Introduction
DNA methylation at cytosines in CpG islands located
in promoters is well known as one of the earliest molecular
alteration occurring during carcinogenesis and specific for the
malignant state (1). Over the past
few years, there have been increasing evidences asserting the role
of DNA methylation at promoter of genes encoding small non coding
microRNAs (MIRs) that act as posttranscriptional regulators
of gene expression (2,3). Aberrant expression of microRNAs
regulated by DNA methylation is involved in many cellular processes
such as DNA repair, cell cycle, apoptosis, through which they
promote cell differentiation, proliferation, malignant
transformation and tumorigenesis (4,5). For
example, the down-regulation of MIR129-2 by DNA methylation
regulates breast cancer cell proliferation and apoptosis (6). Moreover, MIR449c expression,
which was significantly down-regulated by DNA methylation in
osteosarcoma cancer, was negatively correlated with tumor size and
tumor stages (7). In addition,
integrating DNA methylation data with microRNA expression profile
in various types of cancers including lung, colon and breast has
been extensively explored by genome wide analysis recently
(8,9).
Among the huge number of microRNA genes, the
MIR34 gene family, which consists of three genes MIR34a,
MIR34b and MIR34c, has been the focus of numerous
studies in cancer research. All the three MIR34 genes are
transcriptionally regulated by p53 protein, a regulator of cell
cycle and apoptosis; therefore, they act as tumor suppressor genes
by targeting many oncogenes related to proliferation, apoptosis and
invasion (10,11). Numerous studies have demonstrated
the direct link between dysregulations concerning MIR34
family and epigenetic and genetic mechanisms in cancers. For
instance, MIR34a targets the proto-oncogene c-SRC to
attenuate tumor growth in triple-negative breast cancer (12), or programmed death ligand 1 (PDL1)
to modulate the tumor immune response in non small cell lung cancer
(13). Stahlhut and Slack revealed
that combinatorial action of MIR34a and microRNA let-7
effectively synergizes with erlotinib to suppress non-small cell
lung cancer cell proliferation (14). MIR34b and MIR34c,
which share a common primary transcript, function as metastasis
suppressors in lung adenocarcinoma (15,16).
A recent study on circulating MIR34s in 173 patients with
triple-negative breast cancer indicated that MIR34a, MIR34b
and MIR34c expression was respectively correlated with tumor
grade (P=0.038), lymph node positivity (P=0.027) and distant
metastasis (P<0.001) (17).
Importantly, the aberrant low expression of MIR34s was
associated with promoter methylation. Indeed, data integration from
104 studies on microRNAs revealed that MIR34s are silenced
by DNA methylation in the highest number of cancer types (18). Epigenetic inactivation of the
MIR34a was found in ovarian, colorectal, lung, kidney and
breast cancer cell lines (19,20).
The presence of MIR34b/c promoter methylation was
significantly associated with the absence of its transcripts and
with metastasis formation in primary tumors of colon, lung, head
and neck, melanomas and breast cancers (21). Siemens et al (22) showed that MIR34a methylation
is involved in the up-regulation of c-Met, Snail, and β-catenin
proteins, which was associated with the metastasis distance of
colon cancer cells to the liver. Patients after surgery of lung
cancer with aberrant methylation of MIR34b/c had a high
probability of recurrence and poor prognosis (P=0.026) (23). Furthermore, molecular events from
negative surgical margin have been extremely investigated because
of their predictive and prognostic values for tumor progression,
local recurrence, metastasis and overall survival (24). Multi-platform analyses of DNA
defects, epigenetics, and gene expression in cancer-adjacent
tissues have extensively been performed recently to provide
integrative data to the clinic (25,26).
DNA methylation that has occurred at the primary tumor is believed
to progressively spread outwards to surrounding tissues (27,28).
Increased DNA methylation level in ductal carcinoma in situ
is related with future development of invasive breast cancer and
with the cancer metastasis distance (29,30).
Currently, MIR34 methylation profile in normal adjacent and
tumor tissues, and its correlation with clinicopathological
features have been frequently reported to gastrointestinal cancer.
For instance, MIR34 methylation occurs in colorectal normal
adjacent (43.9%) and tumor tissues (79.3%) and correlated with
positive lymph node (P=0.01) (31). Similarly, MIR34b/c
methylation was considerably different between adjacent (22.7%) and
tumors (70%) in gastric cancer (32). However, differences in MIR34
methylation profile in either non-cancerous or normal adjacent
tissues as compared with breast and lung cancer tissues have been
rarely described so far while both types of these cancers are the
first and second common types of cancer and leading cause of cancer
death all over the world (33).
Therefore, investigating the methylation profile of the
MIR34 genes in both types of cancer vs. normal adjacent
tissues and non-cancerous tissues and moreover their association
with the clinicopathological features would provide a comprehensive
evaluation on the synergy of these potential methylation biomarkers
for cancer diagnosis.
In this study, we investigated the methylation
status at the promoter of the genes encoding MIR34a/b/c in
breast cancer vs. normal adjacent tissues, as well as in
non-cancerous lung diseases vs. lung cancer, with samples coming
from Vietnamese patients. The objective is to evaluate the
methylation profiles of these genes both individually and in an
integrative manner in order to establish new integrative
methylation biomarkers for cancer detection. Furthermore, the
comparison of the methylation profiles of these genes in cancer vs.
non-cancerous or normal adjacent tissues has highlighted the
epigenetically concomitant changes of these genes in tissues that
are physiologically different such as breast and lung.
Materials and methods
Sample collection
Surgically resected specimens from breast carcinomas
and matched adjacent tissues were collected from 79 breast cancer
patients having undergone mastectomy at the Department of
Pathology, National Cancer Hospital K (Hanoi, Vietnam) between 2012
and 2013. The corresponding adjacent tissue samples were selected
3–5 cm away from the site at which the primary tumor was obtained.
Breast tumor and corresponding adjacent tissues were snap-frozen in
liquid nitrogen immediately after resection and examination by
pathologists, and stored at −80°C until further used.
Formalin-fixed, paraffin-embedded (FFPE) tissue
specimens were collected from 95 lung cancer patients and 72
patients suffering from non-cancerous lung diseases (whose
classification was examined by pathologists) at the Department of
Pathology, 175 Hospital (Ho Chi Minh, Vietnam) during 2016.
Informed consent was obtained from patients in written form and the
study was approved by the guidelines of the VNU University of
Science ethical committee in Vietnam (106-YS.06-2015.07).
Genomic DNAs extraction and bisulfite
modification
Genomic DNAs were extracted from freshly frozen
breast or FFPE lung tissues by using the QIAampDNA Mini kit or
QIAamp DNA FFPE Tissue kit (Qiagen, Inc., Valencia, CA, USA), and
treated with sodium bisulfite by using the EpiTect Bisulfite kit
(Qiagen, Inc.). During the treatment, the unmethylated cytosines of
the genomic DNAs were converted into uracils, but the methylated
cytosines remained unchanged (34). Polymerase chain reaction (PCR)
realized on native DNA using primer sets specific to methylated
sequences was performed to confirm the accuracy of the primer sets.
PCR realized on the bisulfite-treated DNA using primer sets
specific to unmethylated sequences of the β-globin gene was
performed to determine the efficiency of bisulfite conversion
(35).
Methylation specific PCR (MSP)
The methylation status of the investigated genes was
evaluated by using MSP to amplify bisulfite treated DNA with
primers that distinguish methylated (M) and unmethylated (U) DNA
(36). MIR34a gene locates
on chromosome 1 and is transcribed from the minus strand. The
MIR34a gene structure is described by Tarasov et al
(37) and the CpG island in its
promoter is indicated by Lodygin et al (19). MIR34b/c gene locates on
chromosome 16 and is transcribed from the plus strand. The CpG
island of MIR34b/c promoter locates in the upstream sequence
of the BTG4 gene as described by Toyota et al
(38). Based on these reports, we
look for the sequences corresponding to MIR34a, MIR34b/c
promoters and designed the MSP primer sets used in our study. The
primers for detecting the methylation status of MIR34s were
designed based on the primer designing tool for MSP method
(http://www.urogene.org/methprimer/index1.html). The
primer sequences, MSP conditions and amplicon lengths are shown in
Table I. Bisulfite treated DNAs
were subjected to single or nested PCR depending on the particular
targeted genes. The MSP products were resolved by electrophoresis
in 8% polyacrylamide gel, then stained with ethidium bromide and
imaged with the UVP, LLC (Upland, CA, USA). DNA extracted from
lymphocytes of healthy volunteers and treated with bisulfite was
used as a positive control for unmethylation of the targeted genes
in numerous reports in which the MSP method was also applied
(23,36). Water with no DNA template was
included in each PCR reaction as a control for contamination. All
MSP reactions were performed in duplicate. The methylation status
was confirmed by direct sequencing of the MSP products for a subset
of samples from each assay.
 | Table I.MSP primers for analysis of
MIR34a, MIR34b/c methylation. |
Table I.
MSP primers for analysis of
MIR34a, MIR34b/c methylation.
| Primers | Sequence
(5′-3′) | Amplicon size
(bp) | MSP conditions |
|---|
| MIR34a
EF570049.1 |
|
miR-34a-meF |
TTTTGGGTAGGCGCGTTTCGC | Round 1: 147 | 94°C 5 min, 40
cycles of (94°C 30 sec, 60°C 10 sec, 72°C 15 sec), 72°C 5 min |
|
miR-34a-meR |
CCAATCCCGCCGAACACGAAA |
|
|
|
miR-34a-meF |
TTTTGGGTAGGCGCGTTTCGC | Round 2: 100 | 94°C 5 min, 40
cycles of (94°C 30 sec, 63°C 10 sec, 72°C 15 sec), 72°C 5 min |
|
miR-34a-meR1 |
GCCCCCGCCTAAACTAACG |
|
|
| miR
−34a-unF |
GGTGGTGTTTTGTGATTTAGT GGTGGT | 143 | 94°C 5 min, 40
cycles of (94°C 30 sec, 63°C 10 sec, 72°C 15 sec), 72°C 5 min |
|
miR-34-unR |
CAAAACCAATCCCACCAAACA CAAAATC |
|
|
| MIR34b/c
BC021736 |
|
miR-34b/c-meF |
TCGTTTCGTTTCGCGTTCGTT | 93 | 94°C 1 min, 40
cycles of (94°C 30 sec, 66°C 30 sec, 72°C 30 sec), 72°C 5 min |
|
miR-34b/c-meR |
GCCGCTCTAAACGACCGAAT |
|
|
|
miR-34b/c-unF |
TTGTGGGGTTTTAAGGATGGTT | 160 | 94°C 1 min, 40
cycles of (94°C 30 sec, 66°C |
|
| GGTTGTTT |
| 30 sec, 72°C 30
sec), 72°C 5 min |
|
miR-34b/c-unR |
CCCTTCACCTCCTCAACCCAAAC |
|
|
Statistical analysis
Chi-square test was used to determine the difference
in MIR34s methylation levels between cancer and
non-cancerous/normal adjacent tissues. The Kappa statistic was used
to assess the agreement between two dichotomous variables such as
the concordance between the methylation status of genes analyzed
two by two in a given tissue type or the concordance between the
methylation status of individual gene in tumor and adjacent normal
tissues. Chi-square test and Fisher's exact test was used to
examine the association of the methylation status of individual or
combined genes with clinicoathological characteristics. For all
statistical analyses, a P<0.05 was considered to indicate a
statistically significant difference. All analyses were done by
using the STATA program version 12 (StataCorp LP, College Station,
TX, USA; http://www.stata.com/).
Results
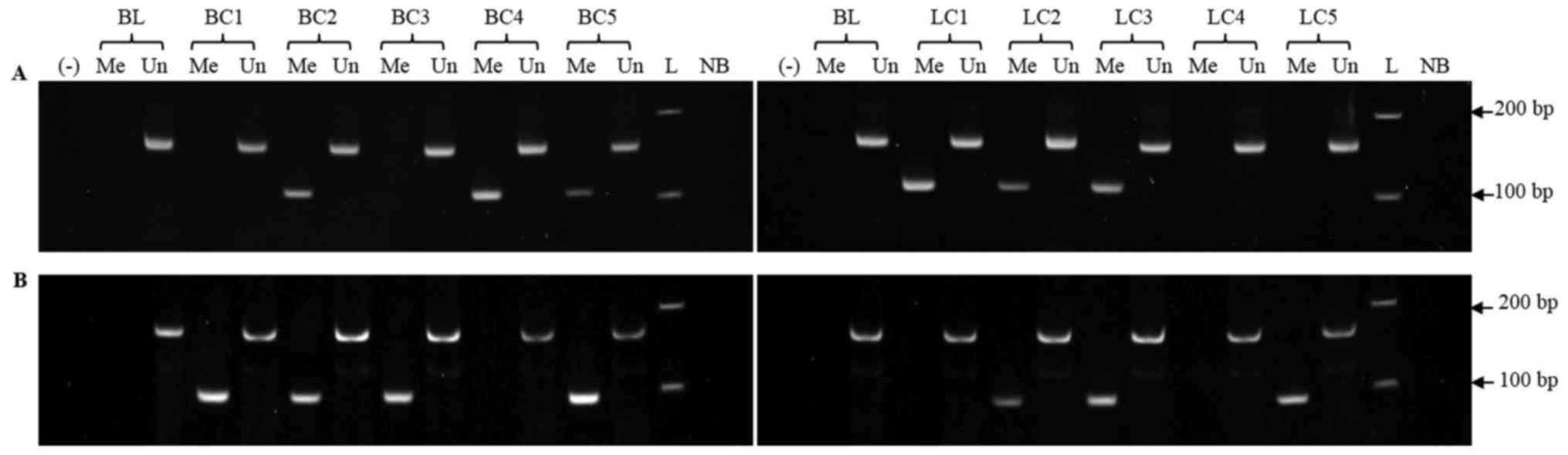
In order to assess the methylation status of
MIR34 genes, we first set up an MSP assay and verified its
specificity, since we have previously shown that false positive
results from the MSP assay could be due to the MSP primers
amplifying nonspecifically untreated genomic DNAs (35). Native DNAs were subjected to MSP
analysis with the primer sets specific to the methylated status of
the targeted sequences MIR34a and MIR34b/c. No MSP
products were amplified from untreated DNAs extracted from
lymphocytes of healthy donors (Fig.
1), confirming that the MSP primers were specific for the
methylated targets and that false positive results have been
avoided.
Genomic DNAs were subsequently treated with
bisulfite and then subjected to the MSP assays. Representative
results of the MSP reactions were illustrated in Fig. 1. The MSP products specific to
methylated sequences of the MIR34 promoters were directly
sequenced. The results showed that all cytosines in the CpG sites
remained cytosines and the cytosines alone were converted to
thymidines (data not shown), indicating the completely conversion
of genomic DNA by bisulfite treatment.
The MSP analysis revealed that methylation of
MIR34 promoters occurred in all analyzed tissues (breast
cancer and adjacent tissues as well as in lung cancer and
non-cancerous tissues). Among 79 pairs of matched breast cancer and
adjacent tissues, MIR34a methylation occurred with the
frequency of 49.37% in cancer tissue, which is significantly higher
than its frequency of 30.38% in normal adjacent tissue (P=0.015).
These frequencies for MIR34b/c methylation were 59.49 and
62.03%, respectively, with no significant difference. The
methylation frequencies of MIR34a and MIR34b/c in
lung cancer (48.42 and 56.84%) were similar to those in
non-cancerous lung diseases (47.22 and 51.39%) (Table II).
| Table II.Methylation profile of the MIR34
genes in breast and lung cancers. |
Table II.
Methylation profile of the MIR34
genes in breast and lung cancers.
|
| Number of
methylated cases (%) |
|---|
|
|
|
|---|
| Tissue | miRa | P-value | miRb/c | P-value |
|---|
| Breast cancer |
| 0.015a |
| 0.745 |
| Tumor
(n=79) | 39 (49.37) |
| 47 (59.49) |
|
|
| [38.34; 60.39] |
| [48.67; 70.32] |
|
| Normal
adjacent tissue (n=79) | 24 (30.38) |
| 49 (62.03) |
|
|
| [20.24; 40.52] |
| [51.32; 72.73] |
|
| Lung cancer |
| 0.878 |
| 0.483 |
| Tumor
(n=95) | 46 (48.42) |
| 54 (56.84) |
|
|
| [38.37; 58.47] |
| [46.88; 66.80] |
|
|
Pulmonary diseases (n=72) | 34 (47.22) |
| 37 (51.39) |
|
|
| [35.69; 58.75] |
| [39.84; 62.93] |
|
As assessed by the calculation of the Kappa
coefficient, the methylation status of MIR34a showed a
significant concordance with that of MIR34b/c in breast
cancer tissue but not in normal adjacent tissues (OR=2.12, 95% CI:
1.04–4.28, P=0.04). On the contrary, a significant concordance of
the methylation status of both MIR34a and MIR34b/c
genes was observed in lung cancer as well as in non-cancerous
pulmonary diseases (P=0.0001; <0.0001) (Table II). Additionally, the methylation
status of each gene promoter was also analyzed in association with
clinicopathological features such as the histological tumor type,
the tumor grade and the metastasis status (Table III). For breast cancer, the
results showed that the methylation frequency of MIR34a was
significantly associated with tumor type IDC (P=0.02). The
univariate logistic analysis indicated that MIR34a was more
methylated in the IDC type than in the other carcinoma types
(OR=6.17, 95% CI: 1.25–30.32, P=0.03). However, no significant
differences in the methylation frequency of the MIR34b/c
gene were associated with clinical features of breast cancer
patients such as the tumor grade and the metastasis status, nor
with the patient' age. As far as lung cancer and non-cancerous
pulmonary diseases are concerned, there was no significant
association of the methylation frequency of MIR34a and
MIR34b/c with any clinical features nor with the patient'
age or sex.
| Table III.Association of the methylation status
of the MIR34s genes with the clinicopathologicalcharacteristics of
the 79 breast cancer, 95 lung cancer and 72 non-cancerous pulmonary
disease patients. |
Table III.
Association of the methylation status
of the MIR34s genes with the clinicopathologicalcharacteristics of
the 79 breast cancer, 95 lung cancer and 72 non-cancerous pulmonary
disease patients.
| A, Breast
cancer |
|---|
|
|---|
|
|
| miR34a |
| miR34b/c |
|
|---|
|
|
|
|
|
|
|
|---|
| Feature | No. of
patients | Un | Me | P-value | Un | Me | P-value |
|---|
| Histological tumor
type | 79 |
|
| 0.015b |
|
| 0.274 |
|
IDC | 67 | 30 | 37 |
| 25 | 42 |
|
|
ILC | 5 | 3 | 2 |
| 2 | 3 |
|
|
Other | 7 | 7 | 0 |
| 5 | 2 |
|
| Tumor grade | 65 |
|
| 0.999 |
|
| 0.382 |
| Grade
1 | 3 | 1 | 2 |
| 0 | 3 |
|
| Grade
2 | 54 | 24 | 30 |
| 22 | 32 |
|
| Grade
3 | 8 | 4 | 4 |
| 2 | 6 |
|
| Metastasis | 79 |
|
| 0.934a |
|
| 0.753a |
| No | 51 | 26 | 25 |
| 20 | 31 |
|
|
Yes | 28 | 14 | 14 |
| 12 | 16 |
|
| Age, years | 79 |
|
| 0.749a |
|
| 0.192a |
|
<50 | 29 | 14 | 15 |
| 9 | 20 |
|
|
≥50 | 50 | 26 | 24 |
| 23 | 27 |
|
|
| B, Lung
cancer |
|
|
|
| miR34a |
|
miR34b/c |
|
|
|
|
|
|
|
|
| Feature | No. of
patients | Un | Me | P-value | Un | Me | P-value |
|
| Histological tumor
type | 95 |
|
| 0.809 |
|
| 0.786 |
|
NSCLC | 72 | 38 | 34 |
| 30 | 42 |
|
|
SCLC | 1 | 0 | 1 |
| 0 | 1 |
|
|
Other | 22 | 11 | 11 |
| 11 | 11 |
|
| Stage | 79 |
|
| 0.739 |
|
| 0.311 |
| I | 2 | 1 | 1 |
| 0 | 2 |
|
| II | 28 | 17 | 11 |
| 11 | 17 |
|
|
III | 49 | 25 | 24 |
| 25 | 24 |
|
| Sex | 94 |
|
| 0.877a |
|
| 0.961a |
|
Female | 21 | 21 | 20 |
| 18 | 23 |
|
|
Male | 28 | 28 | 25 |
| 23 | 30 |
|
| Age, years | 86 |
|
| 0.999 |
|
| 0.999 |
|
<50 | 10 | 5 | 5 |
| 4 | 6 |
|
|
≥50 | 76 | 41 | 35 |
| 35 | 41 |
|
| EGFR mutation | 44 |
|
| 0.480 |
|
| 0.484 |
| No | 11 | 3 | 8 |
| 4 | 7 |
|
|
Yes | 33 | 15 | 18 |
| 16 | 17 |
|
|
| C, Lung
diseases |
|
|
|
| miR34a |
|
miR34b/c |
|
|
|
|
|
|
|
|
| Feature | No. of
patients | Un | Me | P-value | Un | Me | P-value |
|
| Diagnosis | 71 |
|
| 0.768 |
|
| 0.306 |
|
Aspergillus | 9 | 3 | 6 |
| 4 | 5 |
|
|
Tuberculosis | 16 | 8 | 8 |
| 8 | 8 |
|
|
Pneumonia | 22 | 11 | 11 |
| 8 | 14 |
|
|
Pulmonary gas pressures | 17 | 10 | 7 |
| 9 | 8 |
|
| Benign
tumors | 3 | 2 | 1 |
| 2 | 1 |
|
| Other
diseases | 4 | 3 | 1 |
| 4 | 0 |
|
| Sex | 72 |
|
| 0.554a |
|
| 0.118a |
|
Female | 25 | 12 | 13 |
| 9 | 16 |
|
|
Male | 47 | 26 | 21 |
| 26 | 21 |
|
| Age, years | 70 |
|
| 0.863 |
|
| 0.863 |
|
<50 | 46 | 24 | 22 |
| 22 | 24 |
|
|
≥50 | 24 | 12 | 12 |
| 12 | 12 |
|
Discussion
Over the past few years, there have been increasing
evidences asserting the role of small non coding microRNA
genes in different cellular processes promoting cell
differentiation, proliferation, malignant transformation and
tumorigenesis (4,5). Among the huge number of
microRNA genes, the MIR34 genes have been extensively
focused on because they play a key role as tumor suppressors in
cancer (10,11). Currently, clinical trial on cancer
therapy based on MIR34a has already shown antitumor activity
in refractory advanced solid tumor (39). Therefore, investigating the
aberrant expression of the MIR34 family in cancer has been
being an attractive subject. The down regulation of all the three
members of the MIR34 family via promoter methylation, the
correlation between MIR34 methylation with cancer type,
grade, metastasis and survival, as well as the aberrant expression
of MIR34 targeted genes have been extensively reported in
multiple types of cancers including breast and lung cancers
(12,17,19,40).
However, an integrative comparison of MIR34 methylation
between both type of cancers vs. normal tissues adjacent to breast
cancer or non-cancerous pulmonary disease tissues has rarely been
described so far, while both types of these cancers are the first
and second common types of cancer (33). From a more general standpoint over
the literature, the difference in MIR34s methylation
frequency among non-cancerous, cancer and normal adjacent tissues
has been only explored in several types of cancer such as prostate,
colon, gastric and skin cancer (19,31,32).
In this study, we revealed that MIR34 methylation was
frequently found not only in breast cancer but also in normal
tissue adjacent to tumor, with the lowest frequency being around
30% (Table II). Interestingly,
MIR34a methylation occurred with significantly higher
frequency in breast cancer than in adjacent tissues (Table II), and showed a concordance with
MIR34b/c methylation only in breast cancer tissues (Table III). Concerning lung cancer, we
showed here that methylation of MIR34a and MIR34b/c
occurs in non-cancerous lung disease tissues with similar frequency
than in lung cancer (Table II),
suggesting their role in cancerous and non-cancerous lung disease
onset and progression. Indeed, it is worth noting that no
MIR34a methylation and a tiny frequency of MIR34b/c
methylation have been detected in normal tissue adjacent to tumor
of lung cancer (16,40,41).
In addition, the contribution of MIR methylation to the
pathogenesis of pulmonary fibrosis has been described previously
(42). In our study, the
concomitant methylation of MIR34a and MIR34b/c in
breast cancer, lung cancer and non-cancerous lung disease tissues
but not in normal tissue adjacent to breast cancer (Table II) emphasizes the role of
MIR34 methylation in human diseases including cancers.
Recently, Piletič and Kunej have reviewed that epigenetic
regulation of 63 MIR genes including MIR34s was
strongly correlated with 21 human diseases including 11 types of
cancers (43).
Interestingly, MIR34a methylation was
significantly correlated with tumor type IDC that consists of about
85% of all breast cancer types (Table III). Aberrant methylation of
MIR34a has been found to have significant relation with the
tumor grade from triple negative breast cancers or type II ovarian
cancer (17,44). However, we did not find any
association between MIR34a methylation and breast tumor
grade or lymph metastasis (Table
III), even if this latter has been frequently reported in
various cancer types such as colon, gastric and esophageal
carcinomas (22,31,45).
Similarly, there were no association of MIR34a methylation
with clinicopathological features of lung cancer as shown in our
study (Table III), as also shown
in a previous work from Wang and colleagues (23). These observations suggested that
the aberrant MIR34a methylation is preferentially associated
with the development of different types of cancer. This could be
supported by some other findings such as the strong association of
MIR34a methylation with p53 mutation in Li-Fraumeni
syndrome, a highly penetrant cancer predisposition syndrome while
on the contrary, no association with p53 mutation was found
in ovarian cancer (44,46).
On the contrary to MIR34a, MIR34b/c
methylation did not differ from breast cancer to adjacent tissues
as well as from lung cancer to non-cancerous tissues although it
occurred in all investigated tissues in this study. Furthermore, no
significant difference in MIR34b/c methylation was found
associated with clinicopathological features of neither breast nor
lung cancer. There were number of previous reports showing that
MIR34b/c methylation has a strong association with
histologic type, pathologic stage and distance metastasis of breast
and lung cancer (17,21,40,47).
However, it is worth noting that these conclusions have not always
been consistent, since other studies did not detect any correlation
of MIR34b/c methylation with tumor metastasis nor with
clinicopathological features in lung, gastric and colon cancers
(16,31,32).
The inconsistence of our result with previous studies may be
explained by the difference concerning the tumor stage of analyzed
samples. In our study, breast samples at grade 3 represented 12%
[while it was 32% in the study by Zeng et al (17)], and lung cancer samples at stage IV
represented 52% [while no sample of this stage was analyzed in the
study by Kim et al (40)].
In addition, there are limitations inherent to our study design
that should be noted. The statistical analysis was limited to small
samples. Moreover, the clinicopathological characteristics
concerning hormone phenotypes ER/PR/HER2 presented some missing
data. In our future studies, increasing number of fully
characterized samples will be analysed to determine the correlation
between MIR34 promoter methylation and subtypes of
cancers.
To summarize, this study has chosen the non
quantitative MSP method for the preliminary analysis of
MIR34 methylation, a method that has been widely used in
numerous studies (48) given its
simplicity, high sensitivity and low cost. We have shown that the
methylation frequently and concomitantly occurred at the promoters
of MIR34 gene family in breast, lung cancer and pulmonary
diseases. The encouraging results now prompt us to quantitatively
investigate the correlation between MIR34 promoter
methylation and the silencing of their expression, as well as with
the expression level of the mRNAs targeted by MIR34s. In
long term, this would allow optimizing detection techniques that
are suitable for moderately equipped laboratories in developing
countries, using MIR methylation markers in clinical
applications for diagnosis of human disease including cancers.
Acknowledgements
The authors would like to thank Doan H. Van, Nguyen
T. Duc and Thieu M. Thu at the Biomedical Lab, Faculty of Biology,
VNU University of Science, Hanoi, Vietnam.
Funding
This study was financially supported by the Ministry
of Science and Technology, Vietnam (grant no.
106-YS.06-2015.07).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available on reasonable request addressed to the
corresponding author.
Authors' contributions
VTTL conceived, planned the experiments and wrote
the manuscript. VLT contributed to the statistical analysis and
manuscript writing. PATD carried out the statistical analysis. HVS
and NLT contributed to sample preparation. NTT and NTP carried out
the experiments.
Ethics approval and consent to
participate
Informed consent for using tissue materials for
scientific purposes and publication was obtained from patients in
written form and the study was approved by the guidelines of the
VNU University of Science ethical committee in Vietnam (no.
9/2016/108/HDTN, VNU University of Science, Hanoi, Vietnam).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Heyn H and Esteller M: DNA methylation
profiling in the clinic: Applications and challenges. Nat Rev
Genet. 13:679–692. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hao X, Luo H, Krawczyk M, Wei W, Wang W,
Wang J, Flagg K, Hou J, Zhang H, Yi S, et al: DNA methylation
markers for diagnosis and prognosis of common cancers. Proc Natl
Acad Sci USA. 114:7414–7419. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Strmsek Z and Kunej T: MicroRNA silencing
by DNA methylation in human cancer: A literature analysis.
Non-Coding RNA. 1:44–52. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Suzuki H, Maruyama R, Yamamoto E and Kai
M: DNA methylation and microRNA dysregulation in cancer. Mol Oncol.
6:567–578. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kurozumi S, Yamaguchi Y, Kurosumi M, Ohira
M, Matsumoto H and Horiguchi J: Recent trends in microRNA research
into breast cancer with particular focus on the associations
between microRNAs and intrinsic subtypes. J Hum Genet. 62:15–24.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tang X, Tang J, Liu X, Zeng L, Cheng C,
Luo Y, Li L, Qin SL, Sang Y, Deng LM and Lv XB: Downregulation of
miR-129-2 by promoter hypermethylation regulates breast cancer cell
proliferation and apoptosis. Oncol Rep. 35:2963–2969. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li Q, Li H, Zhao X, Wang B, Zhang L, Zhang
C and Zhang F: DNA methylation mediated downregulation of miR-449c
controls osteosarcoma cell cycle progression by directly targeting
oncogene c-Myc. Int J Biol Sci. 13:1038–1050. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
He DX, Gu F, Gao F, Hao JJ, Gong D, Gu XT,
Mao AQ, Jin J, Fu L and Ma X: Genome-wide profiles of methylation,
microRNAs, and gene expression in chemoresistant breast cancer. Sci
Rep. 6:247062016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Su Y, Fang H and Jiang F: Integrating DNA
methylation and microRNA biomarkers in sputum for lung cancer
detection. Clin Epigenetics. 8:1092016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhu J, Zheng Z, Wang J, Sun J, Wang P,
Cheng X, Fu L, Zhang L, Wang Z and Li Z: Different miRNA expression
profiles between human breast cancer tumors and serum. Front Genet.
5:1492014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Agostini M and Knight RA: miR-34: From
bench to bedside. Oncotarget. 5:872–881. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Adams BD, Wali VB, Cheng CJ, Inukai S,
Booth CJ, Agarwal S, Rimm DL, Győrffy B, Santarpia L, Pusztai L, et
al: miR-34a silences c-SRC to attenuate tumor growth in
triple-negative breast cancer. Cancer Res. 76:927–939. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cortez MA, Ivan C, Valdecanas D, Wang X,
Peltier HJ, Ye Y, Araujo L, Carbone DP, Shilo K, Giri DK, et al:
PDL1 regulation by p53 via miR-34. J Natl Cancer Inst. 108:pii:
djv303. 2015.PubMed/NCBI
|
|
14
|
Stahlhut C and Slack FJ: Combinatorial
action of microRNAs let-7 and miR-34 effectively synergizes with
erlotinib to suppress non-small cell lung cancer cell
proliferation. Cell Cycle. 14:2171–2180. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Corney DC, Flesken-Nikitin A, Godwin AK,
Wang W and Nikitin AY: MicroRNA-34b and microRNA-34c are targets of
p53 and cooperate in control of cell proliferation and
adhesion-independent growth. Cancer Res. 67:8433–8438. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Daugaard I, Knudsen A, Kjeldsen TE, Hager
H and Hansen LL: The association between miR-34 dysregulation and
distant metastases formation in lung adenocarcinoma. Exp Mol
Pathol. 102:484–491. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zeng Z, Chen X, Zhu D, Luo Z and Yang M:
Low expression of circulating MicroRNA-34c is associated with poor
prognosis in triple-negative breast cancer. Yonsei Med J.
58:697–702. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Strmsek Z and Kunej T: Data integration of
104 studies related with microRNA epigenetics revealed that miR-34
gene family is silenced by DNA methylation in the highest number of
cancer types. Discov J. 2:e182014. View Article : Google Scholar
|
|
19
|
Lodygin D, Tarasov V, Epanchintsev A,
Berking C, Knyazeva T, Körner H, Knyazev P, Diebold J and Hermeking
H: Inactivation of miR-34a by aberrant CpG methylation in multiple
types of cancer. Cell Cycle. 7:2591–2600. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Vogt M, Munding J, Grüner M, Liffers ST,
Verdoodt B, Hauk J, Steinstraesser L, Tannapfel A and Hermeking H:
Frequent concomitant inactivation of miR-34a and miR-34b/c by CpG
methylation in colorectal, pancreatic, mammary, ovarian,
urothelial, and renal cell carcinomas and soft tissue sarcomas.
Virchows Arch. 458:313–322. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lujambio A, Calin GA, Villanueva A, Ropero
S, Sánchez-Céspedes M, Blanco D, Montuengaf LM, Rossic S, Nicolosoc
MS, Fallerg WJ, et al: A microRNA DNA methylation signature for
human cancer metastasis. Proc Natl Acad Sci USA. 105:13556–13561.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Siemens H, Neumann J, Jackstadt R,
Mansmann U, Horst D, Kirchner T and Hermeking H: Detection of
miR-34a promoter methylation in combination with elevated
expression of c-Met and β-catenin predicts distant metastasis of
colon cancer. Clin Cancer Res. 19:710–720. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Z, Chen Z, Gao Z, Li N, Li B, Tan F,
Tan X, Lu N, Sun Y, Sun J, et al: DNA hypermethylation of
microRNA-34b/c has prognostic value for stage I non-small cell lung
cancer. Cancer Biol Ther. 11:490–496. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Casadio V, Molinari C, Calistri D, Tebaldi
M, Gunelli R, Serra L, Falcini F, Zingaretti C, Silvestrini R,
Amadori D and Zoli W: DNA methylation profiles as predictors of
recurrence in non muscle invasive bladder cancer: An MS-MLPA
approach. J Exp Clin Cancer Res. 32:942013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Troester MA, Hoadley KA, D'Arcy M,
Cherniack AD, Stewart C, Koboldt DC, Robertson AG, Mahurkar S, Shen
H, Wilkerson MD, et al: DNA defects, epigenetics, and gene
expression in cancer-adjacent breast: A study from The Cancer
Genome Atlas. NPJ Breast Cancer. 2:160072016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cao B, Feng L, Lu D, Liu Y, Liu Y, Guo S,
Han N, Liu X, Mao Y, He J, et al: Prognostic value of molecular
events from negative surgical margin of non-small-cell lung cancer.
Oncotarget. 8:53642–53653. 2016.PubMed/NCBI
|
|
27
|
Yan PS, Venkataramu C, Ibrahim A, Liu JC,
Shen RZ, Diaz NM, Centeno B, Weber F, Leu YW, Shapiro CL, et al:
Mapping geographic zones of cancer risk with epigenetic biomarkers
in normal breast tissue. Clin Cancer Res. 12:6626–6636. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Teschendorff AE, Gao Y, Jones A, Ruebner
M, Beckmann MW, Wachter DL, Fasching PA and Widschwendter M: DNA
methylation outliers in normal breast tissue identify field defects
that are enriched in cancer. Nat Commun. 7:104782016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Johnson KC, Koestler DC, Fleischer T, Chen
P, Jenson EG, Marotti JD, Onega T, Kristensen VN and Christensen
BC: DNA methylation in ductal carcinoma in situ related with future
development of invasive breast cancer. Clin Epigenetics. 7:752015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schrijver WA, Jiwa LS, van Diest PJ and
Moelans CB: Promoter hypermethylation profiling of distant breast
cancer metastases. Breast Cancer Res Treat. 151:41–55. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wu XD, Song YC, Cao PL, Zhang H, Guo Q,
Yan R, Diao DM, Cheng Y and Dang CX: Detection of miR-34a and
miR-34b/c in stool sample as potential screening biomarkers for
noninvasive diagnosis of colorectal cancer. Med Oncol. 31:8942014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Suzuki H, Yamamoto E, Nojima M, Kai M,
Yamano HO, Yoshikawa K, Kimura T, Kudo T, Harada E, Sugai T, et al:
Methylation-associated silencing of microRNA-34b/c in gastric
cancer and its involvement in an epigenetic field defect.
Carcinogenesis. 31:2066–2073. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Clark SJ, Harrison J, Paul CL and Frommer
M: High sensitivity mapping of methylated cytosines. Nucleic Acids
Res. 22:2990–2997. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lan VT, Ha NT, Uyen NQ, Duong NT, Huong
NT, Thuan TB, Duong PA and To TV: Standardization of the
methylation-specific PCR method for analyzing BRCA1 and ER
methylation. Mol Med Rep. 9:1844–1850. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Herman JG, Graff JR, Myöhänen S, Nelkin BD
and Baylin SB: Methylation-specific PCR: A novel PCR assay for
methylation status of CpG islands. Proc Natl Acad Sci USA.
93:9821–9826. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tarasov V, Jung P, Verdoodt B, Lodygin D,
Epanchintsev A, Menssen A, Meister G and Hermeking H: Differential
regulation of microRNAs by p53 revealed by massively parallel
sequencing: miR-34a is a p53 target that induces apoptosis and
G1-arrest. Cell Cycle. 6:1586–1593. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Toyota M, Suzuki H, Sasaki Y, Maruyama R,
Imai K, Shinomura Y and Tokino T: Epigenetic silencing of
microRNA-34b/c and B-Cell translocation gene 4 is associated with
CpG Island methylation in colorectal cancer. Cancer Res.
68:4123–4132. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Beg MS, Brenner AJ, Sachdev J, Borad M,
Kang YK, Stoudemire J, Smith S, Bader AG, Kim S and Hong DS: Phase
I study of MRX34, a liposomal miR-34a mimic, administered twice
weekly in patients with advanced solid tumors. Invest New Drugs.
35:180–188. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kim YH, Lee WK, Lee EB, Son JW, Kim DS and
Park JY: Combined effect of metastasis-related microRNA, miR-34 and
miR-124 family, methylation on prognosis of Non-Small-Cell lung
cancer. Clin Lung Cancer. 18:e13–e20. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tan W, Gu J, Huang M, Wu X and Hildebrandt
MA: Epigenetic analysis of microRNA genes in tumors from surgically
resected lung cancer patients and association with survival. Mol
Carcinog. 54 Suppl 1:E45–E51. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Dakhlallah D, Batte K, Wang Y,
Cantemir-Stone CZ, Yan P, Nuovo G, Mikhail A, Hitchcock CL, Wright
VP, Nana-Sinkam SP, et al: Epigenetic regulation of miR-17 ~92
contributes to the pathogenesis of pulmonary fibrosis. Am J Respir
Crit Care Med. 187:397–405. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Piletič K and Kunej T: MicroRNA epigenetic
signatures in human disease. Arch Toxicol. 90:2405–2419. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Schmid G, Notaro S, Reimer D, Abdel-Azim
S, Duggan-Peer M, Holly J, Fiegl H, Rössler J, Wiedemair A, Concin
N, et al: Expression and promotor hypermethylation of miR-34a in
the various histological subtypes of ovarian cancer. BMC Cancer.
16:1022016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Cui X, Zhao Z, Liu D, Guo T, Li S, Hu J,
Liu C, Yang L, Cao Y, Jiang J, et al: Inactivation of miR-34a by
aberrant CpG methylation in Kazakh patients with esophageal
carcinoma. J Exp Clin Cancer Res. 33:202014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Samuel N, Wilson G, Lemire M, Said Id B,
Lou Y, Li W, Merino D, Novokmet A, Tran J, Nichols NE, et al:
Genome-wide DNA methylation analysis reveals epigenetic
dysregulation of microRNA-34A in TP53-associated cancer
susceptibility. J Clin Oncol: pii: JCO676940. 2016. View Article : Google Scholar
|
|
47
|
Tanaka N, Toyooka S, Soh J, Kubo T,
Yamamoto H, Maki Y, Muraoka T, Shien K, Furukawa M, Ueno T, et al:
Frequent methylation and oncogenic role of microRNA-34b/c in
small-cell lung cancer. Lung Cancer. 76:32–38. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kristensen LS and Hansen LL: PCR-based
methods for detecting single-locus DNA methylation biomarkers in
cancer diagnostics, prognostics, and response to treatment. Clin
Chem. 55:1471–1483. 2009. View Article : Google Scholar : PubMed/NCBI
|