Introduction
Lung cancer is one of the most common malignancies,
which is the leading cause of cancer-associated mortality worldwide
(1). Non-small cell lung cancer
(NSCLC), including squamous cell carcinoma, adenocarcinoma and
large cell carcinoma are the most common types of lung cancer
(2). Notably, NSCLC constitutes
~85% of general lung cancers (3,4).
Notable improvements have been made in the treatment of NSCLC,
including surgical resection, chemotherapy, radiation therapy or a
combination of targeted therapy over the past several decades, the
prognosis of patients with NSCLC remains unfavorable; the total
survival rate is ~15% (5,6). Furthermore, the majority of patients
are diagnosed at the later period of lung cancer, which greatly
decreases the survival outcome (7). Thus, investigating the mechanism of
development and progression of NSCLC and identifying novel
therapeutic strategies for NSCLC is urgently required.
MicroRNAs (miRNAs/miRs) function as small non-coding
RNAs of 18–25 nucleotides that negatively regulate gene expression
by affecting translational repression or mRNA cleavage at the
post-transcriptional level via complementarities with the
3′untranslated region (UTR) of their target genes (8,9).
Dysregulation of miRNA expression is often associated with a
variety of human malignancies and increasing evidence indicates
that miRNAs may function as either oncogenes or tumor suppressors
(9,10). At present, numerous miRNAs have
been reported to be upregulated or downregulated in NSCLC (11–13).
For example, Sun et al (14) reported that miR-503-3p inhibits
lung cancer cell viability and induces cell apoptosis by regulating
p21 and cyclin dependent kinase 4 expression in lung cancer cells.
In addition, Li et al (15)
reported that ectopic expression of miR-146b-5p suppresses cell
proliferation, clonogenicity, migration and invasion, and also
induces G1 arrest in vitro, but does not induce cell
apoptosis by regulating matrix metalloproteinase 16 and tumor
necrosis factor receptor associated factor 6 expression in NSCLC.
Furthermore, Huang et al (16) reported that miR-219-5p exerts the
tumor-suppressive function by inhibiting the activation of the
protein kinase B (AKT) and extracellular signal-regulated kinase
(ERK) pathways in NSCLC cells; however, the role and mechanism of
regulation of miR-577 in NSCLC remain unclear.
In the present study, miR-577 was demonstrated to be
downregulated in NSCLC tissues and cell lines; low miR-577
expression levels were associated with larger tumor size, advanced
tumor, node, metastasis (TNM) stage and lymph node metastasis of
patients with NSCLC. Functional analysis revealed that miR-577
overexpression promoted cell proliferation. In addition, Transwell
analysis revealed that the inhibitory effect of miR-577
overexpression on cell migration and invasion functions by
inhibiting the epithelial-mesenchymal transition (EMT) process in
NSCLC cells. Furthermore, Wnt family member 2B (WNT2B) may be a
target of miR-577 and serves the oncogenic role in NSCLC
progression by activating the Wnt/β-catenin signaling pathway.
Collectively, the findings of the present study suggested that
miR-577 may inhibit NSCLC progression via the direct targeting of
WNT2B; the Wnt/β-catenin signaling pathway may be involved in the
regulatory mechanism.
Materials and methods
Tissue samples
A total of 25 NSCLC tissues and the adjacent normal
lung tissues were obtained from patients (n=25; 13 male and 12
female; aged 39–78 years) admitted to Tianjin Huanhu Hospital
(Tianjin, China) between March 2013 and March 2016. All of the
samples were obtained with the patients' informed consent. The
entire investigation conformed to the principles outlined in The
Declaration of Helsinki. The present study was approved by the
ethical review committees of Tianjin Huanhu Hospital.
Cell cultures
Human NSCLC cell lines, including H650, A549, H522,
H1299 and H1155 were purchased from the Cell Bank of Type Culture
Collection of the Chinese Academy of Sciences (Shanghai, China),
and human normal bronchial epithelial cells (HBECs) were purchased
from Shanghai Maisha Biotechnology (http://maishabio.biogo.net/; Shanghai, China). The
cells were routinely grown in Dulbecco's modified Eagle's medium
(DMEM; Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo
Fisher Scientific, Inc.), 1% penicillin/streptomycin mix
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), 2 mM glutamine
(Gibco; Thermo Fisher Scientific, Inc.), 5 mM glucose (Sigma
Aldrich; Merck KGaA) and 1 mM sodium pyruvate (Sigma Aldrich; Merck
KGaA) at 37°C in a humidified atmosphere containing 5%
CO2.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) assay
Total RNA was extracted from cultured cells and
NSCLC tissues using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
miRNAs from cancer specimens or cells were extracted using an
RNeasy kit or miRNeasy mini kit (Qiagen GmbH, Hilden, Germany),
respectively, according to the manufacturer's protocols. miRNAs and
mRNAs were reverse transcribed using a miScript reverse
transcription kit (Qiagen GmbH) following the manufacturer's
protocols. qPCR was performed using a miRNA-specific TaqMan MiRNA
Assay kit (Applied Biosystems; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocols using a Applied
Biosystems 7500 Fast Real-Time PCR system (Thermo Fisher
Scientific, Inc.). The qPCR conditions were as follows: 94°C
pre-denaturation for 5 min, followed by 33 cycles of denaturation
at 94°C for 30 sec, annealing and synthesis at 58°C for 30 sec.
Relative gene expression data was analyzed using the
2−ΔΔCq method (17).
The primers used for RT-qPCR were as follows: miR-577-RT
5′-GTCGTATCCAGTGCAGGGTCCGAGGTGCACTGGATACGACCAGGTA-3′; oligod T
5′-TTTTTTTTTTTTTTTTTT-3′; U6-RT
5′-GTCGTATCCAGTGCAGGGTCCGAGGTGCACTGGATACGACAAAATATGG-3′;
miR-577-qPCR, forward 5′-TGCGGTAGATAAAATATTGG-3′, reverse
5′-GTGCAGGGTCCGAGGT-3′; U6-qPCR, forward
5′-GCTTCGGCAGCACATATACTAAAAT-3′, reverse
5′-CGCTTCACGAATTTGCGTGTCAT-3′; WNT2B-qPCR, forward
5′-GCTGGACCAAACCTGAAC-3′, reverse 5′-CAAGAAGTATCGGGAAGC-3′; and
β-actin-qPCR, forward 5′-CCGTCTTCCCCTCCATCGTGGG-3′, reverse
5′-CGCAGCTCATTGTAGAAGGTGTGG-3′.
Plasmid construction
WNT2B was overexpressed using PCR-amplified cDNA of
H522 cells, which was cloned between the KpnI and XbaI restriction
sites into the pcDNA3 vector (Beyotime Institute of Biotechnology,
Shanghai, China). Overexpression was confirmed by RT-qPCR and
western blot analysis. The pcDNA3 vector alone was used as the
control group. In order to overexpress miR-577, the primary miR-577
was amplified from genomic DNA of H522 cells and cloned into the
pcDNA3 vector between the BamHI and EcoRI restriction sites. To
inhibit the function of miR-577, the 2-O-methyl-modifed antisense
oligonucleotide of miR-577 (ASO-miR-577) and the scramble control
oligonucleotides (ASO-NC) from the Shanghai GenePharma, Co., Ltd.
(Shanghai, China) were used. The pSilencer/shR-WNT2B (shR-WNT2B)
plasmid expressing siRNA targeting WNT2B was constructed by
annealing double-stranded hairpin cDNA and inserting it into the
pSilencer 2.1-U6 neo vector (Ambion; Thermo Fisher Scientific,
Inc.) at the BamHI and EcoRI sites. The primers and sequences used
are listed as follows: Pri-miR-577-sense (S)
5′-CGGGGTACCTGGTAGGTGCCCTGTTGA-3′, pri-miR-577-antisense (AS)
5′-CCGGAATTCTGGAAAGTAACCACGAGA-3′; ASO-miR-577
5′-CAGGUACCAAUAUUUUAUCUA-3′; ASO-NC 5′-UCACAACCUCCUAGAAAGAGUAGA-3′;
WNT2B-S 5′-CGGGGTACCGCCACCATGTTGGATGGCCTTGGAGTG-3′; and WNT2B-AS
5′-TGCTCTAGATCAGGTTTGGTCCAGCCACTCTGCC-3′; shR-WNT2B-S
5′-GATCCCGGACTGATCTTGTCTACTTTCTCGAGAAAGTAGACAAGATCAGTCCGTTTTTGA-3′;
shR-WNT2B-AS
5′-AGCTTCCGGACTGATCTTGTCTACTTTCTCGAGAAAGTAGACAAGATCAGTCCGG-3′. For
transfection, H522 and A549 cells were seeded in 12-well plates at
a density of 1×107 cells/ml for 24 h, and were
subsequently transfected with 2 µg plasmids using
Lipofectamine® 2000 (DNA:Lipofectamine®
2000=1:2; Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocols and incubated for 4 h at 37°C.
Subsequently, the cells were cultured in DMEM supplemented with 10%
FBS for 48 h at 37°C prior to further analysis.
Prediction of miRNA targets
The hypothetical target of miR-577 was predicted
using miRDB (http://www.mirdb.org/) (18), miRNA.org and
TargetScan human7.1 (http://www.targetscan.org/vert_71/), which revealed
that the 3′UTR of WNT2B may be complementarily paired with the seed
sequences of miR-577.
Enhanced green fluorescent protein
(EGFP) reporter assay
The WNT2B 3′UTR was cloned into a pcDNA3/EGFP vector
(Shanghai GeneChem Co., Ltd., Shanghai, China) and mutations were
introduced at potential miR-577 binding sites. To determine that
the 3′UTR of WNT2B mRNA is directly targeted by miR-577,
2×106 H522 and A549 cells were cotransfected with 0.5 µg
pri-miR-577 or 20 nM ASO-miR-577 and 0.5 µg 3′UTR of WNT2B or the
mutant 3′UTR of WNT2Bin in 48-well plates using
Lipofectamine® 2000 (DNA:Lipofectamine®
2000=1:2; Invitrogen; Thermo Fisher Scientific, Inc.). The binding
site of miR-577 in the WNT2B 3′UTR was mutated as follows: 5′
UAACAUUAUUAACAUUUAGAA3′. After 48 h, the EGFP activity was measured
using a spectrophotometer set at 528 nm. Red fluorescent
protein-expressing plasmid was integrated as a transfection
efficiency control.
CCK-8 assay
H522 and A549 cells transfected as aforementioned
were seeded in 96-well plates at a density of 5×103
cells/well. A Cell Counting kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) was used to detect the
viability of H522 and A549 cells according to the manufacturer's
instructions. The absorbance at 450 nm was measured.
Colony formation assay
For the colony formation ability assay, H522 and
A549 cells were counted at 24 h post-transfection and seeded into
24-well plates at 500 cell/well. Culture medium was replaced every
3 days. After ~2 weeks, cells were washed with 1XPBS. Subsequently,
colonies were fixed with 4% paraformaldehyde at room temperature
for 30 min and stained with 1% crystal violet at room temperature
for 20 min. The number of colonies was counted under an inverted
microscope (Leica Microsystems GmbH, Wetzlar, Germany).
Transwell migration and invasion
assay
H522 and A549 cells transfected with the indicated
plasmids were collected and suspended in serum-free medium.
Subsequently, 6×105 cells were added to the upper
chamber covered with 12.5 mg Matrigel (BD Biosciences) in 50 ml
PBS, the lower chamber was filled with medium containing 20% FBS.
Following incubation at 37°C for 48 h, cells below the membrane
were fixed and stained with 0.5% crystal violet at room temperature
for 30 min, washed with 1X PBS, air-dried and observed under an
inverted microscope (Olympus Corporation, Tokyo, Japan). Cell
invasion was assessed using a Transwell system (Corning
Incorporated, Tewksbury, MA, USA). The number of migrated and
invaded cells was counted under a microscope.
Western blot analysis
A total of 6×108 A549 cells were lysed
using the Protein Extraction kit according to the manufacturer's
protocols (http://www.wanleibio.cn/; Wanlei
Biotechnology, Beijing, China); 30 µg total proteins were separated
by 10% SDS-PAGE and transferred onto nitrocellulose membranes (EMD
Millipore, Billerica, MA, USA). The membranes were blocked with 5%
non-fat milk in Tris-buffered saline containing Tween-20 for ~2 hat
room temperature, prior to incubation with the primary antibodies.
The membranes were probed with anti-E-cadherin (1:1,500; cat. no.
ab1416; Abcam, Cambridge, MA, USA), anti-intercellular adhesion
molecule 1 (ICAM1; 1:3,000; cat. no. ab223659; Abcam),
anti-Vimentin (1:2,000; cat. no. ab188499; Abcam), anti-WNT2B
(1:2,000; cat. no. ab50575; Abcam), anti-β-catenin (1:5,000; cat.
no. ab16051; Abcam), anti-cyclin D1 (1:5,000; cat. no. ab15196;
Abcam), anti-c-Myc (1:5,000; cat. no. ab39688; Abcam), p-GSK3β
(1:1,000; cat. no. ab131097; Abcam), total-GSK3β (1:3,000; cat. no.
ab2602; Abcam) and anti-GAPDH (1:5,000; cat. no. ab9485; Abcam)
antibodies overnight at 4°C. Subsequently, membranes were incubated
with a horseradish peroxidase-conjugated secondary antibody
(1:5,000; cat. no. 8889; Cell Signaling Technology, Inc., Danvers,
MA, USA) for 1 h at 37°C. An enhanced chemiluminescence system
(Thermo Fisher Scientific, Inc.) was used to detect the
immunoreactive bands. The relative protein expression levels were
normalized to that of GAPDH. The protein expression levels were
measured using Image Pro Plus software v.6.0 (Media Cybernetics,
Inc., Rockville, MD, USA).
TOP/FOP flash reporter assays
To determine the transcriptional activity of the Wnt
pathway, 6×106 A549 and H522 cells treated as indicated
were co-transfected with either the Wnt signaling reporter TOP
Flash or the negative control FOP Flash (EMD Millipore) according
to the manufacturer's protocols. A549 and H522 cells were
transiently transfected with either 2 µg pTOP flash or pFOP flash
plasmids and 0.5 µg pSV40-Renilla plasmid as an internal
control (Promega Corporation, Madison, WI, USA) using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) at room temperature for 48 h. The dual luciferase
reporter assay system (Dual-Luciferase® Reporter Assay
system; cat. no. E1910; Promega Corporation) was used to assay the
firefly and Renilla luciferase activity ratio.
Immunofluorescence staining
A549 cells transfected with specific plasmids were
seeded in 24-well plates at the indicated time points before
immunofluorescence staining. The cells were washed in PBS and fixed
with 4% paraformaldehyde for 30 min at room temperature. After
cells were washed with PBS, the cells were permeabilized using
0.25% Triton-X-100 for 5 min at room temperature and blocked in 10%
donkey serum (BeyotimeInstitute of Biotechnology; Nanjing, China)
for 30 min. The cells were subsequently incubated with primary
antibodies against β-catenin (1:100; cat. no. ab16051; Abcam)
overnight at 4°C. The following day, cells were washed in PBS and
then incubated at room temperature for 1 h with a
fluorescent-labeled secondary antibody (1:200; cat. no. A0562;
BeyotimeInstitute of Biotechnology), followed by incubation with
DAPI (1:1,000; cat. no. C1002; BeyotimeInstitute of Biotechnology).
Images were captured under a confocal microscope.
Statistical analysis
All analyses were performed using SPSS v. 19.0 for
Windows (IBM Corp., Armonk, NY, USA) and GraphPad Prism v. 5.0 for
Windows (GraphPad Software Inc., La Jolla, CA, USA). For
comparisons of two treatment groups, a Student's t-test was used.
For comparisons of three or more groups, one-way analysis of
variance was followed by the Bonferroni post hoc test for
comparison of two selected treatment groups; the Dunnett's post hoc
test was used for comparisons of the other treatment groups with
the corresponding controls. The Pearson's correlation analysis was
used to determine the r-value. Associations between miR-577
expression and clinicopathological characteristics were assessed
using chi-squared test. Data from at least three independent
experiments are presented as the means ± standard deviation, or
medians with ranges. P<0.05 was considered to indicate a
statistically significant difference.
Results
miR-577 is downregulated in NSCLC
tissues and cell lines
The relative expression levels of miR-577 in NSCLC
tissues and cells were measured via RT-qPCR. The results revealed
that miR-577 expression levels were significantly lower in NSCLC
tissues than that in the adjacent normal tissues (Fig. 1A). In addition, the miR-577
expression levels in NSCLC patients were inversely associated with
the tumor size, TNM stage and lymph node metastasis, but not the
histological grade, age and gender (Table I). Furthermore, the expression
levels of miR-577 were also detected in various human NSCLC cells
lines including H650, A549, H522, H1299 and H1155 cells. As
presented in Fig. 1B, miR-577
expression levels were lower in NSCLC cell lines compared with
HBECs (Fig. 1B). These results
suggested that miR-577 may have a suppressive role in NSCLC
progression.
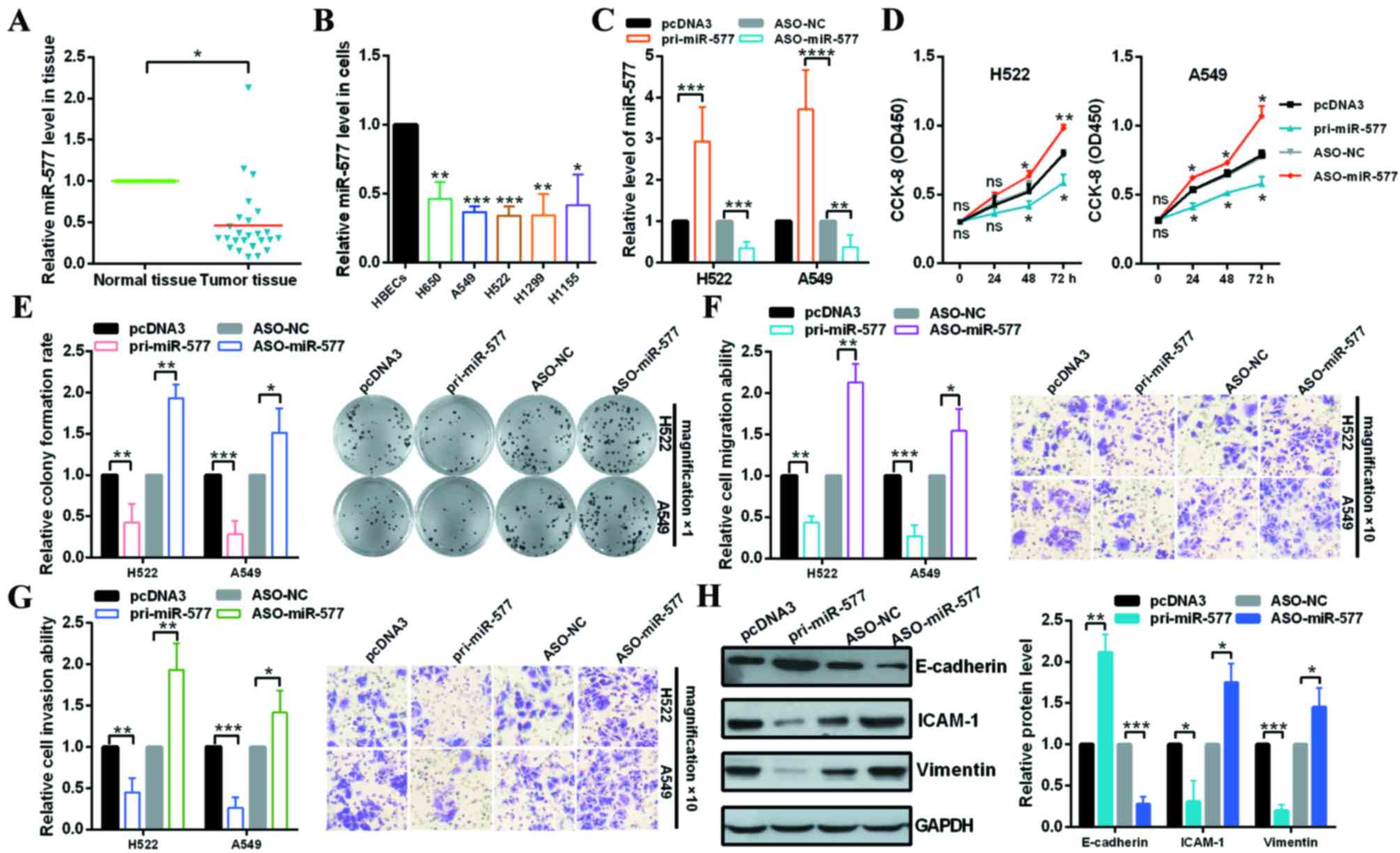 | Figure 1.miR-577 functions as a suppressor gene
in NSCLC cells. (A) Expression levels of miR-577 in NSCLC and
normal tissues were examined by RT-qPCR. (B) Expression levels of
miR-577 in HBECs, H650, A549, H522, H1299 and H1155 cells were
examined by RT-qPCR. (C) Efficiency of pri-miR-577 or ASO-miR-577
was identified by RT-qPCR. (D) Effect of miR-577 on H522 and A549
cellular viabilities were determined by CCK-8 assay. (E) Relative
colony formation rates of H522 and A549 cells with indicated
treatment were determined by colony formation assay. Original
magnification, ×1. (F) Transwell migration assays revealed that
miR-577 suppressed cell migration ability. Original magnification,
×10. (G) Transwell invasion assays demonstrated that miR-577
suppressed cell invasion ability. Original magnification, ×10. (H)
Western blot analysis of the protein expression levels of
E-cadherin, ICAM-1 and Vimentin following transfection with
pri-miR-577 or ASO-miR-577 and the control groups in A549 cells.
*P<0.05; **P<0.01 vs. control. NSCLC, non-small cell lung
cancer; RT-qPCR, reverse transcription-quantitative polymerase
chain reaction; miR, microRNA; HBECs, human normal bronchial
epithelial cells; ASO, antisense; NC, negative control; CCK-8, cell
counting kit-8; ns, no significance; ICAM-1, intercellular adhesion
molecule 1. |
 | Table I.Clinical and pathologic
characteristics of patients with non-small cell lung cancer. |
Table I.
Clinical and pathologic
characteristics of patients with non-small cell lung cancer.
|
|
| microRNA-577 |
|
|
|---|
|
|
|
|
|
|
|---|
| Variables | Total no. (n) | Low (n) | High (n) | χ2 | P-value |
|---|
| Age (years) |
|
|
|
| 0.337 |
|
≥60 | 12 | 6 | 6 | 0.5601 |
|
|
<60 | 13 | 8 | 8 |
|
|
| Gender |
|
|
|
| 0.821 |
|
Male | 13 | 7 | 6 | 0.51 |
|
|
Female | 12 | 7 | 5 |
|
|
| Tumor size
(cm) |
|
|
|
| 0.021 |
| ≥5 | 11 | 9 | 2 | 5.0314 |
|
|
<5 | 14 | 5 | 9 |
|
|
| TNM stage |
|
|
|
| 0.032 |
|
I+II | 10 | 3 | 7 | 4.573 |
|
|
III+IV | 15 | 11 | 4 |
|
|
| Lymph node
metastasis |
|
|
|
| 0.011 |
|
Positive | 16 | 12 | 4 | 6.512 |
|
|
Negative | 9 | 2 | 7 |
|
|
| Histological
type |
|
|
|
| 0.135 |
|
Squamous | 14 | 6 | 8 | 2.231 |
|
|
Adenocarinoma | 11 | 8 | 3 |
|
|
miR-577 inhibits cell proliferation,
cell migration, cell invasion and EMT processes in NSCLC cells
To further confirm whether miR-577 may affect NSCLC
tumorigenesis, pcDNA3, pri-miR-577, ASO-NC or ASO-miR-577 was
transfected into H522 and A549 cells. The results of RT-qPCR
analysis demonstrated that the expression plasmids were effective
(Fig. 1C). CCK-8 and colony
formation assays revealed that the overexpression of miR-577
significantly inhibited cell proliferation and knockdown of miR-577
significantly promoted cell proliferation of H522 and A549 cells
(Fig. 1D and E). Transwell
migration and invasion assays revealed that ectopic expression of
miR-577 significantly reduced the cell migration and invasion
ability of H522 and A549 cells; knockdown of miR-577 increased H522
and A549 cells migration and invasion (Fig. 1F and G). These suggest that miR-577
may have a suppressive effect on NSCLC metastasis. In order to
investigate the regulation of EMT by miR-577, the expression levels
of the EMT markers, E-cadherin, Vimentin and ICAM-1 were analyzed
by western blotting in A549 cells and the results indicated that
the expression levels of Vimentin and ICAM-1 were significantly
decreased by miR-577 overexpression but increased by miR-577
knockdown compared to the control groups; in addition, the
expression levels of E-cadherin were significantly increased by
miR-577 overexpression, but attenuated by miR-577 knockdown
(Fig. 1H).
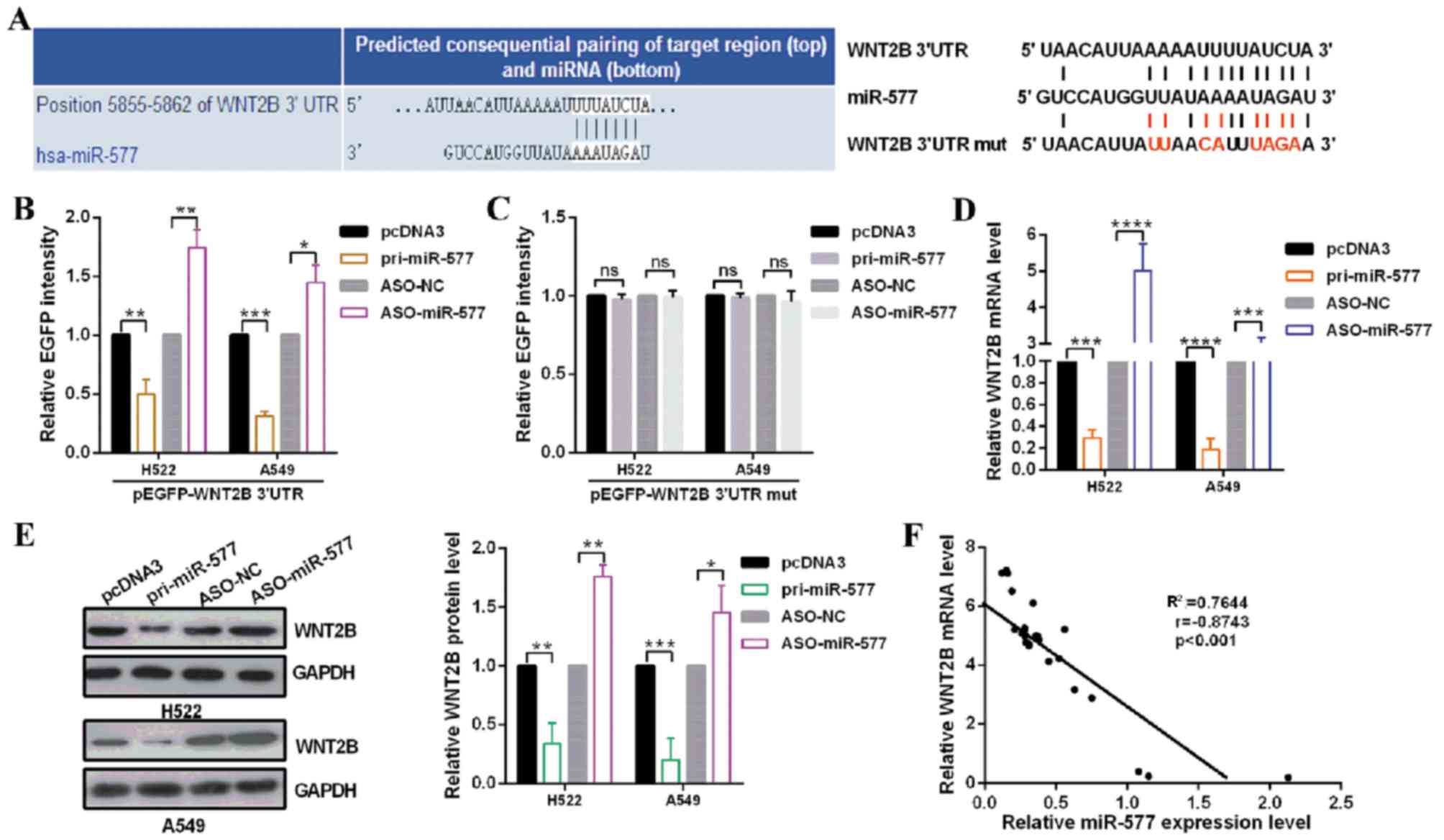
WNT2B is a direct target of
miR-577
To investigate the mechanism of miR-577 affecting
the biological behavior in NSCLC cells, the target genes of miR-577
were predicted using miRDB, miRNA.org and
TargetScan human7.1; WNT2B was predicted to be a target of miR-577
(Fig. 2A). To verify that miR-577
can directly target WNT2B mRNA, EGFP reporter plasmids containing
the 3′UTR or the 3′UTR-mut of WNT2B were constructed. In H522 and
A549 cells, the relative EGFP level was significantly reduced in
the pri-miR-577 and wild type 3′UTR of WNT2B co-transfected group.
Additionally, co-transfection with ASO-miR-577 and wild type 3′UTR
of WNT2B significantly increased the relative EGFP activity
(Fig. 2B); however, no significant
differences in EGFP activity were observed when H522 and A549 cells
were co-transfected with pri-miR-577 or ASO-miR-577 and mutational
EGFP reporter plasmids (Fig. 2C).
To further confirm the regulation of WNT2B by miR-577, RT-qPCR and
western blot assays were performed. The results revealed that
overexpression of miR-577 significantly decreased the mRNA and
protein levels of WNT2B; miR-577 knockdown significantly increased
the expression of WNT2B at the mRNA and protein levels, suggesting
that miR-577 downregulated WNT2B expression at the mRNA and protein
levels in H522 and A549 cells (Fig. 2D
and E). In addition, the present study reported that the mRNA
levels of WNT2B in tumor tissues were negatively correlated with
the miR-577 levels using Pearson's correlation analysis (r=−0.8743,
R2=0.7644; Fig. 2F).
These data indicated that WNT2B may be negatively regulated by
miR-577 in NSCLC cells, which is the novel target of miR-577.
Upregulated WNT2B promotes the
malignancy of NSCLC cells
To determine the role of WNT2B in the aggressiveness
of NSCLC, the mRNA expression levels of WNT2B in NSCLC cell lines
were analyzed by RT-qPCR. The results demonstrated that WNT2B
expression levels were significantly upregulated in NSCLC cells
compared with in HBECs (Fig. 3A).
In addition, the protein expression levels of WNT2B were detected
by western blot assays, which also revealed that WNT2B was
increased in NSCLC cells compared with in HBECs cells (Fig. 3B). As presented in Fig. 3C and D, the overexpression or
knockdown of WNT2B were effective in H522 and A549 cells. At 24, 48
and 72 h post-transfection, cell viabilities of H522 and A549 cells
were significantly increased by WNT2B overexpression and decreased
by WNT2B knockdown (Fig. 3E). In
addition, the relative colony formation rate was significantly
increased by pWNT2B-transfected and decreased by
shR-WNT2B-transfected compared with in the control groups (Fig. 3F). To identify whether WNT2B also
influences the migration and invasion of NSCLC cells, Transwell
migration and invasion assays were performed in H522 and A549
cells. As expected, overexpression of WNT2B significantly promoted
cell migration and invasion ability (Fig. 3G and H). Finally, whether WNT2B may
regulate the protein expression levels of the EMT process was
investigated, including E-cadherin, ICAM-1 and Vimentin. Western
blot analysis demonstrated that the overexpression of WNT2B
significantly decreased E-cadherin expression, but increased ICAM-1
and Vimentin expression levels in A549 cells (Fig. 3I). Collectively, these results
indicated that upregulation of WNT2B promotes NSCLC malignant
behavior.
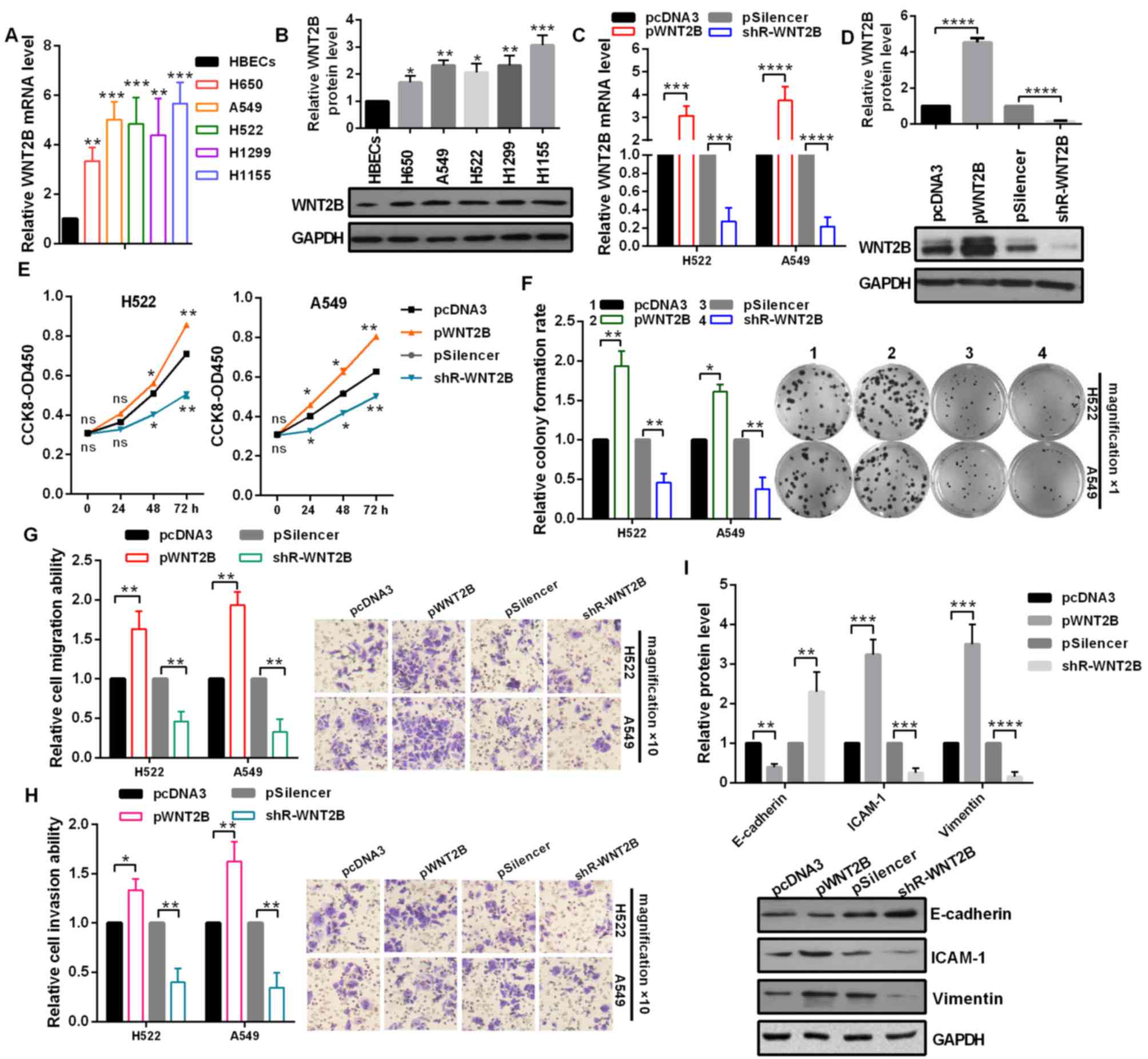 | Figure 3.WNT2B promotes the malignant phenotype
of non-small cell lung cancer cells. (A) mRNA expression levels of
WNT2B in HBECs, H650, A549, H522, H1299 and H1155 cells were
examined by RT-qPCR. (B) Protein expression levels of WNT2B in
HBECs, H650, A549, H522, H1299 and H1155 cells were examined by
western blot analysis. (C) RT-qPCR demonstrated that overexpression
and knockdown of WNT2B was efficient. (D) Western blot analysis
revealed that overexpression and knockdown of WNT2B was efficient
in A549 cells. (E) CCK-8 assay demonstrated that WNT2B
overexpression increased cell viability and WNT2B knockdown
decreased cell viability. (F) Relative colony formation rate was
higher following transfection with pWNT2B and lower with shR-WNT2B
compared with the control groups. Original magnification, ×1. (G)
Transwell migration assays revealed that WNT2B overexpression
promoted cell migration ability. Original magnification, ×10. (H)
Transwell invasion assays revealed that WNT2B overexpression
promoted cell invasion ability. Original magnification, ×10. (I)
Western blot analysis demonstrated the protein expression levels of
E-cadherin, ICAM-1 and Vimentin, following transfection with pWNT2B
or shR-WNT2B in A549 cells. *P<0.05; **P<0.01; ***P<0.001.
RT-qPCR, reverse transcription-quantitative polymerase chain
reaction; WNT2B, Wnt family member 2B; shR, short hairpin RNA;
CCK-8, cell counting kit-8; ns, no significance; ICAM-1,
intercellular adhesion molecule 1. |
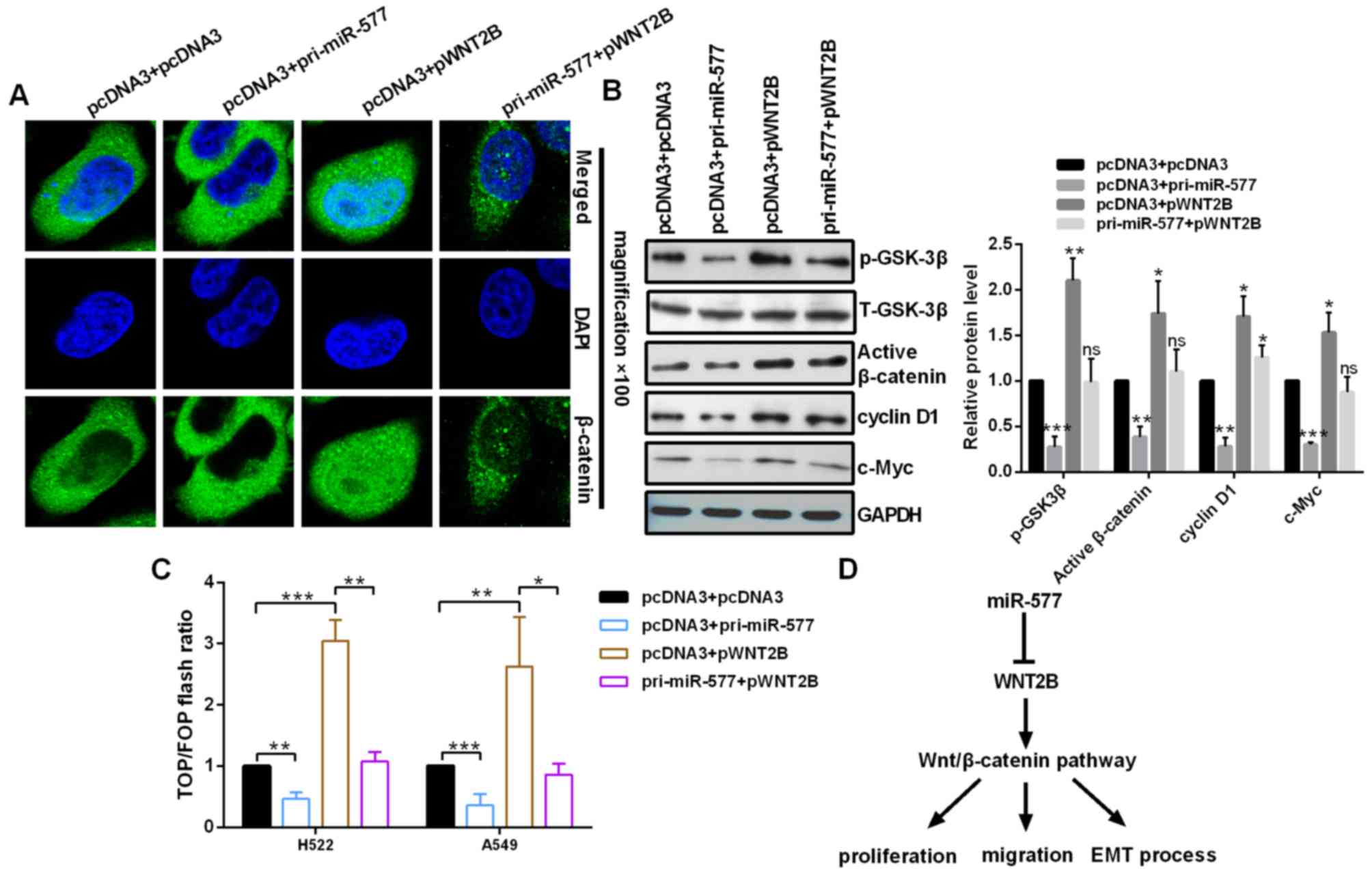
Upregulated miR-577 inhibits the
Wnt/β-catenin signaling pathway by regulating WNT2B in NSCLC
cells
To address the miR-577 whether regulates the
Wnt/β-catenin signaling pathway in NSCLC cells, the expression
levels of WNT-associated genes and the activity of β-catenin were
analyzed in the context of miR-577 overexpression or miR-577
overexpression in conjunction with pWNT2B in NSCLC cells. The
nuclear distribution of β-catenin in A549 cells was decreased in
pri-miR-577-transfected cells and increased in pWNT2B-transfected
cells (Fig. 4A). Western blot
assays revealed that the expression levels of phosphorylated
(p)-GSK3β, activated β-catenin, c-Myc and cyclin D1 were
significantly reduced by miR-577 overexpression and significantly
increased by WNT2B overexpression (Fig. 4B). In addition, a TOP/FOP flash
assay was performed, which is often used to detect Wnt/β-catenin
signaling. The results demonstrated that miR-577 overexpression
decreased the TOP/FOP flash ratio, but WNT2B overexpression
increased it in NSCLC cells, indicating inhibition and activation
of the Wnt/β-catenin signaling pathway, respectively (Fig. 4C).
Discussion
Dysregulation of miRNAs is important in the genesis
and development of various human diseases and cancer, such as NSCLC
(19,20). Previous studies have reported that
miR-577 is downregulated in numerous cancer types. For example,
Jiang et al (21) reported
that miR-577 was downregulated in colorectal cancer (CRC) specimens
and cell lines and miR-577 overexpression suppressed the
proliferation and colony formation and induced a G0/G1 cell cycle
arrest in CRC cells. In addition, Chen et al (22) indicated that miR-577 inhibited
pancreatic β-cell function and survival by targeting fibroblast
growth factor 21 via the activation of ERK1/2 and AKT signaling
pathways in pediatric diabetes. Furthermore, Wang et al
(23) reported that miR-577 mimics
may repress cell proliferation, enhance cell apoptosis and block
the cell cycle in G0/G1 phase in hepatocellular carcinoma. However,
Yuan et al (24) reported
that miR-577 overexpression and testis specific 10 downregulation
promoted cell proliferation and a more rapid G1-S phase transition
in esophageal squamous cell carcinomas. These studies suggested
that miR-577 serves important roles in numerous types of human
cancer and may be considered as a therapeutic target. Thus, it is
important to investigate the role of miR-577 in a variety of
cancers. In the present study, the results revealed significant
downregulation of miR-577 in NSCLC tissues and cell lines. miR-577
overexpression significantly inhibited cell proliferation,
migration and invasion of NSCLC cells in vitro and reduced
the expression levels of ICAM-1 and Vimentin and enhancing those of
E-cadherin. This suggested that this miRNA may also function as a
suppressor gene in NSCLC cells. In addition, miR-577 overexpression
inactivated the Wnt/β-catenin pathway in NSCLC cells.
It has been reported that miRNAs have roles in a
number of cancers via complementary base pairing with the 3′UTR of
their target genes (25). In the
present study, bioinformatics were performed to predict target
genes; WNT2B may be a novel candidate target of miR-577. Notably,
the mRNA expression levels of WNT2B were significantly increased in
H650, A549, H522, H1299 and H1155 cell lines, and were inversely
correlated with miR-577 expression levels in NSCLC cells.
Furthermore, the EGFP reporter assay indicated that miR-577 binds
directly to the 3′UTR of WNT2B mRNA within H522 and A549 cells. The
results from RT-qPCR and western blot analyses in the present study
revealed that miR-577 overexpression significantly reduced the
expression of WNT2B at mRNA and protein levels; miR-577 knockdown
significantly increased the expression of WNT2B at mRNA and protein
levels in H522 and A549 cells.
The canonical Wnt/β-catenin signaling pathway is one
of the most critical developmental pathways that are considered
important for progenitor cell fate, developmental decisions,
proliferation during embryonic development, adult tissue
homeostasis and tumor microenvironment (26–28).
WNT2B has been reported to stimulate the canonical Wnt/β-catenin
signaling pathway and affect various malignant tumor progressions
(29). In nasopharyngeal
carcinoma, WNT2B was significantly upregulated and higher
expression of WNT2B was closely correlated with TNM stage, clinic
stage and cervical lymph node metastasis (30). Wang et al (31) suggested that WNT2B knockdown
inhibited cell colony formation and metastasis and enhanced
chemotherapy sensitivity via the caspase-9/Bcl-2/Bcl-xl and
EMT/p-AKT signaling pathways in ovarian cancer (31). These studies highlighted the role
of WNT2B in malignant processes of tumors; however, the function of
WNT2B in NSCLC remains unclear. The results of the present study
demonstrated that the mRNA and protein expression levels of WNT2B
in NSCLC cell lines were upregulated, and that overexpression of
WNT2B may have promoted cell viability, colony formation ability
and the EMT process in NSCLC cells. In addition, ectopic WNT2B may
activate the Wnt/β-catenin pathway in NSCLC cells.
In conclusion, the present study confirmed that the
downregulation of miR-577 is a common phenomenon in NSCLC tissues
and cell lines, and identified that miR-577 serves an important
role in regulating cell proliferation, migration and invasion, and
the EMT process in NSCLC cells. In addition, miR-577 may have
inhibited WNT2B expression at both the mRNA and protein expression
levels by directly targeting the 3′UTR of the WNT2B mRNA.
Furthermore, miR-577 may inhibit the activation of the
Wnt/β-catenin signaling pathway by regulating WNT2B in NSCLC cells
(Fig. 4D). Collectively, these
findings may provide a novel insight into tumorigenesis and miR-577
may be considered as a potential biomarker for NSCLC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
RJ conceived and supervised the project. BW and RJ
performed the experiments. LS and JL analyzed the data. BW and RJ
wrote the manuscript. All the authors reviewed the manuscript.
Ethics approval and consent to
participate
The present study was approved by the ethical review
committees of Tianjin Huanhu Hospital. All of the samples were
obtained with the patients' informed consent.
Patient consent for publication
All patients provided consent for publication.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Osugi J, Muto S, Matsumura Y, Higuchi M,
Suzuki H and Gotoh M: Prognostic impact of the high-sensitivity
modified Glasgow prognostic score in patients with resectable
non-small cell lung cancer. J Cancer Res Ther. 12:945–951. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Buettner R, Wolf J and Thomas RK: Lessons
learned from lung cancer genomics: The emerging concept of
individualized diagnostics and treatment. J Clin Oncol.
31:1858–1865. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ramnath N, Dilling TJ, Harris LJ, Kim AW,
Michaud GC, Balekian AA, Diekemper R, Detterbeck FC and Arenberg
DA: Treatment of stage III non-small cell lung cancer: Diagnosis
and management of lung cancer, 3rd ed: American college of chest
physicians evidence-based clinical practice guidelines. Chest. 143
5 Suppl:e314S–e340S. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Novaes FT, Cataneo DC, Junior Ruiz RL,
Defaveri J, Michelin OC and Cataneo AJ: Lung cancer: Histology,
staging, treatment and survival. J Bras Pneumol. 34:595–600. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zaporozhchenko IA, Morozkin ES, Skvortsova
TE, Ponomaryova AA, Rykova EY, Cherdyntseva NV, Polovnikov ES,
Pashkovskaya OA, Pokushalov EA, Vlassov VV and Laktionov PP: Plasma
miR-19b and miR-183 as potential biomarkers of lung cancer. PLoS
One. 11:e01652612016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ameres SL and Zamore PD: Diversifying
microRNA sequence and function. Nat Rev Mol Cell Biol. 14:475–488.
2013. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen CZ: MicroRNAs as oncogenes and tumor
suppressors. N Engl J Med. 353:1768–1771. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu N, Zhang Q, Liu Q, Yang J and Zhang S:
A meta-analysis: MicroRNAs' prognostic function in patients with
nonsmall cell lung cancer. Cancer Med. 6:2098–2105. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen Y, Lu L, Feng B, Han S, Cui S, Chu X,
Chen L and Wang R: Non-coding RNAs as emerging regulators of
epithelial to mesenchymal transition in non-small cell lung cancer.
Oncotarget. 8:36787–36799. 2017.PubMed/NCBI
|
|
13
|
Legras A, Pécuchet N, Imbeaud S, Pallier
K, Didelot A, Roussel H, Gibault L, Fabre E, Le Pimpec-Barthes F,
Laurent-Puig P and Blons H: Epithelial-to-mesenchymal transition
and MicroRNAs in lung cancer. Cancers (Basel). 9:pii: E101. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sun Y, Li L, Xing S, Pan Y, Shi Y, Zhang L
and Shen Q: miR-503-3p induces apoptosis of lung cancer cells by
regulating p21 and CDK4 expression. Cancer Biomark. 20:597–608.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li Y, Zhang H, Dong Y, Fan Y, Li Y, Zhao
C, Wang C, Liu J, Li X, Dong M, et al: MiR-146b-5p functions as a
suppressor miRNA and prognosis predictor in non-small cell lung
cancer. J Cancer. 8:1704–1716. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huang LX, Hu CY, Jing L, Wang MC, Xu M,
Wang J, Wang Y, Nan KJ and Wang SH: microRNA-219-5p inhibits
epithelial-mesenchymal transition and metastasis of colorectal
cancer by targeting lymphoid enhancer-binding factor 1. Cancer Sci.
108:1985–1995. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wong N and Wang X: miRDB: An online
resource for microRNA target prediction and functional annotations.
Nucleic Acids Res. 43:(Database Issue). D146–D152. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hou J, Meng F, Chan LW, Cho WC and Wong
SC: Circulating plasma MicroRNAs as diagnostic markers for NSCLC.
Front Genet. 7:193eCollection. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Del Vescovo V and Denti MA: microRNA and
lung cancer. Adv Exp Med Biol. 889:153–177. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jiang H, Ju H, Zhang L, Lu H and Jie K:
microRNA-577 suppresses tumor growth and enhances chemosensitivity
in colorectal cancer. J Biochem Mol Toxicol. 31:2017. View Article : Google Scholar
|
|
22
|
Chen XY, Li GM, Dong Q and Peng H: MiR-577
inhibits pancreatic β-cell function and survival by targeting
fibroblast growth factor 21 (FGF-21) in pediatric diabetes. Genet
Mol Res. 14:15462–15470. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang LY, Li B, Jiang HH, Zhuang LW and Liu
Y: Inhibition effect of miR-577 on hepatocellular carcinoma cell
growth via targeting β-catenin. Asian Pac J Trop Med. 8:923–929.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yuan X, He J, Sun F and Gu J: Effects and
interactions of MiR-577 and TSGA10 in regulating esophageal
squamous cell carcinoma. Int J Clin Exp Pathol. 6:2651–2667.
2013.PubMed/NCBI
|
|
25
|
Farazi TA, Juranek SA and Tuschl T: The
growing catalog of small RNAs and their association with distinct
Argonaute/Piwi family members. Development. 135:1201–1214. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Majidinia M, Aghazadeh J,
Jahanban-Esfahlani R and Yousefi B: The roles of Wnt/β-catenin
pathway in tissue development and regenerative medicine. J Cell
Physiol. 233:5598–5612. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Murillo-Garzón V and Kypta R: WNT
signalling in prostate cancer. Nat Rev Urol. 14:683–696. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nusse R and Clevers H: Wnt/β-catenin
signaling, disease, and emerging therapeutic modalities. Cell.
169:985–999. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Katoh M: Differential regulation of WNT2
and WNT2B expression in human cancer. Int J Mol Med. 8:657–660.
2001.PubMed/NCBI
|
|
30
|
Liu C, Li G, Yang N, Su Z, Zhang S, Deng
T, Ren S, Lu S, Tian Y, Liu Y and Qiu Y: miR-324-3p suppresses
migration and invasion by targeting WNT2B in nasopharyngeal
carcinoma. Cancer Cell Int. 17:22017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang H, Fan L, Xia X, Rao Y, Ma Q, Yang J,
Lu Y, Wang C, Ma D and Huang X: Silencing Wnt2B by siRNA
interference inhibits metastasis and enhances chemotherapy
sensitivity in ovarian cancer. Int J Gynecol Cancer. 22:755–761.
2012. View Article : Google Scholar : PubMed/NCBI
|