Introduction
Cancer is a leading global cause of mortality that
accounts for almost 13% of mortality worldwide (1). According to Globocan
(globocan.iarc.fr/Default.aspx), it is estimated that by 2020,
there will be between 15 and 17 million newly diagnosed cases of
cancer every year, 60% of which will be in developing countries
(1). Asia accounts for 60% of the
world population and half the global burden of cancer. With current
resources, one-third of cancers could be preventable, and another
two-third of newly diagnosed cancer patients could experience
increased survival or early-stage detection (1). Prevention as well as early detection
of cancers leads to both better health outcomes and considerable
savings in treatment costs (2).
Despite this, progress in global cancer control has been slow,
largely due to the weak and fragmented nature of both the global
and national responses to treatment.
Esophageal cancer (EC) is one of the most common
diseases in gastroenterology, and additionally one of the most
fatal (3). EC is the fourth most
common gastrointestinal cancer in the United States of America with
a <20% 5-year survival (4). The
incidence of esophageal adenocarcinoma has risen dramatically over
the past two decades (3). EC also
ranks fourth among cancer-associated mortalities in China (5). Surgical resection is the main
treatment method for EC, with a postoperative 5-year survival rate
of only 34–36% (6). Conventional
treatment strategies including neoadjuvant radiochemotherapy,
preoperative neoadjuvant chemotherapy, and three-field lymph node
dissection are helpful for improving the 5-year survival rate of EC
patients. However, the outcomes are still unsatisfactory, and local
recurrence and distant metastasis results in patient mortality
(7). Therefore, biomarkers can be
used to identify the recurrence or metastasis of EC in order to
facilitate timely diagnosis and treatment strategies, thus
improving the prognosis of EC patients.
MicroRNA (miRNA/miR) is a class of widely
distributed non-coding RNA of 20–22 nucleotides, usually serving a
role in regulation of post-transcriptional gene expression
(8). The biogenesis of miRNA
involves a complex protein system, including members of the
Argonaute family, Polymerase II-dependent transcription and the
RNase III Drosha and Dicer 13 (9).
MiRNA is generally transcribed as a primary transcript (pre-miRNA)
through Polymerase II and is processed to a 70-nucleotide stem-loop
by the Microprocessor (Drosha/Pasha). The resulting pre-miRNA is
transported into the cytoplasm via Exportin-5. Dicer proceeds to
cleave the pre-miRNA to produce a mature miRNA: miRNA duplex. TAR
RNA binding protein/Loquacious is used to bind the miRNA to
Argonaute and RNA-induced silencing complex is formed, then, the
miRNA strand is degraded (8).
MiRNA has been demonstrated to be involved in multiple biological
processes, such as cell differentiation, proliferation,
oncogenesis, angiogenesis, tumor invasion and tumor metastasis. It
has been demonstrated that miRNA serves a pivotal role in human
cancer cell growth, invasion and migration (10). Furthermore, miRNA expression is
associated with a variety of cancers, and miRNA may function as
‘oncogenes’ or ‘tumor suppressor genes’. Approximately 50% of miRNA
is located on the genome at the fragile sites associated with the
tumor, indicating that miRNA serves a crucial role in the
development and progression of tumors.
MiR-675 is derived from long non-coding RNA H19, an
miRNA embedded within H19's first exon, and H19 can generate two
mature microRNAs, miRNA-675-3p and miRNA-675-5p in a Drosha and
Dicer splicing dependent manner. MiRNA-675 is expressed exclusively
in the placenta from the gestational time point when placental
growth normally ceases, but placentas that lack H19 continue to
grow. The main physiological role of H19 is inhibiting the growth
of the placenta prior to birth, by regulating processing of miR-675
(11,12). The inverse association between
miRNA-675 and placenta growth indicates that miRNA-675 can act as a
growth restrictor during embryonic development (11). MiRNA-675 represses the expression
of tumor suppressor retinoblastoma in a classical way and promotes
the proliferation of colon cancer cells (13). In addition, miRNA-675-5p is
expressed in EC and its over-expression in some esophageal squamous
cancer cell lines result in their proliferation and migration. In
the present study, the expression of miR-675-3p was investigated in
a healthy esophageal epithelial cell line (HECC), and its function
on esophageal squamous cell cancer (ESCC) proliferation and
migration was evaluated.
Materials and methods
Ethics statement
Written informed consent was obtained from all
participants, and the study protocol was approved by the ethics
committee of JiangSu Cancer Hospital (Nanjing, China).
Clinical samples and cell lines
ESCC and paired normal esophageal tissues (>5 cm
away from the tumor margin) were obtained from 35 patients (median
age 63.7 years, range 48–76 years, female:male=1:6) who underwent
esophagus resection between September 2016 and June 2017 at JiangSu
Cancer Hospital. All patients had no history of previous
malignancies, chemotherapy and radiotherapy. All the samples were
immediately snap-frozen in liquid nitrogen and stored at −80°C for
RNA extraction. All tumors and paired normal tissues were
histologically confirmed by two pathologists. Human ESCC cell lines
(Te-1 and Kyse-150) were obtained from Chinese Academy of Science
cell bank (Shanghai, China), Human Healthy Esophageal Epithelial
cell line (HECC) were obtained from iCell Bioscience Inc (Shanghai,
China). Cells were cultured and maintained in RPMI 1640 (HyClone;
GE Healthcare, Chicago, IL, USA) supplemented with 10% fetal bovine
serum (FBS, Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) in an incubator with 5% CO2 at 37°C. All culture
materials were purchased from Gibco; Thermo Fisher Scientific,
Inc.
Mir-675-3p overexpression and
interference oligonucleotide transfection assay
Kyse-150 and Te-1 cells were seeded at a density of
5×104 cells/ml in six-well plates and incubated
overnight at 37°C. According to the protocol, they were transiently
transfected with the 20 µmol interference oligonucleotide which
included negative control (NC; 5′-3′ CAGUACUUUUGUGUAGUACAA),
mir-675-3p inhibitor (5′-3′ UGAGCGGUGAGGGCAAUACAG) or mir-675-3p
mimic (sense 5′-3′ CUGUAUGCCCUCACCGCUCA; anti-sense 5′-3′
AGCGGUGAGGGCAUACAGUU) using Lipofectamine® 2000 reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). Following 6 h
transfection at 37°C, the medium was replaced with RPMI1640 media
containing 10% FBS. Three group were used in this experiment, which
included negative control (NC), miR-675-3p inhibitor group and
miR-675-3p inhibitor + miR-675-3p mimic. The group of NC and
miR-675-3p inhibitor were only transfected with NC or miR-675-3p
inhibitor, respectively, and the group of miR-675-3p inhibitor +
miR-675-3p mimic, were transfected with miR-675-3p mimics 6 h
following transfection with miR-675-3p inhibitor.
Total RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
At 24 h post-transfection, cells were treated with
TRIzol (Takara Bio, Inc., Otsu, Japan) to extract total RNA, which
was reverse transcribed to first strand cDNA using Primescript RT
Reagent (Takara Bio, Inc.). The expression levels of miR-675-3p
(sense 5′-CGGAGAGGGCCCACAGTG-3′; anti-sense was an universal primer
which was obtained from GeneCopoeia, USA), MMP2 (sense
5′-GCTGACGGTAAGGACGGACTC-3′; anti-sense
5′-CGTTGCCATTGAACAAGAAGG-3′), MMP9 (sense
5′-TGTGCTACAGGGAGAGATAAGA-3′; anti-sense
5′-GTGGGTGGAGCAGAGTAAATAA-3′) and E-cadherin (sense
5′-CTCAGTTGGAACAGGGTGAATA-3′; anti-sense
5′-GTGCAGGACACTCAAATCAAAG-3′) were examined using RT-qPCR in a Step
one plus system (Roche Molecular Diagnostics, Pleasanton, CA, USA)
using 2−ΔΔCq analysis. In this system, all reactions
were performed in SybrGreen (ChamQ™ SYBR qPCR Master
Mix; Vazyme, Piscataway, NJ, USA), in a volume of 20 ul containing
2 ul cDNA, according to the manufacturer's protocol. All reactions
were performed in triplicate. The thermal cycling conditions
included a step of 5 min at 95°C followed by 40 cycles of 95°C for
10 sec and 60°C for 30 sec. U6 was used as internal control. Final
amounts of target in cells were determined as follows: Target
amount=2−ΔΔCq, where ΔΔCq=[Cq (target)-Cq
(U6)]sample-[Cq (target)-Cq (U6)]internal standard. Final amounts
of target in tissues were determined as target
amount=2−ΔΔCq (14).
Cell proliferation assay
Cell proliferation was monitored by a colorimetric
assay using the Cell Counting Kit-8 (CCK-8, Vazyme) according to
the manufacturer's protocol. Briefly, cells were seeded on 6-well
plates and transfected with NC, miR-675-3p inhibitor or miR-675-3p
inhibitor + miR-675-3p mimics, after 12 h of transfection, the
cells were transferred onto the 96-well plates (8×103
cells/well) and the cell proliferation assays were conducted and
analyzed by a Microplate Reader (ELx808; BIO-TEK Instruments, Inc.,
Winooski, VT, USA) every 12 h for 3 days using CCK-8.
Scratch wound assay
Cells (3×105) were transfected with NC,
miR-675-3p inhibitor or miR-675-3p inhibitor + miR-675-3p mimics
and cultured in a cell culture dish (35×10 mm). Cell layers were
scratched using a pipette tip to form wound gaps. The wound
location in the cell culture dish was marked. Cells were imaged to
record the wound width (0 h). A total of 12 h following this,
images were taken again at the marked wound location to measure the
cell migration ability.
Transwell invasion and migration
ability assay
Cells (3×105) were transfected with NC,
miR-675-3p inhibitor or miR-675-3p inhibitor + miR-675-3p mimics
according to the protocol. Following incubation for 12 h, Kyse-150
cells or Te-1 cells were transferred to the upper Matrigel-coated
or not Matrigel-coated invasion chambers (BD Biosciences, San Jose,
CA, USA) in a serum-free RPMI 1640, and RPMI 1640 containing 10%
FBS was added to the lower chambers. Following 24 h, non-migrated
or non-invaded cells on the upper surface were removed, and the
migrating or invading cells on the underside surface were fixed
with 4% paraformaldehyde, stained with 0.1% crystal violet at room
temperature for 15 min, and imaged. A total of three independent
experiments were performed.
ELISA
Kyse-150 cells (3×105) or Te-1 cells
(3×105) were cultured in six-well plates and following
24 h transfection with NC, miR-675-3p inhibitor or miR-675-3p
inhibitor + miR-675-3p mimics, the related proteins in the cell
culture were extracted by centrifuging at 300 × g for 10 min at 4°C
following the manufacturer's protocol. The protein concentrations
of matrix metalloproteinase (MMP)2, MMP9 and E-cadherin were
measured by ELISA assay (Hangzhou Multisciences, Biotech Co., Ltd,
Zhejiang, China, catalog number: EK1M022-96T, EK1M092-96T,
EK12352-96T, respectively) following the manufacturer's protocol.
All samples were measured in duplicate.
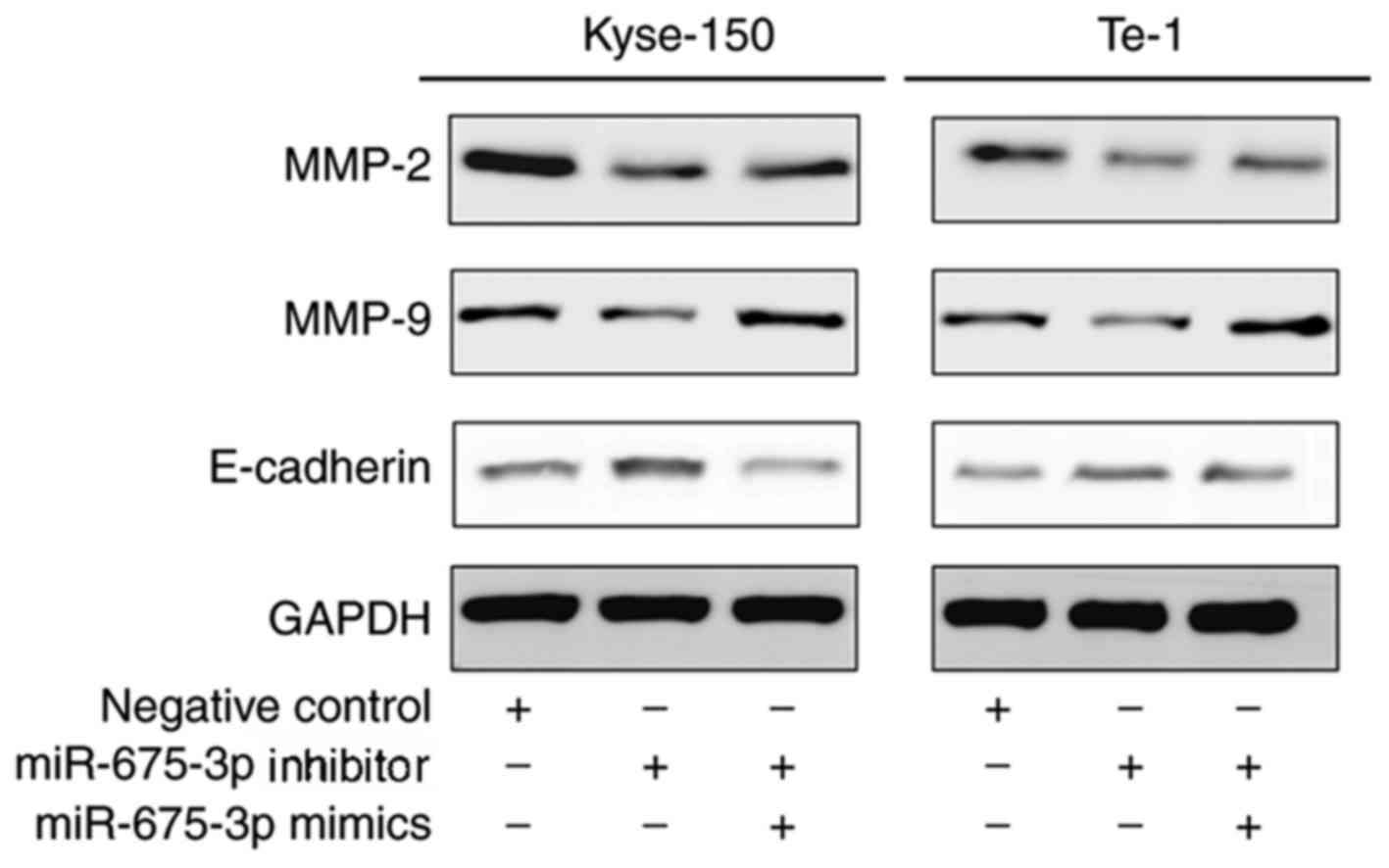
Western blot analysis
Protein levels of MMP2, MMP9 and E-cadherin were
detected by western blotting 24 h following treatment. In brief,
the total cell proteins were extracted from cells using lysis
buffer (Cell Signaling Technology Inc., Danvers, MA, USA). BCA
assay (Thermo Fisher Scientific, Inc.) was carried out to measure
the protein concentrations. Equal amounts of protein samples (25
µg) were separated by 12% SDS-PAGE and then transferred onto
polyvinylidene difluoride membranes. Following this, the membranes
were blocked with 5% skim milk at room temperature for 2 h.
Subsequently, incubation occurred with the primary antibodies
against MMP2 (cat. no. 13132), MMP9 (cat. no. 2270) or E-cadherin
(cat. no. 3195; all 1:1,000; all Cell Signaling Technology Inc.)
overnight at 4°C. The membrane was then incubated with secondary
antibodies (HRP-conjugated goat Anti-Rabbit, cat. no. Ab203-01,
Vazyme, Piscataway, NJ, USA) at room temperature for 2 h. GAPDH
(1:5,000; cat. no. 8884, Cell Signaling Technology Inc.) served as
the internal control. Finally, to visualize the protein bands, an
enhanced chemiluminescence detection system (Super Signal West Dura
Extended Duration Substrate; Thermo Fisher Scientific, Inc.) was
used.
Statistical analysis
Data were analyzed by GraphPad Prism5 (GraphPad
Software, Inc., La Jolla, CA, USA) and were expressed as the mean ±
standard deviation. Comparisons between groups were made using
Student's t-test or one-way analysis of variance followed by
Tukey's test. P<0.05 was considered to indicate a statistically
significant difference.
Results
MiR-675-3p expression in ESCC
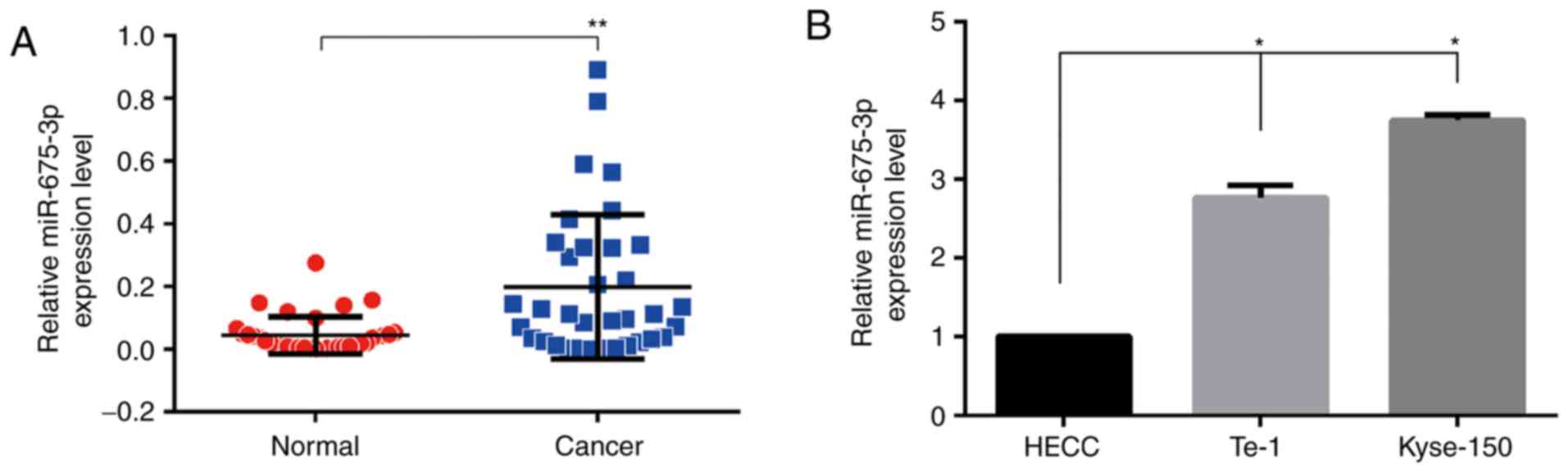
To explore the expression of miR-675-3p in ESCC
tissues, RT-qPCR was used. As demonstrated in Fig. 1A, compared with the normal tissues,
cancer tissues expressed higher levels of miR-675-3p, suggesting
that the expression level of miR-675-3p was upregulated in ESCC
tissues. To further investigate the functional effects of
miR-675-3p in ESCC cells, RT-qPCR was also performed to investigate
miR-675-3p expression level in human ESCC cell lines (Kyse-150 and
Te-1) and the HECC. As demonstrated in Fig. 1B, compared with HECC, both ESCC
cell lines expressed higher levels of miR-675-3p.
Knockdown of miR-675-3p inhibits ESCC
cell growth in vitro
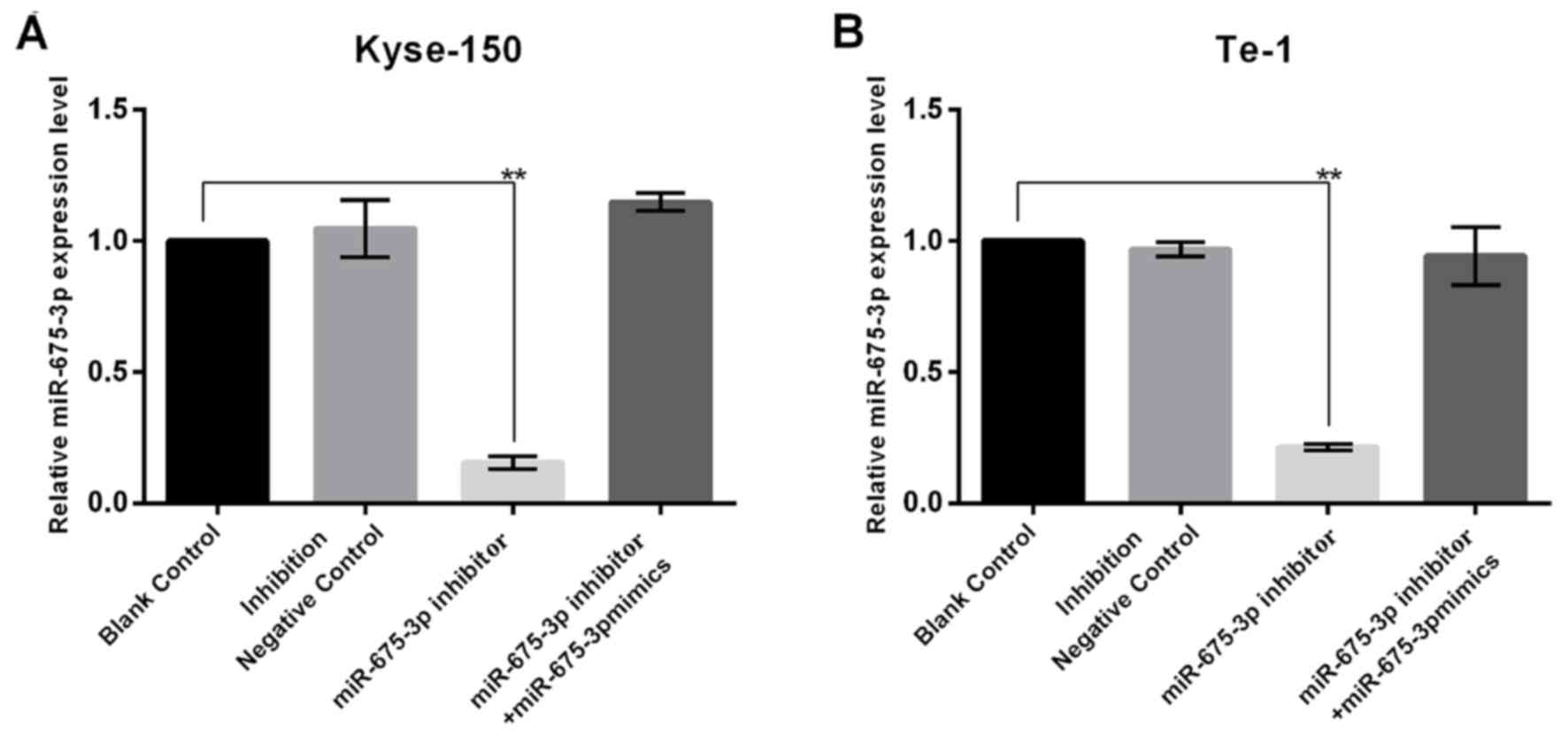
In order to investigate the influence of miR-675-3p
on ESCC cell growth, the present study firstly transfected NC,
miR-675-3p inhibitor, or miR-675-3p inhibitor+miR-675-3p mimics
into cells, and the transfection efficiency was detected using
RT-qPCR. The results indicated that miR-675-3p inhibitor
significantly decreased miR-675-3p expression in KYSE150 and Te-1
cells and this reduction was reversed by miR-675-3p mimics
(Fig. 2). In order to investigate
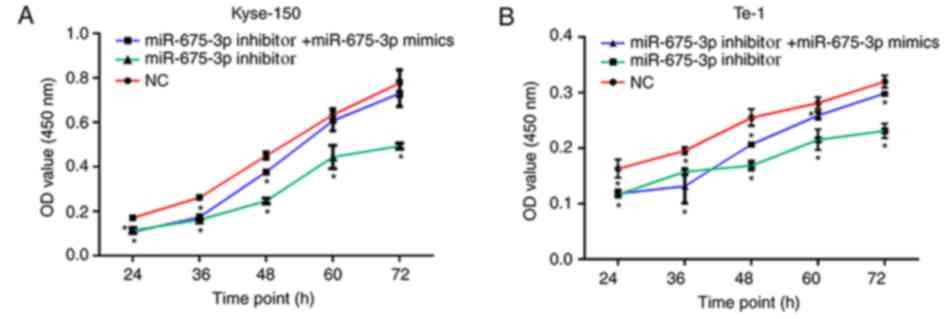
the influence of miR-675-3p on ESCC cell growth, the CCK-8 assay
was conducted. The results demonstrated that the proliferation and
viability of Kyse-150 and Te-1 cells transfected with miR-675-3p
inhibitor was significantly impaired compared with the negative
control (NC), indicating an inhibitory effect. However,
co-transfection with miR-675-3p inhibitor and miR-675-3p mimics
eliminated this effect on Kyse-150 and Te-1 cells, and the
proliferation and viability approached the level of the negative
control (Fig. 3).
Knockdown of miR-675-3p inhibits ESCC
cell migration and invasion
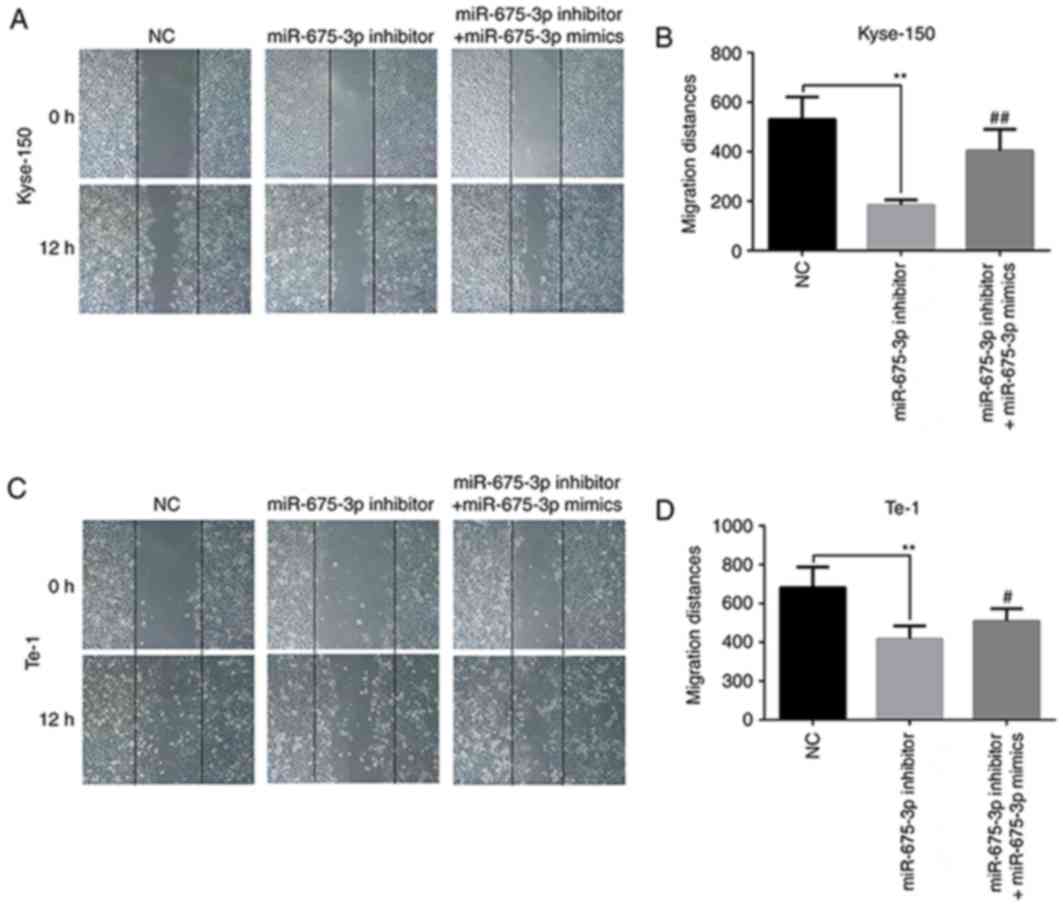
The effect of miR-675-3p on the migration of ESCC
cells was first checked by a wound healing assay. The results of
the in vitro wound healing assay demonstrated that compared
with the negative control cells, the migration of Kyse-150 and Te-1
cells was significantly attenuated by miR-675-3p inhibitor. When
the levels of miR-675-3p were increased by the miR-675-3p mimics,
the migration of Kyse-150 and Te-1 cells recovered to the level of
negative control group (Fig.
4).
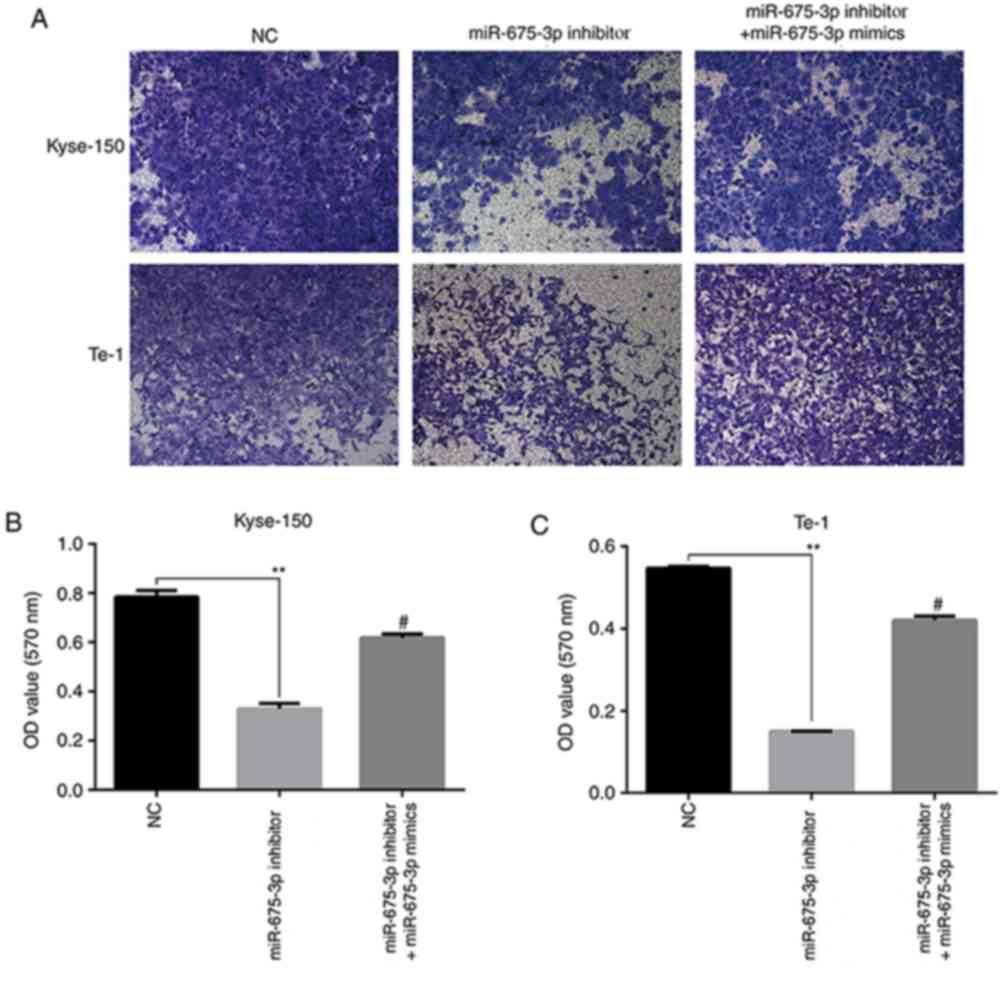
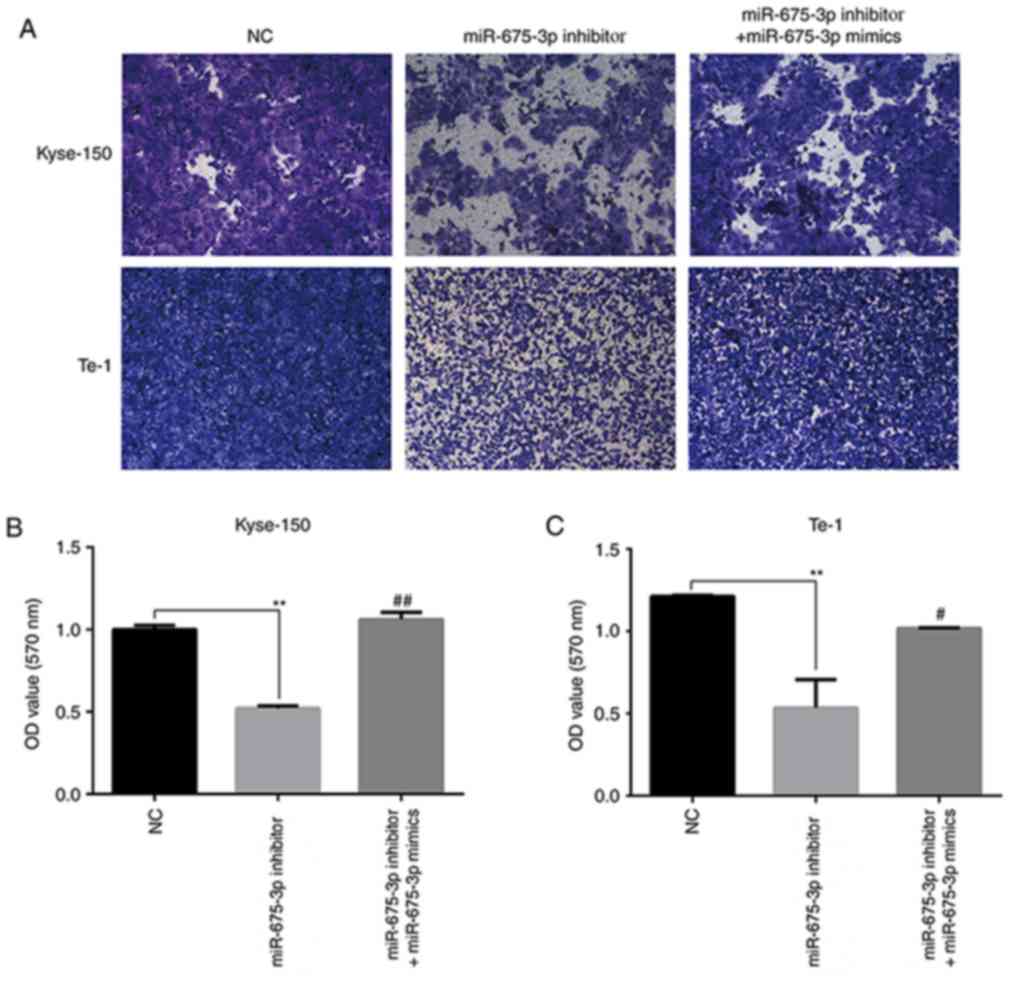
To further verify the effects of miR-675-3p on cell
migration and invasion ability in ESCC, a transwell assay was
performed. An in vitro migration assay revealed that the
migration ability of Kyse-150 and Te-1 cells transfected with
miR-675-3p inhibitor were suppressed compared with the negative
control, but co-transfection with miR-675-3p inhibitor and
miR-675-3p mimics eliminated this effect on Kyse-150 and Te-1 cells
(Fig. 5). Similarly, as presented
in Fig. 6, compared with the
negative control, downregulation of miR-675-3p effectively
repressed the invasion capacity of Kyse-150 and Te-1 cells, however
co-transfection with miR-675-3p inhibitor and miR-675-3p mimics
resulted in migration and invasion capacities that approached the
level of the negative control group (Figs. 5 and 6, respectively). These data may indicate
the oncogenic role of miR-675-3p via the effects on the migration
and invasion of ESCC.
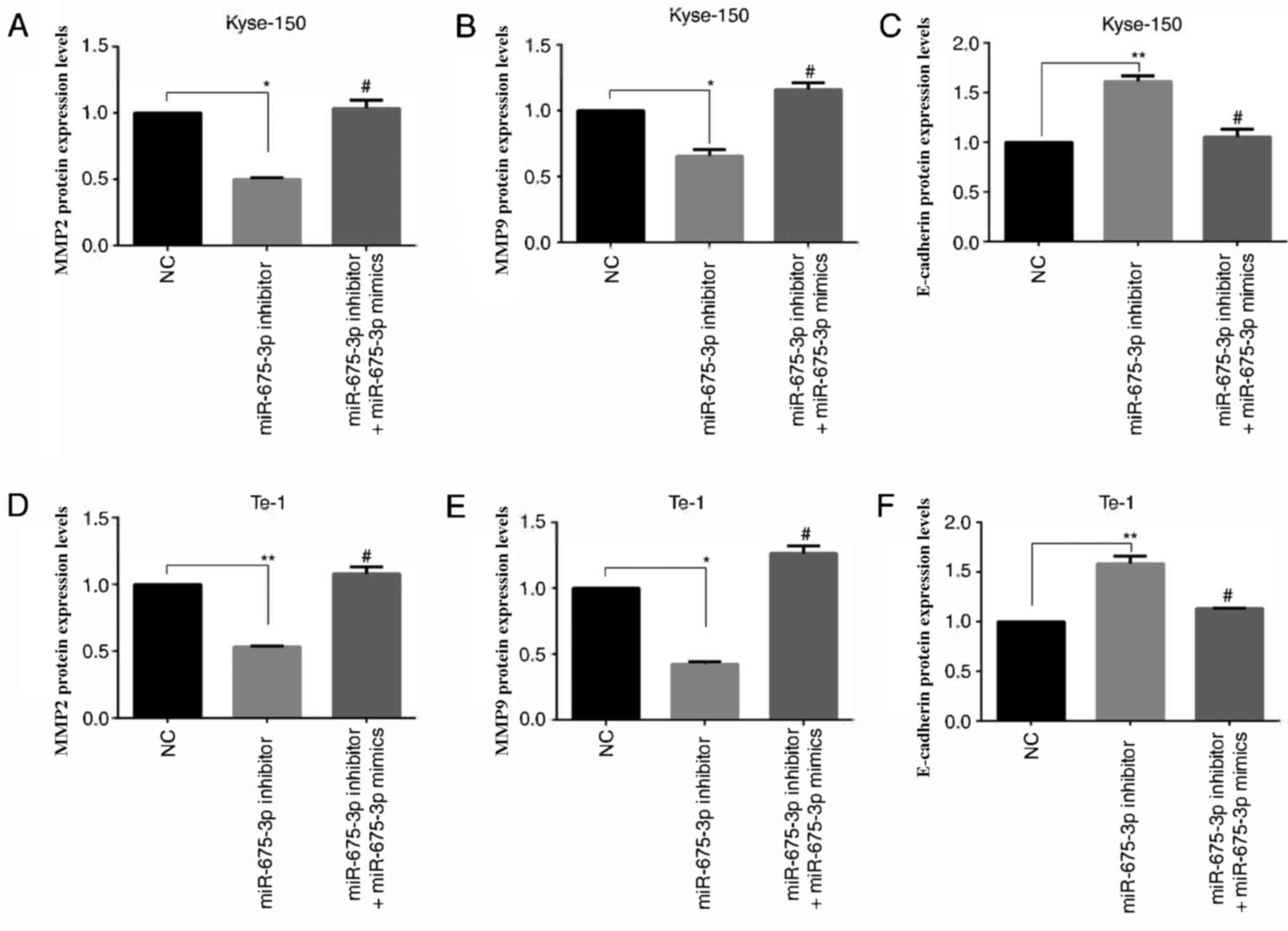
Effect of miR-675-3p on MMP2, MMP9 and
E-cadherin expression in ESCC cell lines
MMP2 (15,16) and MMP9 (17,18)
are involved in many events, such as cancer progression, and
invasion, indicating that they may influence the invasion ability
of cells. E-cadherin, is a calcium-dependent cell adhesion
molecule. Loss of E-cadherin function or expression has been
implicated in cancer progression and metastasis (19–21).
E-cadherin downregulation decreases the strength of cellular
adhesion within a tissue, resulting in an increase in cellular
motility (22–24). This in turn may allow cancer cells
to cross the basement membrane and invade surrounding tissues.
Therefore, expression levels of MMP2, MMP9 and E-cadherin were
analyzed by ELISA (Fig. 7) and
western blot analysis (Fig. 8).
Compared with the negative control, miR-675-3p inhibitor
significantly decreased MMP2 and MMP9 expression whereas E-cadherin
was enhanced. Co-transfection with miR-675-3p inhibitor and
miR-675-3p mimics reversed these effects in Kyse-150 and Te-1 cells
(Figs. 7 and 8).
Discussion
Although great progress regarding the role of miRNA
in cancer pathogenesis has been made, the roles of miRNA in the
carcinogenesis of ESCC are far from being adequately elucidated. In
the present study, miR-675-3p was demonstrated to serve an
important role in promoting the migration and invasion ability of
ESCC cell lines through inhibiting or activating epithelial
mesenchymal transition (EMT) marker expression levels. In the
beginning of the present study, the expression of miR-675-3p was
upregulated in ESCC tissues and cell lines compared with the normal
control. Therefore, the high level of miR-675-3p may be associated
with the progression and development of tumor and the migration and
invasion ability of ESCC cells. Previous studies have highlighted
the prospect that certain miRNAs may be used as biomarkers for
prognosis assessment or tumor therapeutic targets in human cancer
(25–27). MiRNAs are known for their dual role
as oncogenes (28,29) or tumor suppressors (30–33)
and they also have been implicated in the regulatory network of
various cancer types.
MiR-675-5p has been demonstrated to affect tumor
migration or invasion. He et al (34) suggested that miR-675-5p is
downregulated in non-small cell lung cancer tissues compared with
normal tissues, and miR-675-5p inhibition could promote the
migration and invasion ability of non-small cell lung cancer cells.
Zhou et al (35)
demonstrated that downregulation of miR-675-5p inhibits the
migration and invasion ability of the ESCC cells. In the present
study, it was demonstrated that miR-675-3p was highly expressed in
ESCC tumor tissues compared with the normal tissues. Therefore, it
may be hypothesized that miR-675-3p as another variant of miR-675
may also have the biological activity which could affect the
migration and invasion ability of ESCC cells. As demonstrated in
the present results, miR-675-3p affected the migration and invasion
abilities of ESCC cells. When miR-675-3p was downregulated in the
ESCC cells, the migration and invasion ability of ESCC cells was
downregulated, and when miR-675-3p levels were then upregulated,
the migration and invasion ability of the ESCC cells recovered to
levels similar to the control group. These results indicated that
miR-675-3p could serve as a biomarker for tumor progression and
development research, or for clinical diagnosis, but this needs to
be further investigated.
MMP2, MMP9 (36–40)
and E-cadherin, markers of EMT, are usually involved in cancer
progression (24), cancer cell
invasion events (41), and also
may participate in regulating cell migration (42–44)
and invasion ability (45).
Therefore, to reveal whether miR-675-3p participates in the
regulation of ESCC cell migration and invasion, miR-675-3p was
downregulated in ESCC cells using an miR-675-3p inhibitor. It was
demonstrated that inhibition of miR-675-3p in Kyse-150 and Te-1
cells significantly inhibited cell growth, as well as cellular
migration and invasion capabilities. Furthermore, it was also
demonstrated that miR-675-3p may serve an important role in
influencing some proteins, such as MMP2, MMP9, and E-cadherin,
which may affect the ability of migration and invasion in ESCC
cells, however the underlying mechanism has not yet been
investigated. Therefore, understanding the key role of miR-675-3p
in ESCC may lead to the discovery of a novel biomarker, or
identification of novel therapeutic targets for treating esophageal
cancer, however this requires further investigation.
In conclusion, the present results are at the
forefront of research for miR-675-3p, and further investigation is
required for miR-675-3p potential targets and its function in
tumorigenesis.
Acknowledgements
Not applicable.
Funding
The present study was supported by the key
development plan for social development of Jiangsu Province (grant
no. SBE2016750057) and Jiangsu Provincial Key R & D Program
Social Development Clinical Frontier Technology Project
(Application of Image-guided Precise Tumor Surgical Equipment in
Esophageal Cancer Surgery; grant no. BE2016731).
Availability of data and materials
The analyzed data sets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
QX conceived, designed, drafted and performed the
experiments. TC and YW performed some experiments and analyzed the
data. WW participated in analyzing and interpreting the data. YX
provided help in conceiving and designing the study and revising
the manuscript. ZG contributed to the conception, design and guided
assays, gave approval of the version to be published, and
supervised the project. SC conceived the main goal for the study,
designed the experiment, guided assays, provided financial support
of the present study and the research to gave final approval of the
version of the manuscript. All authors have approved the final
manuscript.
Ethics approval and consent to
participate
Written informed consent was obtained from all
participants, and the study protocol was approved by the ethics
committee of JiangSu Cancer Hospital (Nanjing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
López-Gómez M, Malmierca E, de Górgolas M
and Casado E: Cancer in developing countries: The next most
preventable pandemic. The global problem of cancer. Crit Rev Oncol
Hematol. 88:117–122. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sankaranarayanan R, Ramadas K and Qiao YL:
Managing the changing burden of cancer in Asia. BMC Med. 12:32014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Katzka DA: Recent advances in non-invasive
esophageal tissue sampling. Curr Gastroenterol Rep. 19:92017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Peery AF, Crockett SD, Barritt AS, Dellon
ES, Eluri S, Gangarosa LM, Jensen ET, Lund JL, Pasricha S, Runge T,
et al: Burden of gastrointestinal, liver, and pancreatic diseases
in the United States. Gastroenterology. 149:1731–1741.e3. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pennathur A, Gibson MK, Jobe BA and
Luketich JD: Oesophageal carcinoma. Lancet. 381:400–412. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhu ZJ, Hu Y, Zhao YF, Chen XZ, Chen LQ
and Chen YT: Early recurrence and death after esophagectomy in
patients with esophageal squamous cell carcinoma. Ann Thorac Surg.
91:1502–1508. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hayes J, Peruzzi PP and Lawler S:
MicroRNAs in cancer: biomarkers, functions and therapy. Trends in
Mol Med. 20:460–469. 2014. View Article : Google Scholar
|
|
9
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li C, Lei B, Huang S, Zheng M, Liu Z, Li Z
and Deng Y: H19 derived microRNA-675 regulates cell proliferation
and migration through CDK6 in glioma. Am J Transl Res. 7:1747–1764.
2015.PubMed/NCBI
|
|
11
|
Keniry A, Oxley D, Monnier P, Kyba M,
Dandolo L, Smits G and Reik W: The H19 lincRNA is a developmental
reservoir of miR-675 that suppresses growth and Igf1r. Nat Cell
Biol. 14:659–665. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhu M, Chen Q, Liu X, Sun Q, Zhao X, Deng
R, Wang Y, Huang J, Xu M, Yan J and Yu J: lncRNA H19/miR-675 axis
represses prostate cancer metastasis by targeting TGFBI. FEBS J.
281:3766–3775. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tsang WP, Ng EK, Ng SS, Jin H, Yu J, Sung
JJ and Kwok TT: Oncofetal H19-derived miR-675 regulates tumor
suppressor RB in human colorectal cancer. Carcinogenesis.
31:350–358. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jiang Z, Zhang H, Liu C, Yin J, Tong S, Lv
J, Wei S and Wu S: β3GnT8 promotes colorectal cancer cells invasion
via CD147/MMP2/Galectin3 axis. Front Physiol. 9:5882018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kang DY, Sp N, Kim DH, Joung YH, Lee HG,
Park YM and Yang YM: Salidroside inhibits migration, invasion and
angiogenesis of MDA-MB 231 TNBC cells by regulating EGFR/Jak2/STAT3
signaling via MMP2. Int J Oncol. 53:877–885. 2018.PubMed/NCBI
|
|
17
|
Marshall DC, Lyman SK, McCauley S,
Kovalenko M, Spangler R, Liu C, Lee M, O'Sullivan C, Barry-Hamilton
V, Ghermazien H, et al: Selective allosteric inhibition of MMP9 is
efficacious in preclinical models of ulcerative colitis and
colorectal cancer. PLoS One. 10:e01270632015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Grauzam S, Brock AM, Holmes CO, Tiedeken
JA, Boniface SG, Pierson BN, Patterson DG, Coaxum SD, Neskey DM and
Rosenzweig SA: NEDD9 stimulated MMP9 secretion is required for
invadopodia formation in oral squamous cell carcinoma. Oncotarget.
9:25503–25516. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yuan YL, Wang YM, Liu H, Qin GF, Tang AG
and Duan Y: Aberrant expression of E-cadherin in lung tissues of
patients with probable lung cancer. Asian Pac J Cancer Prev.
13:5149–5153. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wong TS, Gao W and Chan JY: Interactions
between E-cadherin and microRNA deregulation in head and neck
cancers: the potential interplay. Biomed Res Int. 2014:1260382014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Repetto O, De Paoli P, De Re V, Canzonieri
V and Cannizzaro R: Levels of soluble E-cadherin in breast,
gastric, and colorectal cancers. Biomed Res Int. 2014:4080472014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Techasen A, Loilome W, Namwat N, Khuntikeo
N, Puapairoj A, Jearanaikoon P, Saya H and Yongvanit P: Loss of
E-cadherin promotes migration and invasion of cholangiocarcinoma
cells and serves as a potential marker of metastasis. Tumour Biol.
35:8645–8652. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang CA, Drasin D, Pham C, Jedlicka P,
Zaberezhnyy V, Guney M, Li H, Nemenoff R, Costello JC, Tan AC and
Ford HL: Homeoprotein Six2 promotes breast cancer metastasis via
transcriptional and epigenetic control of E-cadherin expression.
Cancer Res. 74:7357–7370. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kreiseder B, Orel L, Bujnow C, Buschek S,
Pflueger M, Schuett W, Hundsberger H, de Martin R and Wiesner C:
α-Catulin downregulates E-cadherin and promotes melanoma
progression and invasion. Int J Cancer. 132:521–530. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shin VY and Chu KM: MiRNA as potential
biomarkers and therapeutic targets for gastric cancer. World J
Gastroenterol. 20:10432–10439. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yoshizawa JM and Wong DT: Salivary
microRNAs and oral cancer detection. Methods Mol Biol. 936:313–324.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rocci A, Hofmeister CC and Pichiorri F:
The potential of miRNAs as biomarkers for multiple myeloma. Expert
Rev Mol Diagn. 14:947–959. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pan Y, Liang H, Chen W, Zhang H, Wang N,
Wang F, Zhang S, Liu Y, Zhao C, Yan X, et al: microRNA-200b and
microRNA-200c promote colorectal cancer cell proliferation via
targeting the reversion-inducing cysteine-rich protein with Kazal
motifs. RNA Biol. 12:276–289. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yamada N, Tsujimura N, Kumazaki M,
Shinohara H, Taniguchi K, Nakagawa Y, Naoe T and Akao Y: Colorectal
cancer cell-derived microvesicles containing microRNA-1246 promote
angiogenesis by activating Smad 1/5/8 signaling elicited by PML
down-regulation in endothelial cells. Biochim Biophys Acta.
1839:1256–1272. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jin M, Zhang T, Liu C, Badeaux MA, Liu B,
Liu R, Jeter C, Chen X, Vlassov AV and Tang DG: miRNA-128
suppresses prostate cancer by inhibiting BMI-1 to inhibit
tumor-initiating cells. Cancer Res. 74:4183–4195. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xiao R, Li C and Chai B: miRNA-144
suppresses proliferation and migration of colorectal cancer cells
through GSPT1. Biomed Pharmacother. 74:138–144. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu HT, Xing AY, Chen X, Ma RR, Wang YW,
Shi DB, Zhang H, Li P, Chen HF, Li YH and Gao P: MicroRNA-27b,
microRNA-101 and microRNA-128 inhibit angiogenesis by
down-regulating vascular endothelial growth factor C expression in
gastric cancers. Oncotarget. 6:37458–37470. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tao J, Zhi X, Zhang X, Fu M, Huang H, Fan
Y, Guan W and Zou C: miR-27b-3p suppresses cell proliferation
through targeting receptor tyrosine kinase like orphan receptor 1
in gastric cancer. J Exp Clin Cancer Res. 34:1392015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
He D, Wang J, Zhang C, Shan B, Deng X, Li
B, Zhou Y, Chen W, Hong J, Gao Y, et al: Down-regulation of
miR-675-5p contributes to tumor progression and development by
targeting pro-tumorigenic GPR55 in non-small cell lung cancer. Mol
Cancer. 14:732015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhou YW, Zhang H, Duan CJ, Gao Y, Cheng
YD, He D, Li R and Zhang CF: miR-675-5p enhances tumorigenesis and
metastasis of esophageal squamous cell carcinoma by targeting
REPS2. Oncotarget. 7:30730–30747. 2016.PubMed/NCBI
|
|
36
|
Wang X, Yang B, She Y and Ye Y: The lncRNA
TP73-AS1 promotes ovarian cancer cell proliferation and metastasis
via modulation of MMP2 and MMP9. J Cell Biochem. Jun 15–2018.(Epub
ahead of print).
|
|
37
|
Li H, Zhang Y, Hai J, Wang J, Zhao B, Du L
and Geng X: Knockdown of TRIM31 suppresses proliferation and
invasion of gallbladder cancer cells by down-regulating MMP2/9
through the PI3K/Akt signaling pathway. Biomed Pharmacother.
103:1272–1278. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kalhori V and Törnquist K: MMP2 and MMP9
participate in S1P-induced invasion of follicular ML-1 thyroid
cancer cells. Mol Cell Endocrinol. 404:113–122. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jacob A, Jing J, Lee J, Schedin P, Gilbert
SM, Peden AA, Junutula JR and Prekeris R: Rab40b regulates
trafficking of MMP2 and MMP9 during invadopodia formation and
invasion of breast cancer cells. J Cell Sci. 126:4647–4658. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu J, Ping W, Zu Y and Sun W:
Correlations of lysyl oxidase with MMP2/MMP9 expression and its
prognostic value in non-small cell lung cancer. Int J Clin Exp
Pathol. 7:6040–6047. 2014.PubMed/NCBI
|
|
41
|
Qiu X, Cheng JC, Chang HM and Leung PC:
COX2 and PGE2 mediate EGF-induced E-cadherin-independent human
ovarian cancer cell invasion. Endocr Relat Cancer. 21:533–543.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cai D, Chen SC, Prasad M, He L, Wang X,
Choesmel-Cadamuro V, Sawyer JK, Danuser G and Montell DJ:
Mechanical feedback through E-cadherin promotes direction sensing
during collective cell migration. Cell. 157:1146–1159. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chen D, Wu Z, Luo LJ, Huang X, Qian WQ,
Wang H, Li SH and Liu J: E-cadherin maintains the activity of
neural stem cells and inhibits the migration. Int J Clin Exp
Pathol. 8:14247–14251. 2015.PubMed/NCBI
|
|
44
|
Brett A, Pandey S and Fraizer G: The
Wilms' tumor gene (WT1) regulates E-cadherin expression and
migration of prostate cancer cells. Mol Cancer. 12:32013.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Canel M, Serrels A, Frame MC and Brunton
VG: E-cadherin-integrin crosstalk in cancer invasion and
metastasis. J Cell Sci. 126:393–401. 2013. View Article : Google Scholar : PubMed/NCBI
|