Introduction
Skin defects are often caused by acute and chronic
disease (1). It is difficult to
achieve satisfactory treatment for patients with skin defects that
encompass a large area (2). With
the development of skin tissue engineering, a novel method for
wound healing can be provided (3).
The key to tissue engineering is the choice of seed cells (4). Hair follicle stem cells (HFSCs),
residing in the bulge area, are critical for epidermal regeneration
in response to wounding (5). HFSCs
have the unique capacity to differentiate into keratinocyte cells,
hair follicles, sebaceous glands, endothelial cells and even neural
lineage cells (6–9). However, the differentiation potential
of HFSCs is poor in vitro and in vivo. Small
molecules can be used to promote HFSC activation, which will be
useful for regenerative medicine (10). Resina draconis, a
traditional Chinese herbal medicine, has a long history of clinical
efficacy against wounds, ulcers and other hard-to-heal injuries
(11). Our previous study
indicated that loureirin A, the main active flavonoid ingredient of
Resina draconis, can promote HFSC differentiation by
activating the protein Wnt-9b (Wnt)/β-catenin signaling pathway and
inducing HFSC-seeded tissue-engineered skin to repair skin wounds
(12).
Animal genomes encode hundreds of microRNA
(miRNAs/miRs) genes and recent studies provide new insights into
these short non-coding RNAs, by demonstrating their key roles in
many biological processes, which include cell survival,
proliferation, and differentiation (13,14).
However, aberrant miRNA expression can lead to the development of
disease (15). miRNAs are also
central regulators in epidermal stem cell maintenance and wound
healing (16). miR-24, miR-125b
and miR-214 have unique functions in the maintenance of the hair
follicle in the telogen phase, by inhibiting the transcriptional
program that drives HFSC proliferation, differentiation and hair
shaft assembly (17–19). miR-22 can repress homeobox proteins
DLX-3 and DLX-4 expression to promote keratinocyte differentiation
and hair shaft formation (20).
However, it has not been determined if and how miRNAs modulate
loureirin A-induced HFSC differentiation.
Each miRNA can bind to numerous different mRNAs and
assemble with Argonaute proteins, to form miRNA-induced silencing
complexes that direct post-transcriptional silencing of
complementary mRNA targets (21).
miR-339-5p is a cancer-relevant miRNA that participates in various
cell processes, including cell proliferation, migration, and
invasion (22,23). Additionally, miR-339-5p regulates
neural crest stem cell maintenance and differentiation (24). DLX5 is a transcription factor that
is associated with epithelial cell differentiation (25). DLX5 positively regulates the
Wnt/β-catenin signaling pathway (26), as well as signaling pathways that
modulate stem cells pluripotency (27), based on Gene Ontology enrichment
and Kyoto Encyclopedia of Genes and Genomes pathway analysis in the
Database for Annotation, Visualization and Integrated Discovery
(https://david.ncifcrf.gov/). It has also
been reported that DLX5 is associated with the maintenance of
pluripotent stem cells during developmental processes (28). However, it has not been well
documented whether miR-339-5p targets DLX5 to regulate the
differentiation of HFSCs.
In the present study, a miRNA array of HFSCs treated
with loureirin A was performed. Among the differentially expressed
miRNAs identified, miR-339-5p was further studied as an underlying
suppressor of HFSC differentiation. Notably, DLX5 was identified as
a target gene of miR-339-5p in HFSCs. The present study aimed to
clarify if miR-339-5p targeted DLX5 to inhibit HFSCs
differentiation via the Wnt/β-catenin signaling pathway. This may
provide a potential molecular therapeutic target for the treatment
of skin defects.
Materials and methods
Cell culture and identification
Male Sprague-Dawley rats (3 days old; 20±2 g; n=180)
and mothers (3 months old; 300±20 g; n=5) were provided by the
Laboratory Animal Centre of Guangzhou University of Chinese
Medicine (permit no. SCXK 2013-0020; Guangzhou, China). Rats were
maintained in an environment with controlled humidity (50±5%),
temperature (22±1°C), 12 h light/dark cycle and 0.03%
CO2 conditions, with ad libitum access to milk
from their mothers. The present study was approved by the Ethics
Committee of Guangzhou University of Chinese Medicine (Guangzhou,
China). Male SD rats (20±2 g) were anesthetized with an
intraperitoneal injection of sodium pentobarbital at a dose of 50
mg/kg. The rats were sacrificed by decollation and subsequently
sterilized using 75% ethyl alcohol. HFSCs were isolated and
cultured as previously reported, with some modifications (29). Briefly, hair follicles were
separated from the vibrissae and subsequently digested with 1 g/l
collagenase type I (Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) for 30 min at 37°C, followed by 2.5 g/l trypsin (Gibco;
Thermo Fisher Scientific, Inc.) for 30 min at 37°C. The cells were
cultivated with keratinocyte serum-free medium (K-SFM; Gibco;
Thermo Fisher Scientific, Inc.) and incubated at 37°C in 5%
CO2. Primary rat HFSCs were passaged 6–7 days later.
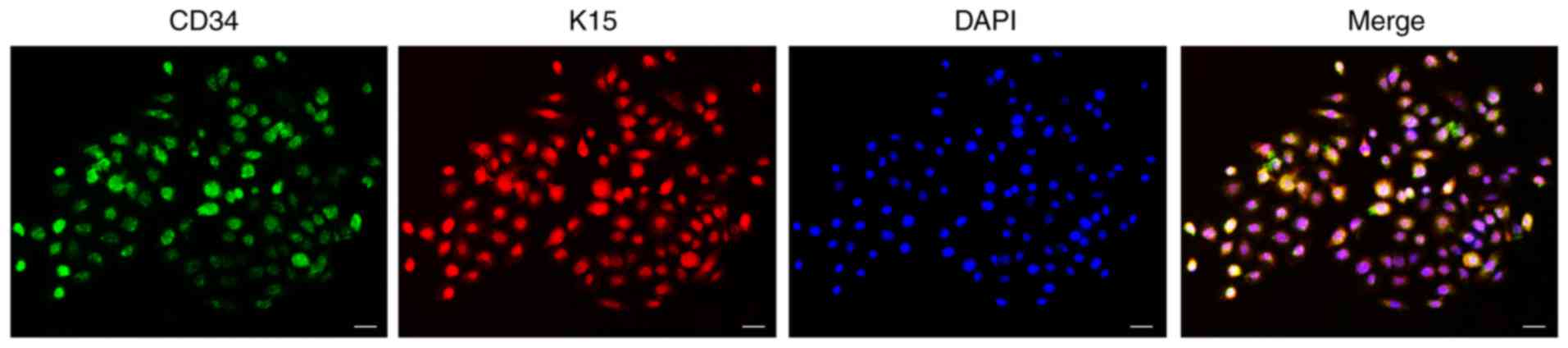
HFSCs were seeded at a density of 1×104
cells/well. When cells reached 80% confluence, hematopoietic
progenitor cell antigen CD34 and keratin, type I cytoskeletal 15
(K15) were assayed by double immunofluorescence staining. Cell
samples were fixed in 4% paraformaldehyde for 30 min at room
temperature and subsequently rinsed with PBS. Cells and nuclear
membranes were permeabilized with 0.3% Triton X-100 (Beyotime
Institute of Biotechnology, Shanghai, China) for 15 min at room
temperature. Following this, cells were overlaid with 5% bovine
serum albumin (Beyotime Institute of Biotechnology) for 30 min at
room temperature, rinsed with PBS and incubated with primary
antibodies against CD34 (1:100; cat no. bsm-10820M; BIOSS, Beijing,
China) and K15 (1:100; cat no. bs-4762R; BIOSS) at 4°C overnight.
Cells were washed with PBS and subsequently incubated with Alexa
Fluor 488-labeled goat anti-mouse IgG secondary antibody (1:200;
cat. no. A0482; Beyotime Institute of Biotechnology) and Alexa
Fluor 555-labeled donkey anti-rabbit IgG secondary antibody (1:200;
cat. no. A0453; Beyotime Institute of Biotechnology) for 1 h at
room temperature. Finally, cells were washed three times with PBS
and counterstained with DAPI (1:1,000; cat. no. C1002; Beyotime
Institute of Biotechnology) for 5 min at room temperature. Slides
were observed with an Olympus IX71 fluorescence microscope (Olympus
Corporation, Tokyo, Japan) at magnification, ×100.
Treatment of HFSCs with loureirin
A
Loureirin A was purchased from the National
Institute for the Control of Pharmaceutical and Biological Products
(cat. no. 200402; Beijing, China). HFSCs were seeded at a density
of 5×104 cells/well in 6-well culture plates. When 80%
confluence was reached, cells were treated with K-SFM containing 20
µg/ml loureirin A for 24 h at 37°C in 5% CO2. HFSCs
cultured with K-SFM only were used as a control. After 24 h, the
cells were extracted for RNA extraction.
RNA extraction and microarray-based
miRNA profiling analysis
Total RNA was extracted from HFSCs with TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.), according to
the manufacturer's instructions. RNA quantity and purity was
assessed using a K5500 micro-spectrophotometer.
A260/A280>1.5 and
A260/A230>1 indicated acceptable RNA
purity and a RNA integrity number value of >7 in the Agilent
2200 RNA TapeStation system (Agilent Technologies, Inc., Santa
Clara, CA, USA) indicated acceptable RNA integrity. Genomic DNA
contamination was evaluated by gel electrophoresis, as described
previously (30). miRNA profiling
was performed at Guangzhou RiboBio Co., Ltd. (Guangzhou, China).
For advanced data analysis, all biological replicates were pooled
and calculated to identify differentially expressed miRNAs based on
the threshold (P<0.05). Data were analyzed by Multiexperiment
Viewer 4.9.0 (http://mev.tm4.org/).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RNA was extracted from HFSCs using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) as described above.
Reverse transcription and qPCR was performed using the Prime
Script™ RT reagent kit and SYBR Premix EX Taq II kit
(both Takara Biotechnology, Co., Ltd., Dalian, China), according to
the manufacturer's instructions. The thermocycling conditions used
in the CFX96™ Real-Time PCR Detection System (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) were as follows: Initial
denaturation at 95°C for 30 sec, followed by 40 cycles of 95°C for
5 sec, annealing at 60°C for 30 sec and final extension at 60°C for
30 sec. miRNA expression was quantified using the 2−ΔΔCq
method (31). The primers for
miRNA were synthesized by Sangon Biotech Co., Ltd. (Shanghai,
China) and the sequences were as follows: miR-3589 forward,
5′-CGGAGGAAACCAGCAAG-3′ and reverse, 5′-GTGCAGGGTCCGAGGT-3′;
miR-347 forward, 5′-TCTCCTGTCCCTCTGGGT-3′ and reverse,
5′-GTGCAGGGTCCGAGGT-3′; miR-203a-3p forward,
5′-CGGCGTGAAATGTTTAGG-3′ and reverse, 5′-GTGCAGGGTCCGAGGT-3′;
miR-339-5p forward, 5′-GTGTCCCTGTCCTCCAGG-3′ and reverse,
5′-GTGCAGGGTCCGAGGT-3′; miR-3099 forward, 5′-GGCGTAGGCTAGAAAGAGG-3′
and reverse, 5′-GTGCAGGGTCCGAGGT-3′; miR-6216 forward,
5′-AGCGGATACACAGAGGCA-3′ and reverse, 5′-GTGCAGGGTCCGAGGT-3′;
miR-3566 forward, 5′-TACCGGCTGCCTAACAA-3′ and reverse,
5′-GTGCAGGGTCCGAGGT-3′; U6 forward, 5′-CTCGCTTCGGCAGCACATAT-3 and
reverse, 5′-TTGCGTGTCATCCTTGCG-3′.
Electrotransfection of HFSCs with
miR-339-5p mimic, miR-339-5p inhibitor or small interfering (si)RNA
targeting DLX5 (siDLX5)
HFSCs were collected with 0.25% trypsin digestion
when cell confluence reached 80%. miRNA sequences were purchased
from Guangzhou RiboBio Co., Ltd.. Cells were electrotransfected
with 0.2 µg of miR-339-5p mimic (5′-UCCCUGUCCUCCAGGAGCUCACG-3′),
miR-339-5p mimic negative control (NC; sequence unavailable),
miR-339-5p inhibitor (5′-CGUGAGCUCCUGGAGGACAGGGA-3′), miR-339-5p
inhibitor NC (sequence unavailable) or siDLX5 using a NEPA21
electroporator (Nepa Gene Co., Ltd., Ichikawa, Japan) in
Opti-minimum essential media (Invitrogen; Thermo Fisher Scientific,
Inc.), according to the manufacturer's electrotransfection
protocol. The sequence of the siDLX5 was:
5′-GCAGCCAGCUCAAUCAAUUTT-3′ and the negative control sequence was:
5′-UUCUCCGAACGUGUCACGUTT-3′. HFSCs were seed at density of
5×105 cells/well in 6-well culture plates. After 48 h,
cells were harvested for western blot and immunofluorescence
analyses.
Western blot analysis
Cells were washed twice with PBS and incubated in
radioimmunoprecipitation assay lysis buffer (Thermo Fisher
Scientific, Inc.) containing phenylmethanesulfonyl fluoride
(Beyotime Institute of Biotechnology) on ice for 30 min. The
supernatant for each protein group was obtained by centrifugation
at 12,000 × g at 4°C for 20 min and protein was quantified with the
bicinchoninic acid protein assay. HFSC protein extracts (20
µg/lane) were separated by 10% SDS-PAGE and electrophoretically
transferred to microporous polyvinylidene difluoride membranes,
followed by blocking with 5% non-fat dry milk in Tris buffered
saline 0.01% Tween-20 at room temperature for 3 h. The blot was
probed with DLX5 (1:10,000; cat. no. ab109737; Abcam, Cambridge,
UK), keratin, type I cytoskeletal 10 (K10; 1:10,000; cat. no.
ab76318; Abcam), involucrin (1:100; cat. no. OM174239; Omnimabs,
Alhambra, CA, USA), protein Wnt-3a (Wnt3a; 1:10,000; cat. no.
ab172612), β-actin (1:50,000; cat. no. ab49900) and GAPDH
(1:10,000; cat. no. ab181602) primary antibodies at 4°C overnight.
Following this, the blot was probed by a horseradish
peroxidase-labeled secondary antibody (1:20,000; cat. no. ab6721;
all Abcam), at room temperature for 1 h and washed with
Tris-buffered saline with 0.1% Tween-20. Bands were visualized
using an enhanced chemiluminescence kit (Beyotime Institute of
Biotechnology) according to the manufacturer's protocol and
analyzed by ImageJ software version 1.4 (National Institutes of
Health, Bethesda, MD, USA). The relative value of the target
protein was calculated by comparison with the corresponding
internal reference gene for each group.
Co-transfection of HFSCs with
pLUC-DLX5 and miR-339-5p mimic or inhibitor and luciferase reporter
assay
HFSCs were collected and electrotransfected with 0.2
µg pLUC-DLX5 plasmid (Shenzen Huaan Pingkang Biological Technology
Co., Ltd., Shenzhen, China), and subsequently electrotransfected
with the miR-339-5p mimic, miR-339-5p inhibitor, or the
corresponding negative controls (Guangzhou RiboBio Co., Ltd.), as
described above. In the pLUC-DLX5 cloning vector, miRNA target
sites predicted by TargetScan software (version 7.1; http://www.targetscan.org/vert_71/) were inserted
after the Renilla luciferase region. Activity was normalized
to firefly luciferase activity. After 48 h, the luminescence of
each well was collected and quantified with the dual luciferase
reporter assay system (Promega Corporation, Madison, WI, USA).
Immunofluorescence analysis
Cells were stained on coverslips. Cell samples were
fixed in 4% paraformaldehyde for 30 min at room temperature and
permeabilized with 0.3% Triton X-100 for 15 min at room
temperature. Cells were subsequently stained with the anti-DLX5
antibody (1:1,000; cat. no. ab109737; Abcam) at 4°C overnight,
followed by incubation with Alexa Fluor 555-labeled donkey
anti-rabbit IgG secondary antibody (1:200; cat. no. A0453; Beyotime
Institute of Biotechnology) for 1 h at room temperature. Coverslips
were counterstained with DAPI (1:1,000) for 5 min at room
temperature for nuclei visualization. Microscopic analysis was
performed with a ZOE™ Fluorescent Cell Imager (Bio-Rad
Laboratories, Inc.). Fluorescence intensity was measured in five
viewing areas for 200–300 cells per coverslip and analyzed using
ImageJ software version 1.4.
Statistical analysis
Data are presented as the mean ± standard deviation
of three independent experiments. All statistical analyses were
performed using GraphPad Prism 5.0 software (GraphPad Software,
Inc., La Jolla, CA, USA). P-values were calculated using the
unpaired Student's t-test, or one-way analysis of variance followed
by the Bonferroni procedure. P<0.05 was considered to indicate a
statistically significant difference.
Results
Morphology and identification of
HFSCs
The cultured cells were cobblestone-like and
exhibited the typical morphology of stem cells. Cultured cells
proliferated well and were confluent after approximately 7 days.
Cells stained positive for both CD34 and K15 (Fig. 1).
miR-339-5p is a negative regulator of
loureirin A-induced HFSC differentiation
The present study aimed to determine if differential
miRNA expression existed in HFSCs following treatment with
loureirin A. Microarray-based miRNA profiling analysis is an
effective method for the prediction of the mechanisms underlying
the effects of Chinese medicine monomers, including loureirin A
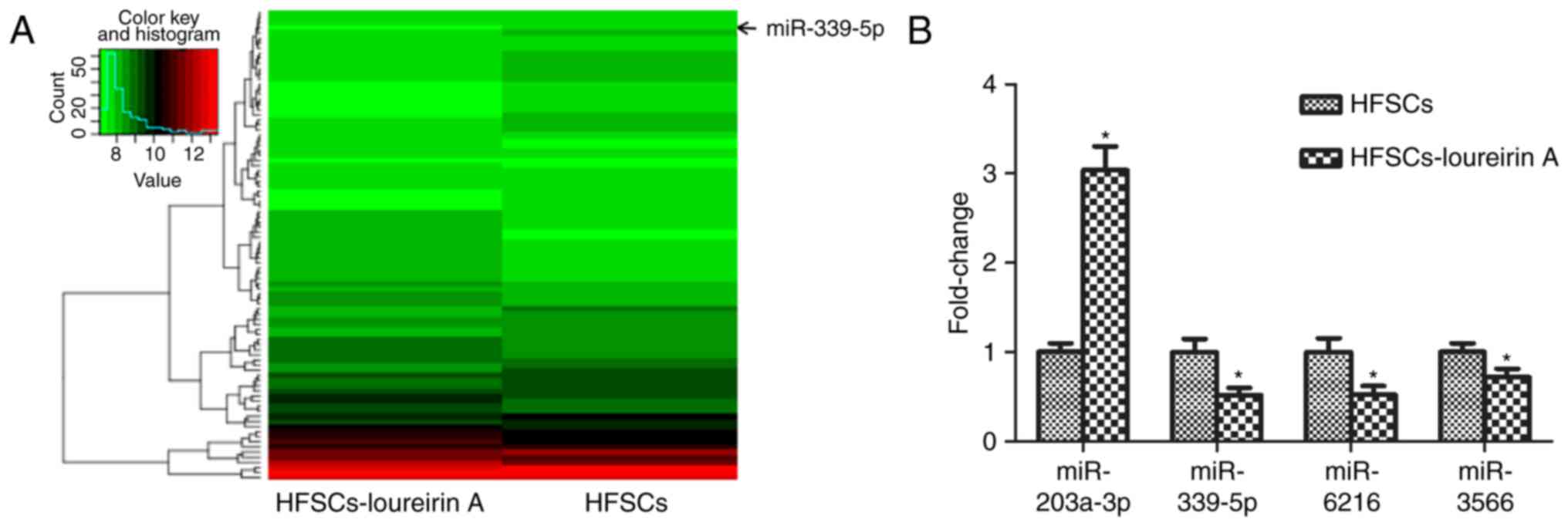
(32). A total of 92
differentially expressed miRNAs were identified between the control
and loureirin A groups. Compared with the control, 41 miRNAs were
upregulated and 51 miRNAs were downregulated in the loureirin A
group. Data were analyzed and a cluster diagram was created, with
P<0.05 taken as the filter condition of differential gene
expression (Fig. 2A). In
particular, miR-203a-3p, miR-339-5p, miR-6216 and miR-3566
exhibited prominent differences; these were also confirmed with
RT-qPCR (Fig. 2B). By contrast to
the control group, miR-339-5p, miR-6216 and miR-3566 were
down-regulated, while miR-203a-3p was upregulated. The function of
miR-339-5p in HFSCs is largely unknown, so further in vitro
analysis of miR-339-5p was conducted.
Downregulation of miR-339-5p promotes
HFSC differentiation
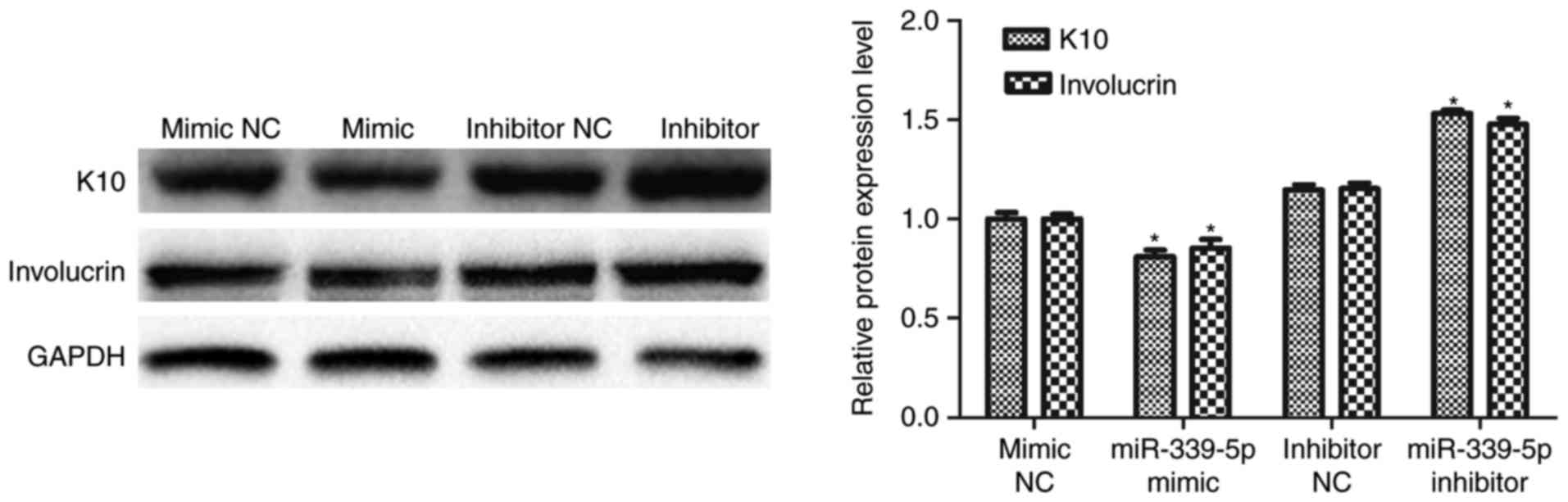
K10 and involucrin are markers of epidermal
differentiation. miR-339-5p was transfected into HFSCs and western
blot analysis was performed to determine if miR-339-5p was involved
in the regulation of K10 and involucrin expression. The result
revealed that K10 and involucrin expression decreased in the
miR-339-5p mimic group, compared with the mimic NC group and
increased in the miR-339-5p inhibitor group compared with the
inhibitor NC group (Fig. 3).
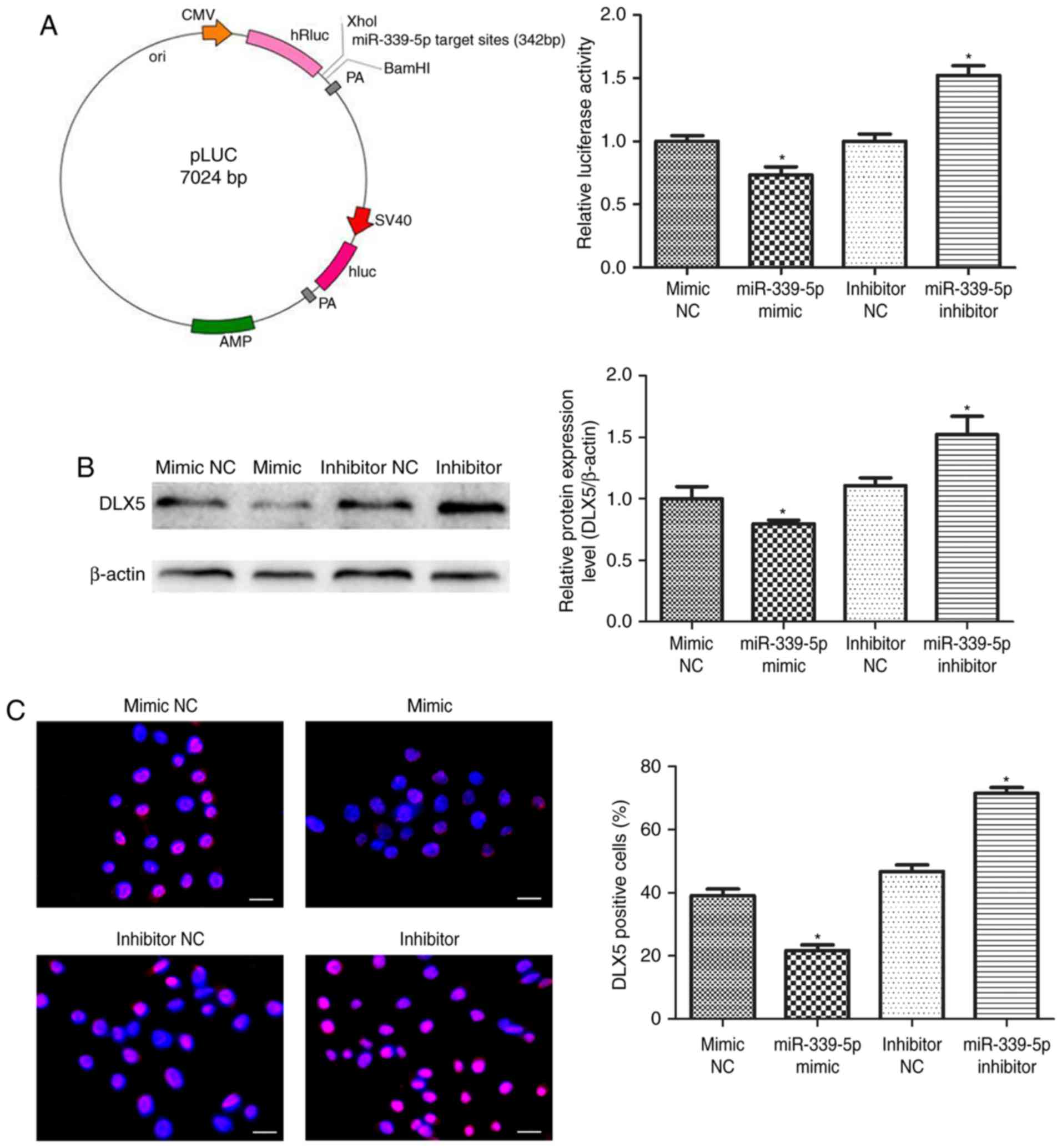
DLX5 is a potential target of
miR-339-5p
HFSCs were co-transfected with the luciferase
reporter carrying miR-339-5p target sites in the pLUC-DLX5 cloning
vector (Fig. 4A, left), as well as
the miR-339-5p mimic, miR-339-5p inhibitor or negative controls.
The luciferase activity of pLUC-DLX5, containing the DLX5
3′untranslated region (UTR), was significantly reduced by the
miR-339-5p mimics and markedly increased with the miR-339-5p
inhibitor (Fig. 4A, right).
Western blot and immunofluorescence analyses revealed that reduced
endogenous DLX5 expression was due to increased miR-339-5p
expression in the mimic group, and increased endogenous DLX5
expression was due to reduced miR-339-5p expression in the presence
of the miR-339-5p inhibitor (Fig. 4B
and C). These results suggest that miR-339-5p directly
regulates the expression of DLX5.
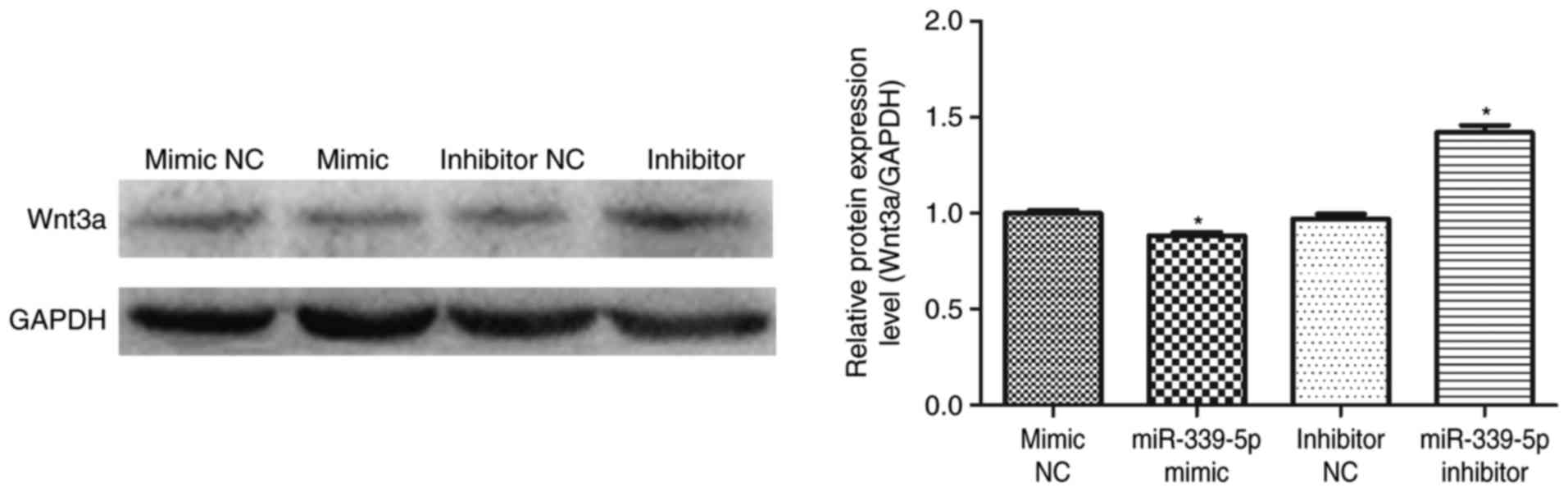
Downregulation of miR-339-5p increases
Wnt3a protein expression in HFSCs
miR-339-5p was transfected into HFSCs and western
blot analysis was performed to determine if miR-339-5p was involved
in the regulation of Wnt3a expression. The results demonstrated
that Wnt3a expression decreased in the miR-339-5p mimic group
compared with the mimic NC group. Furthermore, Wnt3a levels
increased in the miR-339-5p inhibitor group compared with the
inhibitor NC group (Fig. 5).
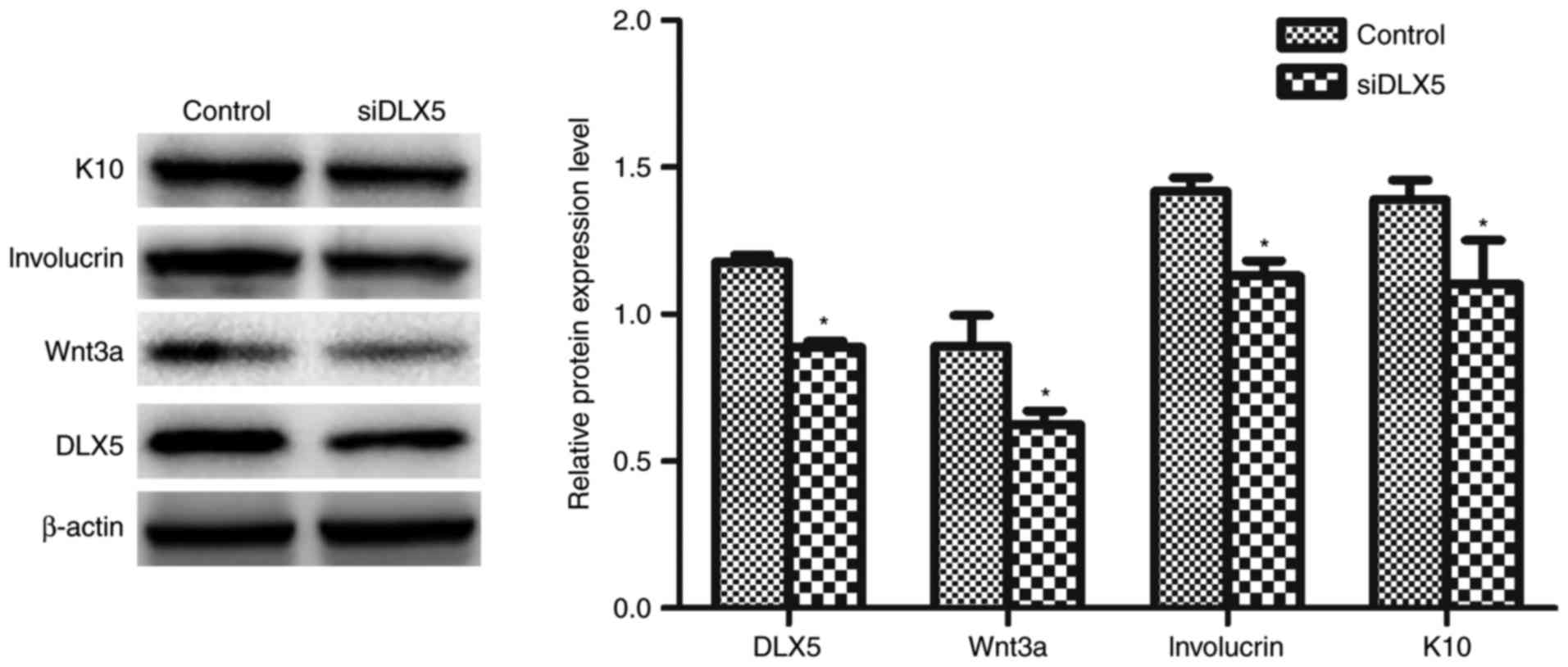
Downregulation of DLX5 inhibits HFSCs
differentiation
siDLX5 was transfected into HFSCs and western blot
analysis was performed to determine if DLX5 was involved in the
regulation of Wnt3a, involucrin and K10 expression. The results
revealed that Wnt3a, involucrin and K10 expression was
significantly decreased in the siDLX5 group compared with the
control group (Fig. 6).
Discussion
HFSCs are located in the bulge region of the outer
root sheath of the hair follicle and exhibit high proliferative and
differential capacity. It is widely accepted that CD34 and K15 are
markers of HFSCs (33,34). In the present study, the cultured
hair follicle cells positive expression of CD34 and K15 was
confirmed, indicating that the separated epithelioid cells were
HFSCs. Our previous study demonstrated that HFSC-seeded
tissue-engineered skin could repair full-thickness skin defects
(35), and that loureirin A
accelerates the healing process via the Wnt/β-catenin signaling
pathway (12). However, if and how
miRNAs modulate loureirin A-induced HFSC differentiation remains
unclear. The present study revealed that downregulation of
miR-339-5p expression promoted loureirin A-induced HFSC
differentiation; DLX5 was a potential target of miR-339-5p; and the
Wnt/β-catenin signaling pathway may be involved in the regulatory
effects of miR-339-5p/DLX5. These findings suggest that miR-339-5p
and DLX5 may be key targets in drug development for wound
healing.
The present study demonstrated that miR-339-5p
expression downregulation promoted loureirin A-induced HFSC
differentiation. Microarray data and RT-qPCR analyses identified
that miR-339-5p was downregulated in HFSCs induced by loureirin A.
These results are consistent with the hypothesis that certain
miRNAs can be differently expressed in the skin (36). A previous study indicated that
miR-339-5p is involved in the development and metastasis of certain
cancers (37). However, little is
known about the role of miR-339-5p in HFSCs. The results of the
present study demonstrate that miR-339-5p was downregulated in
HFSCs treated with loureirin A. Additionally, it was revealed that
the expression of K10 and involucrin decreased with miR-339-5p
mimic transfection, and expression increased with miR-339-5p
inhibitor transfection. Therefore, it was hypothesized that HFSC
differentiation may be associated with miR-339-5p expression.
In order to further identify the function of
miR-339-5p in HFSC differentiation, miR-339-5p mimics and
inhibitors were transfected into HFSCs. Western blot analysis was
used to detect the expression of proteins associated with HFSC
differentiation. K10 and involucrin are epidermal differentiation
markers (38,39). The expression of both these
proteins was significantly reduced in the miR-339-5p mimic
transfection group, and significantly increased in the miR-339-5p
inhibitor transfected group. These data indicate that miR-339-5p
may have negatively regulated the differentiation of HFSCs.
In the current study, it was revealed that DLX5 was
targeted by miR-339-5p. DLX5 is a transcription factor that can
modulate epithelial cell differentiation. Therefore, the focus of
the present study was on DLX5. The pLUC-DLX5 cloning vector, which
contained miR-339-5p target sites in the DLX5 3′UTR, was
constructed to co-transfect with the miR-339-5p mimic or inhibitor.
The results revealed that the luciferase activity of pLUC-DLX5 was
significantly reduced by miR-339-5p mimic, and markedly increased
by the miR-339-5p inhibitor. To further confirm this result,
miR-339-5p mimics and inhibitor were transfected into HFSCs and
DLX5 protein expression was detected by western blot and
immunofluorescence analyses. DLX5 expression was significantly
reduced in the miR-339-5p mimic transfection group, compared with
the mimic NC group. Similarly, DLX5 expression was significantly
increased in the miR-339 inhibitor transfection group, compared
with the inhibitor NC group. Taken together, these results support
the hypothesis that DLX5 was targeted by miR-339-5p, which may be
directly responsible for HFSC differentiation.
Notably, the present study demonstrated that the
Wnt/β-catenin signaling pathway was involved in the regulatory
effects of miR-339-5p/DLX5. Wnt3a expression significantly
decreased in the miR-339-5p mimic group and significantly increased
in the miR-339-5p inhibitor group. siDLX5 was transfected into
HFSCs and western blot analysis revealed that Wnt3a, involucrin and
K10 expression was significantly downregulated. Wnt3a, an essential
component of the Wnt/β-catenin pathway (40), induces the differentiation of
mesenchymal stem cells into epidermal-like cells through activation
of the classic Wnt signaling pathway (41). Furthermore, our previous results
reported that loureirin A promotes HFSC differentiation by
activating the Wnt/β-catenin signaling pathway (12). From these data, it may be
hypothesized that miR-339-5p decreased HFSC differentiation by
regulating DLX5 expression, resulting in Wnt/β-catenin signaling
pathway inhibition. However, the present study was conducted in
vitro. The precise role of miR-339-5p/DLX5 in the
differentiation of HFSCs must be fully elucidated in
vivo.
In conclusion, the present study demonstrated that
miR-339-5p is a negative regulator in loureirin A-induced HFSC
differentiation. Downregulation of miR-339-5p expression results in
increased DLX5 expression. The present study provided evidence for
miR-339-5p and DLX5 as potential therapeutic molecular targets for
the treatment of skin wounds.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural
Science Foundation of Guangdong Province (grant no. 2017A030312009)
and the Educational Commission of Guangdong Province (grant no.
2016KTSCX020).
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
DC and AL conceptualized and developed the study
design. XL, YW and FX performed most of the experiments. FZ and SZ
prepared the cells for experimentation. JZ participated in western
blot and immunofluorescence assays. FZ, SZ and JZ acquired and
analyzed the data. XL, FZ, SZ and JZ discussed the results and
wrote the manuscript. DC and AL made comments, suggested
appropriate modifications and made corrections. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Guangzhou University of Chinese Medicine.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
miR-339-5p
|
microRNA-339-5p
|
|
HFSCs
|
hair follicle stem cells
|
|
DLX5
|
homeobox protein DLX-5
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
K10
|
keratin, type I cytoskeletal 10
|
|
K15
|
keratin, type I cytoskeletal 15
|
References
|
1
|
Yeh DD, Nazarian RM, Demetri L, Mesar T,
Dijkink S, Larentzakis A, Velmahos G and Sadik KW:
Histopathological assessment of OASIS Ultra on critical-sized wound
healing: A pilot study. J Cutan Pathol. 44:523–529. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hartmann-Fritsch F, Marino D and Reichmann
E: About ATMPs, SOPs and GMP: The hurdles to produce novel skin
grafts for clinical use. Transfus Med Hemothr. 43:344–352. 2016.
View Article : Google Scholar
|
|
3
|
Hirsch T, Rothoeft T, Teig N, Bauer JW,
Pellegrini G, De Rosa L, Scaglione D, Reichelt J, Klausegger A,
Kneisz D, et al: Regeneration of the entire human epidermis using
transgenic stem cells. Nature. 551:327–332. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang Y, Zhang W, Li Y, Fang G and Zhang K:
Scalded skin of rat treated by using fibrin glue combined with
allogeneic bone marrow mesenchymal stem cells. Ann Dermatol.
26:289–295. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Strong AL, Neumeister MW and Levi B: Stem
cells and tissue engineering regeneration of the skin. Clin Plast
Surg. 44:635–650. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Veijouyeh Joulai S, Mashayekhi F, Yari A,
Heidari F, Sajedi F, Ghoroghi Moghani F and Nobakht M: In vitro
induction effect of 1,25(OH)2D3 on
differentiation of hair follicle stem cell into keratinocyte.
Biomed J. 40:31–38. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang H, Zhao H, Qiao J, Zhang S, Liu S,
Li N, Lei X, Ning L, Cao Y and Duan E: Expansion of hair follicle
stem cells sticking to isolated sebaceous glands to generate in
vivo epidermal structures. Cell Transplant. 25:2071–2082. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Quan R, Du W, Zheng X, Xu S, Li Q, Ji X,
Wu X, Shao R and Yang D: VEGF165 induces differentiation of hair
follicle stem cells into endothelial cells and plays a role in in
vivo angiogenesis. J Cell Mol Med. 21:1593–1604. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Najafzadeh N, Sagha M, Tajaddod Heydari S,
Golmohammadi MG, Oskoui Massahi N and Moghaddam Deldadeh M: In
vitro neural differentiation of CD34+ stem cell
populations in hair follicles by three different neural induction
protocols. In Vitro Cell Develop Biol-Anim. 51:192–203. 2015.
View Article : Google Scholar
|
|
10
|
Flores A, Schell J, Krall AS, Jelinek D,
Miranda M, Grigorian M, Braas D, White AC, Zhou JL, Graham NA, et
al: Lactate dehydrogenase activity drives hair follicle stem cell
activation. Nat Cell Biol. 19:1017–1026. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xu J, Xiong T, Yang Y, Li J and Mao J:
Resina draconis as a topical treatment for pressure ulcers:
A systematic review and meta-analysis. Wound Repair Regen.
23:565–574. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu A, Du B, Yi H, Tang Y, Li X, Zhang S,
Zhou J and Chen D: Loureirin A activates Wnt/β-catenin pathway to
promote wound with follicle stem cell-seeded tissue-engineered skin
healing. J Biomater Tiss Eng. 6:427–432. 2016. View Article : Google Scholar
|
|
13
|
Chandra S, Vimal D, Sharma D, Rai V, Gupta
SC and Chowdhuri DK: Role of miRNAs in development and disease:
Lessons learnt from small organisms. Life Sci. 185:8–14. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hu Y, Rao SS, Wang ZX, Cao J, Tan YJ, Luo
J, Li HM, Zhang WS, Chen CY and Xie H: Exosomes from human
umbilical cord blood accelerate cutaneous wound healing through
miR-21-3p-mediated promotion of angiogenesis and fibroblast
function. Theranostics. 8:169–184. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Reza AMMT, Choi YJ and Kim JH: MicroRNA
and transcriptomic profiling showed miRNA-dependent impairment of
systemic regulation and synthesis of biomolecules in rag2 ko mice.
Molecules. 23:E5272018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ojeh N, Pastar I, Tomic-Canic M and
Stojadinovic O: Stem cells in skin regeneration, wound healing, and
their clinical applications. Int J Mol Sci. 16:25476–25501. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Amelio I, Lena AM, Bonanno E, Melino G and
Candi E: miR-24 affects hair follicle morphogenesis targeting
Tcf-3. Cell Death Dis. 4:e9222013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang L, Ge Y and Fuchs E: miR-125b can
enhance skin tumor initiation and promote malignant progression by
repressing differentiation and prolonging cell survival. Genes Dev.
28:2532–2546. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ahmed MI, Alam M, Emelianov VU,
Poterlowicz K, Patel A, Sharov AA, Mardaryev AN and Botchkareva NV:
MicroRNA-214 controls skin and hair follicle development by
modulating the activity of the Wnt pathway. J Cell Biol.
207:549–567. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yuan S, Li F, Meng Q, Zhao Y, Chen L,
Zhang H, Xue L, Zhang X, Lengner C and Yu Z: Post-transcriptional
regulation of keratinocyte progenitor cell expansion,
differentiation and hair follicle regression by miR-22. PLoS Genet.
11:e10052532015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jonas S and Izaurralde E: Towards a
molecular understanding of microRNA-mediated gene silencing. Nat
Rev Genet. 16:421–433. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jansson MD, Damas ND, Lees M, Jacobsen A
and Lund AH: miR-339-5p regulates the p53 tumor-suppressor pathway
by targeting MDM2. Oncogene. 34:1908–1918. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhou C, Liu G, Wang L, Lu Y, Yuan L, Zheng
L, Chen F, Peng F and Li X: MiR-339-5p regulates the growth, colony
formation and metastasis of colorectal cancer cells by targeting
PRL-1. PLoS One. 8:e631422013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ichi S, Costa FF, Bischof JM, Nakazaki H,
Shen YW, Boshnjaku V, Sharma S, Mania-Farnell B, McLone DG, Tomita
T, et al: Folic acid remodels chromatin on Hes1 and Neurog2
promoters during caudal neural tube development. J Biol Chem.
285:36922–36932. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hu B, Wang Q, Wang YA, Hua S, Sauvé CG,
Ong D, Lan ZD, Chang Q, Ho YW, Monasterio MM, et al: Epigenetic
activation of WNT5A drives glioblastoma stem cell differentiation
and invasive growth. Cell. 167:1281–1295. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rakowiecki S and Epstein DJ: Divergent
roles for Wnt/β-catenin signaling in epithelial maintenance and
breakdown during semicircular canal formation. Development.
140:1730–1739. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Babaie Y, Herwig R, Greber B, Brink TC,
Wruck W, Groth D, Lehrach H, Burdon T and Adjaye J: Analysis of
Oct4-dependent transcriptional networks regulating self-renewal and
pluripotency in human embryonic stem cells. Stem Cells. 25:500–510.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wolfrum K, Wang Y, Prigione A, Sperling K,
Lehrach H and Adjaye J: The LARGE principle of cellular
reprogramming: Lost, acquired and retained gene expression in
foreskin and amniotic fluid-derived human iPS cells. PLoS One.
5:e137032010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Huang E, Lian X, Chen W, Yang T and Yang
L: Characterization of rat hair follicle stem cells selected by
vario magnetic activated cell sorting system. Acta Histochem
Cytochem. 42:129–136. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang G, Zhao T, Wang L, Hu B, Darabi A,
Lin J, Xing MM and Qiu X: Studying different binding and
intracellular delivery efficiency of ssDNA single-walled carbon
nanotubes and their effects on LC3-related autophagy in renal
mesangial cells via miRNA-382. ACS Appl Mater Interfaces.
7:25733–25740. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zheng J, Yu L, Chen W, Lu X and Fan X:
Circulating exosomal microRNAs reveal the mechanism of fructus
meliae toosendan-induced liver injury in mice. Sci Rep.
8:28322018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ge W, Zhao Y, Lai FN, Liu JC, Sun YC, Wang
JJ, Cheng SF, Zhang XF, Sun LL, Li L, et al: Cutaneous applied
nano-ZnO reduce the ability of hair follicle stem cells to
differentiate. Nanotoxicology. 11:465–474. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chacon-Martinez CA, Klose M, Niemann C,
Glauche I and Wickstrom SA: Hair follicle stem cell cultures reveal
self-organizing plasticity of stem cells and their progeny. EMBO J.
36:151–164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu A, Chen X, Yi H, Li F and Chen D:
Accelerating the healing of skin defects transplanted FSC-seeded
tissue-engineered skin. J Biomater Tiss Eng. 5:574–578. 2015.
View Article : Google Scholar
|
|
36
|
Aberdam D, Candi E, Knight R and Melino G:
miRNAs, ‘stemness’ and skin. Trends Biochem Sci. 33:583–591. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yan H, Zhao M, Huang S, Chen P, Wu WY,
Huang J, Wu ZS and Wu Q: Prolactin inhibits BCL6 expression in
breast cancer cells through a microRNA-339-5p-dependent pathway. J
Breast Cancer. 19:26–33. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pilehvar-Soltanahmadi Y, Nouri M, Martino
MM, Fattahi A, Alizadeh E, Darabi M, Rahmati-Yamchi M and Zarghami
N: Cytoprotection, proliferation and epidermal differentiation of
adipose tissue-derived stem cells on emu oil based electrospun
nanofibrous mat. Exp Cell Res. 357:192–201. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sundaramurthi D, Krishnan UM and
Sethuraman S: Epidermal differentiation of stem cells on
poly(3-hydroxybutyrate-co-3-hydroxyvalerate) (PHBV) nanofibers. Ann
Biomed Eng. 42:2589–2599. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shimomura Y, Agalliu D, Vonica A, Luria V,
Wajid M, Baumer A, Belli S, Petukhova L, Schinzel A, Brivanlou AH,
et al: APCDD1 is a novel Wnt inhibitor mutated in hereditary
hypotrichosis simplex. Nature. 464:1043–1047. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sun TJ, Tao R, Han YQ, Xu G, Liu J and Han
YF: Wnt3a promotes human umbilical cord mesenchymal stem cells to
differentiate into epidermal-like cells. Eur Rev Med Pharmacol Sci.
19:86–91. 2015.PubMed/NCBI
|