Introduction
Non-alcoholic fatty liver disease (NAFLD) is a
pathological syndrome characterized by fatty liver unassociated
with alcohol consumption. NAFLD unassociated with genetic
and-environment-associated metabolic stress diseases, including
simple non-alcoholic fatty liver (NAFL), non-alcoholic
steatohepatitis (NASH) and liver cirrhosis. NAFLD is often
associated with obesity, hyperglycemia, hyperlipidemia,
hyperinsulinemia, hypertension and atherosclerosis (1).
With lifestyle changes in modern society, the
incidence of NAFLD increases each year and shows high prevalence at
young age (2). The prevalence of
NAFLD in Western countries is 20–30% (3) whereas in the Asia Pacific region, the
prevalence is up to 24% (4).
Italian 10-year cohort study showed that the prevalence of NAFLD
was increased by 2% each year (5).
In China, NAFLD affects over a quarter of the general population,
and its prevalence is rapidly increasing with lifestyle changes and
the aging population (6,7).
In spite of this, the pathogenesis of NAFLD has not
yet been fully elucidated (8). In
1998, Day and James (9) proposed a
hypothesis called ‘secondary strike’ for the pathogenesis of NAFLD.
According to the hypothesis, the strike is primarily a reactive
product increased of oxidative metabolism, leading to lipid
peroxidation with cytokines, and mitochondrial uncoupling protein.
Their study found that abnormal metabolism of fatty acids leads to
generation of reactive oxygen species (ROS) (9). These functional abnormalities may
lead to endoplasmic reticulum stress (ERS). It has been known that
the endoplasmic reticulum plays an important role in both fatty
acid synthesis and cholesterol metabolism (10). In addition, the endoplasmic
reticulum is the site of triglyceride formation (11). The potential role of the
endoplasmic reticulum in the regulation of lipid droplet number,
composition, size and in lipogenesis and lipolysis remains an
important but unexplored area (12). When the endoplasmic reticulum
becomes stressed due to the accumulation of newly synthesized
unfolded proteins, stress signaling systems are activated,
including the c-Jun-NH2 terminal kinase (JNK) pathways, as well as
networks activated by oxidative stress, which can influence lipid
metabolism. Previous studies have demonstrated that disturbed
endoplasmic reticulum homeostasis-induced apoptosis (13) and lipid accumulation (14) in the liver may contribute to the
development of NAFLD (15).
Despite this, there is a lack of ideal experimental
animal models for NAFLD, which limits research into its etiology,
pathogenesis, and prevention. The current ob/ob/db/db mouse is the
most used NAFLD model for research. The ob/ob mouse is genetically
deficient in the leptin gene, while the db/db mouse has a natural
mutation in the leptin receptor gene. Normal feed can cause fatty
liver, but not in the form of NASH (16,17).
Congenital and genetically modified models are difficult to obtain
and expensive to purchase. Lack of methionine and choline diet in
rats can cause NAFLD, but these diet feeding reduced muscle and fat
with body weight loss, which is different from human NAFLD
(18,19). NAFLD rat models induced by CCl4
suffer from high mortality, while this model lacks obesity and IR.
Compared with mice and rats, rabbits have unique features of lipid
metabolism similar to those of humans (20,21).
Unlike mice in which high density lipoproteins (HDLs) is the
predominant plasma lipoproteins, rabbit lipoprotein profiles are
LDL-rich, which is similar to those of humans. Secondly, it has
been reported that cholesteryl ester transfer protein (CETP) plays
a central role in lipid metabolism and the atherosclerotic process.
Rabbits have abundant CETP activity in the plasma as do humans,
whereas mice do not have an endogenous CETP gene. In addition, like
humans, rabbits do not have hepatic apoB mRNA editing activity, so
rabbit apoB-48 is only present in chylomicrons. However, apoB-48 is
present in all apoB-containing particles such as very low-density
lipoproteins (VLDL), LDLs and chylomicrons in mice. Therefore,
rabbits provide a unique system to study lipid metabolism. In the
present study, we attempted to generate a rabbit NAFLD model by
feeding with a diet containing high cholesterol or high cholesterol
with high fat to mimic human NAFLD.
Materials and methods
Animals and diets
Japanese white rabbits were provided by the
Laboratory Animal Center of Xi'an Jiaotong University. A total of
20 male rabbits (16 weeks old, about 2.0 kg body weight) were
randomly divided into three groups: The control group (n=4) was fed
with a chow diet, HCD group (n=8) was fed with a chow diet
supplemented with 1% cholesterol and high fat and high cholesterol
diet (HFCD) group (n=8) was fed a chow diet containing 6.7% lard
and 1% cholesterol for 12 weeks. Rabbits were fed with a restricted
diet (100 g/day/animal) with free access to water.
Cholesterol was purchased from Wako Pure Chemical
Industries, Ltd., (Osaka, Japan). The experimental protocols were
approved by the Animal Administration Committee of Xi'an Medical
University (Shaanxi, China) and performed according to the Xi'an
Medical University Guidelines for Animal Experimentation and the
Guide for the Care and Use of Laboratory Animals Published by the
US National Institutes of Health (NIH publication no. 85-23,
revised 1996).
Determination of plasma lipid levels
and glucose tolerance test
Rabbits were fasted for 16 h prior to blood
collection. Blood samples were collected via the ear artery and put
into tubes containing EDTA and the plasma separated by
centrifugation at 2,000 rpm/min (20 min, 4°C). Plasma total
cholesterol (TC) and triglyceride (TG) levels were measured
biweekly using commercial assay kits (BioSino Bio-technology and
Science, Inc., Beijing, China). At 12 weeks, blood samples were
collected to measure plasma free fatty acid (FFA) levels using
commercial assay kits (BioVision, Inc., Milpitas, CA, USA). At the
same time, plasma alanine aminotransferase (ALT) and aspartate
aminotransferase (AST) were measured using commercial assay kits
(Pars Azmun, Tehran, Iran).
To evaluate the effect of high cholesterol diet
(HCD) on glucose metabolism, rabbits were fasted overnight and an
intravenous glucose tolerance test (IVGTT) was performed using the
method as described previously (22). A bolus of glucose (0.6 g/kg body
weight) was injected through the ear vein and blood samples were
collected through the ear artery at 5, 10, 15, 20, 30, 45, 60, 75
and 120 min. Plasma glucose levels were measured using commercial
assay kits (Wako Pure Chemical Industries, Ltd., Osaka, Japan). The
incremental area under the curve (AUC) was calculated according to
the trapezium rule as described previously (23).
Analysis of lipid metabolism and
ERS-related gene expression of the liver
At the end of the experiment, the rabbits were
euthanized by overdose of injection of sodium barbital solution. A
piece of liver was quickly put into liquid nitrogen for homogenate
and RNA extraction. About 100 mg liver tissue was homogenized in
0.75 ml of methanol and chloroform (2:1), and lipids were extracted
from the chloroform fraction and N2-dried to remove the chloroform.
Then, the hepatic TC and TG contents were determined using Wako
kits (Wako Pure Chemical Industries, Ltd.). For the hepatic FFA
assay, liver tissue was homogenized in 0.2 ml of chloroform with 1%
Triton X-100. Fatty acids were extracted in the chloroform fraction
and N2-dried to remove the chloroform. Then, the hepatic tissue FFA
contents were determined using a FFA quantification kit (BioVision,
Mountain View, CA, USA) (24). For
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) reactions, total RNA was extracted from liver samples
using TRIzol Plus (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) according to manufacturer's instructions. A total
of 10 µg total RNA was reverse transcribed into cDNA using Takara
SuperScript™ RT reagent kit with gDNA eraser (cat. no. RR047A;
Takara Biotechnology Co., Ltd., Dalian, China). A mixture of dNTPs
were included in the PrimeScript RT enzyme Mix, and OligodT primers
and random 6 mers were included in the RT primer mix of the
SuperScript™ RT reagent kit. ERS-associated gene expression was
measured by RT-qPCR. Forward and reverse primer sequences of
ERS-associated gene are presented in Table II. GAPDH was used as the reference
gene and the primer sequences are included in Table II. The thermocycling conditions
used for qPCR were as follows: 95°C for 30 sec, followed by 40
cycles of 95°C for 5 sec and 60°C for 30 sec, and then, 95°C for 15
sec, 60°C for 30 sec and 95°C for 15 sec. RT-qPCR analysis was
performed using Takara SYBR Green kit (cat. no. DRR820A; Takara
Biotechnology Co., Ltd.) by TaKaRa TP800 (Takara Bio, Inc., Otsu,
Japan). Values of the quantification cycle (Cq) were used for
quantification with the 2−ΔΔCq method, as described
previously (25).
 | Table II.Primer sequences for reverse
transcription-quantitative polymerase chain reaction. |
Table II.
Primer sequences for reverse
transcription-quantitative polymerase chain reaction.
| Genes | Primer sequence
(5′-3′) |
|---|
| GRP78 | F:
TGGCTACTCCTCTCTGAACAAC |
| GRP78 | R:
CGAGAATCATGGCGGAGATCT |
| JNK | F:
CACCACCAAAGATTCCTGACA |
| JNK | R:
GGCTGCCCTCGTATAACTCC |
| CHOP | F:
AGAACTGGGACGAAGAGACC |
| CHOP | R:
AGCTAGCTGTGCCACTTTCC |
| Caspase-12 | F:
GGAAGGTAGGCAAGACTGGTT |
| Caspase-12 | R:
TCAATAGTGGGCATCTGGGTC |
| GAPDH | F:
CCACTTTGTGAAGCTCATTTCCT |
| GAPDH | R:
TCGTCCTCCTCTGGTGCTCT |
Western blotting analysis
Fresh liver samples were homogenized in lysis buffer
at 4°C followed by centrifugation at 10,000 rpm at 4°C for 10 min.
The resultant supernatants were collected and subjected to western
blotting. Briefly, 15 µg lysates were fractionated on 10%
SDS-polyacrylamide gels and then transferred to Sequi-Blot
polyvinylidene fluoride membranes (Bio-Rad, Hercules, CA, USA). The
membranes were incubated with each primary antibody (Ab) (1:1,000)
at 4°C overnight, as recommended in the manufacturer's
instructions. After washing 3 times, they were incubated with
horseradish peroxidase conjugated secondary Ab for 2 h. Signals
were detected using the Immobilon reagent (Millipore, Billerica,
MA, USA) and visualized using an LAS-400 Lumino Image Analyzer
(Fujifilm, Co., Tokyo, Japan) (26). Primary Abs of CHOP (L63F7), JNK
(9251L) and Caspase-12 (2202P) were purchased from Cell Signaling
Technology, Inc., (Danvers, MA, USA). Anti-GRP78 (ab89789) was
purchased from Abcam (Cambridge, UK).
Pathological analysis
At the end of the experiment, a piece of liver was
stocked in liquid nitrogen and embedded in optimal cutting
temperature (OCT) compound and cut in 8-µm sections followed by oil
red O staining. The areas stained by oil red O were quantified to
assess the areas of connective tissue and lipid deposition,
respectively, using an image analysis system (WinROOF v6.5)
(24,27). At the same time, formalin-fixed
liver was embedded in paraffin and cut in 4-µm thick sections and
stained with hematoxylin and eosin (HE).
Statistical analysis
Data are expressed as the mean ± standard error of
the mean. Statistical analysis was performed using analysis of
variance and Dunnett's post hoc test, and the data with an equal F
value, using GraphPad Prism 5 software (GraphPad Software, Inc., La
Jolla, CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Plasma lipid levels, liver function
and glucose tolerance test
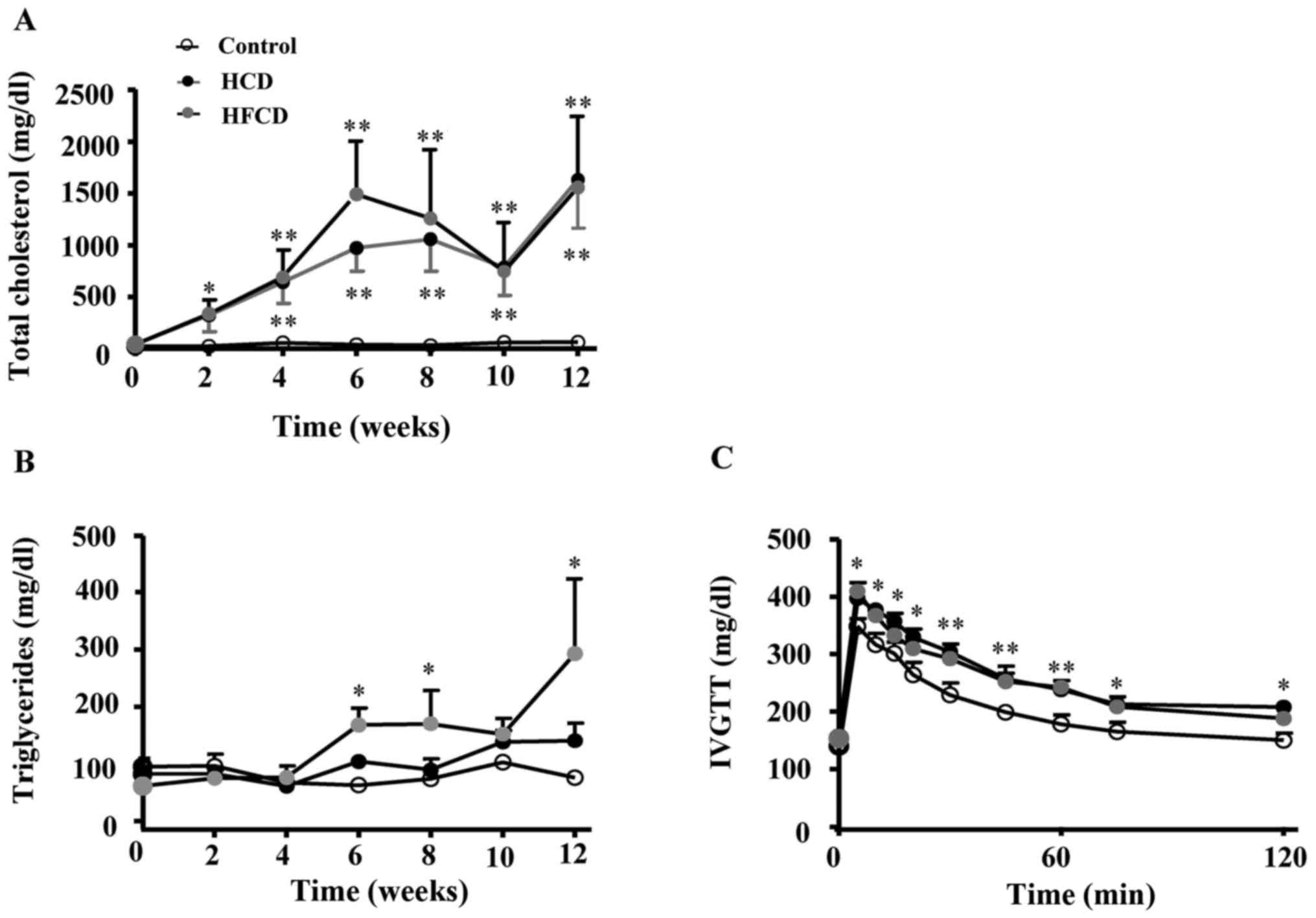
Plasma TC levels were remarkably increased in both
HCD and HFCD groups compared with control group (P<0.05 or
P<0.01). Plasma TG significantly increased in HFCD group
compared with control group (P<0.05). At the end of the
experiment, plasma FFA was measured and the results showed that FFA
was significantly increased in HCD and HFCD groups compared with
control group (P<0.05). We also performed a glucose tolerance
test and found that glucose clearance was delayed in HCD and HFCD
groups compared with control group (P<0.05 or P<0.01;
Fig. 1). Plasma ALT and AST levels
were not remarkably different among the three groups (Table I).
| Table I.Test results of liver function. |
Table I.
Test results of liver function.
| Group | ALT (U/l) | AST (U/l) |
|---|
| Control | 43±10 | 40±12 |
| HCD | 46±11 | 81±13 |
| HFCD | 42±7 | 78±9 |
Pathological analysis of liver
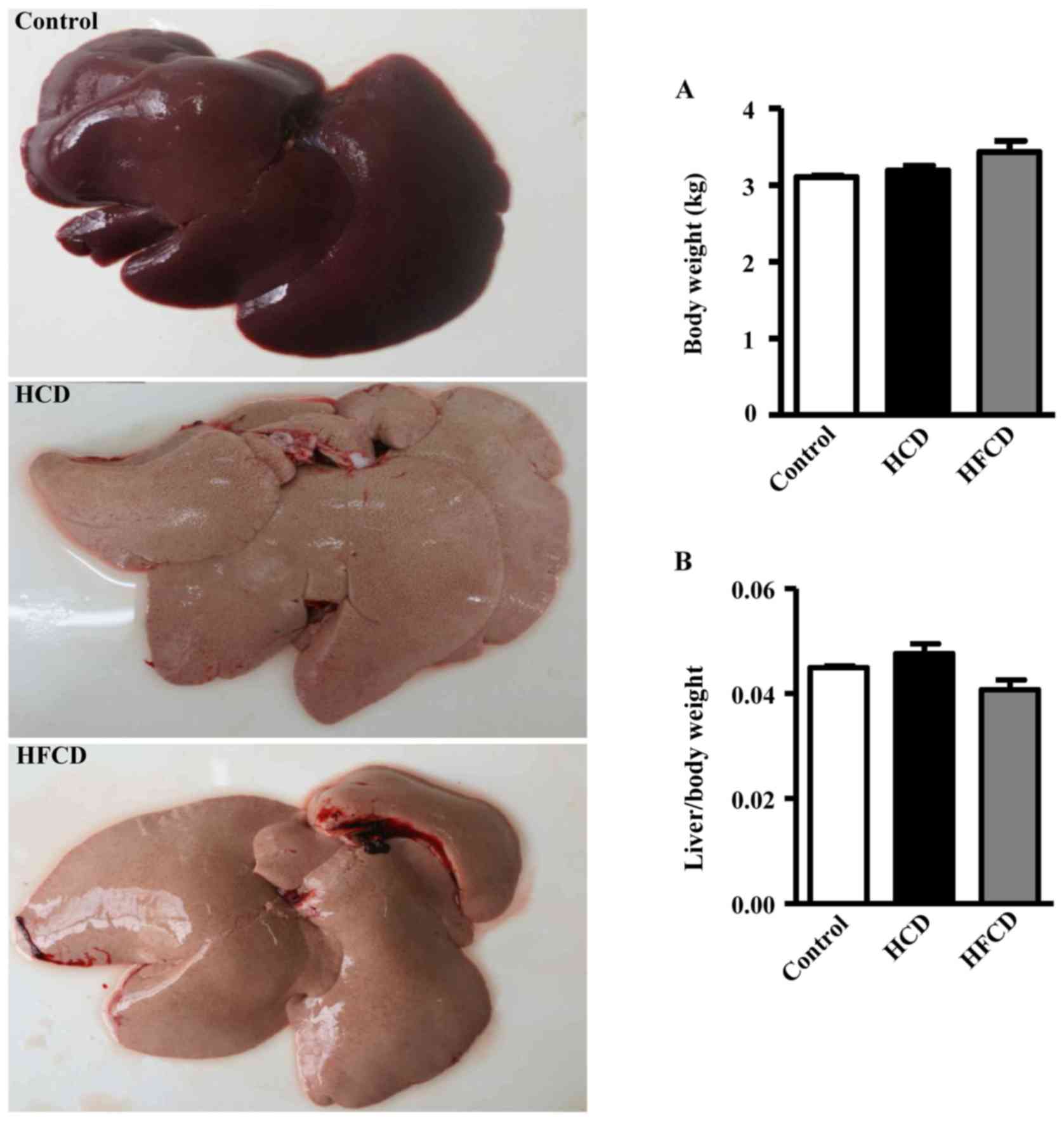
Compared with the liver of the control group, the
surface of the liver of HCD and HFCD groups appeared obviously
yellow and greasy (Fig. 2, left).
Body weight and the ratio of the liver weight to body weight were
not statistically significant in the three groups (Fig. 2A and B).
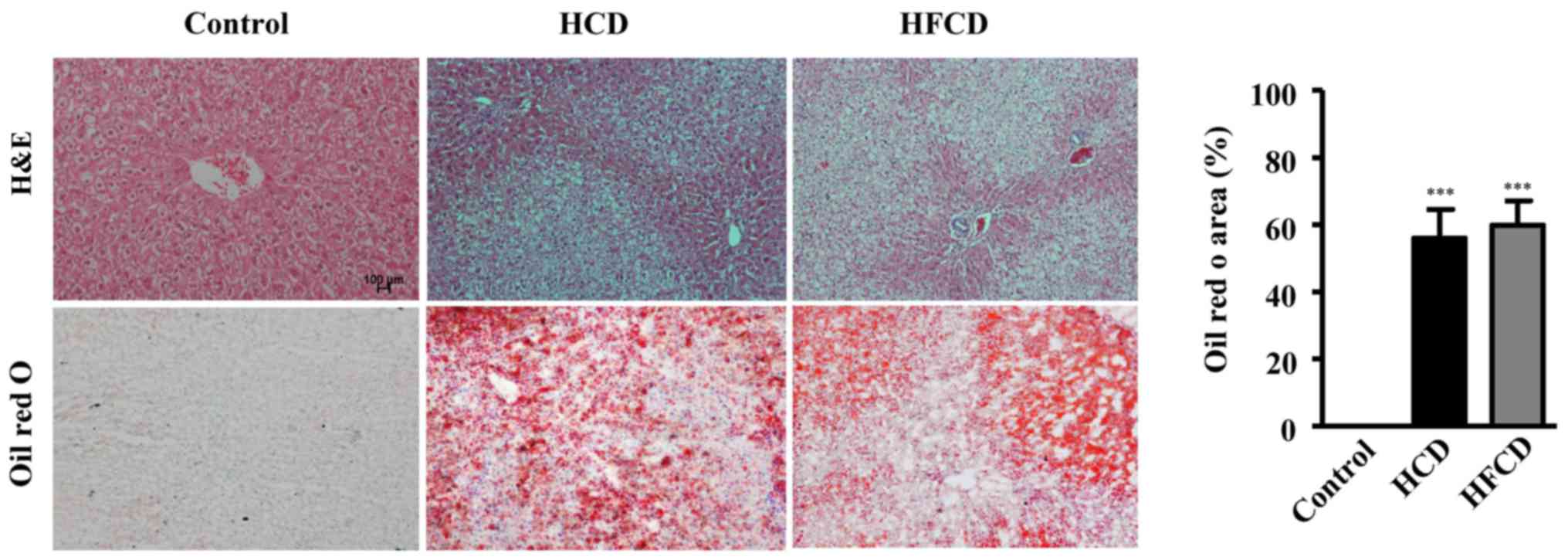
The analysis of liver histology revealed that there
were a large number of vacuoles in the hepatocytes of the HCD and
HFCD groups compared with control group as demonstrated by both HE
and oil red O staining (Fig. 3).
The areas stained by oil red O were measured to evaluate the
severity of connective tissue and lipid deposition as previously
described (24,26) and the results showed that NAFLD
model was successfully established in both HCD and HFCD groups even
though liver histological features were not significantly different
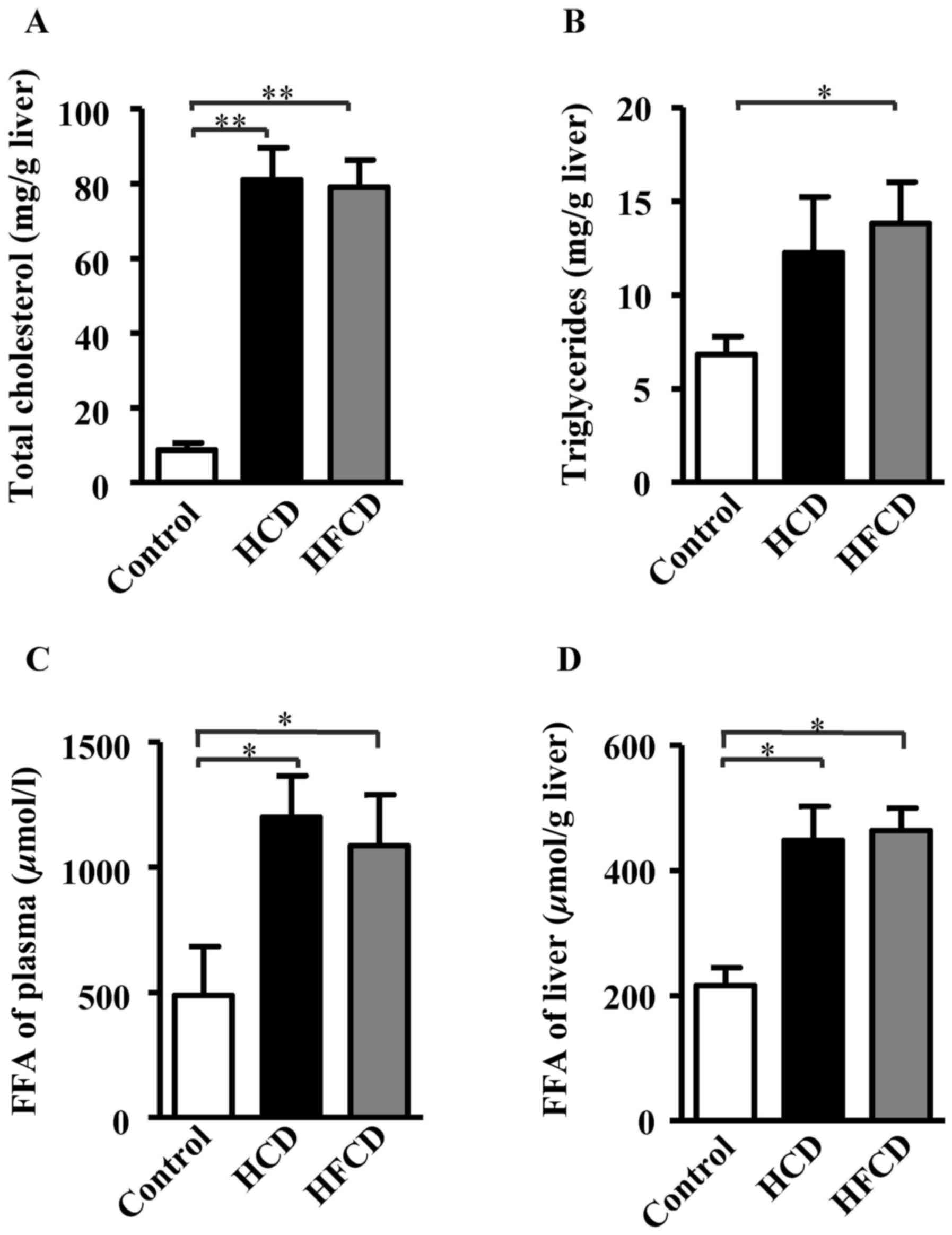
(Fig. 3). To evaluate lipid
compositions of liver, TC, TG and FFA contents were measured using
liver homogenate. The results showed that hepatic TC and FFA
contents were significantly increased in HCD and HFCD groups
compared with control group, and hepatic TG content was
significantly increased in HFCD group compared with control group.
However significant differences were not observed between the HCD
and HFCD groups (P<0.05 or P<0.01; Fig. 4).
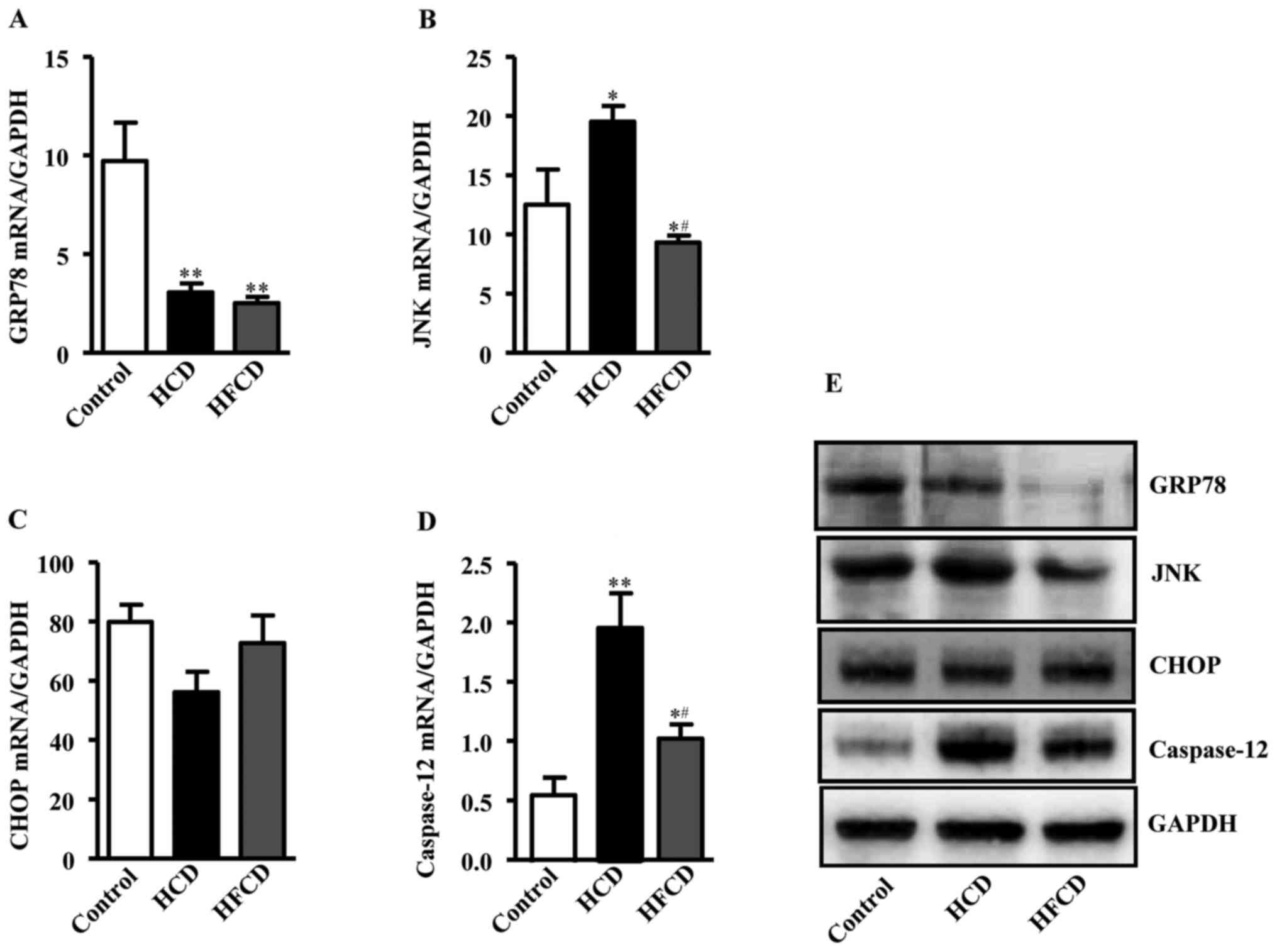
ERS indicators
Expression levels of ERS-associated liver genes were
compared by RT-qPCR to investigate whether ERS plays any role in
the pathogenesis of NAFLD. The genes and primer sequence were
showed in Table II. Glucose
regulation protein 78 (GRP78) mRNA expression was significantly
decreased in the HCD and HFCD groups compared with the control
group. JNK mRNA expression was increased in HCD group compared with
control and HFCD group, while there was no significant difference
between HFCD and control groups. Caspase-12 mRNA expression was
also increased in the HCD and HFCD groups compared with control,
and there was a significant difference between the HCD and HFCD
groups (P<0.05 or P<0.01). CCAAT/enhancer-binding protein
homologous protein (CHOP) mRNA expression were not statistically
significant in the three groups. The protein expression levels of
the ERS indicators were measured and the results exhibited a
similar trend to the mRNA expression levels (Fig. 5).
Discussion
At present, NAFLD has become a serious challenges
for human health. Lack of an ideal animal models hampers the study
of NAFLD mechanisms and development of therapeutics. Based on the
advantages of rabbit lipid metabolism features being similar to
humans, this experiment established the NAFLD model using
cholesterol-fed JW rabbits. Two types of diets were used and both
HCD and HFCD could induce NAFLD features in rabbits. The analysis
of liver histology showed that HCD and HFCD groups were not
significantly different in terms of hepatic steatosis. This result
suggested that HCD is the major factor for inducing NAFLD in rabbit
model, and a high fat in the diet is unable to accelerate the
formation of NAFLD in a rabbit model. Therefore, high cholesterol
is a key factor in establishing NAFLD in rabbits.
TC, TG and FFA levels of liver tissue homogenate
were significantly increased in HCD and HFCD groups compared with
the control group. This result revealed that there was accumulation
of excessive cholesterol and other lipids in the liver compatible
with the observation that a large number of vacuoles or oil red O
stained lipid droplets were found in the livers of HCD and HFCD
groups even though no difference was observed between the HCD and
HFCD groups.
The endoplasmic reticulum plays an important role in
lipid metabolism. When a large amount of lipid accumulates in the
endoplasmic reticulum, stress signaling systems may be activated
thus influencing lipid metabolism. Previous studies have found that
fatty acid metabolism in the endoplasmic reticulum will weaken
cellular autophagy functions (28), producing a certain amount of ROS
(9). In the early stage of ERS,
expression of endoplasmic reticulum molecular chaperone protein and
GRP78 will increase and assist cell deal with unfolded protein.
However, prolonged ERS can cause apoptosis (5), the cell apoptosis process was
mediated by ROS and ERS (9). The
cell apoptosis caused by ERS is called the endoplasmic reticulum
associated death (ERAD) pathway. The ERAD pathway consists of ERS
induced CHOP which activates the JNK pathway and/or caspase-12
proteolytic enzyme (29). In our
study, we found that GRP78 mRNA expression was significantly
decreased in both HCD and HFCD groups. Prolonged ERS induced by
lipid accumulation can cause hepatocyte apoptosis. Therefore, with
the increase in cell death, GRP78 expression may be decreased
accordingly. JNK and caspase-12 expression levels were notably
increased in the HCD group. Caspase-12 expression was notably
increased in the HFCD group while CHOP expression differences were
not statistically significant among the three groups. In the
current study, the results suggested that the cell apoptosis may
have been caused by the activation of the JNK and caspase-12
pathways rather than the CHOP pathway, which may determine the
severity of hepatic steatosis and apoptosis. These results suggest
that accumulation of lipids in hepatocytes may lead to ERS by which
hepatocyte apoptosis may be enhanced although this hypothesis
remains to be verified in the future.
Acknowledgements
Not applicable.
Funding
The present study was supported in part by a
research grant from Shaanxi Science and Technology Hall Fund
Project of China (grant nos. 2016SF-107, 2017JZ028 and 17JS116),
Research Initiation Funds for Doctors of Xi'an Medical University
(grant no. 2016DOC25) and the National Natural Science Foundation
of China (grant nos. 81770457 and 81773795).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YW and PZ performed the administration and feed of
experimental animals, and analyzed plasma lipids levels and the
gene expression of ERS indicators in liver. XS performed the
glucose tolerance test and analyzed the data. QY performed the
histological examination of the liver and analyzed the data. YC and
HG analyzed the protein expression of ERS indicators in the liver
and revised the figures. EL contributed to the experimental design
and drafted the manuscript. JF was a major contributor in designing
the experiment, revising the manuscript critically for important
intellectual content and gave final approval of the version to be
published. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The experimental protocols were approved by the
Animal Administration Committee of Xi'an Medical University
(Shaanxi, China) and performed according to the Xi'an Medical
University Guidelines for Animal Experimentation and the Guide for
the Care and Use of Laboratory Animals Published by the US National
Institutes of Health (NIH Publication no. 85-23, revised 1996).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
DeFilippis AP, Blaha MJ, Martin SS, Reed
RM, Jones SR, Nasir K, Blumenthal RS and Budoff MJ: Nonalcoholic
fatty liver disease and serum lipoproteins: The multi-ethnic study
of atherosclerosis. Atherosclerosis. 227:429–436. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kwak JH, Jun DW, Lee SM, Cho YK, Lee KN,
Lee HL, Lee OY, Choi HS and Yoon BC: Lifestyle predictors of obese
and non-obese patients with nonalcoholic fatty liver disease: A
cross-sectional study. Clin Nutr pii. S0261-5614:303012017.
|
|
3
|
Sayiner M, Koenig A, Henry L and Younossi
ZM: Epidemiology of nonalcoholic fatty liver disease and
nonalcoholic steatohepatitis in the United States and the rest of
the world. Clin Liver Dis. 20:205–214. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chitturi S, Farrell GC, Hashimoto E,
Saibara T, Lau GK and Sollano JD: Asia-pacific working party on
NAFLD: Non-alcoholic fatty liver disease in the Asia-Pacific
region: Definitions and overview of proposed guidelines. J
Gastroenterol Hepatol. 22:778–787. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bedogni G, Miglioli L, Masutti F,
Tiribelli C, Marchesini G and Bellentani S: Prevalence of and risk
factors for nonalcoholic fatty liver disease: The Dionysos
nutrition and liver study. Hepatology. 42:44–52. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dai H, Wang W, Chen R, Chen Z, Lu Y and
Yuan H: Lipid accumulation product is a powerful tool to predict
non-alcoholic fatty liver disease in Chinese adults. Nutr Metab
(Lond). 14:492017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li Z, Xue J, Chen P, Chen L, Yan S and Liu
L: Prevalence of nonalcoholic fatty liver disease in mainland of
China: A meta-analysis of published studies. J Gastroenterol
Hepatol. 29:42–51. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cheung O and Sanyal AJ: Recent advances in
nonalcoholic fatty liver disease. Curr Opin Gastroenterol.
25:230–237. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Day C and James O: Steatohepatitis: A tale
of two ‘hits’? Gastroenterology. 114:842–845. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hotamisligil GS: Endoplasmic reticulum
stress and the inflammatory basis of metabolic disease. Cell.
140:900–917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wolins NE, Brasaemle DL and Bickel PE: A
proposed model of fat packaging by exchangeable lipid droplet
proteins. FEBS Lett. 580:5484–5491. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gregor MG and Hotamisligil GS: Thematic
review series: Adipocyte Biology. Adipocyte stress: The endoplasmic
reticulum and metabolic disease. J Lipid Res. 48:1905–1914. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wei Y, Wang D, Topczewski F and
Pagliassotti MJ: Saturated fatty acids induce endoplasmic reticulum
stress and apoptosis independently of ceramide in liver cells. Am J
Physiol Endocrinol Metab. 291:E275–E281. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Werstuck GH, Lentz SR, Dayal S, Hossain
GS, Sood SK, Shi YY, Zhou J, Maeda N, Krisans SK, Malinow MR and
Austin RC: Homocysteine-induced endoplasmic reticulum stress causes
dysregulation of the cholesterol and triglyceride biosynthetic
pathways. J Clin Invest. 107:1263–1273. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jung TW, Kim HC, El-Aty Abd AM and Jeong
JH: Maresin 1 attenuates NAFLD by suppression of endoplasmic
reticulum stress via AMPK-SERCA2b pathway. J Biol Chem.
293:3981–3988. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Catmiel-Haggai M, Cederbaum AI and Nieto
N: A high-fat diet leads to the progression of non-alcoholic fatty
liver disease in obese rats. FASEB J. 19:136–138. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wortham M, He L, Gyamfi M, Copple BL and
Wan YJ: The transition from fatty liver to NASH associates with
SAMe depletion in db/db mice fed a methionine choline-deficient
diet. Dig Dis Sci. 53:2761–2774. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Diehl AM: Lessons from animal models of
NASH. Hepatol Res. 33:138–144. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
London RM and Geroge J: Pathogenesis of
NASH: Animal models. Clin Liver Dis. 11(55–74): viii2007.
|
|
20
|
Fan J, Kitajima S, Watanabe T, Xu J, Zhang
J, Liu E and Chen YE: Rabbit models for the study of human
atherosclerosis: From pathophysiological mechanisms to
translational medicine. Pharmacol Ther. 146:104–119. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fan J and Watanabe T: Transgenic rabbits
as therapeutic protein bioreactors and human disease models.
Pharmacol Ther. 99:261–282. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhao S, Zhang C, Lin Y, Yang P, Yu Q, Chu
Y, Yang P, Fan J and Liu E: The effects of rosiglitazone on aortic
atherosclerosis of cholesterol-fed rabbits. Thromb Res.
123:281–287. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu E, Kitajima S, Higaki Y, Morimoto M,
Sun H, Watanabe T, Yamada N and Fan J: High lipoprotein lipase
activity increases insulin sensitivity in transgenic rabbits.
Metabolism. 54:132–138. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ogawa T, Fujii H, Yoshizato K and Kawada
N: A human-type nonalcoholic steatohepatitis model with advanced
fibrosis in rabbits. Am J Pathol. 177:153–165. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guan H, Lin Y, Bai L, An Y, Shang J, Wang
Z, Zhao S, Fan J and Liu E: Dietary cocoa powder improves
hyperlipidemia and reduces atherosclerosis in apoE deficient mice
through the inhibition of hepatic endoplasmic reticulum stress.
Mediators Inflamm. 2016:19375722016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hübscher SG: Histological assessment of
non-alcoholic fatty liver disease. Histopathology. 49:450–465.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Deng X, Pan X, Cheng C, Liu B, Zhang H,
Zhang Y and Xu K: Regulation of SREBP-2 intracellular trafficking
improves impaired autophagic flux and alleviates endoplasmic
reticulum stress in NAFLD. Biochim Biophys Acta. 1862:337–350.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhao L and Ackerman SL: Endoplasmic
reticulum stress in health and disease. Curr Opin Cell Biol.
18:444–452. 2006. View Article : Google Scholar : PubMed/NCBI
|