Introduction
Atopic dermatitis (AD) is a common chronic
inflammatory and pruritic skin disease characterized by eczematous,
dry, and chapped skin (1,2). AD is caused by the invasion of
inflammatory cells such as the mast cells, eosinophils,
monocytes/macrophages and T lymphocytes into the skin barrier
(3). AD is also reported that the
development is increased levels of circulating eosinophils and
serum immunoglobulin E (IgE) owing to elevated manufacture of
interleukin (IL)-4, IL-5, and IL-13 by the Th2 cells in most
patients (4,5). IgE plays an important role in
instantaneous hypersensitivity responses by cross-linking to the
high-affinity IgE receptor (FcεRI) expressed on the mast cells and
basophils. The antigen-specific IgE binding to FcRI is the binding
of multivalent antigens of allergens that contribute to allergic
diseases, leading to the release of inflammatory mediators such as
histamine, arachidonic acid metabolites and cytokines by activating
mast cells (4,5).
Mast cells, inflammatory cells that respond to
innate and adaptive immunity, can be described as allergic effector
cells that secrete numerous cytokines associated with chronic skin
inflammation (6,7). They induce the production of
inflammatory mediators, such as histamine, chemokines, and
cytokines, under degranulation and immunostimulation (8). These inflammatory mediators
significantly influence the development of AD (9). It has been demonstrated in several
studies that thymic stromal lymphopoietin (TSLP) is associated with
the development, maintenance, and enhancement of AD (10–12).
It has been identified as an activation factor for itching; a
characteristic of AD. TSLP plays a critical role in the activation
of dendritic cells that prime human CD4+ T cells as Th2
cytokine-generating cells within the local lymph nodes (13–15).
TSLP signaling in the CD4+ T cells is required for the formation of
memory following Th2 sensitization (16). It activates the innate lymphoid
cells, a significant players in the pathogenesis of several
inflammatory skin diseases, including AD (17).
Filaggrin plays an important structural and
functional role in the epidermis, and has a significant effect on
skin homeostasis (18). It is
instrumental in maintaining skin hydration, by preserving the
integrity of the stratum corneum, and in the production of natural
moisturizing factors (19).
Inherited or acquired filaggrin deficiency has been described as a
key contributor to the pathogenesis of AD (18). Thus, the inhibition of production
of these components constitutes a promising and important modality
in the treatment of patients with allergic disease, especially
those with AD.
A number of treatments are available for AD,
including the use of emollients, dexamethasone, topical
glucocorticoids, calcineurin inhibitors, and immunosuppressants,
such as cyclosporine A; all of which are effective in reducing
inflammation. However, they are associated with various
side-effects (1,2). Therefore, the need for the
development of new pharmacological agents that control the
inflammatory responses, with reduced side-effects and higher
efficacy, has been identified.
Bee venom (BV) is a natural toxin produced by
honeybees (Apis mellifera L.). BV contains various peptides
including melittin, apamin, adolapin, and mast cell degranulating
peptide along with enzymes, biological amines, and non-peptide
components (20). BV is widely
used as a traditional medicine for various diseases (20,21).
The treatment of inflammatory diseases has been investigated in
several studies (22–24). The anticancer properties of BV have
also been demonstrated in lung and breast cancer, hepatocellular
carcinoma, and prostate cancer cells (25–27).
In previous study, we demonstrated that melittin of main BV
components has beneficial effects on atopic-dermatitis (28). However, the anti-inflammatory
effects of BV on ovalbumin (OVA)-induced inflammatory skin disease
in an animal model have not been reported to date. Thus, the
current study objective was to evaluate the therapeutic effects of
BV as an alternative to anti-inflammatory therapy for the treatment
of AD.
Materials and methods
BV collection
Animal care and all experimental procedures were
approved by the Institutional Animal Care and Use Committee of
Catholic University of Daegu (Daegu, Korea; approval no.
DCIAFCR-160428-1-Y) and experiments were performed in accordance
with these institutional guidelines. The natural honeybee colonies
used in the present study were maintained at the National Academy
of Agricultural Science (Suwon, Korea). BV was collected with the
aid of a collecting device (Chung Jin Biotech Co., Ltd., Ansan,
Korea) in a sterile manner under strict laboratory conditions. In
brief, the BV collector device was placed in the hive and the bees
were administered electric shocks to caused them to sting a glass
plate from which dried BV was later removed by scraping. The
collected venom was purified using the method described by Han
et al (29) The BV was then
stored in a refrigerator for later use.
Animals and induction of AD
Six-week-old female BALB/c mice (n=25) were
purchased from Samtako (Osan, Korea) and housed at 22±2°C and 55%
humidity in a 12 h light-dark cycle, and allowed food and water ad
libitum. The mice were equilibrated for seven days prior to the
induction of AD and randomly divided into five groups (5 mice per
group). These were divided into 5 groups as follows: the normal
control (NC), OVA-induced AD with no treatment (OVA), and
OVA-induced AD with various concentrations (1, 10 and 100 µg/kg of
weight) of BV-treatment (BV1, BV10 and BV100).
Induction of AD was performed according to the
method given by Kim et al (28) Briefly, All the mice, with the
exception of those in the NC group, were intraperitoneally
inoculated mixed with 10 µg of chicken OVA (grade V)
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) mixed with 4 mg of
aluminum hydroxide (ImjectAlum®; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) in a volume of 200 µl three
time at intervals of one-week intervals (i.e., on days 0, 7, and
14). They were then anesthetized by isoflurane inhalation
(Ifran®; HANA Pharm, Seoul, Korea). During anesthesia,
the dorsal skin was shaved. After shaving, the mice were sensitized
with OVA patches on day 14. The OVA patches were prepared as 100 µg
of OVA in phosphate-buffered saline (PBS). The OVA patches were
attached to the shaved skin for seven days (i.e., from days 14 to
20) and changed daily. Thereafter, the indicated concentration of
BV (1, 10, and 100 µg/kg of weight) was administrated twice a week
intraperitoneally. Following the BV treatment, the OVA patches were
reattached for a week. After the OVA treatment with patches, their
blood was collected by cardiac puncture, and the mice were
sacrificed by CO2 asphyxiation. After sacrifice, their
dorsal skins were excised for the next experiments.
Histologic analysis
The dorsal skin specimens were fixed in 10% neutral
buffered formalin for at least 24 h at room temperature. The fixed
skin specimens were dissected, dehydrated, and embedded in
paraffin. Thereafter, thin sections (4 µm) were mounted on glass
slides and stained with hematoxylin and eosin (H&E) and giemsa.
All slides were examined under a Pannoramic® MIDI slide
scanner (3DHISTECH Ltd., Budapest, Hungary). The number of
infiltrated and degranulation mast cells was counted at
magnification, ×400 with CaseViewer® 1.4 software
(3DHISTECH Ltd.).
Immunohistochemical staining
The paraffin-embedded tissue sections were
deparaffinized with xylene, dehydrated in gradually decreasing
concentrations of ethanol, and then subsequently treated with 3%
hydrogen peroxidase in methanol for 10 min to block endogenous
peroxidase activity. The tissue sections were immersed in 10 mM
sodium citrate buffer (a pH of 6.0) for 5 min at 95°C. The final
step was repeated using 10 mM sodium citrate solution (a pH of
6.0). The sections in the same solution were left to cool at room
temperature for 20 min, rinsed in PBS, and then incubated with
primary antibody (1:100 dilution) for 1 h at 37°C. Anti-CD4 and
anti-CD11b antibodies (Abcam, Cambridge, UK) were used as the
primary antibody. The signal was visualized using an Envision
System® (Dako; Agilent Technologies, Inc., Santa Clara,
CA, USA) for 30 min at 37°C. 3,3′-diaminobenzidine
tetrahydrochloride (DAB) was used as the coloring reagent, and
hematoxylin was used as a counter stain. The slides were inspected
with a Pannoramic MIDI slide scanner and the integrated optical
density was analyzed with the i-Solution DT software.
Immunofluorescence staining
The paraffin-embedded dorsal skin specimens were
deparaffinized with xylene and dehydrated in gradually decreasing
concentrations of ethanol. The tissue sections were subsequently
placed in blocking serum (5% bovine serum albumin in PBS) at room
temperature for 1 h. The primary antibody (1:100 dilution) was
added followed by overnight incubation at 4°C, and secondary
antibody (1:200 dilution) was performed for 4 h at room
temperature. The nuclei were stained with Hoechst 33342®
solution for 20 min. The antibodies included filaggrin (cat. no.
ENZ-ABS181; Enzo Life Sciences, Lausen, Switzerland) and
anti-rabbit-biotinylated secondary antibodies conjugated with
fluorescein isothiocyanate (Invitrogen; Thermo Fisher Scientific,
Inc.). The slides were mounted using a fluorescence mounting medium
(Dako; Agilent Technologies, Inc.). The stained slides were imaged
using a NIKON® A1+ confocal microscope (Nikon, Tokyo,
Japan).
Enzyme-linked immunosorbent assay
(ELISA)
The concentrations of IgE, tumor necrosis factor
(TNF)-α, and TSLP in the collected sera of the blood were
determined by ELISA kit, according to the manufacturer's
instructions (IgE; Bethyl Laboratories, Montgomery, TX, USA) and
R&D Systems (TNF-α and TSLP). The optical density was measured
at 450 nm using an ELISA reader (BMG Labtech, Baden-Württemberg,
Germany).
Statistical analysis
All the data were presented as the mean ± standard
error of the mean. Statistical significance was determined by
one-way analysis of variance with Tukey's multiple comparison test
using GraphPad Prism 5.0 (GraphPad Software, Inc., La Jolla, CA,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Effect of BV treatment on dorsal skin
thickness according to histopathologic analysis
As shown in Fig.
1A, the AD-like skin lesions induced by the OVA were clearly
evident on the dorsal skin of the Balb/c mice. Various pathological
features, such as bleeding, erythema, eczema, and dryness, were
observed in the OVA group. By contrast, it was macroscopically
confirmed that pathological features were significantly reduced in
the BV group. Inflammatory cells infiltration and skin thickness
were also measured in the epidermis and dermis of the mice skin by
H&E staining (Fig. 1B and C).
As a result, infiltration by the inflammatory cells in the OVA
group was significantly increased compared with that in the NC
group, and the skin thickness was also significantly increased. By
contrast, the extent of inflammatory cell infiltration and skin
thickness was significantly decreased in the BV treatment group,
and constituted the greatest decrease, especially in BV 100 group
(Fig. 1C). Animal care and all
experimental procedures were approved and implemented in accordance
with the guidelines of the Institutional Animal Care and Use
Committee Catholic University of Daegu (approval no.
DCIAFCR-160428-1-Y).
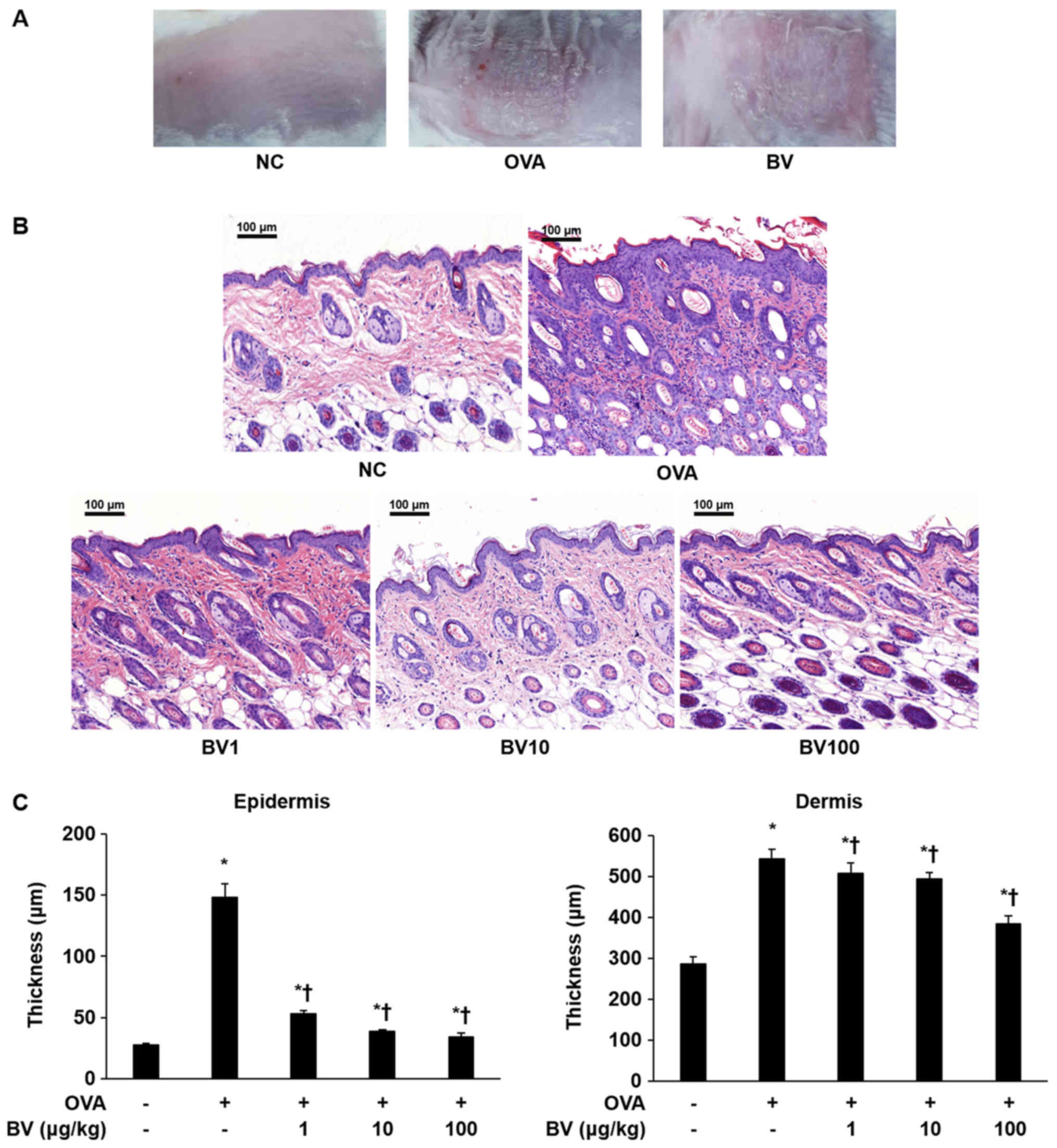 | Figure 1.BV inhibits inflammation in
OVA-induced inflammatory skin disease. OVA was used to induce
atopic dermatitis-like inflammatory skin disease through
intraperitoneal inoculation and patch attachment. BV was
intraperitoneally inoculated during OVA induction. (A) Skin lesions
of each group. (B) Hematoxylin and eosin staining: Representative
images of histological analysis, exhibiting increased epidermal and
dermal thickness, and increased number of invasive inflammatory
cells in the dorsal skin specimens; however, this increase was
reduced in the BV administration group. Magnification, ×200; scale
bars, 100 µm. (C) The thickness of the epidermis and dermis was
measured. The results are expressed as the mean ± standard error of
the mean of least 10 random fields per section. *P<0.05 vs. NC
group; †P<0.05 vs. OVA group. BV, bee venom; OVA,
ovalbumin; BV group, OVA with BV treatment (1, 10 and 100 µg/kg);
NC, normal control; OVA, ovalbumin; BV, bee venom. |
Effect of BV treatment on mast cell
and IgE expression
To investigate the inhibitory effect of the BV
treatment, infiltration and degranulation of mast cells was
examined, and IgE levels were measured. The extent to which mast
cell infiltration and degranulation occurred was measured at
magnification, ×400 (Fig. 2A). It
was determined that the extent of mast cell infiltration and
degranulation was significantly increased in the OVA group, but
diminished in the BV treatment group, according to the
concentration level applied. Notably, it was observed that the BV
100 group was significantly reduced (Fig. 2B). The escalation in the serum IgE
level was shown to dramatically increased in the OVA group using
ELISA. Similar to the measurement results of mast cell infiltration
and degranulation extent, the serum IgE levels identified by ELISA
were found to be reduced in the BV 100 group (Fig. 2C).
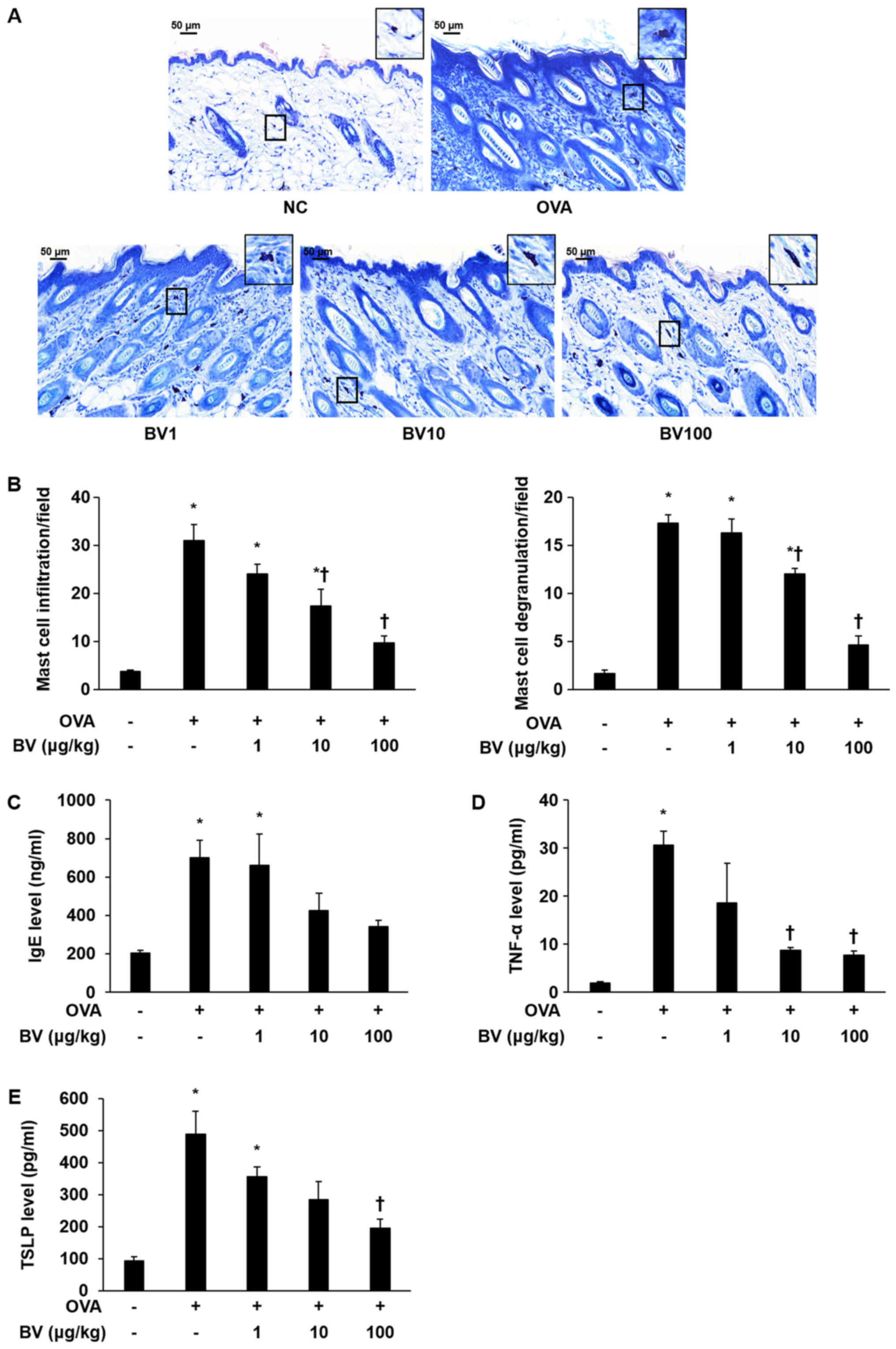 | Figure 2.BV reduces OVA-induced mast cells,
serum IgE and pro-inflammatory cytokine expression. (A) Giemsa
staining: Representative images of histological analysis,
presenting increased infiltration and degranulation of the mast
cells. However, it was reduced under BV treatment. Magnification,
×400; scale bars, 50 µm. (B) Infiltration and degranulation were
evaluated by assessing the number of mast cells from at least 3
random fields per section at 400-fold magnification. (C) The ELISA
results demonstrated that BV inhibited the increased levels of
serum-IgE induced by OVA. In addition, BV inhibited the increased
expression of inflammatory cytokines such as (D) TNF-α and (E) TSLP
induced by OVA. The results are expressed as the mean ± standard
error of the mean of 3 independent determinations. *P<0.05 vs.
NC group; †P<0.05 vs. OVA group. BV, bee venom; OVA,
ovalbumin; BV group, OVA with BV treatment (1, 10 and 100 µg/kg);
NC, normal control; IgE, immunoglobulin; TNF-α, tumor necrosis
factor-α; TSLP, thymic stromal lymphopoietin. |
Effect of BV treatment on
pro-inflammatory cytokine expression
TNF-α and TSLP levels were measured in the mouse
serum of the NC, OVA, and BV-treatment group to establish whether
or not BV treatment would induce alterations to the
pro-inflammatory cytokines involved in OVA-induced skin
inflammation. The levels of TNF-α and TSLP were higher in the
OVA-treated group than in the NC group. However, TNF-α, and TSLP
levels were significantly decreased in 100 group (Fig. 2D and E). IL-1β was reduced in BV100
but not statistically significant.
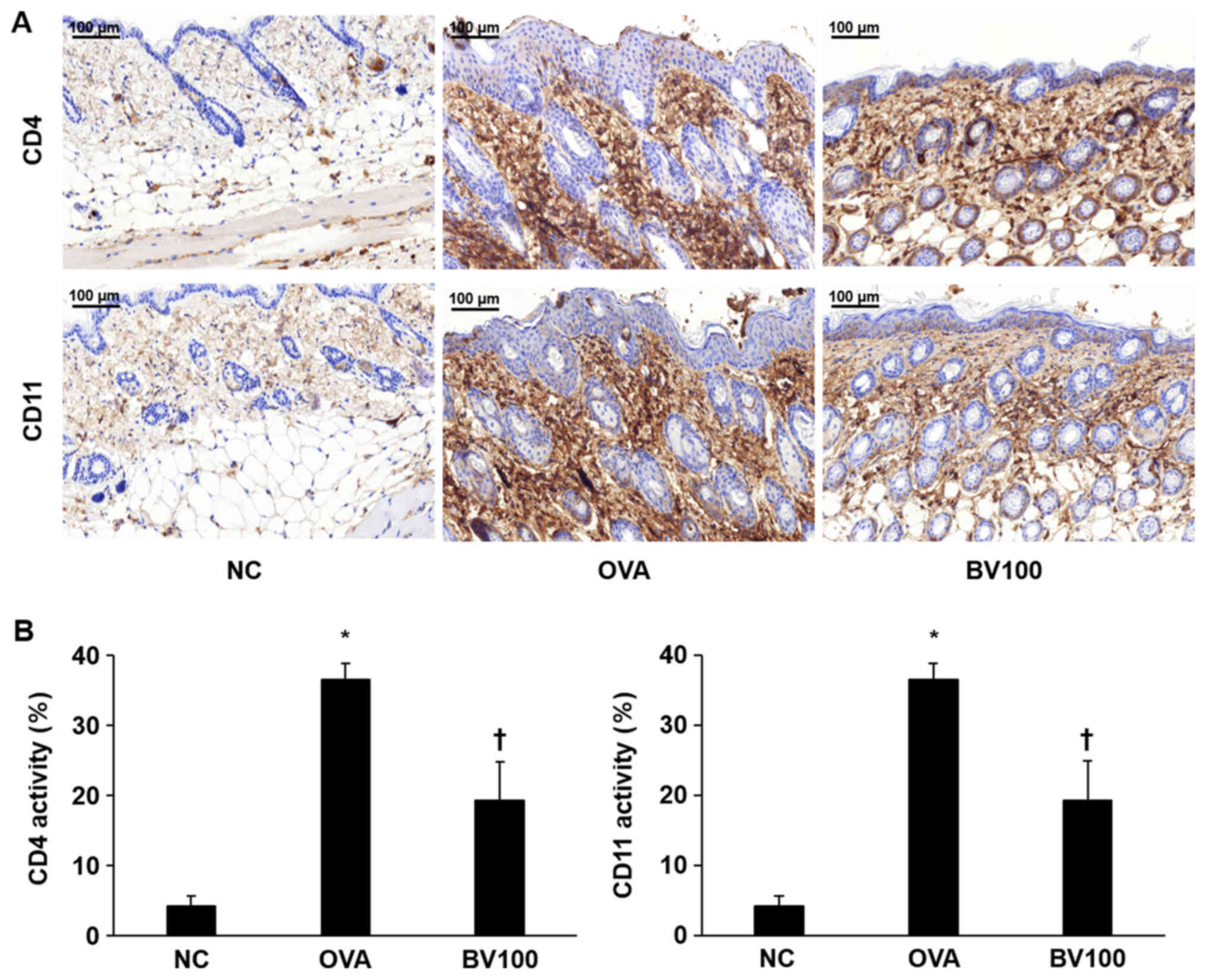
Effect of BV treatment on CD4+ and
CD11b+ expression
Immunohistochemical staining confirmed the extent of
infiltration of the CD4+ T helper cells and CD11b+ known as
monocyte antigen. As shown in Fig.
3A, the expression of CD4+ was significantly increased in the
OVA group. On the other hand, the expression level of CD4+ was
significantly decreased in the BV 100 group compared with OVA
group. Similar to the results of the expression of CD4+, the
expression level of CD11b+ was significantly increased in the OVA
group, and it was also confirmed that the expression level of CD11b
was also significantly decreased in the BV 100 group (Fig. 3B).
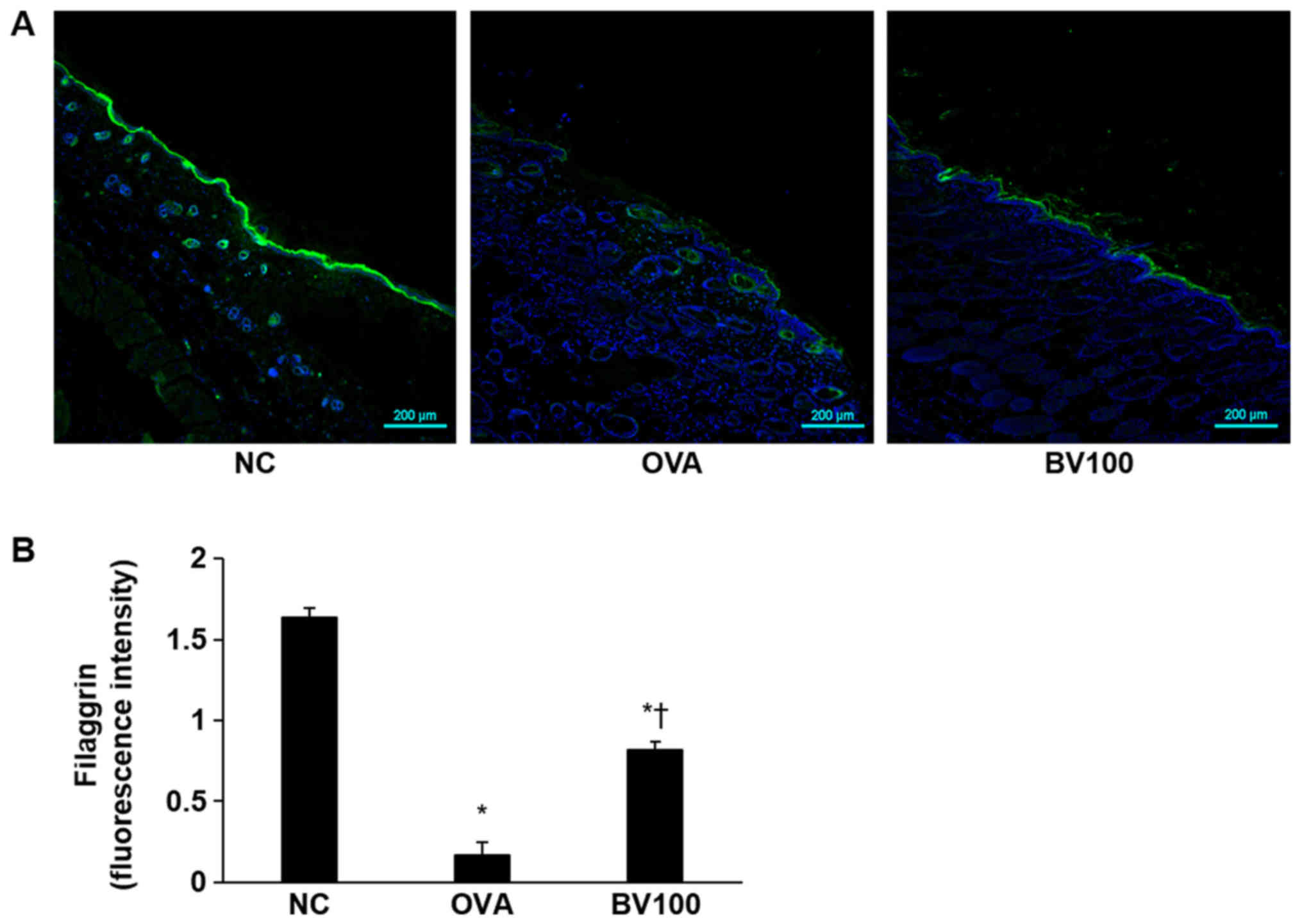
Effect of BV treatment on filaggrin
deficiency
Immunofluorescent staining was performed to
determine the extent of filaggrin deficiency, which plays an
important role in skin homeostasis. As shown in Fig. 4, the expression of filaggrin was
strongly expressed in the NC group, but deficiency was confirmed in
the OVA group. However, the expression of filaggrin was found to
increase in the BV100 group.
Discussion
BV therapy has been used in Oriental medicine since
ancient times to treat and relieve the pain associated with
inflammatory diseases, such as rheumatoid arthritis and multiple
sclerosis (21). Relatively BV of
high concentrations (≥2.5 µg/ml) induce the release of inflammatory
cytokines from the human keratinocytes (30). By contrast, BV of low
concentrations (≤100 ng/ml) reduced pro-inflammatory cytokines in
human keratinocytes infected with bacteria (31).
In previous studies, we reported that BV inhibits
atherosclerotic lesions induced by an intraperitoneal injection of
lipopolysaccharide (32). In
addition, the in vitro antimicrobial activity of BV against
inflammation induced by Propionibacterium acnes was
demonstrated (33). However, the
anti-inflammatory effects of BV on OVA-induced inflammatory skin
disease in an animal model have not yet been reported. Therefore,
we examined whether there were the anti-inflammatory properties of
BV in a mouse model of OVA-induced skin inflammation.
AD has been reported to be characterized by
hyperkeratosis, spongiosis, epidermal hyperplasia, and accumulation
of lymphocyte and mast cell (34).
Therefore, our study examined the effect of BV on the
characteristics of AD. We measured epidermal thickness to assess
the extent of hyperkeratosis, spongiosis, and epidermal
hyperplasia. The markedly increased epidermal thickness in AD-like
skin lesions by OVA has been shown to decrease through BV
treatment. These results suggest that BV inhibits characteristic
symptoms of AD in OVA-induced AD-like skin lesions.
Filaggrin, a major protein involved in the terminal
differentiation of epidermal keratinocytes, has an important
structural and functional role in the epidermis, which is
influential in homeostasis of the skin (18). Variations in the filaggrin
genotype, environmental factors, and skin microenvironment have
been reported as major contributors to reduced levels of filaggrin
(18). Filaggrin deficiency in AD
patients is known to affect epidermal function and increase the
risk of microbial infection or other atopic diseases (18,35).
Therefore, an attempt was made in the current study to determine
whether or not BV treatment would reduce the symptoms of
OVA-induced skin diseases. Cha et al (36) showed that the increase in filaggrin
expression contributes to the maintenance and enhancement of the
skin barrier function. Similarly, our study showed that the
expression of filaggrin deficient by OVA-induced improved the
expression of filaggrin deficient by BV treatment. Based on this
fact, we suggested that BV has an effect that contributes to
maintenance and enhancement of skin barrier function. Based on this
fact, increased expression of filaggrin resulting from BV treatment
was considered to contribute to the maintenance and enhancement of
skin barrier function and was observed to contribute to inhibition
of OVA-induced AD-like skin lesions.
Mast cells are generally known to contribute to
allergic reactions, including atopic skin lesions (34), and are caused by FcεRI-activated
allergens that bind to serum IgE-FcεRI complexes (6,37).
The mast cells, activated by immunologic stimulation, undergo
degranulation and release inflammatory mediators, including
histamine, chemokines, and cytokines (8). Therefore, low IgE levels and
inactivation of the mast cells are important in the alleviation of
AD. It was observed in the current study that IgE expression and
the number of mast cells involved in the inflammatory responses
decreased in the BV-treated, OVA-induced mice AD model. Several
studies have shown that elevated levels of serum IgE and
pro-inflammatory cytokines in AD have been reported (38,39).
In addition, pro-inflammatory cytokines, such as IL-1β, IL-6, and
TNF-α, are known to induce the production of inflammatory
mediators, such as histamine, and protease and chemotactic factors,
in the mast cells (40).
Pro-inflammatory cytokines are produced by inflammatory dendritic
cells, i.e., differentiated mononuclear cells recruited during the
acute AD phase. In our Previously, we demonstrated that inhibition
of pro-inflammatory cytokines by BV treatment in P.
acnes-induced acne skin inhibited inflammatory skin disease
(41). Similar to these results,
BV treatment was demonstrated to reduce the expression of
pro-inflammatory cytokines in OVA-induced AD-like skin lesions in
the current study. These data suggest that it is likely that BV
would be an effective treatment in AD-like skin disease.
It has been shown in several studies that
pro-inflammatory cytokine expression, i.e., interferon (IFN)-γ,
IL-4, IL-13, IL-17, and TSLP, increases in AD skin lesions
(42,43). TSLP, the keratinocyte-derived
cytokine, has been shown to play an important role in the early
stages of allergic inflammation. It was identified as a sensitive
product of keratinocytes released before the lesion skin developed
In AD patients (44). In addition,
it has been shown to activator of sensory neurons that causes
itching; a vital feature of atopic skin (45). Elsewhere, it was demonstrated that
the down-regulation of TSLP improved AD (46). This was similar to the finding of
the current study, where a reduction in TSLP expression was
observed following BV treatment. Therefore, the modulation of
AD-associated TSLP and cytokine expression could result in
therapeutic efficacy. BV was demonstrated to suppress TSLP and
pro-inflammatory cytokine expression in the OVA-induced, AD-like
mice model in the present study. Therefore, the use of BV has
considerable potential as an alternative AD therapy for AD
lesions.
In addition, TSLP binds to the dendritic cell
receptor and promotes the differentiation of CD4+ T cells into Th2
cells (14). CD4+ T cells are key
components of allergic inflammatory disease. CD11b+ is a monocyte
antigen that is expressed on many leukocyte surfaces, including
granulocytes, monocytes, and macrophages (47). Thus, it was deemed worthwhile to
investigate whether or not BV could alleviate AD-like skin lesions
(especially allergic diseases) through an evaluation of CD4+ T and
CD11b+ expression. In this study, it was confirmed that CD4+ T and
CD11b+ expression was markedly decreased following BV treatment.
Thus, BV treatment was demonstrated to have a therapeutic effect on
AD-like inflammatory disease. It can be used to effectively
maintain skin barrier function, prevent the destruction of the skin
barrier, regulate proinflammatory cytokine levels, inhibit mast
cell activation, and reduce inflammation.
BV has an anti-inflammatory effect on AD in this
experiment. In previous study, melittin which is a major component
of BV also showed an anti-inflammatory function for AD (28). It seems to be due to melittin, the
main component of BV. Thus, the pharmacological function of BV
could be similar to that of melittin.
In conclusion, it was demonstrated in the present
study that the administration of BV via intraperitoneal inoculation
effectively suppressed the onset of atopic skin lesions. BV
inhibited cytokines that are influential in AD via a decline in
IgE, TNF-α, and TSLP in vivo, and demonstrated effective
anti-inflammatory activity. Therefore, we propose that BV is an
alternative therapy for the treatment of anti-inflammatory AD.
However, the exact anti-inflammatory mechanism of BV components
requires further investigation.
Acknowledgements
Not applicable.
Funding
The present study was conducted with the support of
the ‘Cooperative Research Program for Agriculture Science and
Technology Development (project no. PJ01316601)’ Rural Development
Administration, Republic of Korea.
Availability of data and materials
The datasets used or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HG, W-HK and K-KP conceived and supervised the
study. HG, H-JA, J-YK, W-HK and M-GG performed the experiments. HG,
SMH and JL performed the data analysis. HG and K-KP drafted the
manuscript. All authors discussed, revised and approved the
manuscript.
Ethics approval and consent to
participate
Animal care and all experimental procedures were
approved by the Institutional Animal Care and Use Committee of
Catholic University of Daegu (approval no. DCIAFCR-160428-1-Y) and
experiments were performed in accordance with these institutional
guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Leung DY and Guttman-Yassky E: Deciphering
the complexities of atopic dermatitis: Shifting paradigms in
treatment approaches. J Allergy Clin Immunol. 134:769–779. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schlapbach C and Simon D: Update on skin
allergy. Allergy. 69:1571–1581. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lim SJ, Kim M, Randy A, Nam EJ and Nho CW:
Effects of Hovenia dulcis Thunb. extract and methyl vanillate on
atopic dermatitis-like skin lesions and TNF-α/IFN-γ-induced
chemokines production in HaCaT cells. J Pharm Pharmacol.
68:1465–1479. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Galli SJ, Tsai M and Piliponsky AM: The
development of allergic inflammation. Nature. 454:445–454. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Owen CE: Immunoglobulin E: Role in asthma
and allergic disease: Lessons from the clinic. Pharmacol Ther.
113:121–133. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kawakami T, Ando T, Kimura M, Wilson BS
and Kawakami Y: Mast cells in atopic dermatitis. Curr Opin Immunol.
21:666–678. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Stone KD, Prussin C and Metcalfe DD: IgE,
mast cells, basophils, and eosinophils. J Allergy Clin Immunol. 125
2 Suppl 2:S73–S80. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kempuraj D, Castellani ML, Petrarca C,
Frydas S, Conti P, Theoharides TC and Vecchiet J: Inhibitory effect
of quercetin on tryptase and interleukin-6 release, and histidine
decarboxylase mRNA transcription by human mast cell-1 cell line.
Clin Exp Med. 6:150–156. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Leung DY, Boguniewicz M, Howell MD, Nomura
I and Hamid QA: New insights into atopic dermatitis. J Clin Invest.
113:651–657. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Miazgowicz MM, Headley MB, Larson RP and
Ziegler SF: Thymic stromal lymphopoietin and the pathophysiology of
atopic disease. Expert Rev Clin Immunol. 5:547–556. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ziegler SF: The role of thymic stromal
lymphopoietin (TSLP) in allergic disorders. Curr Opin Immunol.
22:795–799. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wilson SR, Thé L, Batia LM, Beattie K,
Katibah GE, McClain SP, Pellegrino M, Estandian DM and Bautista DM:
The epithelial cell-derived atopic dermatitis cytokine TSLP
activates neurons to induce itch. Cell. 155:285–295. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Soumelis V, Reche PA, Kanzler H, Yuan W,
Edward G, Homey B, Gilliet M, Ho S, Antonenko S, Lauerma A, et al:
Human epithelial cells trigger dendritic cell mediated allergic
inflammation by producing TSLP. Nat Immunol. 3:673–680. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Watanabe N, Hanabuchi S, Soumelis V, Yuan
W, Ho S, de Waal Malefyt R and Liu YJ: Human thymic stromal
lymphopoietin promotes dendritic cell-mediated CD4+ T cell
homeostatic expansion. Nat Immunol. 5:426–434. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ebner S, Nguyen VA, Forstner M, Wang YH,
Wolfram D, Liu YJ and Romani N: Thymic stromal lymphopoietin
converts human epidermal Langerhans cells into antigen-presenting
cells that induce proallergic T cells. J Allergy Clin Immunol.
119:982–990. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang Q, Du J, Zhu J, Yang X and Zhou B:
Thymic stromal lymphopoietin signaling in CD4(+) T cells is
required for TH2 memory. J Allergy Clin Immunol. 135:781–791.e3.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim BS, Siracusa MC, Saenz SA, Noti M,
Monticelli LA, Sonnenberg GF, Hepworth MR, Van Voorhees AS, Comeau
MR and Artis D: TSLP elicits IL-33-independent innate lymphoid cell
responses to promote skin inflammation. Sci Transl Med.
5:170ra162013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cabanillas B and Novak N: Atopic
dermatitis and filaggrin. Curr Opin Immunol. 42:1–8. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Levin J, Friedlander SF and Del Rosso JQ:
Atopic dermatitis and the stratum corneum: Part 1: The role of
filaggrin in the stratum corneum barrier and atopic skin. J Clin
Aesthet Dermatol. 6:16–22. 2013.PubMed/NCBI
|
|
20
|
Son DJ, Lee JW, Lee YH, Song HS, Lee CK
and Hong JT: Therapeutic application of anti-arthritis,
pain-releasing, and anti-cancer effects of bee venom and its
constituent compounds. Pharmacol Ther. 115:246–270. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Billingham ME, Morley J, Hanson JM,
Shipolini RA and Vernon CA: Letter: An anti-inflammatory peptide
from bee venom. Nature. 245:163–164. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kwon YB, Lee HJ, Han HJ, Mar WC, Kang SK,
Yoon OB, Beitz AJ and Lee JH: The water-soluble fraction of bee
venom produces antinociceptive and anti-inflammatory effects on
rheumatoid arthritis in rats. Life Sci. 71:191–204. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kwon YB, Lee JD, Lee HJ, Han HJ, Mar WC,
Kang SK, Beitz AJ and Lee JH: Bee venom injection into an
acupuncture point reduces arthritis associated edema and
nociceptive responses. Pain. 90:271–280. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Stieger M, Wüthrich B, Wyss S and Kopper
E: Clinical picture and diagnosis of bee-venom allergy. A
comparison between skin tests and RAST determinations. Hautarzt.
29:632–637. 1978.(In German).
|
|
25
|
Ip SW, Liao SS, Lin SY, Lin JP, Yang JS,
Lin ML, Chen GW, Lu HF, Lin MW, Han SM and Chung JG: The role of
mitochondria in bee venom-induced apoptosis in human breast cancer
MCF7 cells. In Vivo. 22:237–245. 2008.PubMed/NCBI
|
|
26
|
Oršolić N: Bee venom in cancer therapy.
Cancer Metastasis Rev. 31:173–194. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Park MH, Choi MS, Kwak DH, Oh KW, Yoon DY,
Han SB, Song HS, Song MJ and Hong JT: Anti-cancer effect of bee
venom in prostate cancer cells through activation of caspase
pathway via inactivation of NF-κB. Prostate. 71:801–812. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim WH, An HJ, Kim JY, Gwon MG, Gu H, Jeon
M, Sung WJ, Han SM, Pak SC, Kim MK and Park KK: Beneficial effects
of melittin on ovalbumin-induced atopic dermatitis in mouse. Sci
Rep. 7:176792017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Han SM, Lee GG and Park KK: Acute dermal
toxicity study of bee venom (Apis mellifera L.) in rats. Toxicol
Res. 28:99–102. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dombrowski Y, Peric M, Koglin S,
Kaymakanov N, Schmezer V, Reinholz M, Ruzicka T and Schauber J:
Honey bee (Apis mellifera) venom induces AIM2 inflammasome
activation in human keratinocytes. Allergy. 67:1400–1407. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim JY, Lee WR, Kim KH, An HJ, Chang YC,
Han SM, Park YY, Pak SC and Park KK: Effects of bee venom against
Propionibacterium acnes-induced inflammation in human keratinocytes
and monocytes. Int J Mol Med. 35:1651–1656. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee WR, Kim SJ, Park JH, Kim KH, Chang YC,
Park YY, Lee KG, Han SM, Yeo JH, Pak SC and Park KK: Bee venom
reduces atherosclerotic lesion formation via anti-inflammatory
mechanism. Am J Chin Med. 38:1077–1092. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Han S, Lee K, Yeo J, Baek H and Park K:
Antibacterial and anti-inflammatory effects of honeybee (Apis
mellifera) venom against acne-inducing bacteria. J Med Plants Res.
4:459–464. 2010.
|
|
34
|
Lee YJ, Kim JE, Kwak MH, Go J, Kim DS, Son
HJ and Hwang DY: Quantitative evaluation of the therapeutic effect
of fermented soybean products containing a high concentration of
GABA on phthalic anhydride-induced atopic dermatitis in
IL-4/Luc/CNS-1 Tg mice. Int J Mol Med. 33:1185–1194. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
van den Oord RA and Sheikh A: Filaggrin
gene defects and risk of developing allergic sensitisation and
allergic disorders: Systematic review and meta-analysis. BMJ.
339:b24332009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cha HY, Ahn SH, Cheon JH, Park SY and Kim
K: Hataedock treatment has preventive therapeutic effects for
atopic dermatitis through skin barrier protection in
Dermatophagoides farinae-induced NC/Nga mice. J Ethnopharmacol.
206:327–336. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Amin K: The role of mast cells in allergic
inflammation. Respir Med. 106:9–14. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Choi EJ, Debnath T, Tang Y, Ryu YB, Moon
SH and Kim EK: Topical application of Moringa oleifera leaf extract
ameliorates experimentally induced atopic dermatitis by the
regulation of Th1/Th2/Th17 balance. Biomed Pharmacother.
84:870–877. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu FT, Goodarzi H and Chen HY: IgE, mast
cells, and eosinophils in atopic dermatitis. Clin Rev Allergy
Immunol. 41:298–310. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sae-Wong C, Mizutani N, Kangsanant S and
Yoshino S: Topical skin treatment with Fab fragments of an
allergen-specific IgG1 monoclonal antibody suppresses
allergen-induced atopic dermatitis-like skin lesions in mice. Eur J
Pharmacol. 779:131–137. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
An HJ, Lee WR, Kim KH, Kim JY, Lee SJ, Han
SM, Lee KG, Lee CK and Park KK: Inhibitory effects of bee venom on
Propionibacterium acnes-induced inflammatory skin disease in an
animal model. Int J Mol Med. 34:1341–1348. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
You CE, Moon SH, Lee KH, Kim KH, Park CW,
Seo SJ and Cho SH: Effects of emollient containing bee venom on
atopic dermatitis: A double-blinded, randomized, base-controlled,
multicenter study of 136 patients. Ann Dermatol. 28:593–599. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liu YJ: Thymic stromal lymphopoietin:
Master switch for allergic inflammation. J Exp Med. 203:269–273.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Comeau MR and Ziegler SF: The influence of
TSLP on the allergic response. Mucosal Immunol. 3:138–147. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jeong HJ, Hong SH, Lee DJ, Park JH, Kim KS
and Kim HM: Role of Ca(2+) on TNF-alpha and IL-6 secretion from
RBL-2H3 mast cells. Cell Signal. 14:633–639. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Han NR, Moon PD, Kim HM and Jeong HJ:
Tryptanthrin ameliorates atopic dermatitis through down-regulation
of TSLP. Arch Biochem Biophys. 542:14–20. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Solovjov DA, Pluskota E and Plow EF:
Distinct roles for the alpha and beta subunits in the functions of
integrin alphaMbeta2. J Biol Chem. 280:1336–1345. 2005. View Article : Google Scholar : PubMed/NCBI
|