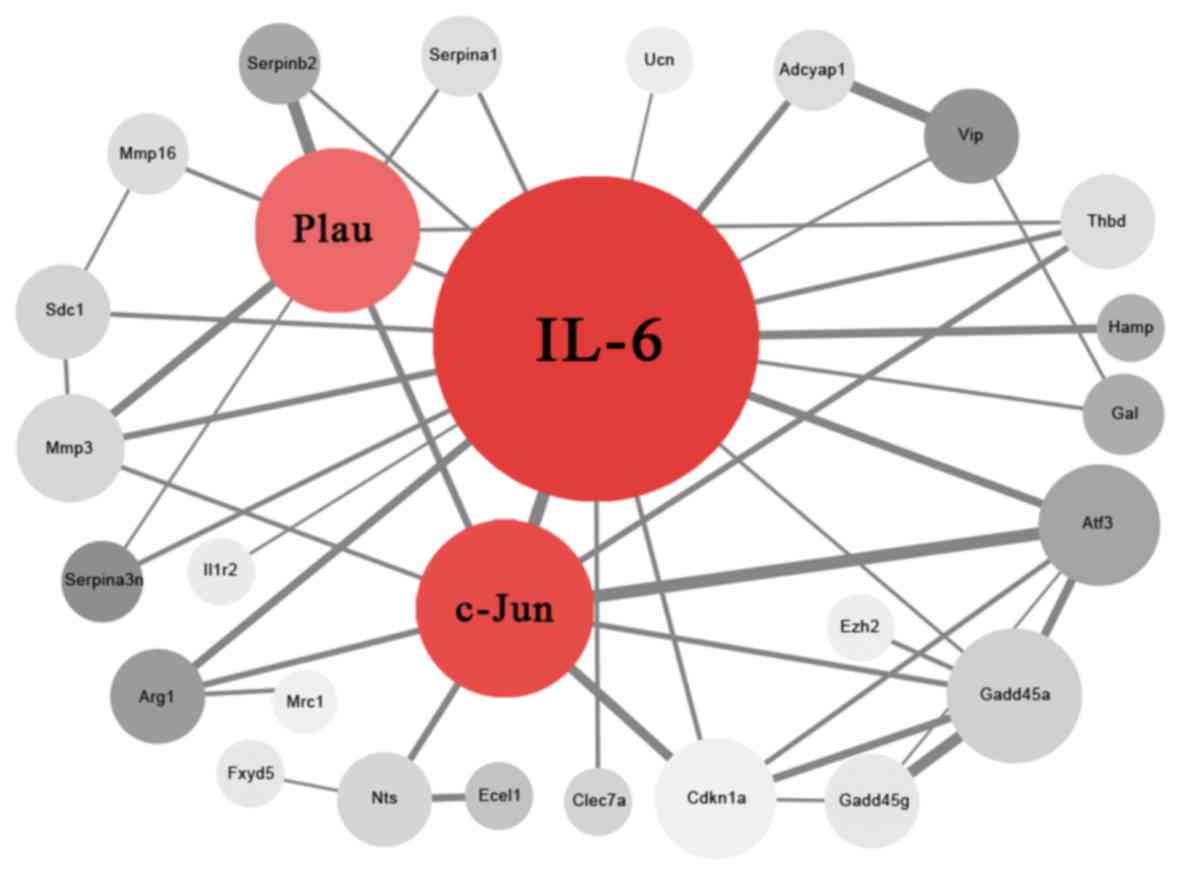
In the present study, 27 nodes (proteins) and 47 PPI
edges (interactions) were obtained following deletion of the
disconnected nodes in the network (Fig. 3). IL-6, c-Jun and Plau were
considered as hub proteins and key genes (Table III) that may have an important
role in the development of neuropathic pain.
| Table III.Hub proteins in the protein-protein
interaction network. |
Table III.
Hub proteins in the protein-protein
interaction network.
| Proteins | Degree |
|---|
| IL-6 | 20 |
| Jun | 9 |
| Plau | 8 |
| Gadd45α | 6 |
| Atf3 | 5 |
| Cdkn1a | 5 |
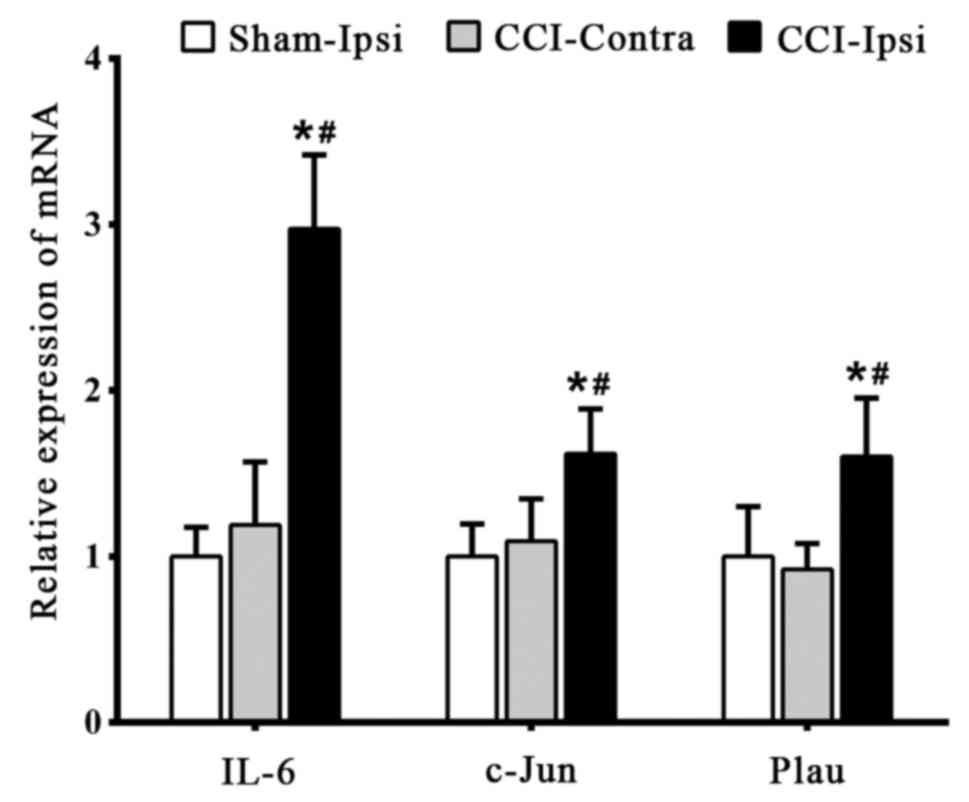
RT-qPCR validation
RT-qPCR was used to detect the expression of the
identified key genes to confirm the reliability of bioinformatics
analysis. Compared with the CCI-Ipsi group, the expression levels
of IL-6, c-Jun and Plau were significantly decreased (P<0.05) in
the Sham-Ipsi and CCI-Contra group 7 days post-surgery; no
significant difference (P>0.05) was detected between the
Sham-Ipsi group and CCI-Contra group (Fig. 4).
Discussion
At present, the mechanisms underlying neuropathic
pain remain unclear. The development of chronic pain is thought to
be due to peripheral and/or central nerve injury. Several studies
have reported that nerve injury induces alteration in the
peripheral immune system and glial cells (24–26).
Additionally, it has been demonstrated that the activation of
microglia is associated with the early stages of neuropathic pain
in the spinal dorsal horn where the nerve is injured (27–29).
Therefore, identifying key genes involved may be helpful in further
understanding these mechanisms underlying the development of
neuropathic pain.
Vega-Avelaira et al (10) used the microarray expression
profile GSE15041 to identify DEGs associated with neuropathic pain.
However, the expression profile required re-analysis, as the gene
chip quality was not verified. The present study obtained different
results following correct quality control. Microarray quality
control has a significant impact on the results of the analysis and
unverified microarrays may lead to incorrect results. In the
present study, mRNA expression was analyzed in a mouse model of
neuropathic pain 7 days post-surgery to confirm the bioinformatics
analysis results. RT-qPCR was performed following 7 days as mice
have the lowest paw withdrawal threshold and this is accompanied by
significant changes in molecular biology (30).
A total of 86 common DEGs among the Sham, Contra and
Ipsi groups were identified following quality control of the
microarray and removal of the disqualified sample. The pivotal
roles of IL-6, Jun and Plau in neuropathic pain were subsequently
identified.
IL-6 is an inflammatory cytokine with a wide range
of biological effects. It has been reported that unilateral sciatic
nerve injury increases the expression of IL-6 mRNA (31), and mechanical allodynia induced by
spinal nerve lesion is attenuated and delayed in IL-6 knockout mice
(32). Furthermore, intrathecal
injection of anti-IL-6 antibody reduces mechanical allodynia
induced by spinal nerve transection (33). These data indicate the involvement
of IL-6 in the initiation and development of neuropathic pain.
Immediate early genes (IEGs) may be transiently and rapidly
activated in response to a wide variety of cellular stimuli.
Well-characterized IEGs include c-Jun, and the proto-oncogenes
c-Fos and c-Myc. Double labeling with the tracers fast blue and
horseradish peroxidase-labeled gold underwent retrograde transport
from the site of injury and revealed that c-Jun protein expression
was confined to injured neurons (34). Activated c-Jun in the dorsal root
ganglion (DRG) contributes to the pathogenesis of neuropathic pain
induced by CCI (35), and
neuropathic pain is alleviated by inhibiting the mitogen-activated
protein kinase 8 (JNK)/c-Jun signal pathway (36,37).
Furthermore, Plau mRNA is abundantly expressed by many neurons in
the peripheral and central nervous system (38). Plau expression is upregulated in
rat DRGs following nerve injury (39). The aforementioned data suggests the
involvement of IL-6, c-Jun and Plau in neuropathic pain.
Common DEGs were revealed to be enriched in the
injury and inflammation-associated biological processes through GO
enrichment analysis. Pathway analysis revealed that DEGs were
typically enriched in tuberculosis, rheumatoid arthritis, HTLV-1
and MAPK signaling pathways. Spinal tuberculosis may induce varying
degrees of damage to the spinal cord nerve roots (40), resulting in neuropathic pain. A
clinical study revealed that a third of patients with rheumatoid
arthritis have symptoms of neuropathic pain (41), and a review reported that
approximately 53.1% of HTLV-1 carriers have symptoms of neuropathic
pain (42). The conventional MAPKs
include the p38 isoforms, extracellular signal-regulated kinases
1/2 (ERK1/2) and JNKs (43). MAPKs
regulate numerous physiological activities, including inflammation,
apoptosis, cancer, tumor cell invasion and metastasis. Active MAPK
signaling also has an important role in neuropathic pain. It has
been reported that IL-6 is involved in the maintenance of
neuropathic pain by activating ERK; and the JNK/c-Jun pathway also
participates in the maintenance of neuropathic pain (43). It has been demonstrated that p38
MAPK activation in the ipsilateral spinal dorsal horn is
significantly increased following CCI (44). Additionally, MAPK activity is
significantly decreased following an intrathecal injection of
anti-IL-6 antibody (45). These
results indicate that IL-6 and c-Jun have an important role in
neuropathic pain that is mediated through the MAPK signaling
pathway. This is consistent with the bioinformatics findings of the
present study and reflect the reliability of the bioinformatics
analysis performed.
In the present study, the results of the
bioinformatics analysis revealed that there was a significant
difference between the Sham and Ipsi group (P<0.05), however no
difference was observed between the Sham and Contra group
(P>0.05). RT-qPCR analysis was consistent with this. The sciatic
nerve was only injured in the CCI-Ipsi group, not the CCI-Contra or
Sham-Ipsi group, and the nerve was only exposed in the Sham-Ipsi
group. Sciatic nerve injury induces the inflammatory response in
the ipsilateral spinal cord, whereas the contralateral spinal cord
was not, compared with the sham group, as previously reported
(46). Nerve injury induces a set
of progressive alterations in ipsilateral spinal cord microglia
with increased secretion of cytokines. This does not occur in the
contralateral spinal cord following nerve injury, or the uninjured
ipsilateral spinal cord. Therefore, significant changes in
neurophysiology in spinal cord of the Sham and Contra group may not
be induced. DEG analysis revealed that there were numerous DEGs
detected in the Ipsi group compared with the Contra and Sham
groups, and not between the Contra and Sham groups. This may be due
to only the Ipsi group exhibiting nerve injury, whereas the other
two groups did not. However, DEGs may be detected between the
Contra and Sham group only in the local tissue of the surgical
incision, rather than the spinal cord. Thus, many DEGs were
identified in the Ipsi group compared with Sham and Contra group by
bioinformatics analysis.
In summary, 86 DEGs were identified and IL-6, c-Jun,
Plau were identified as key genes in neuropathic pain through
pathway and PPI network analysis, which may aid in further
understanding of the underlying pain mechanisms. Following the
bioinformatics analysis, the expression of key genes was confirmed
with RT-qPCR in a mouse model of neuropathic pain. However, the
current study has certain limitations. Although the results
revealed that IL-6, c-Jun and Plau have an important role in
neuropathic pain, further studies are required to confirm the
results at the molecular level.
Acknowledgements
The authors of the present study would like to thank
their colleagues for their support and help.
Funding
The present study supported by the National Natural
Science Foundation of China (grant nos. 81471129, 81371207,
81171047 and 81171048), the Natural Science Foundation of Jiangsu
Province of China (grant no. BK20170654), the Department of Health
of Jiangsu Province of China (grant nos. XK201140 and RC2011006)
and the Fundamental Research Funds for the Central Universities
(grant nos. 0214-14380338 and 0214-14380271).
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Author's contributions
Study Design: HL, TX, ZM and XG; data collection:
HL; statistical analysis: HL and TX; animal experiment: FX and HL;
manuscript preparation: HL, TX and XG; funds collection: TX, ZM and
XG.
Ethics approval and consent to
participate
All animal experiments were approved by the
Experimental Animals Welfare and Ethical Inspection of Nanjing Drum
Tower Hospital (Nanjing, China).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Woolf CJ and Mannion RJ: Neuropathic pain:
Aetiology, symptoms, mechanisms, and management. Lancet.
353:1959–1964. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
van Hecke O, Austin SK, Khan RA, Smith BH
and Torrance N: Neuropathic pain in the general population: A
systematic review of epidemiological studies. Pain. 155:654–662.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bouhassira D, Lanteri-Minet M, Attal N,
Laurent B and Touboul C: Prevalence of chronic pain with
neuropathic characteristics in the general population. Pain.
136:380–387. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Attal N, Lanteri-Minet M, Laurent B,
Fermanian J and Bouhassira D: The specific disease burden of
neuropathic pain: Results of a French nationwide survey. Pain.
152:2836–2843. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Finnerup NB, Haroutounian S, Kamerman P,
Baron R, Bennett DL, Bouhassira D, Cruccu G, Freeman R, Hansson P,
Nurmikko T, et al: Neuropathic pain: An updated grading system for
research and clinical practice. Pain. 157:1599–1606. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Attal N and Bouhassira D: Pharmacotherapy
of neuropathic pain: Which drugs, which treatment algorithms? Pain.
156 Suppl 1:S104–S114. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gewandter JS, Dworkin RH, Turk DC, Farrar
JT, Fillingim RB, Gilron I, Markman JD, Oaklander AL, Polydefkis
MJ, Raja SN, et al: Research design considerations for chronic pain
prevention clinical trials: IMMPACT recommendations. Pain.
156:1184–1197. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Finnerup NB, Sindrup SH and Jensen TS:
Recent advances in pharmacological treatment of neuropathic pain.
F1000 Med Rep. 2:522010.PubMed/NCBI
|
|
9
|
Luscombe NM, Greenbaum D and Gerstein M:
What is bioinformatics? A proposed definition and overview of the
field. Methods Inf Med. 40:346–358. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vega-Avelaira D, Géranton SM and
Fitzgerald M: Differential regulation of immune responses and
macrophage/neuron interactions in the dorsal root ganglion in young
and adult rats following nerve injury. Mol Pain. 5:702009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Carlson M: Rat2302.db: Affymetrix Rat
Genome 230 2.0 Array annotation data (chip rat2302). R package.
version 3.2.3. 2016.
|
|
12
|
Gautier L, Cope L, Bolstad BM and Irizarry
RA: Affy-analysis of Affymetrix GeneChip data at the probe level.
Bioinformatics. 20:307–315. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wilson CL and Miller CJ: Simpleaffy: A
BioConductor package for Affymetrix quality control and data
analysis. Bioinformatics. 21:3683–3685. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Irizarry RA, Hobbs B, Collin F,
Beazer-Barclay YD, Antonellis KJ, Scherf U and Speed TP:
Exploration, normalization, and summaries of high density
oligonucleotide array probe level data. Biostatistics. 4:249–264.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gentleman R: Annotate: Annotation for
microarrays. R package. version 1.56.1. 2017.
|
|
16
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: Limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Smyth GK: Linear models and empirical
bayes methods for assessing differential expression in microarray
experiments. Stat Appl Genet Mol Biol. 3:Article32004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The Gene
Ontology Consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Szklarczyk D, Franceschini A, Kuhn M,
Simonovic M, Roth A, Minguez P, Doerks T, Stark M, Muller J, Bork
P, et al: The STRING database in 2011: Functional interaction
networks of proteins, globally integrated and scored. Nucleic Acids
Res. 39:D561–D568. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Smoot ME, Ono K, Ruscheinski J, Wang PL
and Ideker T: Cytoscape 2.8: New features for data integration and
network visualization. Bioinformatics. 27:431–432. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bennett GJ and Xie YK: A peripheral
mononeuropathy in rat that produces disorders of pain sensation
like those seen in man. Pain. 33:87–107. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Scholz J and Woolf CJ: The neuropathic
pain triad: Neurons, immune cells and glia. Nat Neurosci.
10:1361–1368. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Milligan ED and Watkins LR: Pathological
and protective roles of glia in chronic pain. Nat Rev Neurosci.
10:23–36. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Thacker MA, Clark AK, Marchand F and
McMahon SB: Pathophysiology of peripheral neuropathic pain: Immune
cells and molecules. Anesth Analg. 105:838–847. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Inoue K and Tsuda M: Microglia and
neuropathic pain. Glia. 57:1469–1479. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dominguez E, Rivat C, Pommier B, Mauborgne
A and Pohl M: JAK/STAT3 pathway is activated in spinal cord
microglia after peripheral nerve injury and contributes to
neuropathic pain development in rat. J Neurochem. 107:50–60. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tsuda M, Shigemoto-Mogami Y, Koizumi S,
Mizokoshi A, Kohsaka S, Salter MW and Inoue K: P2X4 receptors
induced in spinal microglia gate tactile allodynia after nerve
injury. Nature. 424:778–783. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lai CY, Hsieh MC, Ho YC, Lee AS, Wang HH,
Cheng JK, Chau YP and Peng HY: Growth Arrest and
DNA-damage-inducible protein 45β-mediated DNA demethylation of
voltage-dependent t-type calcium channel 3.2 subunit enhances
neuropathic allodynia after nerve injury in rats. Anesthesiology.
126:1077–1095. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Luo X, Tai WL, Sun L, Pan Z, Xia Z, Chung
SK and Cheung CW: Crosstalk between astrocytic CXCL12 and
microglial CXCR4 contributes to the development of neuropathic
pain. Mol Pain. 12:2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ramer MS, Murphy PG, Richardson PM and
Bisby MA: Spinal nerve lesion-induced mechanoallodynia and
adrenergic sprouting in sensory ganglia are attenuated in
interleukin-6 knockout mice. Pain. 78:115–121. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Arruda JL, Sweitzer S, Rutkowski MD and
DeLeo JA: Intrathecal anti-IL-6 antibody and IgG attenuates
peripheral nerve injury-induced mechanical allodynia in the rat:
Possible immune modulation in neuropathic pain. Brain Res.
879:216–225. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Leah JD, Herdegen T and Bravo R: Selective
expression of Jun proteins following axotomy and axonal transport
block in peripheral nerves in the rat: Evidence for a role in the
regeneration process. Brain Res. 566:198–207. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Son SJ, Lee KM, Jeon SM, Park ES, Park KM
and Cho HJ: Activation of transcription factor c-jun in dorsal root
ganglia induces VIP and NPY upregulation and contributes to the
pathogenesis of neuropathic pain. Exp Neurol. 204:467–472. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang C, Kong X, Zhu C, Liu C, Sun D, Xu Q,
Mao Z, Qin Q, Su H, Wang D, et al: Wu-tou decoction attenuates
neuropathic pain via suppressing spinal astrocytic IL-1R1/TRAF6/JNK
signaling. Oncotarget. 8:92864–92879. 2017.PubMed/NCBI
|
|
37
|
Jiang L, Pan CL, Wang CY, Liu BQ, Han Y,
Hu L, Liu L, Yang Y, Qu JW and Liu WT: Selective suppression of the
JNK-MMP2/9 signal pathway by tetramethylpyrazine attenuates
neuropathic pain in rats. J Neuroinflammation. 14:1742017.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sumi Y, Dent MA, Owen DE, Seeley PJ and
Morris RJ: The expression of tissue and urokinase-type plasminogen
activators in neural development suggests different modes of
proteolytic involvement in neuronal growth. Development.
116:625–637. 1992.PubMed/NCBI
|
|
39
|
Yamanaka H, Obata K, Fukuoka T, Dai Y,
Kobayashi K, Tokunaga A and Noguchi K: Tissue plasminogen activator
in primary afferents induces dorsal horn excitability and pain
response after peripheral nerve injury. Eur J Neurosci. 19:93–102.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wouda EMN, Stienstra Y, van der Werf TS,
Kerstjens H, de Lange WCM, Coppes M, Kuijlen J, Tepper M and
Akkerman OW: Neurological and functional recovery in tuberculosis
patients with spinal cord injury in the Netherlands.
NeuroRehabilitation. 40:439–445. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yesil H, Sungur U, Akdeniz S, Gurer G,
Yalcin B and Dundar U: Association between serum vitamin D levels
and neuropathic pain in rheumatoid arthritis patients: A
cross-sectional study. Int J Rheum Dis. 21:431–439. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
San-Martin DL, Santos DN and Baptista AF:
Pain Study Group: Pain prevalence, characteristics and associated
factors in human T-cell lymphotropic virus type 1 infected
patients: A systematic review of the literature. Braz J Infect Dis.
20:592–598. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ding CP, Guo YJ, Li HN, Wang JY and Zeng
XY: Red nucleus interleukin-6 participates in the maintenance of
neuropathic pain through JAK/STAT3 and ERK signaling pathways. Exp
Neurol. 300:212–221. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhao WX, Wang PF, Song HG and Sun N:
Diosgenin attenuates neuropathic pain in a rat model of chronic
constriction injury. Mol Med Rep. 16:1559–1564. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zaringhalam J, Tekieh E, Manaheji H and
Akhtari Z: Cellular events during arthritis-induced hyperalgesia
are mediated by interleukin-6 and p38 MAPK and their effects on the
expression of spinal mu-opioid receptors. Rheumatol Int.
33:2291–2299. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Banno T, Omura T, Masaki N, Arima H, Xu D,
Okamoto A, Costigan M, Latremoliere A, Matsuyama Y and Setou M:
Arachidonic acid containing phosphatidylcholine increases due to
microglial activation in ipsilateral spinal dorsal horn following
spared sciatic nerve injury. PLoS One. 12:e01775952017. View Article : Google Scholar : PubMed/NCBI
|