Introduction
Prostate cancer is a common malignant tumor that
occurs in the prostate epithelium and is the second leading cause
of cancer-associated mortalities among older men worldwide
(1). Prostate cancer begins with
intraepithelial neoplasia, and locally invasive prostate cancer
gradually progresses to castration resistant prostate cancer, which
represents the leading cause of prostate cancer-associated
mortalities at present (2). This
is a complex process involving alterations in the extracellular
matrix microenvironment to support the invasion and increase of
cell motility (3). However, the
exact underlying mechanism of prostate cancer progression is
unclear and determining the molecular mechanism is crucial for
clinical diagnosis and therapy.
Nuclear factor (NF)-κB represents an important
member of the transcription factor family, which widely exists in
eukaryotes to participate in multiple physiological and
pathological processes (4). It has
been demonstrated that NF-κB may adjust cell apoptosis in a number
of different ways, involving regulation of the expression of
apoptosis-associated genes and interacting with other signaling
pathways (5). A previous study
also reported that in epidermal cells, the activation of NF-κB
suppressed cell proliferation (6).
Nuclear factor-erythroid 2-related factor 2 (Nrf2) serves an
important role in the anti-oxidative signaling pathway in organisms
(7). Nrf2 can be activated by the
external stimuli from free radicals and is then transferred to the
nucleus to combine with the antioxidant response element (8,9). It
has been reported previously that the activity of antioxidases from
Nrf2 knockdown mice was significantly reduced, while the expression
of correlative cytokines was markedly enhanced, involving
interleukin (IL)-6, IL-1β, tumor necrosis factor-α, inducible
nitric oxide synthase and cyclo-oxygenase 2 (10,11).
In addition, a large number of investigations have suggested that
interactions exist between NF-κB and Nrf2. For example, NF-κB may
suppress the activity of Nrf2 by selectively preventing the
combination of Nrf2 and cyclic adenosine 3′,5′-monophosphate
response element binding protein (12,13).
However, to the best of our knowledge, the roles and mechanisms of
the NF-κB/Nrf2 signaling pathway in prostate cancer have not been
fully elucidated.
Brain-type glycogen phosphorylase (PYGB), as a
glycogen phosphorylase, metabolizes glycogen. It is generally
considered to provide energy for organisms in the emergency state
(14–16). PYGB has been investigated
previously in tumors of the gastrointestinal system and in lung
carcinomas (17–20). In addition, it has been
demonstrated that PYGB may inhibit the production of reactive
oxygen species (ROS) and suppress the apoptosis of Escherichia
coli (E. coli) cells (21). To the best of our knowledge, the
effect of PYGB on prostate cancer tissues and cells has not yet
been reported.
The present study analyzed the association between
PYGB expression and prostate cancer tissues together with
clinicopathological data. In addition, the present study explored
the effect of PYGB silencing and the NF-κB/Nrf2 signaling pathway
in the growth and apoptosis of prostate cancer cells.
Materials and methods
Tissue samples and clinical data
collection
Patients that had been pathologically diagnosed with
prostate cancer were included in the present study. Patients were
excluded from the experiment if androgen or radiation therapy had
been received. A total of 50 male patients (mean age, 70.5)
undergoing resection therapy at the East Hospital, Tongji
University School of Medicine (Shanghai, China) from June 2010 to
August 2015 were included in this study. All patients recruited to
the present study provided written informed consent for the
utilization of their tissue samples for clinical research. The
project protocol was approved by the Institutional Review Board of
East Hospital, Tongji University School of Medicine.
Clinicopathological data, involving patient age and sex,
histological grade and tumor, nodes, metastasis (TNM) stage, was
obtained from the patient database. Matched adjacent normal
prostate tissues were also collected as negative controls.
Following resection, all of the tissue samples were snap-frozen in
liquid nitrogen and stored at −80°C immediately for the subsequent
experiments.
Cell culture, genes and plasmids
Human prostate cancer cell lines, including LNCap,
PC3 and DU145, and the normal prostate cell line PrEC were obtained
from the Type Culture Collection of The Chinese Academy of Sciences
(Shanghai, China). Cells were maintained in RPMI-1640 medium
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) in a 5% CO2
atmosphere at 37°C. PYGB small interfering (si)-RNA
(5′-AGGCCTGGGAAATCACGAAG-3′) or negative siRNA control
(5′-GGAACGGAGGCGAAGAGATAT-3′) was cloned into the pcDNA3.1(+) empty
vector (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). si-PYGB or siRNA-NC was sub-cloned into the pSilencer 2.1-U6
vector (Thermo Fisher Scientific) for gene interference. The
expression of PYGB was detected 20 h post-transfection.
Cell viability analysis
Cell Counting Kkit-8 (CCK-8; Beyotime Institute of
Biotechnology, Haimen, China) assay was performed to determine cell
viability. A total of 6×104 cells/ml PC3 cells in the
logarithmic phase were seeded into the wells of 96-well plates and
maintained in an incubator (37°C and 5% CO2) for 12 h.
Following this, cells were transfected with siRNA negative vector
(1 µg; mock group) or PYGB siRNA vector (1 µg; si-PYGB group) using
Lipofectamine 3000 (Invitrogen; Thermo Fisher Scientific, Inc.) for
20 h. The untreated cells served as a control. Cells were then
maintained in the incubator (37°C, 5% CO2) for 12, 24
and 48 h. Subsequently, 10 µl CCK reagent was supplemented into
each well. Cells were then maintained in 5% CO2 at 37°C
for 3 h. A microplate reader (Bio-Rad Laboratories, Inc., Hercules,
CA, USA) was used to read the absorbance at a wavelength of 450 nm.
Cell viability was evaluated by calculating the percentage of cell
survival compared with the control.
Apoptosis assay
Flow cytometry (FCM) was used to assess the cell
apoptosis. Following washing with PBS, PC3 cells were trypsinized
using 0.25% Trypsin (Beyotime Institute of Biotechnology). An
Annexin V-FITC/PI Apoptosis Detection kit was purchased from
Invitrogen (Thermo Fisher Scientific, Inc.). Cells were centrifuged
at 224 × g for 1 min at 4°C, and the supernatant was discarded.
Subsequently, cells were suspended in Annexin-V binding buffer at a
density of 1×106 cells/ml. Annexin V-fluorescein
isothiocyanate and propidium iodide were then maintained with cells
at room temperature in the dark for 15 min. Cell apoptosis was
evaluated by using a FACSCalibur flow cytometer with BD CellQuest
Pro 3.3 (BD Biosciences, Franklin Lakes, CA, USA).
Western blot analysis
Tissues were rapidly grinded in liquid nitrogen.
Cells were lysed with NP40 lysis buffer (Beyotime) and protein was
isolated using a Tissue or Cell Total Protein Extraction kit
(Solarbio, China). Protein concentration was measured by BCA assay
kit (Thermo, USA). Equal amount of proteins (25 µg/lane) were
separated by 12% SDS-PAGE and then transferred onto a
polyvinylidene difluoride membrane (EMD Millipore, Billerica, MA,
USA). The membranes were blocked with 5% skimmed milk at room
temperature for 1 h. The blotting was performed using specific
primary antibodies at 4°C overnight, including anti-PYGB (1:2,000;
Abcam, Cambridge, UK; cat. no. ab154969; rabbit anti-human),
anti-poly (adenosine diphosphate-ribose) polymerase (PARP; 1:2,000;
Abcam; cat. no. ab32138; rabbit anti-human), anti-cleaved-PARP
(1:5,000; Abcam; cat. no. ab32064; rabbit anti-human),
anti-cleaved-caspase-3 (1:1,000; Abcam; cat. no. ab2302; rabbit
anti-human), anti-NF-κB (1:500; Abcam; cat. no. ab31412; rabbit
anti-human), anti-Nrf2 (1:1,000; Abcam; cat. no. ab89443; mouse
anti-human), anti-B-cell lymphoma-2 (Bcl-2; 1:1,000; Abcam; cat.
no. ab32124; rabbit anti-human), anti-Bcl-2-associated X protein
(Bax; 1:1,000; Abcam; cat. no. ab32503; rabbit anti-human) and
anti-actin (1:500; Abcam; cat. no. ab205; mouse anti-human). A
horseradish peroxidase-conjugated secondary antibody (1:1,000; cat.
no. bs-0293M; BIOSS, Beijing, China) was incubated with the blots
at room temperature for 1 h. Enhanced chemiluminescent reagents
(EMD Millipore) using an ECL system (GE Healthcare, Chicago, IL,
USA) was used for visualization. The density of the blots was
determined with Quantity One 4.6.2 software (Bio-Rad Laboratories,
Inc.).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from tissues or cultured
cell lines (PrEC, LNCap, PC3 and DU145) with TRIzol reagent (Thermo
Fisher Scientific, Inc.). RNA was reverse transcribed to cDNA using
a Transcriptor High Fidelity cDNA Synthesis kit (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) according to the manufacturer's
protocol. The temperature protocol consisted of 25°C for 5 min,
37°C for 60 min and 85°C for 5 min. RT-qPCR analysis was performed
using SYBR Green qPCR Master mix (MedChemExpress, Princeton, NJ,
USA) on an ABI 7500 thermocycler (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The qPCR thermocycling conditions were as
follows: 5 min pretreatment at 95°C, followed by 35 cycles of 95°C
for 15 sec and 60°C for 30 sec, a final extension at 72°C for 10
min and then held at 4°C. Actin was used as the control of the
input RNA level. The primers used in this analysis were designed by
Invitrogen (Thermo Fisher Scientific, Inc.) and were as follows:
PYGB, forward, 5′-AGGCCTGGGAAATCACGAAG-3′ and reverse,
5′-CCATGTTGATCCGCTTGCAG-3′ (product: 237 bp); NF-κB, forward,
5′-GCGAGAGGAGCACAGATACC-3′ and reverse, 5′-AGGGGTTGTTGTTGGTCTGG-3′
(product: 279 bp); Nrf2, forward, 5′-TGAGGTTTCTTCGGCTACGTT-3′ and
reverse, 5′-AGCTCCTCCCAAACTTGCTC-3′ (product: 174 bp); Bax,
forward, 5′-CAGCTCTGAGCAGATCATGAAGACA-3′ and reverse,
5′-GCCCATCTTCTTCCAGATGGTGAGC-3′ (product: 235 bp); Bcl-2, forward,
5′-ACTTGTGGCCCAGATAGGCACCCAG-3′ and reverse,
5′-CGACTTCGCCGAGATGTCCAGCCAG-3′ (product: 214 bp); and Actin,
forward, 5′-CACAATGTGCGACGAAGACG-3′ and reverse,
5′-ATGATGCCGTGCTCGATAGG-3′ (product: 237 bp). Gene expression was
quantified according to the 2−ΔΔCq method (22).
Statistical analysis
The results of the present study are presented as
the mean ± standard deviation of at least three independent
experiments. All experimental data was analyzed by a Student's t
test, or one-way analysis of variance followed by Tukey's post-hoc
test. Kaplan-Meier survival analysis was performed and P-values
were calculated with log-rank test. The differences between the
categorial variables were determined with the Chi-square test.
P<0.05 was considered to indicate a statistically significant
difference. GraphPad Prism 6.0 (GraphPad Software, Inc., La Jolla,
CA, USA) was used to perform the data analysis.
Results
PYGB is upregulated in prostate cancer
tissues and is associated with disease progression
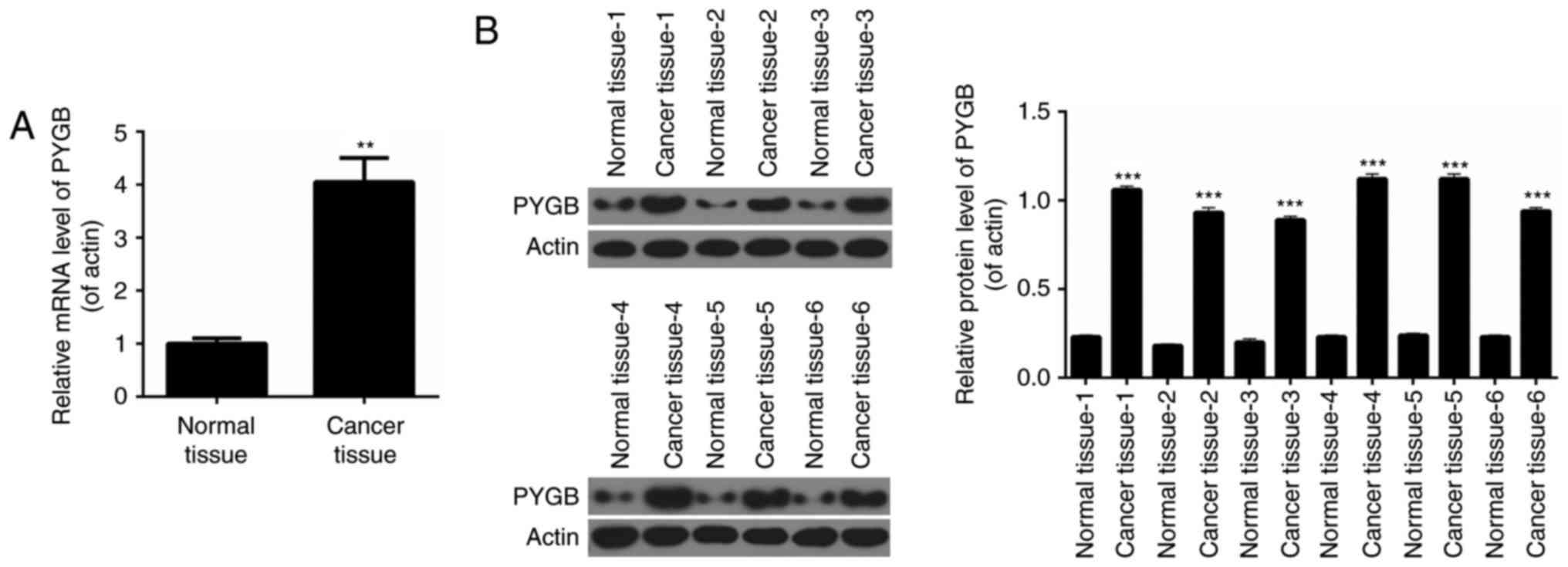
RT-qPCR analysis was performed to determine the
expression levels of PYGB in pairs of human prostate cancer tissues
and matched adjacent normal prostate tissues from 50 prostate
cancer patients. It was revealed that the expression levels of PYGB
in prostate cancer tissues were significantly higher than those
observed in pair-matched adjacent non-tumorous tissues (Fig. 1A; P<0.01). Western blot analysis
was performed to evaluate PYGB protein expression in 6 randomly
selected pairs of human prostate cancer tissues and matched
adjacent normal prostate tissues from prostate cancer patients. The
results indicated that compared with pair-matched adjacent
non-tumorous tissues, PYGB expression was significantly enhanced in
prostate cancer tissues (Fig. 1B;
P<0.001). These results implied that PYGB expression was
upregulated in prostate cancer tissues. Clinicopathological
analysis revealed that PYGB was associated with the degree of
histological differentiation (P<0.05), TNM stage (P<0.01) and
metastasis (P<0.01) of prostate cancer tissues (Table I). However, there was no
significant association between PYGB and other clinicopathological
characteristics, involving age and gender (Table I; P>0.05). In addition, the
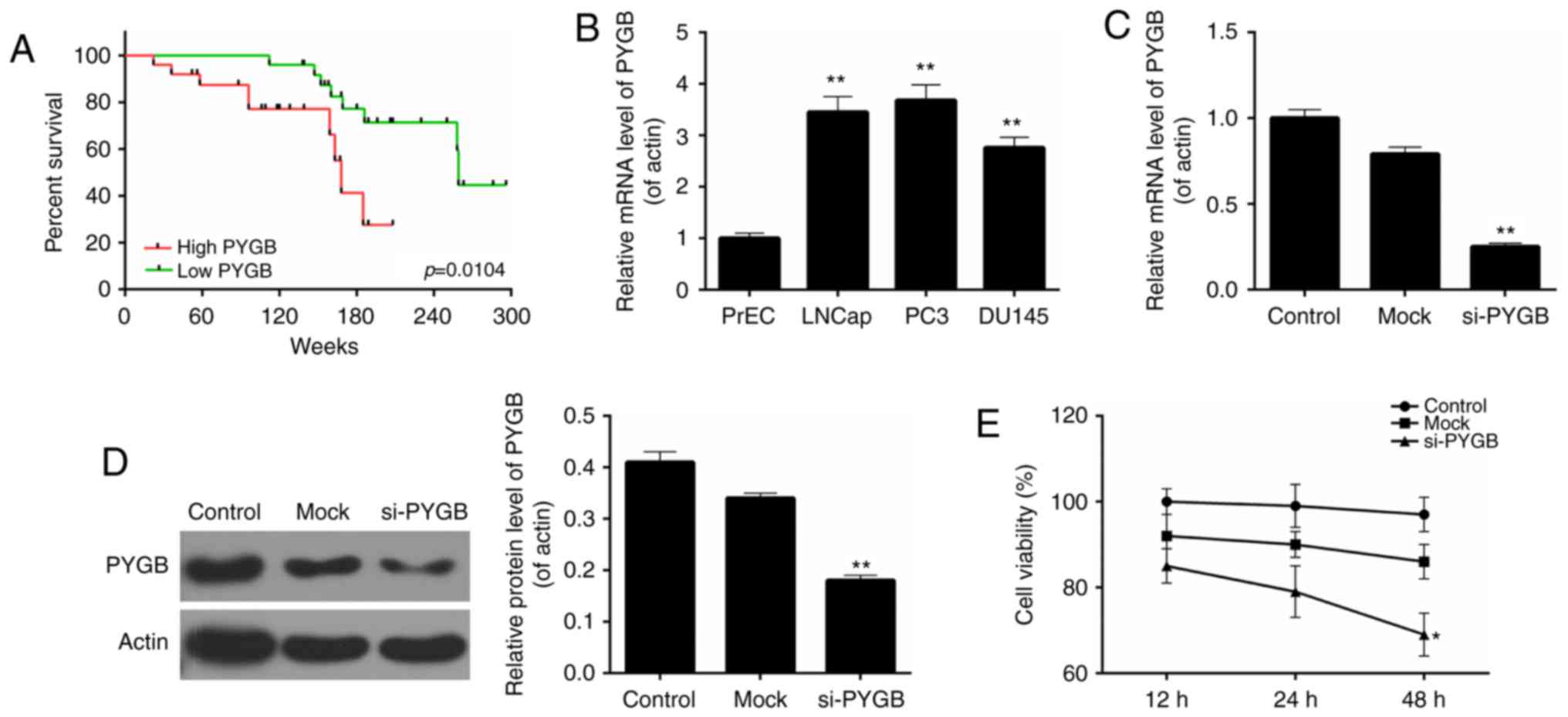
present study also assessed the association between PYGB expression
in prostate cancer and patient survival to understand the
prognostic significance of PYGB upregulation in prostate cancer. It
was demonstrated that high expression levels of PYGB were
significantly associated with a poor 5-year survival rate in
prostate cancer patients (Fig. 2A;
P<0.05). Low PYGB expression was associated with a higher 5-year
survival rate in prostate cancer patients. Collectively, these
results indicated that high PYGB expression may be associated with
prostate cancer progression.
 | Table I.Associations between brain-type
glycogen phosphorylase expression levels and the clinical data of
prostate cancer patients. |
Table I.
Associations between brain-type
glycogen phosphorylase expression levels and the clinical data of
prostate cancer patients.
| Factor | Number of
patients | Low PYGB
expression, n (%) | High PYGB
expression, n (%) | P-value |
|---|
| Sex |
|
|
|
|
|
Male | 50 | 21 (100.0) | 29 (100.0) |
|
| Age (years) |
|
|
| 0.054 |
|
<55 | 18 | 4 (20.0) | 14 (46.7) |
|
|
≥55 | 32 | 16 (80.0) | 16 (53.3) |
|
| Histological
grade |
|
|
| 0.012a |
|
Well | 5 | 2 (13.3) | 3 (8.6) |
|
|
Moderately | 35 | 10 (66.7) | 25 (71.4) |
|
|
Poorly | 10 | 3 (20.0) | 7 (20.0) |
|
| TNM stage |
|
|
| 0.008b |
|
I/II | 23 | 5 (20.8) | 18 (75.0) |
|
|
III/IV | 27 | 19 (79.2) | 6 (25.0) |
|
| Metastasis |
|
|
| <0.001 |
| No | 38 | 9 (47.4) | 29 (93.5) |
|
|
Yes | 12 | 10 (52.6) | 2 (6.5) |
|
PYGB silencing suppresses cell
viability of PC3 cells
Based on the above results, the present study
evaluated the cell viability of three different prostate cancer
cell lines, including LNCap, PC3 and DU145, together with a normal
prostate cell line PrEC. The results revealed that, when compared
with the normal prostate cell line, the PYGB expression was
significantly higher in the three prostate cancer cell lines
(Fig. 2B). The highest PYGB
expression was detected in the PC3 cell line. Therefore, the PC3
cell line was chosen as the research target of prostate cancer
cells in the subsequent studies. The knockdown efficiency was ~75%
in PC3 cells following cells being transfected with si-PYGB
(Fig. 2C). According to the
western blotting results, it was revealed that following
transfection with si-PYGB, the expression level of the PYGB protein
was significantly reduced (Fig.
2D; P<0.01). Thus, the CCK-8 assay was performed to
determine the cell viability of PC3 cells for 12, 24 and 48 h
following siRNA transfection to further evaluate the potential
effects of PYGB silencing. The results showed that, when compared
with the non-transfected control (control) and non-targeting
transfected control (mock), the cell viability of PC3 cells treated
with si-PYGB for 48 h was significantly reduced (Fig. 2E; P<0.05). However, no
significant difference was revealed between the control and mock
groups at each time point (Fig.
2E). These results suggested that PYGB silencing suppressed
cell viability of PC3 cells.
PYGB silencing promotes the apoptosis
of PC3 cells
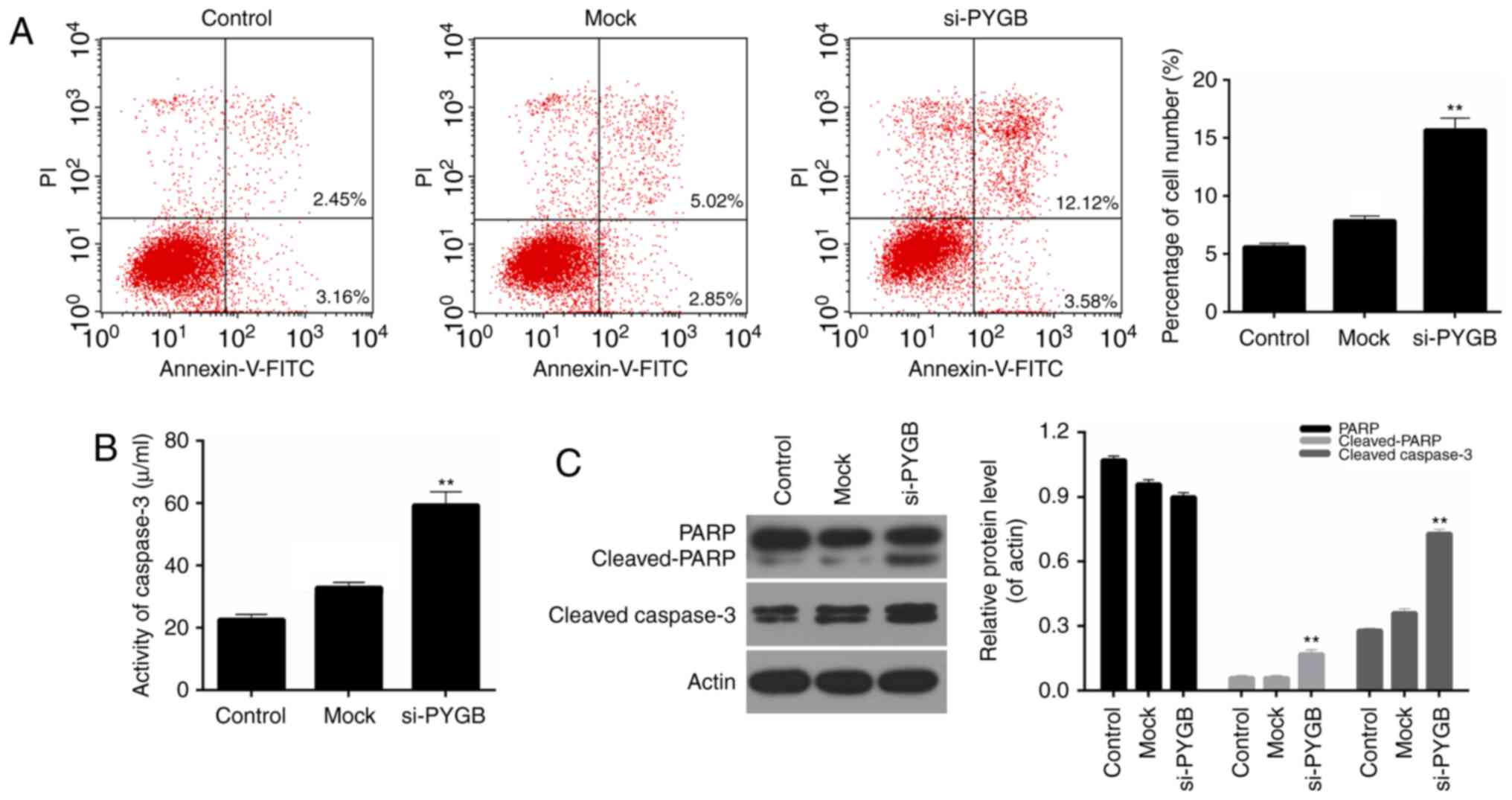
In addition, the cell apoptosis of PC3 cells was
assessed by FCM in the present study. As the FCM results revealed
(Fig. 3A), the percentage of
apoptotic PC3 cells in the si-PYGB group was 15.70%, which was
markedly higher than the control and mock groups (5.61 and 7.87%,
respectively; P<0.05). These results suggested that PYGB
silencing significantly strengthened the apoptotic capacity of PC3
cells. In addition, the activity of caspase-3 in PC3 cells was
evaluated. It was noted that the activity of caspase-3 in PC3
treated with si-PYGB was significantly enhanced (Fig. 3B; P<0.01). The present study
also investigated the expression levels of apoptotic-associated
proteins in PC3 cells. According to the RT-qPCR and western
blotting data, it was demonstrated that PARP expression in the
three groups was not significantly different, whereas an increase
in cleaved-PARP expression in PC3 cells was observed in the si-PYGB
group (Fig. 3C; P<0.05). It was
also revealed that there were high expression levels of
cleaved-caspase-3 in PC3 cells treated with si-PYGB (Fig. 3C; P<0.05). In addition, the
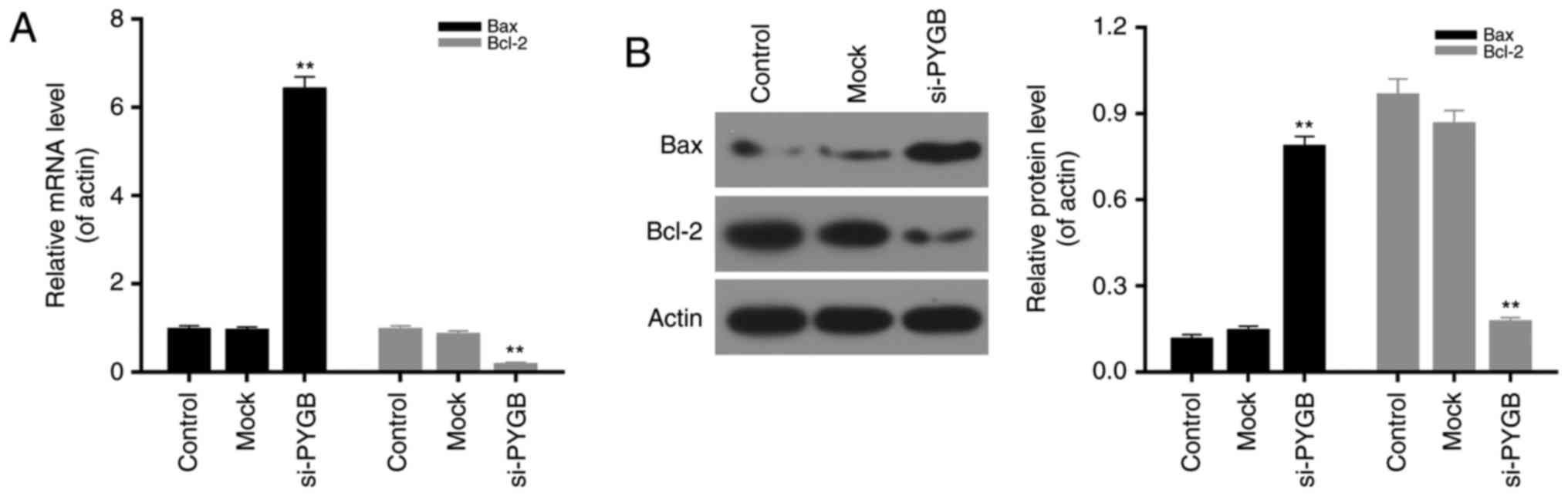
present study measured the expression levels of Bax and Bcl-2 in
PC3 cells transfected with si-PYGB to further determine the
associated apoptotic mechanisms. The results revealed that PTGB
silencing significantly upregulated the mRNA and protein expression
levels of Bax, and reduced Bcl-2 expression levels in PC3 cells
(Fig. 4; P<0.05). Based on
these results, it was concluded that PYGB silencing promoted the
apoptosis of PC3 cells via regulation of cleaved-PARP,
cleaved-caspase-3, Bax and Bcl-2 expression.
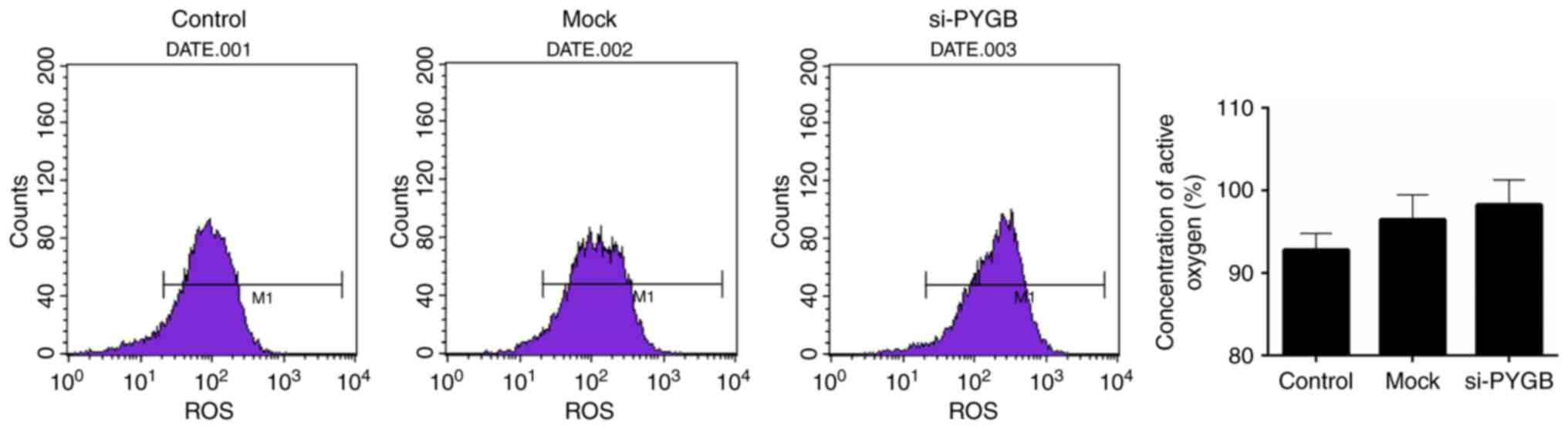
PYGB silencing increases the ROS
content in PC3 cells
The present study also assessed the ROS content in
PC3 cells. FCM results revealed that the ROS content appeared
enhanced in the si-PYGB group when compared with the control group
(Fig. 5); however, this was not
significant.
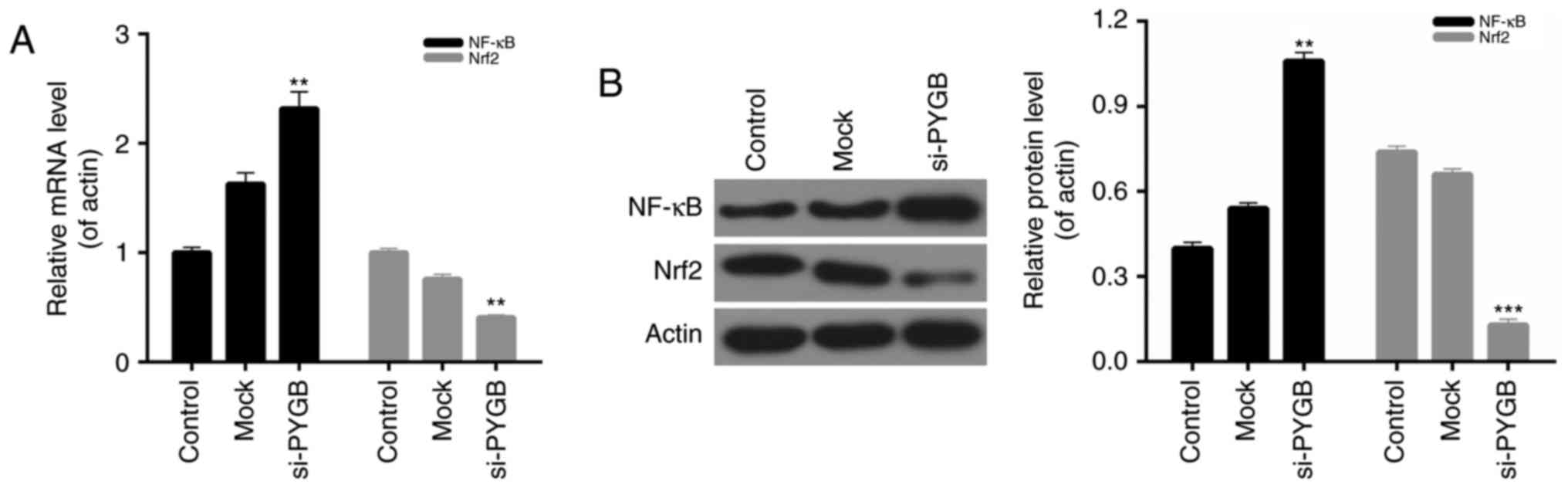
PYGB silencing affects the NF-κB/Nrf2
signaling pathway in PC3 cells
The present study also evaluated the expression of
NF-κB and Nrf2 in PC3 cells from each group. The RT-qPCR and
western blotting results indicated that the mRNA and protein
expression levels of NF-κB in PC3 cells transfected with si-PYGB
were significantly higher than the control and mock groups.
However, a decrease in Nrf2 expression was observed in the PC3
cells treated with si-PYGB (Fig.
6; P<0.01). Therefore, it was concluded that the NF-κB/Nrf2
signaling pathway in PC3 cells may be affected by PYGB
silencing.
Discussion
PYGB is a glycogen phosphorylase and is primarily
localized in adult brain tissues and embryo liver tissues, whose
function is to provide energy to organisms (15). Increasing evidence has suggested
that PYGB is frequently expressed in cancer, including gastric
carcinoma (23), lung cancer
(24) and renal cell cancer
(25). For example, it has been
previously demonstrated that PYGB expression was significantly
upregulated in hepatocellular carcinoma, whereas the expression
levels of PYGB in normal liver cells were low or not expressed
(26). However, the PYGB
expression levels in prostate cancer tissues or cells are currently
not clear. Thus, the present study assessed PYGB expression in
human prostate cancer tissues and matched adjacent normal prostate
tissues from patients with prostate cancer. The results suggested
that the expression levels of PYGB in prostate cancer tissues were
markedly enhanced, while low PYGB expression was observed in normal
tissues. In addition, the present study also evaluated PYGB
expression in a number of different prostate cancer cell lines, and
the results revealed that PYGB expression was relevant to the
malignant degree of prostate cancer cells. These consequences were
consistent with the previous studies in regard to PYGB expression
in other cancer cells (17–20),
and indicated that PYGB may act as a pathological diagnostic index
for malignant prostate neoplasia's. According to the
clinicopathological analysis data, the present study confirmed that
PYGB was associated with the degree of histological
differentiation, TNM stage and metastasis of prostate cancer
tissues. Furthermore, via survival curve analysis of the expression
levels of PYGB and the survival time of prostate cancer patients,
it was observed that high PYGB expression was associated with a
poor 5-year survival rate and low PYGB expression was associated
with an improved 5-year survival rate in the prostate cancer
patients. These results also implied that PYGB may serve as a
prognostic indicator of prostate cancer. Overall, the present study
proposed that PYGB may become an important target in the detection
and therapy of prostate cancer.
In order to further investigate the role and
underlying mechanism of PYGB in prostate cancer, the present study
selected the PC3 cell line as the research target. The results of
the present study demonstrated that the expression levels of PYGB
in PC3 cells were higher than in the other prostate cancer cell
lines and the normal prostate cell line (PrEC). In addition, an
siRNA vector target of PYGB, si-PYGB, was constructed in the
present study. To the best of our knowledge, the present study is
the first to investigate the PYGB silencing effect on cell growth
and apoptosis of prostate cancer cells. Following treatment with
si-PYGB, the expression levels of PYGB in PC3 cells were reduced,
which indicated that a good knockdown efficiency was achieved. The
present study further evaluated the cell viability of PC3 cells
transfected with si-PYGB, and the results demonstrated that PYGB
silencing suppressed the cell viability of PC3 cells. In addition,
the apoptotic capacity of PC3 cells treated with the empty vector
and si-PYGB was assessed. The data revealed that when compared with
other groups, PYGB silencing promoted the apoptosis of PC3 cells.
Furthermore, based on the previous studies (27,28),
the present study also measured the activity of caspase-3 and
investigated the expression levels of several apoptotic-associated
proteins in PC3 cells transfected with the empty vector and
si-PYGB. The results indicated that PYGB silencing enhanced the
activity of caspase-3 in PC3 cells. It was also demonstrated that
PYGB silencing upregulated the expression levels of cleaved-PARP,
cleaved-caspase-3 and Bax, and reduced the Bcl-2 expression levels
in PC3 cells. Therefore, it was concluded that PYGB silencing
promoted the apoptosis of PC3 cells by modulating the expression
levels of cleaved-PARP, cleaved-caspase-3, Bax and Bcl-2. It has
been previously demonstrated that PYGB inhibited the production of
ROS, and suppressed the apoptosis and necrosis of E. coli
cells (21). In the present study,
it was confirmed that PYGB silencing promoted the apoptosis of PC3
cells; however, whether PYGB silencing induces ROS production
required further investigation. Therefore, ROS content in PC3 cells
transfected with the empty vector and si-PYGB was assessed. The
results showed that PYGB silencing increased the production of ROS
in PC3 cells, and these data also verified the cell apoptosis
results. Thus, it was concluded that PYGB silencing increased ROS
production in PC3 cells, which may further cause increased cell
apoptosis of PC3 cells.
It has previously been reported that NF-κB is
involved in the growth, invasion and apoptosis of human prostate
cancer cells (29–32). Additionally, previous studies have
also demonstrated the roles that Nrf2 served in prostate cancer
(33–35). It was also demonstrated that NF-κB
and Nrf2 possessed anti-inflammatory and anti-oxidative activities
(36). Furthermore, the NF-κB and
Nrf2 signaling pathways have been confirmed to contribute to the
inhibition of colorectal carcinogenesis and prevent breast cancer
(37,38). However, the roles and underlying
mechanism of the NF-κB/Nrf2 signaling pathway in prostate cancer
are not clear. Thus, the present study investigated the expression
levels of NF-κB and Nrf2 in PC3 cells treated with si-PYGB. It was
revealed that PYGB silencing significantly upregulated the
expression levels of NF-κB in PC3 cells. Nrf2 expression in PC3
cells was reduced by PYGB silencing. Therefore, it was concluded
that PYGB silencing affected the NF-κB/Nrf2 signaling pathway in
PC3 cells.
Taken together, the results of the present study
demonstrated that PYGB silencing suppressed the growth and promoted
the apoptosis of prostate cancer cells by affecting the NF-κB/Nrf2
signaling pathway. This may provide a novel research focus for
understanding the pathogenesis of prostate cancer, and may aid the
diagnosis and therapy of prostate cancer.
However, further study is required to confirm the
role of PYGB in prostate cancer. For example, the data of the
present study would be supported by PYGB overexpression
experiments, to further examine its effect on the growth and
apoptosis of prostate cancer cells in vitro, or by exploring
the effect of PYGB on prostate cancer progression in vivo.
Furthermore, studies with larger sample sizes should be
performed.
In conclusion, the present study highlighted that
PYGB silencing suppressed the growth and promoted the apoptosis of
prostate cancer cells by affecting the NF-κB/Nrf2 signaling
pathway. The findings of the present study may influence the
understanding of the underlying mechanisms of PYGB and prostate
cancer cells. The potential effects of PYGB on the growth and
apoptosis of prostate cancer cells suggested that PYGB may be an
effective target for anti-tumor therapies.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
ZW and GH wrote the manuscript. ZW, GH, QL and WZ
performed the experiments. ZW and JW designed the study. GH and QL
performed the data analysis. ZW, GH and JW revised the manuscript.
All authors reviewed the manuscript.
Ethics approval and consent to
participate
All patients recruited to the present study provided
written informed consent for the utilization of their tissue
samples for clinical research. The project protocol was approved by
the Institutional Review Board of East Hospital, Tongji University
School of Medicine (Shanghai, China).
Patient consent for publication
Informed consent was obtained from all participants
for the publication of their data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen W, Zheng R, Zeng H, Zhang S and He J:
Annual report on status of cancer in China, 2011. Chin J Cancer
Res. 27:2–12. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cai C, Chen S, Ng P, Bubley GJ, Nelson PS,
Mostaghel EA, Marck B, Matsumoto AM, Simon NI, Wang H, et al:
Intratumoral de novo steroid synthesis activates androgen receptor
in castration-resistant prostate cancer and is upregulated by
treatment with CYP17A1 inhibitors. Cancer Res. 71:6503–6513. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Steeg PS: Metastasis suppressors alter the
signal transduction of cancer cells. Nat Rev Cancer. 3:55–63. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Baldwin AS Jr: The NF-kappa B and I kappa
B proteins: New discoveries and insights. Annu Rev Immunol.
14:649–683. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dutta J, Fan Y, Gupta N, Fan G and Gélinas
C: Current insights into the regulation of programmed cell death by
NF-kappaB. Oncogene. 25:6800–6816. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Clarkson RW and Watson CJ: NF-kappaB and
apoptosis in mammary epithelial cells. J Mammary Gland Biol
Neoplasia. 4:165–175. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ma Q: Role of nrf2 in oxidative stress and
toxicity. Annu Rev Pharmacol Toxicol. 53:401–426. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Baird L and Dinkova-Kostova AT: The
cytoprotective role of the Keap1-Nrf2 pathway. Arch Toxicol.
85:241–272. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li W and Kong AN: Molecular mechanisms of
Nrf2-mediated antioxidant response. Mol Carcinog. 48:91–104. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Boyanapalli SS, Paredes-Gonzalez X,
Fuentes F, Zhang C, Guo Y, Pung D, Saw CL and Kong AN: Nrf2
knockout attenuates the anti-inflammatory effects of phenethyl
isothiocyanate and curcumin. Chem Res Toxicol. 27:2036–2043. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cheung KL, Lee JH, Khor TO, Wu TY, Li GX,
Chan J, Yang CS and Kong AN: Nrf2 knockout enhances intestinal
tumorigenesis in Apc(min/+) mice due to attenuation of
anti-oxidative stress pathway while potentiates inflammation. Mol
Carcinog. 53:77–84. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu GH, Qu J and Shen X: NF-kappaB/p65
antagonizes Nrf2-ARE pathway by depriving CBP from Nrf2 and
facilitating recruitment of HDAC3 to MafK. Biochim Biophys Acta.
1783:713–727. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ziady AG, Sokolow A, Shank S, Corey D,
Myers R, Plafker S and Kelley TJ: Interaction with CREB binding
protein modulates the activities of Nrf2 and NF-κB in cystic
fibrosis airway epithelial cells. Am J Physiol Lung Cell Mol
Physiol. 302:L1221–L1231. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Newgard CB, Hwang PK and Fletterick RJ:
The family of glycogen phosphorylases: Structure and function. Crit
Rev Biochem Mol Biol. 24:69–99. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rich SS, Goodarzi MO, Palmer ND, Langefeld
CD, Ziegler J, Haffner SM, Bryer-Ash M, Norris JM, Taylor KD,
Haritunians T, et al: A genome-wide association scan for acute
insulin response to glucose in Hispanic-Americans: The insulin
resistance atherosclerosis family study (IRAS FS). Diabetologia.
52:1326–1333. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shimada S, Maeno M, Akagi M, Hatayama I,
Sato T and Sato K: Immunohistochemical detection of glycogen
phosphorylase isoenzymes in rat and human tissues. Histochem J.
18:334–338. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Barbosa AJ and Castro LP: BGP expression
in gastric epithelium and early gastric cancer. Gastric Cancer.
5:123–124. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lee MK, Kim JH, Lee CH, Kim JM, Kang CD,
Kim YD, Choi KU, Kim HW, Kim JY, Park DY, et al:
Clinicopathological significance of BGP expression in
non-small-cell lung carcinoma: Relationship with histological type,
microvessel density and patients' survival. Pathology. 38:555–560.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shiomori K, Shimada S, Marutsuka T,
Hatayama I and Ogawa M: Genetic pathways of ‘de novo’ colorectal
carcinomas with reference to fetal-type glycogen phosphorylase
positive foci. Int J Oncol. 22:65–74. 2003.PubMed/NCBI
|
|
20
|
Tashima S, Shimada S, Yamaguchi K, Tsuruta
J and Ogawa M: Expression of brain-type glycogen phosphorylase is a
potentially novel early biomarker in the carcinogenesis of human
colorectal carcinomas. Am J Gastroenterol. 95:255–263. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim SY, Nishioka M, Hayashi S, Honda H,
Kobayashi T and Taya M: The gene yggE functions in restoring
physiological defects of Escherichia coli cultivated under
oxidative stress conditions. Appl Environ Microbiol. 71:2762–2765.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Uno K, Shimada S, Tsuruta J, Matsuzaki H,
Tashima S and Ogawa M: Nuclear localization of brain-type glycogen
phosphorylase in some gastrointestinal carcinoma. Histochem J.
30:553–559. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee CH, Son HI, Kim JH, Choi KU, Kim JY,
Park DY and Sol MY: Clinicopathologic significance of brain-type
glycogen phosphorylase expression in non-small-cell lung
carcinomas. Lung Cancer. S41–S42. 2004.PubMed/NCBI
|
|
25
|
Takashi M, Koshikawa T, Kurobe N and Kato
K: Elevated concentrations of brain-type glycogen phosphorylase in
renal cell carcinoma. Jpn J Cancer Res. 80:975–980. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sato K, Morris HP and Weinhouse S:
Phosphorylase: A new isozyme in rat hepatic tumors and fetal liver.
Science. 178:879–881. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Royle JS, Ross JA, Ansell I, Bollina P,
Tulloch DN and Habib FK: Nitric oxide donating nonsteroidal
anti-inflammatory drugs induce apoptosis in human prostate cancer
cell systems and human prostatic stroma via caspase-3. J Urol.
172:338–344. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tang DG, Li L, Zhu Z and Joshi B:
Apoptosis in the absence of cytochrome c accumulation in the
cytosol. Biochem Biophys Res Commun. 242:380–384. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hafeez BB, Siddiqui IA, Asim M, Malik A,
Afaq F, Adhami VM, Saleem M, Din M and Mukhtar H: A dietary
anthocyanidin delphinidin induces apoptosis of human prostate
cancer PC3 cells in vitro and in vivo: Involvement of nuclear
factor-kappaB signaling. Cancer Res. 68:8564–8572. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lü L, Tang D, Wang L, Huang LQ, Jiang GS,
Xiao XY and Zeng FQ: Gambogic acid inhibits TNF-α-induced invasion
of human prostate cancer PC3 cells in vitro through PI3K/Akt and
NF-κB signaling pathways. Acta Pharmacol Sin. 33:531–541. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Raj GV, Sekula JA, Guo R, Madden JF and
Daaka Y: Lysophosphatidic acid promotes survival of
androgen-insensitive prostate cancer PC3 cells via activation of
NF-kappaB. Prostate. 61:105–113. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shaikh IA, Brown I, Schofield AC, Wahle KW
and Heys SD: Docosahexaenoic acid enhances the efficacy of
docetaxel in prostate cancer cells by modulation of apoptosis: The
role of genes associated with the NF-kappaB pathway. Prostate.
68:1635–1646. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jayakumar S, Kunwar A, Sandur SK, Pandey
BN and Chaubey RC: Differential response of DU145 and PC3 prostate
cancer cells to ionizing radiation: Role of reactive oxygen
species, GSH and Nrf2 in radiosensitivity. Biochim Biophys Acta.
1840:485–494. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nair S, Barve A, Khor TO, Shen GX, Lin W,
Chan JY, Cai L and Kong AN: Regulation of Nrf2- and AP-1-mediated
gene expression by epigallocatechin-3-gallate and sulforaphane in
prostate of Nrf2-knockout or C57BL/6J mice and PC-3 AP-1 human
prostate cancer cells. Acta Pharmacol Sin. 31:1223–1240. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang C, Su ZY, Khor TO, Shu L and Kong
AN: Sulforaphane enhances Nrf2 expression in prostate cancer TRAMP
C1 cells through epigenetic regulation. Biochem Pharmacol.
85:1398–1404. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Surh YJ: NF-kappa B and Nrf2 as potential
chemopreventive targets of some anti-inflammatory and antioxidative
phytonutrients with anti-inflammatory and vantioxidative
activities. Asia Pac J Clin Nutr. 17 Suppl 1:S269–S272. 2008.
|
|
37
|
Mandal A, Bhatia D and Bishayee A:
Anti-Inflammatory Mechanism Involved in Pomegranate-Mediated
Prevention of Breast Cancer: The Role of NF-κB and Nrf2 Signaling
Pathways. Nutrients. 9(pii): E4362017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yao J, Zhao L, Zhao Q, Zhao Y, Sun Y,
Zhang Y, Miao H, You QD, Hu R and Guo QL: NF-κB and Nrf2 signaling
pathways contribute to wogonin-mediated inhibition of
inflammation-associated colorectal carcinogenesis. Cell Death Dis.
5:e12832014. View Article : Google Scholar : PubMed/NCBI
|