Introduction
The skeletal system is one of the most important
systems in the human body. It serves as the body's structural
support, provides a framework for attachment of tissues, protects
vital organs and helps direct the forces necessary for movement
(1,2). Bone homeostasis depends on a dynamic
balance between osteoblasts and osteoclasts (2). An imbalance in bone homeostasis leads
to metabolic diseases such as osteoporosis, osteopetrosis, and
Paget's diseases (3). Therefore,
the components and mechanisms of bone homeostasis have been widely
investigated. Osteoblast proliferation and differentiation are
important in maintaining bone metabolic balance, and these
processes are regulated by numerous signals and pathways.
Platelet-derived growth factor-BB (PDGF-BB) can
promote the proliferation and differentiation of human osteoblasts
(4), and stimulate the synthesis
of interleukin-6 in osteoblasts (5), thus promoting bone formation and
regeneration (6). Following the
binding of PDGF-BB to its receptor (PDGFR-β), endogenous tyrosine
phosphorylating activity and intercellular signaling are activated
(7). PDGF-BB secreted by
preosteoclasts promotes bone formation through stimulating the
migration and angiogenesis of endothelial progenitor cells and MSCs
(8).
Src interacts with PDGFR through its Src homology 2,
coiled-coil and FCH (Fer/Fes/Fps/Cip4 homology) domains (9). Src not only serves key functions in
physiological and pathological processes, including cell survival,
cell differentiation, inflammation (10), tumorigenesis, invasion and
metastasis (11), but it also
serves a critical function in bone metabolism. Although western
blot analysis has indicated that PDGF-BB induces the
phosphorylation of Src and Janus kinase 2 (JAK2) in pancreatic
stellate cells in a time-dependent manner (7), the interaction between Src and
PDGF-BB in osteoblast cells is poorly understood.
The JAK family consists of four members, JAK1, JAK2,
JAK3 and Tyk2, which are activated by cytokines binding their
receptor. This activation leads to the subsequent phosphorylation
and activation of signal transducer and activation of transcription
(STAT) transcription factors (3,12).
JAK2 was previously demonstrated to be associated with MC3T3-E1
cell proliferation in conditioned media from mouse osteosarcoma
cells (13).
The aim of the present study was to investigate the
complex associations between PDGF-BB, Src and JAK2, and to
investigate their interaction in MC3T3-E1 cells.
Materials and methods
Reagents
Antibodies against phosphorylated (p-)PDGFRβ (cat.
no. 3161), PDGFRβ (cat. no. 3175), p-Src (cat. no. 2105), Src (cat.
no. 2110), p-JAK2 (cat. no. 3771), JAK2 (cat. no. 3230; all
dilution, 1:1,000), GAPDH (cat. no. 5174), β-actin (cat. no. 8457;
both dilution, 1:2,500) and Runt-related transcription factor 2
(RUNX2; cat. no. 98059; dilution, 1:2,000), were purchased from
Cell Signaling Technology, Inc. (Danvers, MA, USA). The OSTERIX
(cat. no. ab22552) and anti-COL1α1 (cat. no. ab166606; both
dilution, 1:2,000) antibodies were purchased from Abcam (Cambridge,
UK). Dexamethasone, ascorbic acid, β-glycerophosphate, AG1295 and
hexadecylpyridinium chloride monohydrate were purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). SU6656 and AG490
were purchased from Calbiochem (Merck KGaA). PDGF-BB was purchased
from Peprotech, Inc. (Rocky Hill, NJ, USA).
MC3T3-E1 cell culture
Murine osteoblast-like MC3T3-E1 cells (China Center
for Type Culture Collection, Wuhan, China) were cultured in α-MEM
complete medium supplemented with 10% fetal bovine serum (FBS) and
1% penicillin-streptomycin, at 37°C, in a humidified atmosphere of
95% air and 5% CO2. The cell culture medium was changed
every 2–3 days. Cells were divided into five groups, including
bovine serum albumin (BSA), PDGF-BB, PDGF-BB +20 µmol/l AG1295,
PDGF-BB + 2 µmol/l SU6656 and PDGF-BB + 10 µmol/l AG490 10 µmol/l).
All PDGF-BB used in this research was 25 ng/ml.
Western blot analysis
The MC3T3-E1 cells were cultured in a 6-well plate,
grown to 70–80% confluence, and then starved in serum-free medium
for 24 h. The cells were lysed in radioimmunoprecipitation assay
(RIPA) lysis buffer (Sigma-Aldrich; Merck KGaA) at 4°C for 30 min.
Equal amounts of protein quantified by BCA (60 µg) were loaded onto
10% SDS-polyacrylamide gels. Following electrophoresis, proteins
were transferred to polyvinylidene fluoride membranes (EMD
Millipore, Billerica, MA, USA). Blots were blocked with 5% skimmed
milk (Difco; BD Biosciences, Franklin Lakes, NJ, USA) at room
temperature for 1 h, then incubated with the aforementioned primary
antibodies overnight at 4°C. Afterwards, the blots were washed and
incubated with horseradish peroxidase-conjugated secondary
antibodies (Beijing TDY Biotech Co., Ltd., Beijing China; dilution,
1:5,000) for 2 h. Blots were detected using an ECL kit (GE
Healthcare, Chicago, IL, USA) according to the manufacturer's
protocol. Western blot bands were quantified by densitometric
analysis using Photoshop CS6 (Adobe Systems, Inc., San Jose, CA,
USA).
Osteoblast differentiation
Murine osteoblast-like MC3T3-E1 cells were cultured
at 5×104 cells/cm2 in osteogenic medium
containing 10% FBS, 0.1 µM dexamethasone, 10 mM β-glycerophosphate
and 50 µg/ml ascorbic acid for 21 days. At days 3 and 7, cells were
harvested for reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) analysis or western blotting. Cells were fixed
and alkaline phosphatase (ALP) staining was performed at days 3 and
7, whereas Alizarin Red staining was performed at days 14 and
21.
RNA isolation and RT-qPCR
Total RNA was extracted from cells with TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. Total RNA (1 µg) was used for cDNA
synthesis using the RevertAid First Strand cDNA Synthesis kit
(Eppendorf, Hamburg, Germany). qPCR was performed in triplicate
using SYBR Green Master mix (Takara Biotechnology Co., Ltd.,
Dalian, China) on an Applied Biosystems 7500 Real-Time PCR system
(Thermo Fisher Scientific, Inc.) as follows: 95°C for 30 sec,
followed by 40 cycles of 95°C for 5 sec, 60°C for 31 sec and 95°C
for 15 sec, then 60°C for 1 min and 95°C for 15 sec. The qPCR
results were automatically analyzed using the Applied Biosystems
7500 system. The 2−ΔΔCq method (14) was used to calculate the relative
gene expression level. The primers used are presented in Table I.
 | Table I.Primers used for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primers used for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|
| GAPDH |
TATCGGACGCCTGGTTAC |
CTGTGCCGTTGAACTTGC |
| RUNX2 |
TCATTCAGTGACACCACCAGG |
TGTAGGGGCTAAAGGCAAAA |
| OSTERIX |
AGAAGCCATACACTGACCTTTC |
GGTGGGTAGTCATTGGCATAG |
| ALP |
GAGATGGTATGGGCGTCTC |
GTTGGTGTTGTACGTCTTGGA |
| OCN |
GACAAGTCCCACACAGCAACT |
GGACATGAAGGCTTTGTCAGA |
| OPN |
CCCATCTCAGAAGCAGAATCTT |
GTCATGGCTTTCATTGGAGTTG |
| COL1α1 |
GACATGTTCAGCTTTGTGGACCTC |
GGGACCCTTAGGCCATTGTGTA |
ALP staining
Cells were washed twice with PBS, fixed with 4%
paraformaldehyde for 10 min, rinsed with deionized water and
stained with a 5-bromo-4-chloro-3′-indolyphosphate/nitro-blue
tetrazolium ALP color development kit (Beyotime Institute of
Biotechnology, Haimen, China) for 12 h away from direct light,
according to the manufacturer's protocol. Images were subsequently
obtained.
Alizarin Red staining and
mineralization assay
Cells were washed twice with cold PBS and fixed with
4% paraformaldehyde for 10 min. They were then stained with 50 mM
Alizarin Red S (pH 4.2) for 10 min at room temperature, and images
were obtained. In order to determine the extent of calcium
deposition, the mineralization of calcium nodules was quantified.
After staining, the cells were washed three times with PBS.
Hexadecylpyridinium chloride monohydrate (10%) was added and
incubated for 20 min at room temperature. The absorbance of the
supernatant was measured at 562 nm in triplicate using a Multiskan
EX plate reader (Power Wave XS2; Thermo Fisher
Scientific, Inc.). Finally, cells were washed in PBS and lysed with
RIPA buffer. and protein content quantified by BCA and calcium
levels were normalized to the total protein content.
Cell proliferation assay
Proliferation activity was measured using a Cell
Counting kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan) assay. MC3T3-E1 cells were seeded into a 96-well
plate at 2×104 cells/cm2. The medium was
removed following adherence, and the cells were continuously
cultured for 72 h. CCK-8 solution (10 µl) was added to 100 µl of
medium in each well, and incubated with the cells for 2 h, before
measuring the optical density (OD) at 450 nm. All experiments were
performed in six replicates.
Statistical analysis
All quantitative data are expressed as the mean ±
standard deviation. Analysis was performed with GraphPad Prism 5
(GraphPad Software, Inc., La Jolla, CA, USA). Comparisons were
evaluated with one-way or two-way analysis of variance followed by
Tukey's post-hoc test. P<0.05 was considered to indicate a
statistically significant difference. Each experiment was repeated
≥3 times, and representative images are presented.
Results
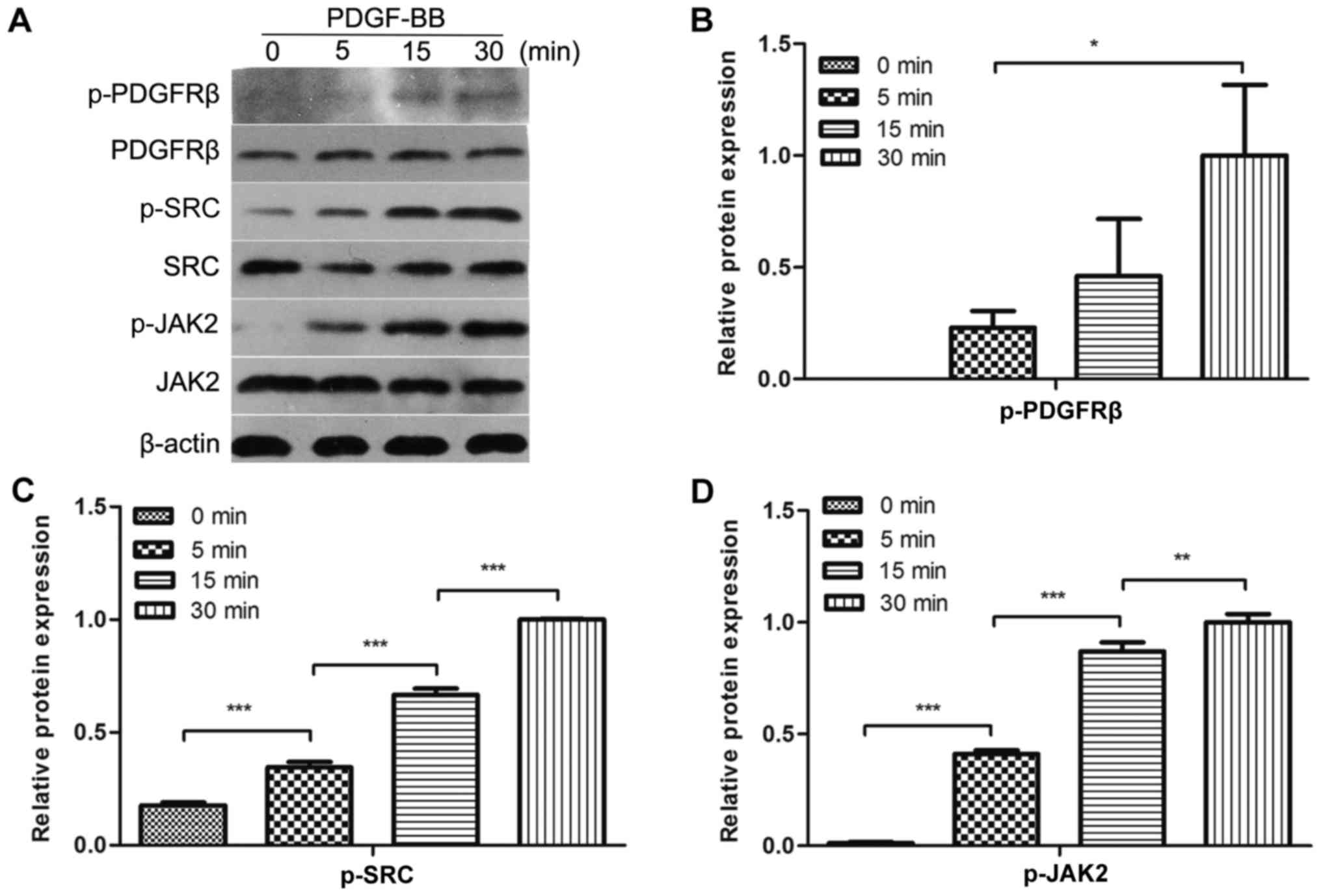
PDGF-BB activates PDGFRβ, Src and JAK2
in MC3T3-E1 cells
To evaluate whether PDGF-BB could activate PDGFRβ,
Src and JAK2 in MC3T3-E1 cells, the cells were treated with PDGF-BB
at a range of different durations, followed by the analysis of
PDGFRβ, Src and JAK2 phosphorylation. It was identified that
PDGFRβ, Src and JAK2 were rapidly phosphorylated by PDGF-BB at 5
min and their phosphorylation levels increased over time for the
first 30 min (Fig. 1A-D). These
results indicated that PDGFRβ, Src and JAK2 were expressed in
MC3T3-E1 cells and could be activated by PDGF-BB in a
time-dependent manner within 30 min.
Src lies upstream of JAK2 in MC3T3-E1
cells
Subsequently, it was investigated whether the
phosphorylation of Src was performed directly by PDGFRβ, and
whether Src was an upstream molecule of JAK2, in MC3T3-E1 cells.
Therefore, the effects of low molecular mass inhibitors against
PDGFRβ (AG1295), Src family kinases (SU6656) and JAK2 (AG490) on
Src phosphorylation were determined. Inhibition of PDGFRβ and Src
kinase activity resulted in the inhibition of JAK2 phosphorylation
(Fig. 2A and D). However, the
inhibition of JAK2 activity did not affect PDGF-BB-induced Src
phosphorylation (Fig. 2A and C).
SU6656 and AG490 inhibited the activity of PDGFRβ (Fig. 2B), while previous study showed that
AG490 had no effect on PDGFRβ (9).
Therefore, we hypothesized that Src and JAK2 may be involved in
other pathways that affect the phosphorylation of PDGFRβ in
MC3T3-E1 cells, and that Src was upstream of JAK2 in MC3T3-E1
cells.
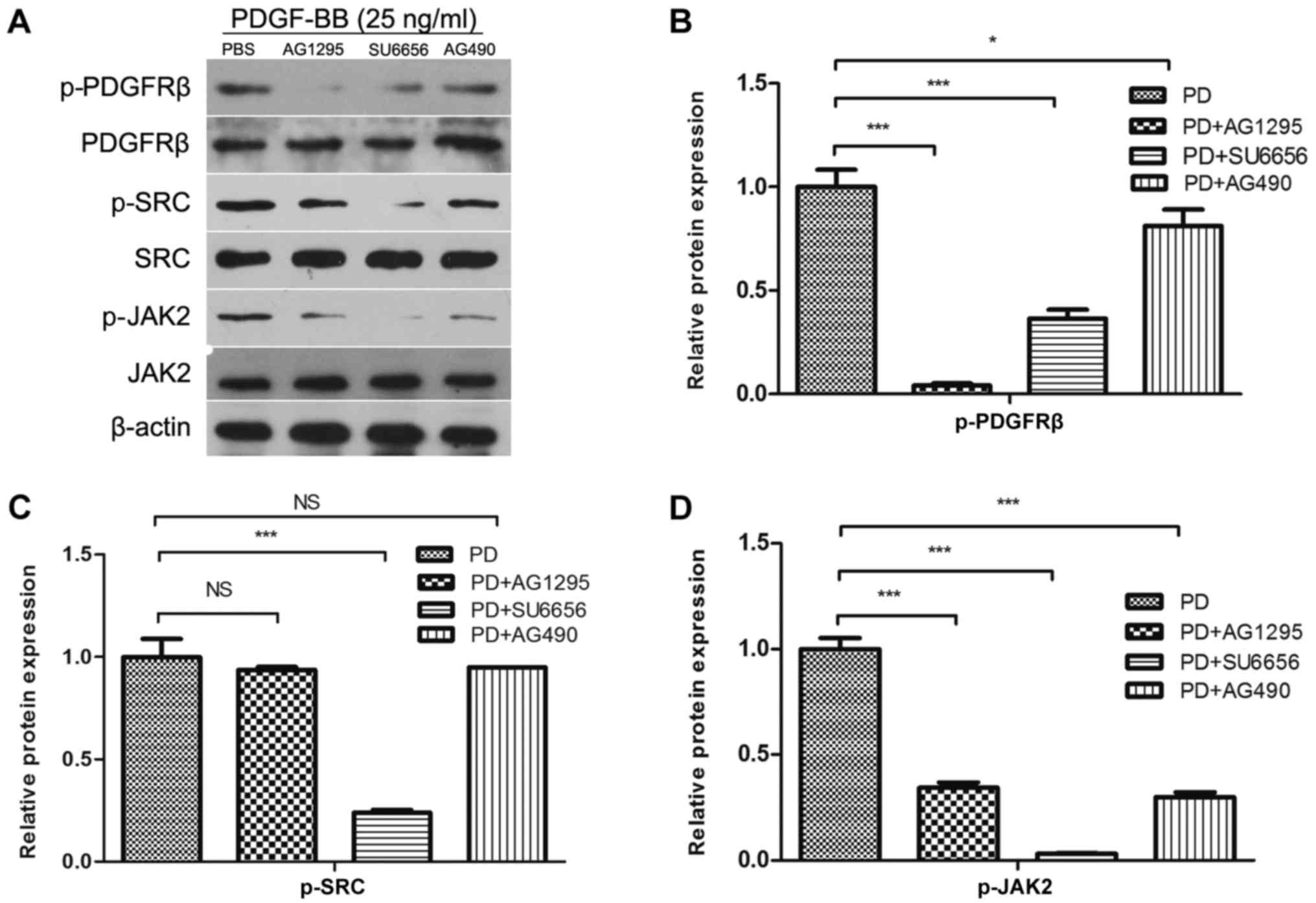 | Figure 2.Src lies in the upstream of JAK2 in
MC3T3-E1 cells. (A) MC3T3-E1 cells were serum-starved overnight and
pretreated for 30 min with inhibitors (AG1295 20 µmol/l, SU6656 2
µmol/l and AG490 10 µmol/l) as indicated, and stimulated with 25
ng/ml PDGF-BB at the same time. Densitometric analysis of the
associated bands for (B) PDGFRβ, (C) Src and (D) JAK2 were
expressed as relative optical density of the bands, corrected using
β-actin as control (normalization band). Data are presented as the
mean ± standard deviation (n=3). *P<0.05 and ***P<0.001, as
indicated. PDGF-BB, platelet-derived growth factor-BB; PDGFRβ,
PDGFR-BB receptor β; JAK2, Janus kinase 2; NS, not significant; p-,
phosphorylated. |
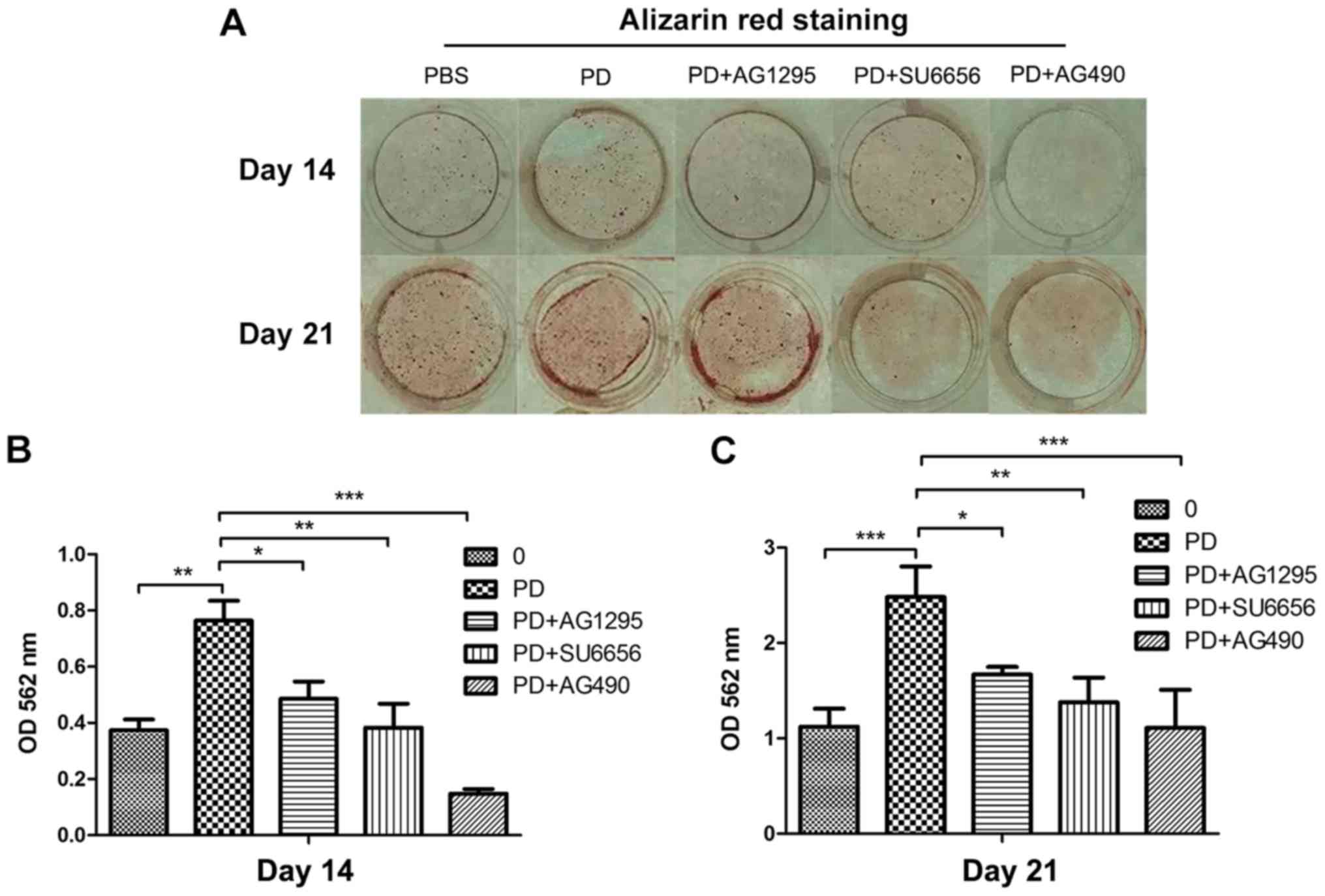
PDGF-BB promotes the differentiation
and mineralization of MC3T3-E1 cells via the Src/JAK2 signaling
pathway
In order to evaluate whether the Src/JAK2 signaling
pathway served a function in MC3T3-E1 cell differentiation and
proliferation, the effects of PDGF-BB were measured in MC3T3-E1
cells cultured in osteogenic differentiation media. As identified
with, the expression of osteoblast differentiation-associated
genes, including Runx2, a key transcription factor (15), and Osterix, ALP and Col1α1,
increased in the PDGF-BB group compared with the control, PDGF-BB +
AG1295 and PDGF-BB + SU6656 groups following culture in osteogenic
differentiation media for 3 days (Fig.
3A). The same trend was observed for the expression of
osteocalcin (OCN) and osteopontin (OPN) at 7 days (Fig. 3B). The expression levels of these
genes increased 2–5 fold. In addition, western blot analysis
indicated an increase in Runx2 and Col1α1 at 3 days (Fig. 4A and B) and 7 days (Fig. 4C and D) in the PDGF-BB group
compared with the other groups. A significantly enhanced ALP signal
was observed in the PDGF-BB group compared with the control,
PDGF-BB + AG1295, PDGF-BB + SU6656 and PDGF-BB +AG490 groups at 3
and 7 days (Fig. 5A). The Alizarin
Red staining intensity was greatly enhanced in the PGDF-BB group
compared with the other groups at 14 and 21 days (Fig. 6A-C).
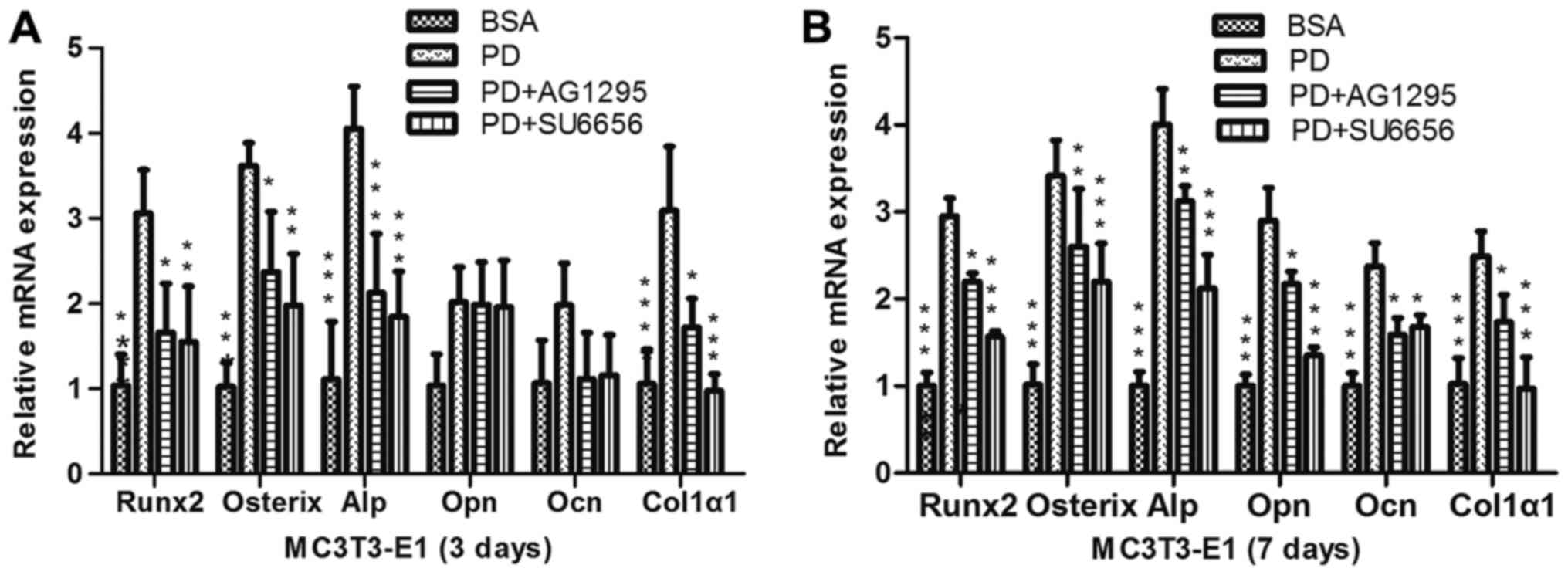 | Figure 3.PDGF-BB promotes osteogenic relative
gene expression via the Src/JAK2 signaling pathway. (A) MC3T3-E1
cells were treated with BSA (25 ng/ml), PDGF-BB (25 ng/ml), PDGF-BB
(25 ng/ml)+AG1295 (20 µmol/l) and PDGF-BB (25 ng/ml)+SU6656 (2
µmol/l), in osteogenic media for 3 days or (B) for 7 days. Data are
presented as the mean ± standard deviation (n=3). *P<0.05,
**P<0.01 and ***P<0.001, vs. PD. BSA, bovine serum albumin;
PDGF-BB, platelet-derived growth factor-BB; JAK2, Janus kinase 2;
NS, not significant; Runx2, Runt-related transcription factor 2;
Alp, alkaline phosphatase; Ocn, osteocalcin; Opn, osteopontin;
Col1α1, collagen type 1α1. |
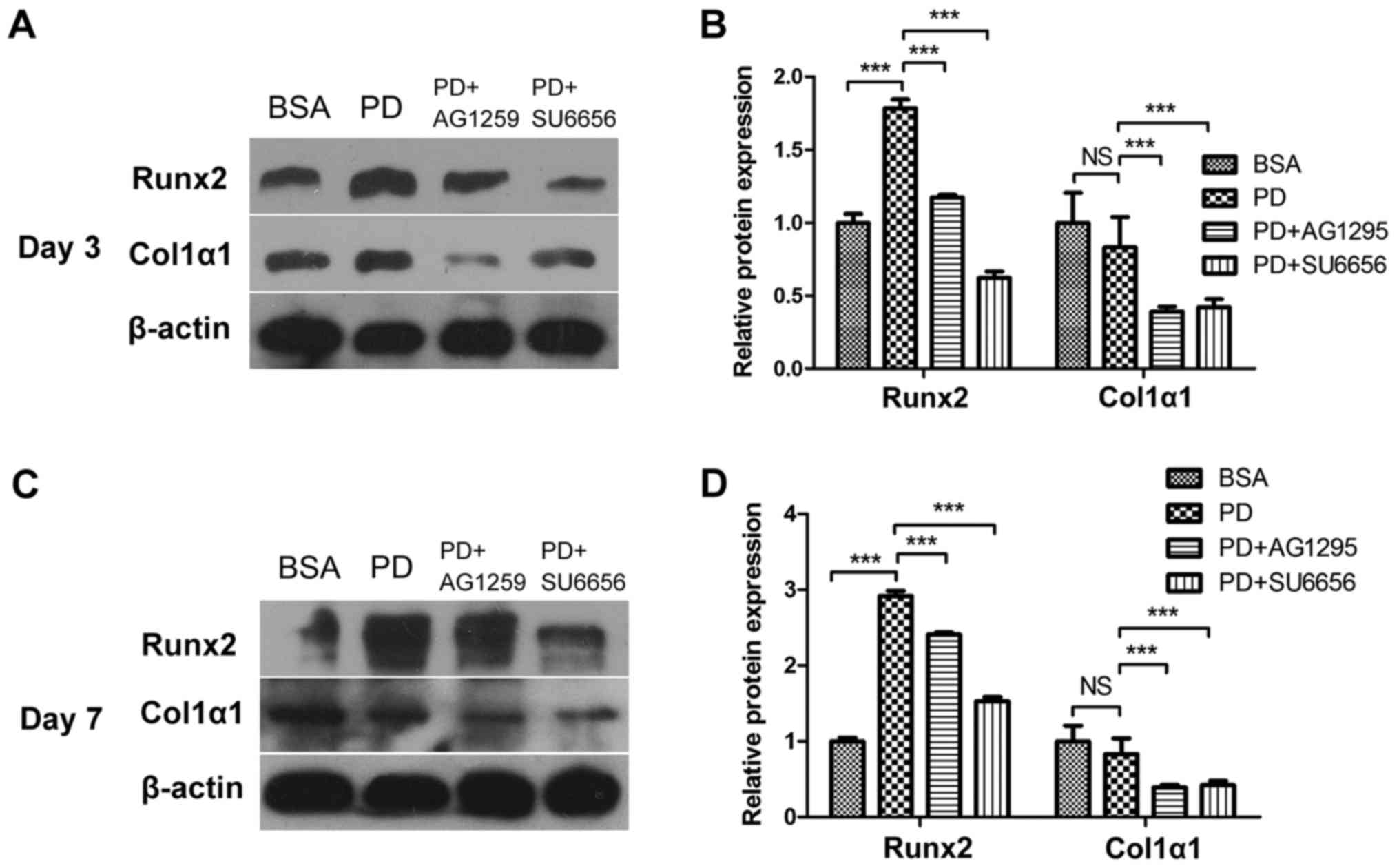 | Figure 4.PDGF-BB promotes osteogenic relative
protein expression via the Src/JAK2 signaling pathway. (A) MC3T3-E1
cells were treated with BSA (25 ng/ml), PDGF-BB (25 ng/ml), PDGF-BB
(25 ng/ml)+AG1295 (20 µmol/l) and PDGF-BB (25 ng/ml)+SU6656 (2
µmol/l), in osteogenic media for 3 days or (C) 7 days. (B and D)
Densitometric analysis of the associated bands were expressed as
the relative optical density of the bands, corrected using β-actin
as the control (normalization bands). Data are presented as the
mean ± standard deviation (n=3). ***P<0.001, as indicated.
PDGF-BB, platelet-derived growth factor-BB; JAK2, Janus kinase 2;
NS, not significant; BSA, bovine serum albumin; Runx2, Runt-related
transcription factor 2; Col1α1, collagen type 1α1. |
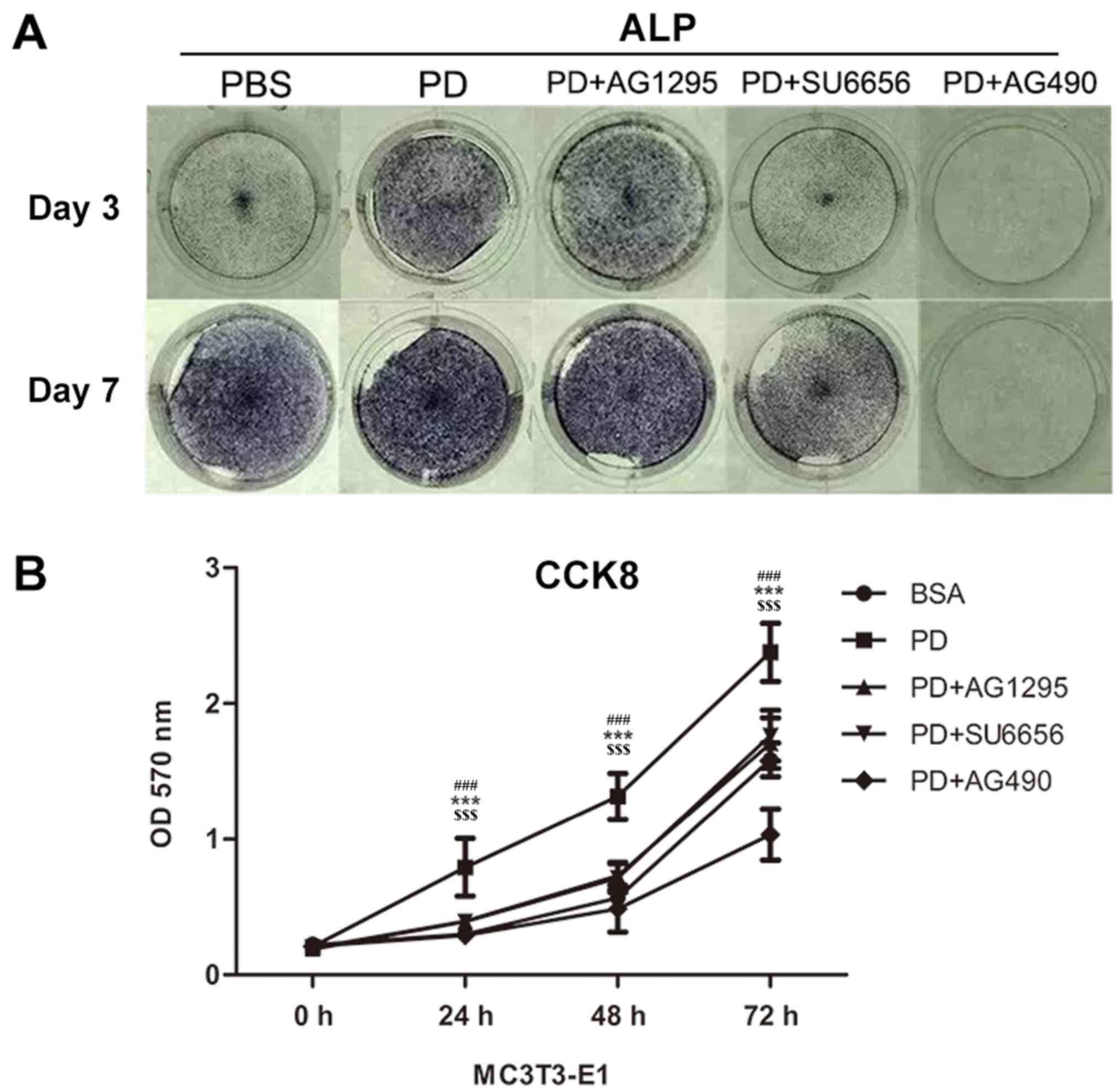 | Figure 5.PDGF-BB promotes MC3T3-E1 cell
differentiation and proliferation via the Src/JAK2 signaling
pathway. (A) Images of M3T3-E1 cells treated with BSA (25 ng/ml),
PDGF-BB (25 ng/ml), PDGF-BB (25 ng/ml)+AG1295 (20 µmol/l), PDGF-BB
(25 ng/ml)+SU6656 (2 µmol/l) and PDGF-BB (25 ng/ml)+AG490 (10
µmol/l) in osteogenic media stained by alkaline phosphatase at 3
and 7 days. (B) The OD values at 450 nm were then measured for 72
h, and the data were expressed as the mean ± standard deviation
(n=3) with six replicates. ***P<0.001 vs. PDGF-BB+AG1295;
###P<0.001 vs. PDGF-BB+SU6656;
$$$P<0.001 vs. PDGF-BB+AG490. PDGF-BB,
platelet-derived growth factor-BB; BSA, bovine serum albumin; JAK2,
Janus kinase 2; OD, optical density; CCK, Cell Counting kit; ALP,
alkaline phosphatase. |
PDGF-BB promotes the proliferation of
MC3T3-E1 cells via the Src/JAK2 signaling pathway
It was not previously determined whether PDGF-BB
promotes the proliferation of MC3T3-E1 cells through the Src/JAK2
signaling pathway. Thus, the proliferation rate of cultured
MC3T3-E1 cells was quantified with a CCK-8 assay in the present
study. It was identified that the OD570 was markedly increased in
the PDGF-BB group compared with the control, PDGF-BB + AG1295,
PDGF-BB + SU6656 and PDGF-BB + AG490 groups (Fig. 5B).
Discussion
The association between PDGF-BB and bone formation
has been widely studied. PDGF-BB serves a function in osteoblast
migration, proliferation and differentiation (4,6,8,16).
Previous studies have indicated that PDGF-BB is associated with the
apoptosis resistance of chondrocytes through the Src/PI3K/AKT
signaling pathway (17) and that
PDGF-BB induces metanephric mesenchymal cell migration by
activating Src (18). In addition,
PDGF-BB promotes pancreatic cancer cell proliferation through the
JAK2-STAT3 signaling pathway (7).
This indicates that there are interactions between PDGF-BB, Src and
JAK2.
In the present study, it was determined whether
PDGFRβ, Src and JAK2 were activated in MC3T3-E1 cells following
PDGF-BB treatment. The results indicated that PDGF-BB induced the
phosphorylation of PDGFRβ, Src and JAK2 in a time-dependent manner.
Next, it was examined whether Src was upstream of JAK2. To evaluate
this, AG1295, SU6656 and AG490, specific inhibitors of PDGFRβ, Src
and JAK2, respectively, were applied. It was identified that
SU6656, a specific inhibitor of Src, also suppressed JAK2. However,
AG490, a specific inhibitor of JAK2, did not inhibit Src. These
inhibitors were also used to determine whether PDGF-BB could
promote MC3T3-E1 cell differentiation and proliferation through the
Src/JAK2 signaling pathway. Subsequently, it was demonstrated that
SU6656 suppressed the activity of Src and JAK2, while AG490 had no
effect on Src. Therefore, we hypothesized that Src is upstream of
JAK2, which is consistent with a previous study, in which it was
demonstrated that c-Src is an upstream molecule mediating
thrombin-induced JAK2 and STAT3 activation in WI-38 cells, by the
transfection of cells with c-SrcDN (19).
There are conflicting reports regarding the ability
of PDGF-BB to promote osteogenesis. A number of studies have
indicated that PDGF-BB inhibits the osteogenic differentiation of
mesenchymal stem cells by regulating miRNA-138 (20), while others have reported that
PDGF-BB could promote host cell migration into artificial bones
without inhibiting osteoblastogenesis (21) and enhance DNA synthesis in MC3T3-E1
cells (22). In order to evaluate
the potential role of the Src/JAK2 signaling pathway in
PDGF-BB-induced osteogenesis, the effect of PDGF-BB on the
differentiation and mineralization of MC3T3-E1 cells was evaluated.
The results of qPCR indicated that PDGF-BB increased the expression
of osteogenic genes, including Runx2, Osterix, ALP, OCN, OPN and
Col1α1. In addition, SU6656, an inhibitor of Src, significantly
reduced the effect of PDGF-BB on these genes. The western blot
analysis of Runx2 and Col1α1 expression supported these results.
These findings indicated that PDGF-BB increases the expression of
osteogenic genes and proteins, and that Src and JAK2 are involved
in the process. Consistent with these results, it was also
identified that PDGF-BB enhanced ALP staining. This effect was
reduced by the suppression of Src or JAK2 signaling pathway, with
similar results for Alizarin Red staining. These data suggest that
PDGF-BB promotes the osteogenic differentiation of MC3T3-E1 cells,
and that the Src/JAK2 signaling pathway was involved in this
process.
As PDGF-BB was reported to promote the proliferation
of human osteoblasts (4), the
effects and potential mechanisms of PDGF-BB on MC3T3-E1 cells were
investigated in the current study. The results indicated that
PDGF-BB promoted MC3T3-E1 cell proliferation compared with the
control and inhibitor treatment groups. This suggested that
suppressing the Src/JAK2 signaling pathway also suppressed the
PDGF-BB induction of MC3T3-E1 cell proliferation. It was therefore
hypothesized that the Src/JAK2 signaling pathway may be involved in
the PDGF-BB-induced increase in MC3T3-E1 cell proliferation.
In addition to its effects on bone formation and
regeneration (6,23), the ability of PDGF-BB to promote
migration has also been extensively studied in osteoblasts
(16,24) and epithelial mesenchymal cells
(25). Numerous studies have
indicated that PDGF-BB may act on vascular smooth muscle cells, as
an example, to promote migration (16,26)
and proliferation (27,28), and ability of PDGF-BB to induce
angiogenesis has also been demonstrated (8,29–32).
Therefore, PDGF-BB may serve a central function in the healing of
bone fractures, as it can not only induce osteoblast
differentiation, a key factor in osteogenesis, but also vascular
smooth muscle cell migration and angiogenesis, which is critical
for the survival of new bone. This illustrates the important
function of PDGF-BB in osteogenesis and the necessity of further
research into the mechanism of PDGF-BB in inducing MC3T3-E1 cell
differentiation and proliferation.
In summary, the present study demonstrated that
PDGF-BB could promote MC3T3-E1 cell differentiation and
proliferation through the Src/JAK2 signaling pathway. This suggests
that PDGF-BB could be an important bone regulatory factor and
provides a better understanding into the molecular mechanisms of
MC3T3-E1 cells. These results may inform a novel strategy for the
treatment of patients with bone fractures or osteoporosis.
Acknowledgements
Not applicable.
Funding
The present study was supported by National Natural
Science Foundation of China (grant nos. 81470718 and 81771051) for
the design of the study, data collection and analysis, and
manuscript writing; and by Hubei Province Health and Family
Planning Scientific Research Project (grant no. WJ2017M046) for
data collection, analysis and interpretation.
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ cultured the cells, and collected, analyzed and
interpreted the data from the cell proliferation assay, histology
test and RT-qPCR. QL performed the western blotting and analyzed
the data, and was a major contributor in writing the manuscript. ZL
conceived, designed and supervised the study. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The application of the cell line MC3T3-E1 used in
the present study was approved by the Ethics Committee of School
and Hospital of Stomatology, Wuhan University (Hubei, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
PDGF-BB
|
platelet-derived growth factor-BB
|
|
PDGFRβ
|
platelet-derived growth factor-BB
receptor β
|
|
JAK2
|
Janus kinase 2
|
|
STAT
|
signal transducers and activators of
transcription
|
|
CCK8
|
Cell Counting kit-8
|
References
|
1
|
Li J: JAK-STAT and bone metabolism.
JAKSTAT. 2:e239302013.PubMed/NCBI
|
|
2
|
Luo J, Yang Z, Ma Y, Yue Z, Lin H, Qu G,
Huang J, Dai W, Li C, Zheng C, et al: LGR4 is a receptor for RANKL
and negatively regulates osteoclast differentiation and bone
resorption. Nat Med. 22:539–546. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li CH, Zhao JX, Sun L, Yao ZQ, Deng XL,
Liu R and Liu XY: AG490 inhibits NFATc1 expression and STAT3
activation during RANKL induced osteoclastogenesis. Biochem Biophys
Res Commun. 435:533–539. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vordemvenne T, Paletta JR, Hartensuer R,
Pap T, Raschke MJ and Ochman S: Cooperative effects in
differentiation and proliferation between PDGF-BB and matrix
derived synthetic peptides in human osteoblasts. BMC Musculoskelet
Disord. 12:2632011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Takai S, Matsushima-Nishiwaki R, Adachi S,
Natsume H, Minamitani C, Mizutani J, Otsuka T, Tokuda H and Kozawa
O: (−)-Epigallocatechin gallate reduces platelet-derived growth
factor-BB-stimulated interleukin-6 synthesis in osteoblasts:
Suppression of SAPK/JNK. Mediators Inflamm. 2008:2918082008.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Caplan AI and Correa D: PDGF in bone
formation and regeneration: New insights into a novel mechanism
involving MSCs. J Orthop Res. 29:1795–1803. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Masamune A, Satoh M, Kikuta K, Suzuki N
and Shimosegawa T: Activation of JAK-STAT pathway is required for
platelet-derived growth factor-induced proliferation of pancreatic
stellate cells. World J Gastroenterol. 11:3385–3391. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xie H, Cui Z, Wang L, Xia Z, Hu Y, Xian L,
Li C, Xie L, Crane J, Wan M, et al: PDGF-BB secreted by
preosteoclasts induces angiogenesis during coupling with
osteogenesis. Nat Med. 20:1270–1278. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lennartsson J, Ma H, Wardega P, Pelka K,
Engström U, Hellberg C and Heldin CH: The Fer tyrosine kinase is
important for platelet-derived growth factor-BB-induced signal
transducer and activator of transcription 3 (STAT3) protein
phosphorylation, colony formation in soft agar, and tumor growth in
vivo. J Biol Chem. 288:15736–15744. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rucci N, Susa M and Teti A: Inhibition of
protein kinase c-Src as a therapeutic approach for cancer and bone
metastases. Anticancer Agents Med Chem. 8:342–349. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ishizawar R and Parsons SJ: c-Src and
cooperating partners in human cancer. Cancer Cell. 6:209–214. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Waibel M, Solomon VS, Knight DA, Ralli RA,
Kim SK, Banks KM, Vidacs E, Virely C, Sia KC, Bracken LS, et al:
Combined targeting of JAK2 and Bcl-2/Bcl-xL to cure mutant
JAK2-driven malignancies and overcome acquired resistance to JAK2
inhibitors. Cell Rep. 5:1047–1059. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mori K, Blanchard F, Charrier C, Battaglia
S, Ando K, Duplomb L, Shultz LD, Redini F and Heymann D:
Conditioned media from mouse osteosarcoma cells promote MC3T3-E1
cell proliferation using JAKs and PI3-K/Akt signal crosstalk.
Cancer Sci. 99:2170–2176. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ge X, Chen SY, Liu M, Liang TM and Liu C:
Evodiamine inhibits PDGF-BB-induced proliferation of rat vascular
smooth muscle cells through the suppression of cell cycle
progression and oxidative stress. Mol Med Rep. 14:4551–4558. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wei J, Shimazu J, Makinistoglu MP, Maurizi
A, Kajimura D, Zong H, Takarada T, Lezaki T, Pessin JE, Hinoi E and
Karsenty G: Glucose uptake and Runx2 synergize to orchestrate
osteoblast differentiation and bone formation. Cell. 161:1576–1591.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Colciago A, Celotti F, Casati L, Giancola
R, Castano SM, Antonini G, Sacchi MC and Negri-Cesi P: In vitro
effects of PDGF isoforms (AA, BB, AB and CC) on migration and
proliferation of SaOS-2 osteoblasts and on migration of human
osteoblasts. Int J Biomed Sci. 5:380–389. 2009.PubMed/NCBI
|
|
17
|
Montaseri A, Busch F, Mobasheri A,
Buhrmann C, Aldinger C, Rad JS and Shakibaei M: IGF-1 and PDGF-bb
suppress IL-1β-induced cartilage degradation through
down-regulation of NF-κB signaling: Involvement of Src/PI-3K/AKT
pathway. PLoS One. 6:e286632011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wagner B and Gorin Y: Src tyrosine kinase
mediates platelet-derived growth factor BB-induced and
redox-dependent migration in metanephric mesenchymal cells. Am J
Physiol Renal Physiol. 306:F85–F97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bai KJ, Chen BC, Pai HC, Weng CM, Yu CC,
Hsu MJ, Yu MC, Ma HP, Wu CH, Hong CY, et al: Thrombin-induced CCN2
expression in human lung fibroblasts requires the c-Src/JAK2/STAT3
pathway. J Leukoc Biol. 93:101–112. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qu B, Xia X, Wu HH, Tu CQ and Pan X:
PDGF-regulated miRNA-138 inhibits the osteogenic differentiation of
mesenchymal stem cells. Biochem Biophys Res Commun. 448:241–247.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yoshida S, Iwasaki R, Kawana H, Miyauchi
Y, Hoshi H, Miyamoto H, Mori T, Kanagawa H, Katsuyama E, Fujie A,
et al: PDGFBB promotes PDGFRα-positive cell migration into
artificial bone in vivo. Biochem Biophys Res Commun. 421:785–789.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tsukamoto T, Matsui T, Fukase M and Fujita
T: Platelet-derived growth factor B chain homodimer enhances
chemotaxis and DNA synthesis in normal osteoblast-like cells
(MC3T3-E1). Biochem Biophys Res Commun. 175:745–751. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen W, Baylink DJ, Brier-Jones J, Neises
A, Kiroyan JB, Rundle CH, Lau KH and Zhang XB: PDGFB-based stem
cell gene therapy increases bone strength in the mouse. Proc Natl
Acad Sci USA. 112:pp. E3893–E3900. 2015; View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hengartner NE, Fiedler J, Ignatius A and
Brenner RE: IL-1β inhibits human osteoblast migration. Mol Med.
19:36–42. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang L, Lin C and Liu ZR: P68 RNA helicase
mediates PDGF-induced epithelial mesenchymal transition by
displacing Axin from beta-catenin. Cell. 127:139–155. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kang H, Ahn DH, Pak JH, Seo KH, Baek NI
and Jang SW: Magnobovatol inhibits smooth muscle cell migration by
suppressing PDGF-Rβ phosphorylation and inhibiting matrix
metalloproteinase-2 expression. Int J Mol Med. 37:1239–1246. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Park S, Kim JK, Oh CJ, Choi SH, Jeon JH
and Lee IK: Scoparone interferes with STAT3-induced proliferation
of vascular smooth muscle cells. Exp Mol Med. 47:e1452015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yan G, Wang Q, Hu S, Wang D, Qiao Y, Ma G,
Tang C and Gu Y: Digoxin inhibits PDGF-BB-induced VSMC
proliferation and migration through an increase in ILK signaling
and attenuates neointima formation following carotid injury. Int J
Mol Med. 36:1001–1011. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rykala J, Przybylowska K, Majsterek I,
Pasz-Walczak G, Sygut A, Dziki A and Kruk-Jeromin J: Angiogenesis
markers quantification in breast cancer and their correlation with
clinicopathological prognostic variables. Pathol Oncol Res.
17:809–817. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Drela E, Kulwas A, Jundziłł W, Góralczyk
B, Boinska J, Drewniak W, Gadomska G and Rość D: VEGF-A and
PDGF-BB-angiogenic factors and the stage of diabetic foot syndrome
advancement. Endokrynol Pol. 65:306–312. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Han H, Cao FL, Wang BZ, Mu XR, Li GY and
Wang XW: Expression of angiogenesis regulatory proteins and
epithelial-mesenchymal transition factors in platelets of the
breast cancer patients. ScientificWorldJournal. 2014:8782092014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yin T, He S, Su C, Chen X, Zhang D, Wan Y,
Ye T, Shen G, Wang Y, Shi H, et al: Genetically modified human
placenta-derived mesenchymal stem cells with FGF-2 and PDGF-BB
enhance neovascularization in a model of hindlimb ischemia. Mol Med
Rep. 12:5093–5099. 2015. View Article : Google Scholar : PubMed/NCBI
|