Introduction
Hemangiomas (HAs) are benign neoplasms that
proliferate rapidly, they are known to be the most common tumor to
present in infancy (1). It is
reported that 1/10 children will develop HAs, most of which are in
the head or neck region (2). A
previous research revealed that HAs is associated with low birth
weight and premature infants (3).
They examined 973 preterm infants and the results showed that the
frequency of HAs is 12.7%, for the low-birth-weight infants below
1,000 g and below 1,500 g the rate was 15.6 and 22.9%,
respectively. The development of HAs usually include 3 stages, an
early proliferative phase (0–12 month), a plateau phase and an
involution phase (1–8 years). Although HAs are benign, they
required treatment because they might develop complications and
resulted in mortality (4,5). To date, the molecular mechanisms
underlying the occurrence and development of HAs are still
unclear.
MicroRNAs (miRNAs) are a class of 19–24 nt small
non-coding RNAs. It is clear that miRNAs negatively regulate mRNA
translation by binding to the 3′-untranslated region (3′ UTR) of
target mRNAs. Since the first miRNA was identified in 1993, more
and more miRNAs have been discovered; and nearly 1,500 miRNA
sequences have reportedly been found (6). miRNAs play critical role in various
biological processes, including cell growth (7), proliferation (8), differentiation (9) and apoptosis (10). During the last decade, a growing
body of literature has confirmed that the dysregulation of miRNAs
is involved in almost all diseases. A few studies have reported
that miRNAs are aberrantly expressed in HAs. For example, Li et
al (11) found that miR-382 is
upregulated in infantile HA, and that the inhibition of miR-382 by
propranolol prominently inhibits the progression of cells derived
from infantile HA; A meta-analysis identified that miR-939, miR-9
and let-7 family are involved in regulating infantile HA (12); Venneti et al (13) found that miR-9 and miR-200a are
dysregulated in HAs; And miR-143 was found to act as a suppressor
of HA growth by targeting Bcl-2 (14). These findings presented a new
perspective for understanding the molecular mechanisms underlying
the development of HAs.
miR-424 has an important role in regulating
cell-autonomous angiogenesis (15), and studies have reported that
downregulation of miR-424 contributes to the abnormal angiogenesis
in senile HA (16). In this study,
we examined the expression of miR-424, finding that miR-424 was
reduced in HA-derived endothelial cells (HemECs), and demonstrating
that miR-424 might be used to treat HAs, as the restoration of
miR-424 levels in HemECs could effectively inhibit cell growth and
induce apoptosis. Furthermore, we demonstrated that vascular
endothelial growth factor receptor (VEGFR)-2, an important
pro-angiogenic receptor, was a direct target of miR-424. VEGF-R2
was upregulated in HemECs and the overexpression of miR-424 clearly
decreased VEGFR-2 and inhibited the activation of protein kinase B
(AKT)/extracellular signal-regulated kinase (ERK) signaling
cascades. Thus, our data demonstrated that miR-424 serves a
critical role in HA pathogenesis and indicate that it may represent
a therapeutic target in the treatment of HAs.
Materials and methods
Preparation of HA specimens
A case of proliferating-phase infantile HA specimen
was collected from the Department of General Surgery, Affiliated
with Xinhua Hospital (Shanghai, China). The tissue was washed with
pre-cooled PBS (0°C) and HemECs cells were isolated. The
experimental protocol was approved by the Ethics Committee of
Xinhua Hospital. Written informed consent was obtained from the
parents of the patient.
Cell extraction, isolation and
culture
Fresh HA sample was washed with PBS for three times,
and minced with scissors. Then, 0.2% collagenase A was added and
the mixture was maintained at 37°C. After 1 h, the mixture was
removed from the incubator and homogenized using glass
homogenizers. Then the homogenate was filtered through a 40 µm cell
strainer and the suspension was collected. HemECs were isolated
from the suspension using CD31 immuno-magnetic beads and plated on
cell culture plates in Endothelial Cell Medium (ScienCell Research
Laboratories, Inc., San Diego, CA, USA). Human umbilical vein
endothelial cells (HUVECs) used in the experiments were purchased
from the Cell Bank of the Chinese Academy of Sciences (Shanghai,
China), and also cultured and maintained in Endothelial Cell
Medium.
Cell transfection
Human miR-424 mimics, miR-424 inhibitors and the
corresponding negative control miRNA (miR-Ctrl) were purchased from
GenePharma (Shanghai, China). Cells were seeded onto 6-well plates
(2×105 cells per well), 24 h later the cells were transfected with
miR-424 mimics, miR-424 inhibitors or miR-Ctrl using
Oligofectamine™ transfection reagent (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) according to the manufacturer's
protocol.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
For total RNA extraction TRIzol reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) was used. For miRNA extraction, a
mirPremier® microRNA Isolation kit (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) was used. The RNA was quantified by
assessing the absorbance at 260 and 280 nm. Following this,
reverse-transcription was performed using M-MLV reverse
transcriptase (Invitrogen; Thermo Fisher Scientific, Inc.). RT-qPCR
was performed on ABI 7300 (Applied Biosystems, Foster, CA, USA)
using the SYBR Green qPCR Master Mix (Tiangen, Shanghai, China).
GAPDH mRNA or U6 snRNA was used as the endogenous control. All
primers used in our study were as followed: VEGFR-2 forward,
5′-CACTGGTTGTACCTCAGCAC-3′ and reverse, 5′-CGTACCAGAAGACACTTCGT-3′;
GAPDH forward, 5′-GTGAACCATGAGAAGTATGACAA-3′ and reverse,
5′-CATGAGTCCTTCCACGATAC-3 (GenePharma). The thermocycling
conditions for qPCR were as follows: 10 min at 95°C, followed by 40
cycles of 95°C for 15 sec, 56°C for 30 sec, and 70°C for 30 sec.
The 2-∆∆Cq method was used to analyze relative gene expression
(17).
Construction of expression
vectors
3′UTR of VEGFR-2 was amplified using PCR and cloned
into a psiCHECK-2 vector to generate the psiCHECK2-VEGFR-2-3′UTR
(Luc-VEGFR-2-wt). The following primers were used to clone VEGFR-2
3′UTR forward, 5′-CTCGAGAAGGAAGCATCCACACC-3′ and reverse,
5′-GCGGCCGCGTTGTCGAAATGAAAATC-3′. The PCR products were digested
with Xho I and Not I and inserted into a psiCHECK vector. To
generate the psiCHECK-VEGFR-2-mut-3′UTR (Luc-VEGFR-2-mut), a
QuikChange mutagenesis kit (Stratagene; Agilent Technologies GmbH,
Waldbronn, Germany) was used to perform site-directed mutagenesis
of the miR-424 target sites in the VEGFR-2 3′UTR.
Cell proliferation assay
Cell proliferation was assessed using a Cell
Counting kit-8 (Dojindo, Kumamoto, Japan), according to the
manufacturer's instructions. 48 h after transfection with the
indicated miRNAs, cell culture supernatants were discarded, and 10
µl CCK-8 solution was added, After 2 h, the absorbance values at
450 nm were measured. All the experiments were carried out in
triplicate and repeated three times.
Dual luciferase reporter assay
For the dual luciferase reporter assay, HEK293T
cells were seeded onto 96 well-plate (5×103 per well),
After 24 h, the cells were co-transfected with pRL-TK (Promega,
Madison, WI, USA), Luc-VEGFR2-wt or Luc-VEGFR2-mut and miR-424 or
control mimics using lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). 24 h later dual-luciferase assay was performed
according to specification (Promega). Luciferase activity was
measures using a Victor Luminometer (Perkin Elmer, Waltham, MA,
USA). The plasmid of pRL-TK containing Renilla luciferase was used
as internal control. The experiments were carried out in triplicate
and repeated 3 times.
Western blotting analysis
Cells were harvested and lysed in RIPA lysis buffer
(Beyotime Institute of Biotechnology, Shanghai, China). Lysates
were prepared from 1×107 cells by dissolving the cell pellets in
100 µl lysis buffer. Lysates were then centrifuged at 12,000 rpm
for 10 min, and supernatants were transferred to fresh Eppendorf
tubes. The protein concentration was determined using a BCA protein
assay kit (Beyotime Institute of Biotechnology). 30–50 µg protein
was subjected to 10% SDS-PAGE and then electrophoretically
transferred to a PVDF membrane (Millipore, Billerica, MA, USA).
After blocking with blocking buffer (Beyotime Institute of
Biotechnology) for 1 h at room temperature, the membrane was
incubated with the indicated primary antibodies overnight at 4°C.
This was followed by incubation with HRP-conjugated secondary
antibodies for 1 h. Positive signals were visualized using the ECL
Advanced Solution (Bioworld, St. Louis, USA). Actin was used as the
loading control. The antibodies used in this study were as follows:
VEGFR-2 (1:2,000), ERK1/2 (1:2,000), p-ERK1/2 (1:2,000), p-AKT
(1:2,000), AKT (1:2,000), and ACTIN (1:2,000). These antibodies
were purchased from Cell Signaling Technology (Danvers, MA, USA).
All the experiments were repeated three times and the results are
presented as the mean ± SEM.
Apoptosis analysis
After been treated as indicated, the cells were
collected, washed and subjected to apoptosis analysis using an
Annexin V-fluorescein isothiocyanate (FITC) kit (Beyotime Institute
of Biotechnology), according to the manufacturer's instructions.
Cells were analyzed using a FACScan flow cytometer with CellQuest
software (BD Biosciences, Franklin Lakes, NJ, USA). Experiment was
repeated three times and the results are presented as the mean ±
SEM.
Statistical analysis
Differences between two groups were analyzed for
statistical significance by using analysis of variance with a
Tukey-Kramer post hoc test or the Student's t-test. Statistical
analyses were conducted using Prism GraphPad software version 4
(GraphPad Software, Inc., La Jolla, CA, USA). Data are expressed as
the mean ± standard error of the mean. P<0.05 was considered to
indicate a statistically significant difference.
Results
miR-424 expression is downregulated in
HemECs
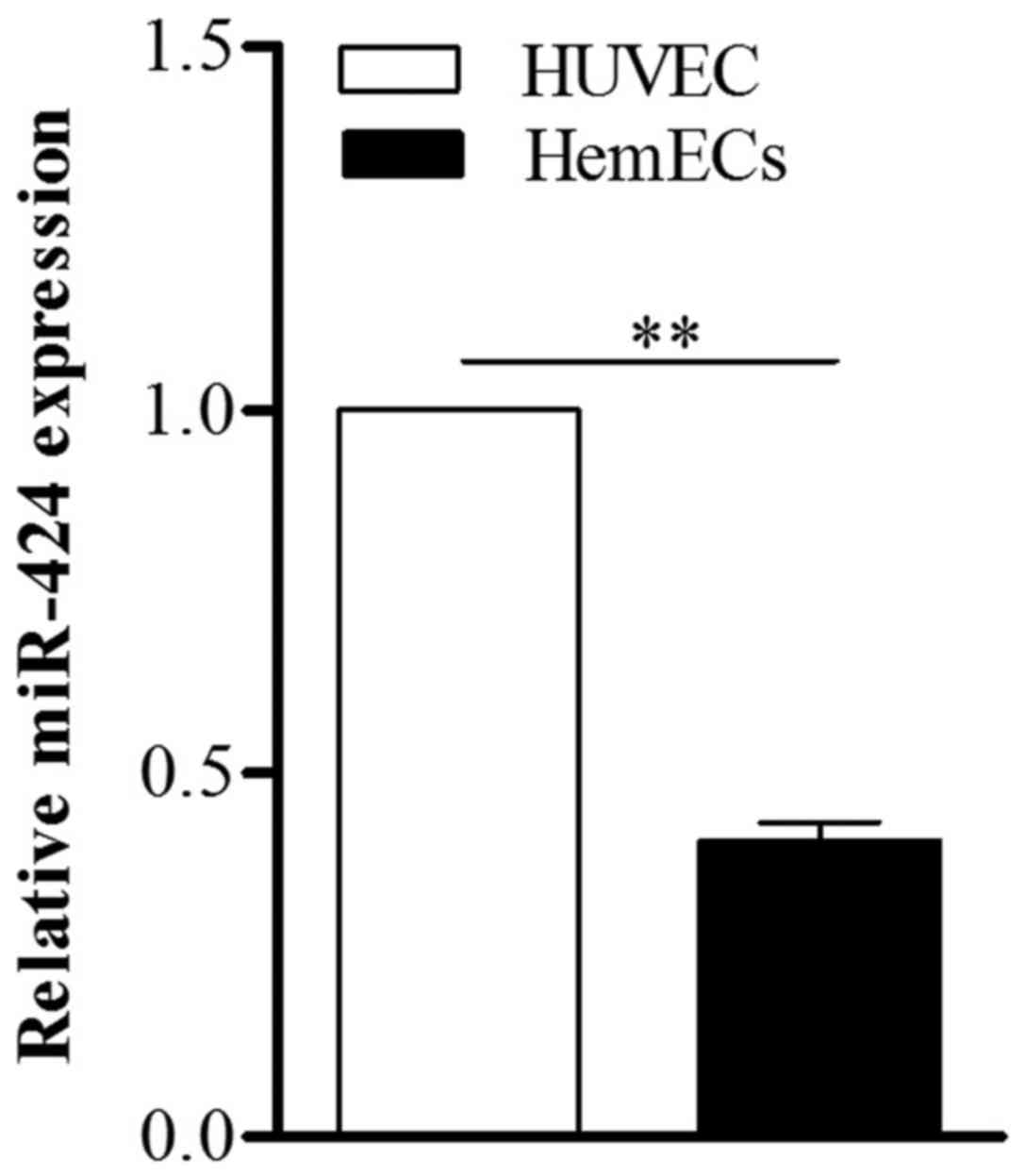
To determine the role of miR-424 in HemECs, we first
used qPCR to detect the expression of miR-424 in HemECs collected
from tissue specimens obtained from a patient with
proliferating-phase infantile HA. Compared to HUVECs, miR-424 was
markedly low in HemECs (Fig.
1).
miR-424 overexpression inhibits cell
proliferation and promotes cell apoptosis in HemECs
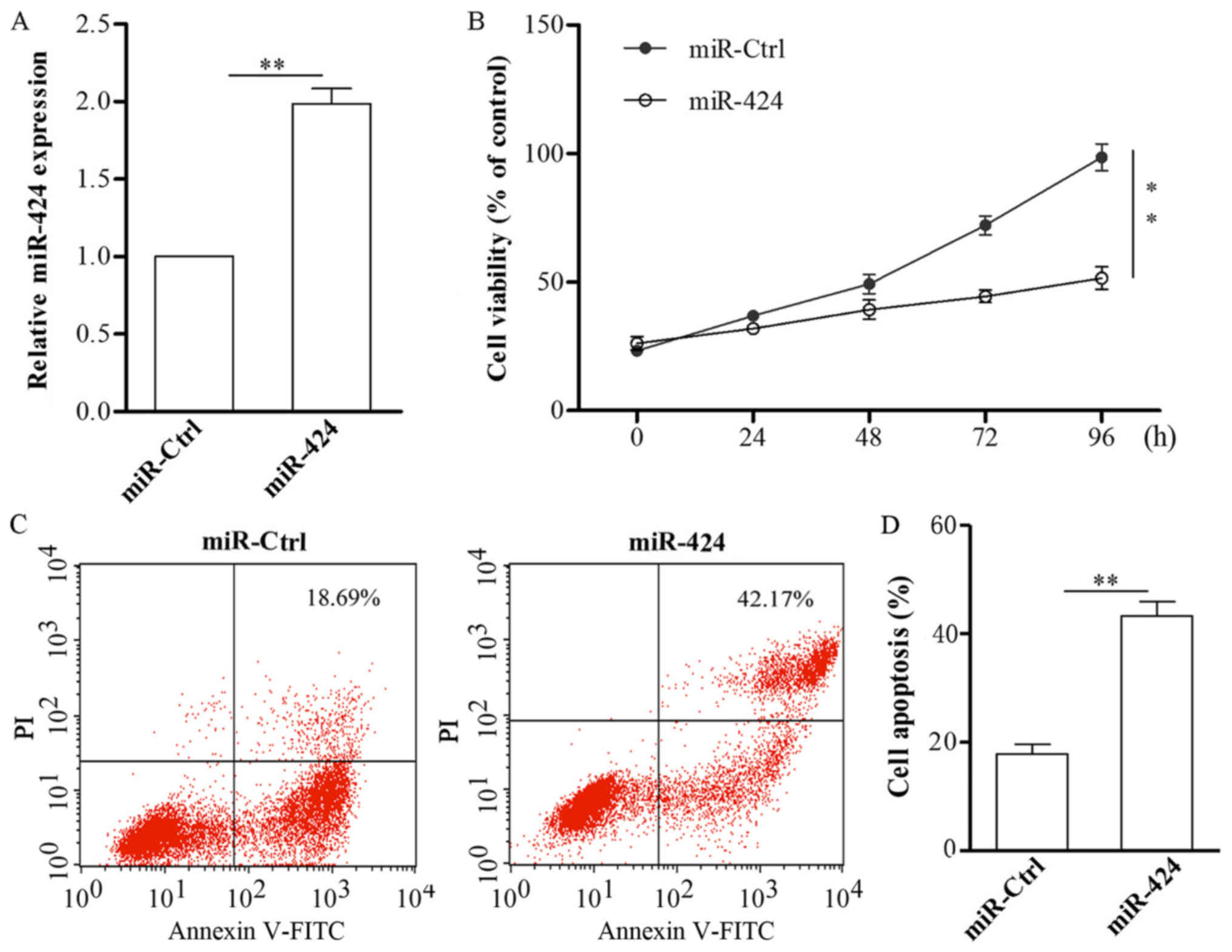
To further examine the possible function of miR-424
during HA development, miR-424 mimics or miR-ctrl were transfected
into HemECs. After transfection with miR-424 mimics, the cells were
collected and subjected to qPCR, the results of which showed that
miR-424 was effectively upregulated in HemECs (Fig. 2A). The results of CCK-8 assay
demonstrated that the upregulation of miR-424 resulted in decreased
HemECs cell viability (Fig.
2B).
Additionally, we explored the possible role of
miR-424 in HemECs apoptosis. miR-424 mimics or miR-ctrl were
transfected into HemECs for 48 h, following which the cells were
collected and subjected to an apoptosis assay. The results
indicated that the overexpression of miR-424 in HemECs markedly
promoted apoptosis (Fig. 2C).
VEGFR2 is a direct target of
miR-424
Next, we explored how miR-424 exerted an inhibitory
effect on HemECs proliferation, we used online miRNA target
prediction databases (www.targetscan.org) to identify the targets of miR-424
in HemECs. The results showed that VEGFR-2 might be one such target
(Fig. 3A). To further study
whether VEGFR-2 was a direct target of miR-424, we performed a
luciferase reporter gene assay. The wt-VEFGR2-3′UTR plasmid,
mut-VEFGR2-3′UTR plasmid and miR-424/miR-Ctrl were co-transfected
into HEK293T cells. As shown in Fig.
3B, compared with the cells co-transfected with miR-Ctrl, the
luciferase activities of miR-424-transfected cells were suppressed
by ~68%, while those cells co-transfected with the mut-VEFGR2-3′UTR
plasmid did not exhibit altered luciferase activity. The
aforementioned data indicate that miR-424 regulates VEGFR-2
expression through direct targeting the 3′UTR of VEGFR2.
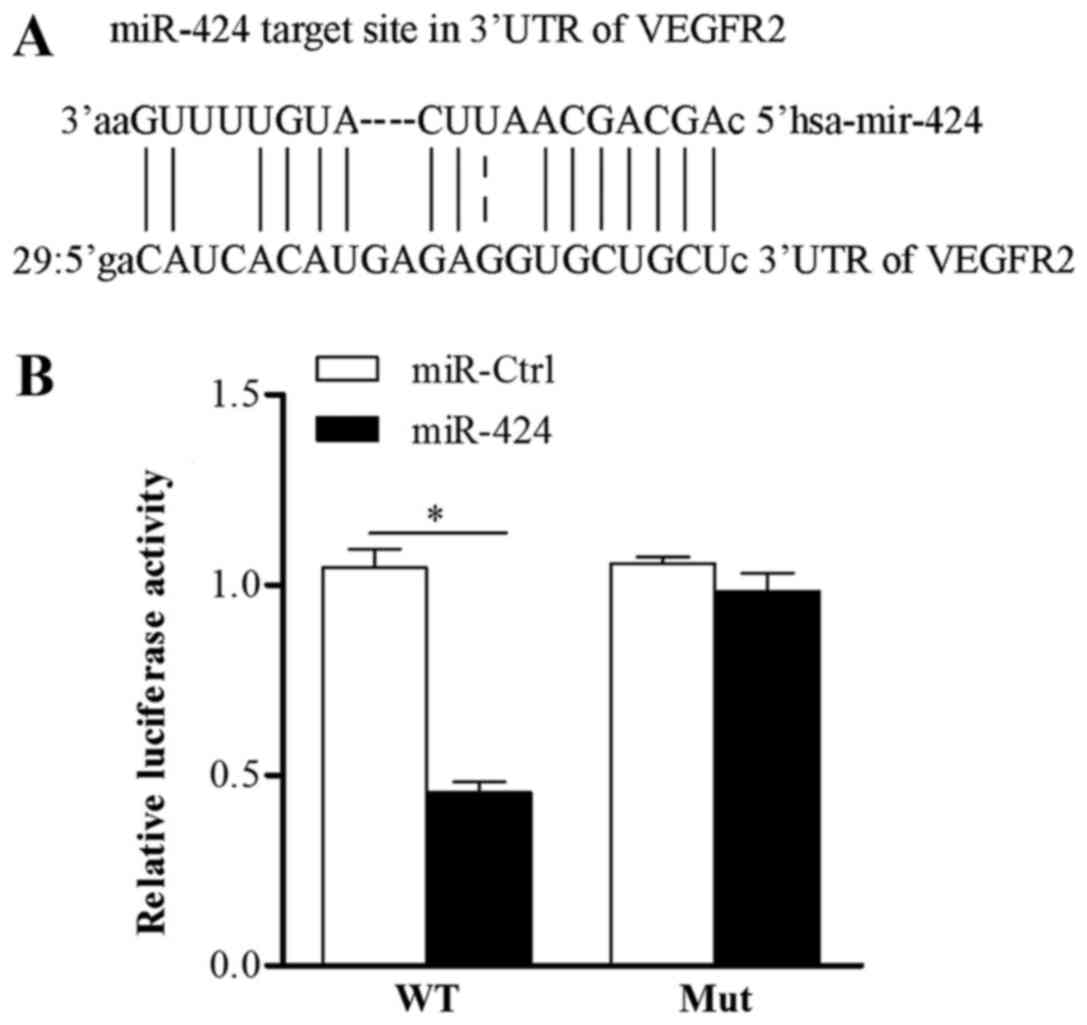 | Figure 3.VEGFR2 is a direct target of miR-424.
(A) VEGFR-2 may be a target of miR-424. (B) 293T cells were
co-transfected with Luc-VEGFR-2-wt or Luc-VEGFR-2-mut, together
with pRL-TK, control miRNA or miR-424 mimics, as indicated.
Following 48 h, firefly luciferase activity was measured and
normalized to Renilla luciferase activity. *P<0.05, as
indicated. VEGFR2, vascular endothelial growth factor receptor 2;
miR, microRNA; Luc, luciferase; WT, wild type; Mut, mutant; Ctrl,
control. |
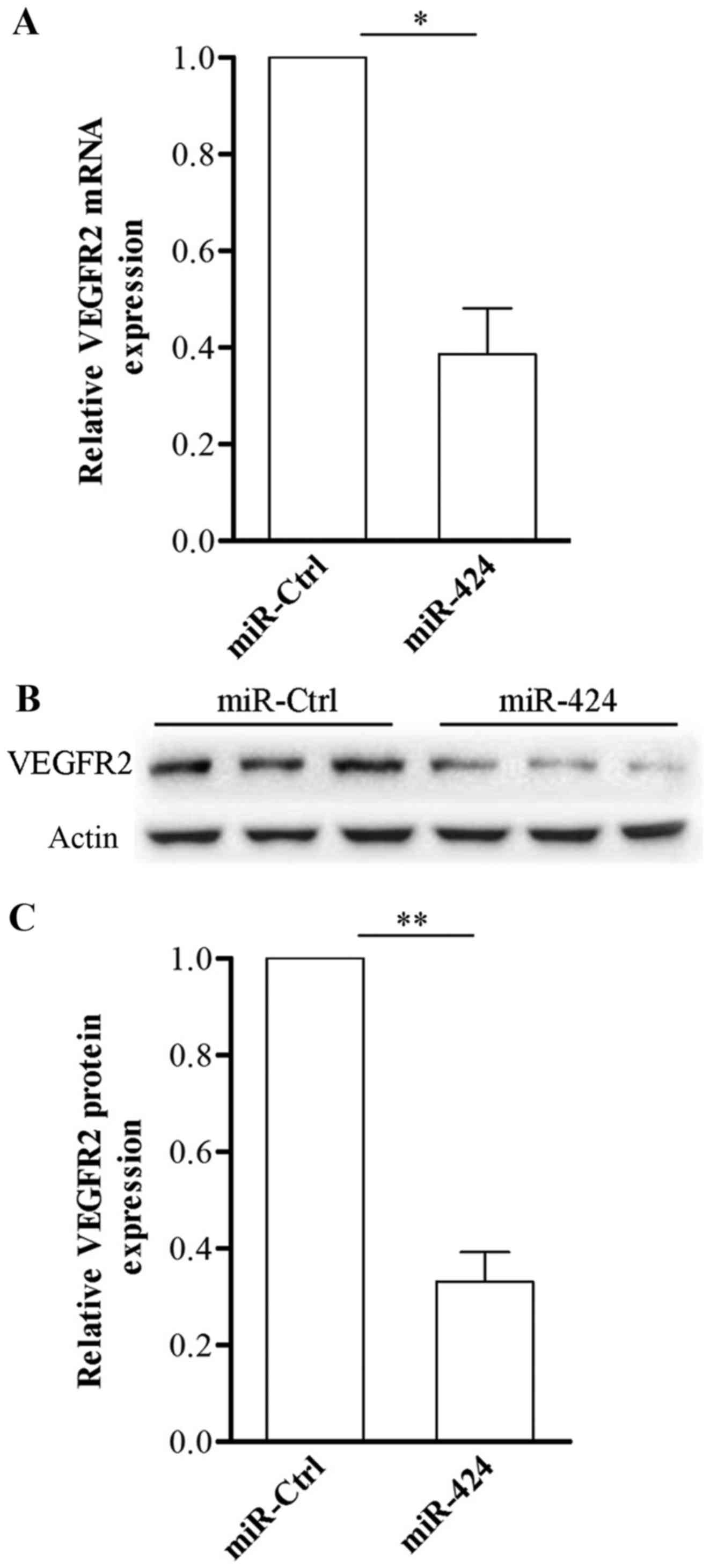
miR-424 downregulates VEGFR2
expression in HemECs
VEGF is an endothelial cell mitogen and survival
factor, involved in regulating vascular development during
embryogenesis and blood-vessel formation in adults (18). VEGFR-2 is the most biologically
important receptor for VEGF. Herein, we found that VEGFR-2 is a
target of miR-424, following the induced overexpression of miR-424
in HemECs. HAs regulates endothelial cell migration, proliferation
and survival. Here we found that VEGFR-2 was a target of miR-424,
then we overexpressed miR-424 in HemECs, As shown in Fig. 4, compared with HemECs transfected
with miR-Ctrl, VEGFR-2 mRNA levels were significantly suppressed in
those HemECs transfected with miR-424 mimics (Fig. 4A); and the overexpression of
miR-424 significantly decreased the expression of VEGFR2 in HemECs
(Fig. 4B). The results
demonstrated that high expression of VEGFR-2 in HemECs might be
attributed to miR-424 downregulation. These results further
verified that VEGFR2 is a bona fide target of miR-424.
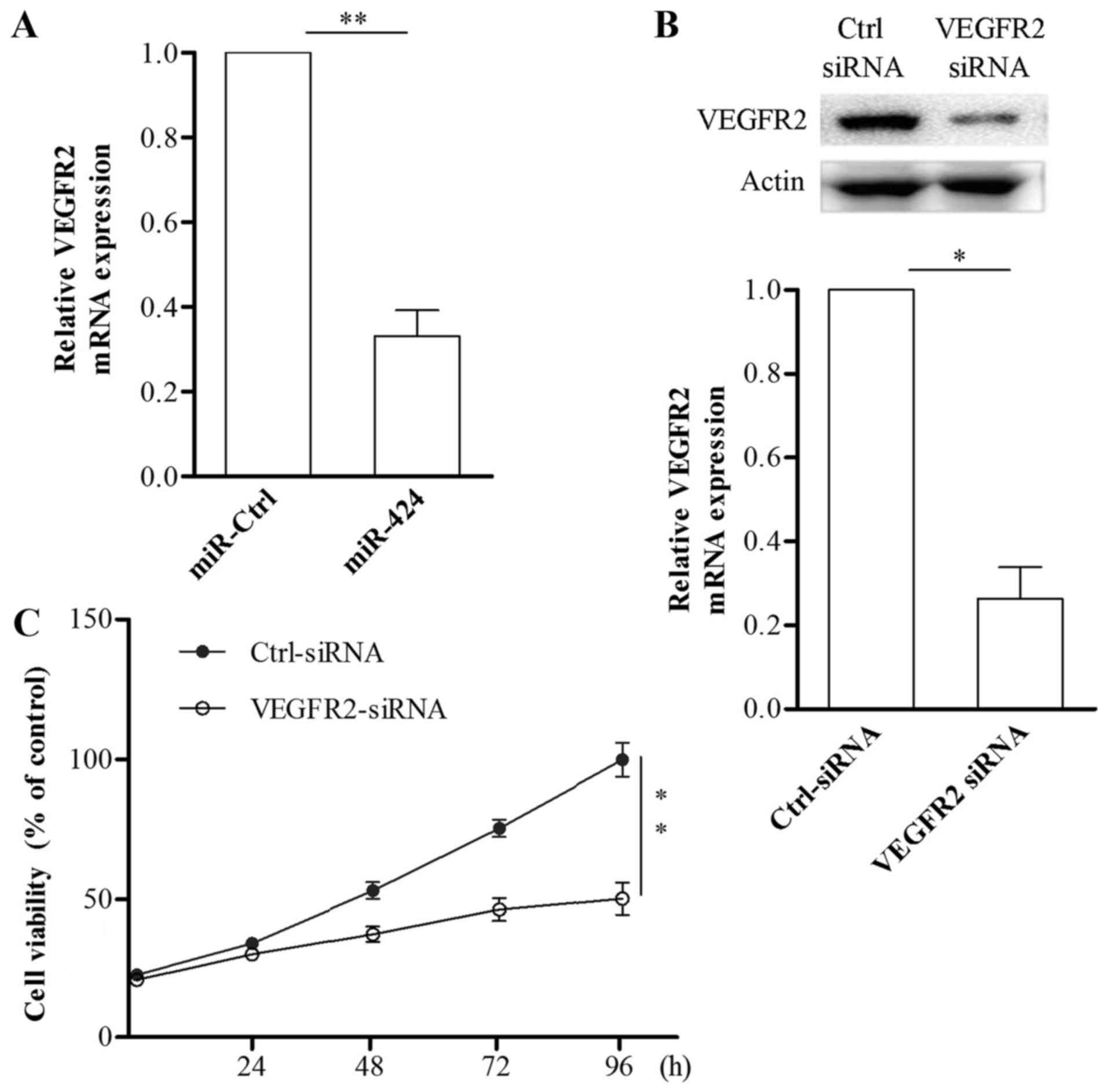
Inhibition of VEGFR2 is responsible
for the suppressive effects of miR-424 in HemECs
To explore whether miR-424 inhibited HemECs growth
is mediated by VEGFR-2, we induced VEGFR-2 knockdown in HemECs
using RNAi. The results showed that, following the transfection of
HemECs with VEGFR2 siRNA, VEGFR2 mRNA and protein expression was
effectively inhibited (Fig. 5A and
B). Results from the CCK-8 assay showed that HemECs growth was
inhibited by the suppression of VEGFR-2 (Fig. 5C). These data demonstrated that
miR-424 suppresses the proliferation of HemECs via targeting
VEGFR-2.
miR-424 inhibits the phosphorylation
of AKT and ERK in HemECs
To determine whether miR-424 suppresses the
downstream signaling of VEGFR-2 in HemECs, we transfected miR-424
mimics or miR-Ctrl into HemECs and then detected the expression of
AKT, p-AKT, ERK1/2 and p-ERK1/2 via western blotting. As shown in
Fig. 6, the upregulation of
miR-424 clearly inhibited phosphorylation of AKT and ERK1/2. These
data reveal that overexpression of miR-424 suppresses HemECs growth
partly via regulation of the AKT and ERK signaling pathways.
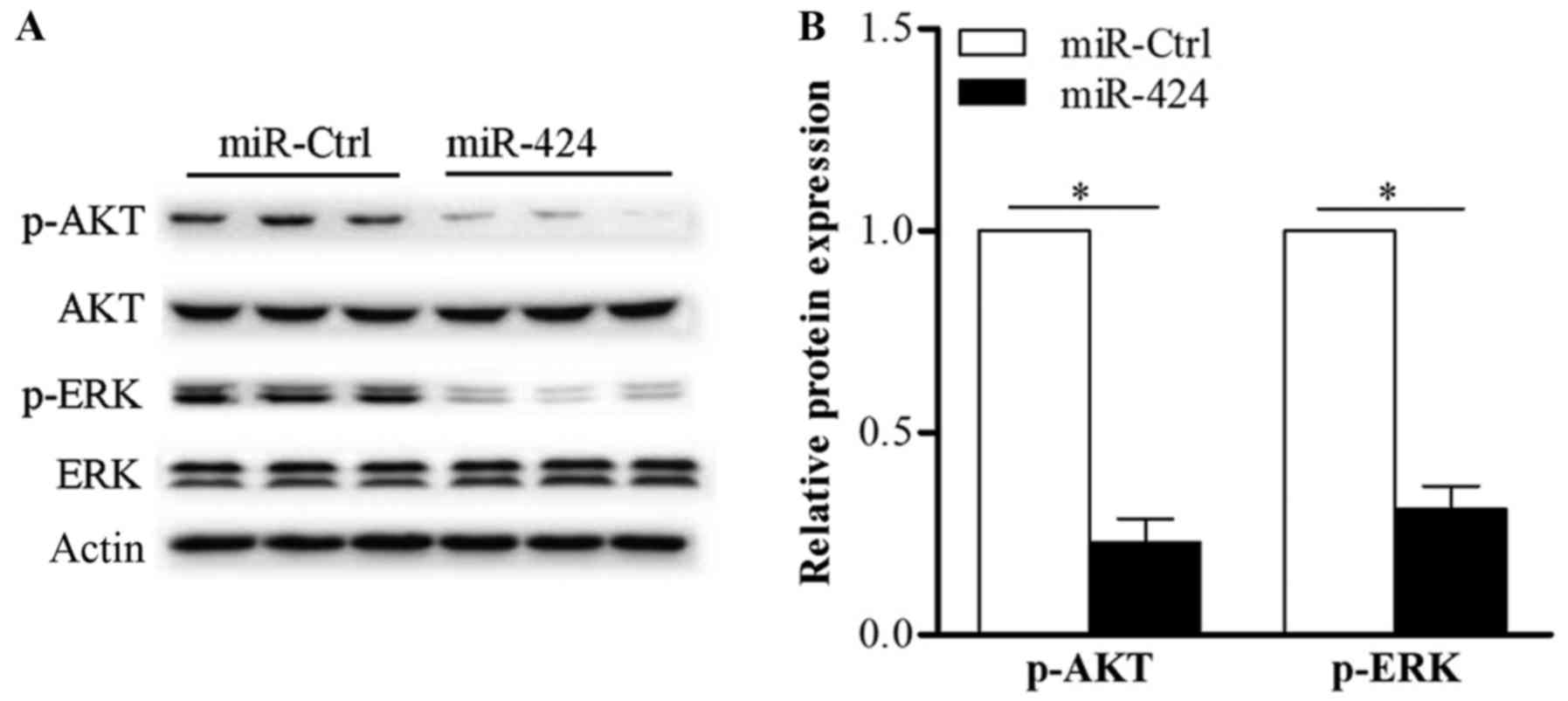 | Figure 6.miR-424 inhibits the AKT and ERK
signaling pathways in HemECs. (A) Following the transfection of
miR-424 mimics or miR-ctrl into HemECs, the expression of AKT,
p-AKT, ERK1/2 and p-ERK1/2 was determined via western blotting. (B)
Upregulation of miR-424 inhibited the phosphorylation of AKT and
ERK in HemECs. Actin was used as an internal control. *P<0.05,
as indicated. miR, microRNA; siRNA, small interfering RNA; HemECs,
hemangioma-derived endothelial cells; Ctrl, control; AKT, protein
kinase B; ERK, extracellular signal-regulated kinase; p-,
phosphorylated. |
Discussion
miRNAs are a group of endogenously expressed small
non-coding RNAs that negatively regulate protein expression by
targeting the 3′ UTR of target mRNAs. It was reported that 30% of
the protein-coding genes in human are regulated by miRNAs (19,20).
There have been increasing number of reports describing how the t
dysregulation of miRNAs contributes to tumorigenesis and tumor
development (21,22). Herein, we evaluated the
downregulation of miR-424 in HemECs, determining that overexpressed
miR-424 in HemECs markedly inhibited cell growth and promoted
apoptosis. Further experiments demonstrated that VEGFR-2 is a
direct target of miR-424, and that miR-424 inhibits HemECs
development through VEGFR-2-mediated ERK signaling pathway. These
findings indicate that miR-424 could represent a therapeutic target
for treatment of HA.
miR-424 has been identified as aberrant in various
tumor types, and many researchers have declared that miR-424 exerts
tumor suppressor role (16,23).
For example, Yu et al (24)
found that miR-424 is downregulated in hepatocellular carcinoma,
and that the overexpression of miR-424 serves a tumor suppressor
role through targeting proto-oncogene c-Myb. In chronic lymphocytic
leukemia, the ectopic expression of miR-424 resulted in
significantly decreased expression of the oncogene PLAG1 (25,26).
It was also reported that miR-424 in cervical cancer cells
suppressed cell growth, migration and invasion (27). Concordant with these discoveries,
miR-424 was found to be dysregulation in senile HA (16). Studies have stated that low miR-424
expression levels contribute to the upregulation of MEK1 or cyclin
E1 in senile HA, which may then cause abnormal tumor cell
proliferation.
Yang et al (28) revealed that miR-424 inhibit cell
proliferation, migration and tube formation capabilities and the
development of infantile skin HA. They further demonstrated that
miR-424 explore the role through suppressing the bFGF/FGFR1/ERK1/2
pathway. Here we also found that the expression of miR-424 was
lowed in HemECs, and that the overexpression of miR-424 in HemECs
clearly inhibited HemECs growth and induced apoptosis. To elucidate
the molecular mechanism underlying the growth suppressive role of
miR-424 in HemECs, we predicted the possible targets of miR-424
using bioinformatics analysis and verified that VEGFR-2 is a direct
target of miR-424 in HemECs. Transfection of miR-424 into HemECs
resulted in significantly decreased VEGFR-2 expression; miR-424
also significantly inhibited VEGFR2 3′ UTR luciferase reporter
activity, while showing no effect on the mut-VEGFR-3′ UTR reporter
activity. These data demonstrate that miR-424 may suppress HA
development partly through reducing VEGFR-2 expression and this is
differs from Yang (28) and her
colleagues' report, which means miR-424 exert inhibition role in HA
via many ways.
Angiogenesis is considered as a critical event in
tumor progression (29,30). Currently, the inhibition of
angiogenesis is a strategy widely proposed for treatment of various
diseases, particularly malignancies (31–34).
Angiogenesis is a physiological process that involves multiple
cellular processes, including endothelial cell proliferation,
migration, and morphological differentiation, and is regulated by
various growth factors and intracellular signaling pathways
(35). VEGFs serve key roles in
regulating angiogenesis. VEGFR-2 is a major receptor in the VEGF
signaling pathway, and regulates cell migration, proliferation, and
angiogenesis (36). It is well
known that HAs are tumors formed by hyper-proliferation of vascular
endothelial cells, which is caused by elevated VEGF signaling
transduction through VEGFR-2. In our previous study, we
demonstrated that increased VEGFR-2 expression is involved in the
development of primary HemECs, though the underlying mechanisms
remain unknown. The results of our investigation revealed that
VEGFR-2 is a direct target of miR-424, and that miR-424 can
negatively regulate the expression of VEGFR-2; these discoveries
may indicate novel mechanism of post-transcriptional control of
VEGFR-2.
In our study, we also examined whether miR-424
affects downstream signaling pathway of VEGFR-2. The results
demonstrated that overexpressed miR-424 in HemECs inhibited
phosphorylation of AKT and ERK1/2. These data demonstrate that
miR-424 inhibits HemECs progression by targeting VEGFR-2 through
the AKT/ERK signaling pathway.
In conclusion, our study suggest that the
downregulation of miR-424 contributes to HA development. Our data
also indicate that miR-424 is clearly downregulated in HemECs, and
that the restoration of miR-424 inhibits HemECs growth and induced
apoptosis partly through suppressing the VEGFR-2 pathway. These
results help us to understand the molecular mechanisms of HA
development and allow us to propose miR-424 as a potential
biomarker and therapeutic target for HA. While, all data above is
based on only one patient, this is a shortage of the study.
Acknowledgements
Not applicable.
Funding
The present study was supported by by the National
Natural Science Foundation of China (grant no. 81572673), the
Science and Technology Commission Foundation of Shanghai, China
(grant no. 13140903802), the Medicine and Engineering Cross
Foundation of Shanghai Jiaotong University (grant no. YG2012MS33)
and Shanghai Science and Technology Department (grant no.
15140901600).
Availability of data and materials
The datasets used or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZF and JO wrote the manuscript. ZQ and YL treated
the patient and collected the clinical samples. ZF, MQ, XQ, YD, SW,
ZQ and YL performed the experiments. ZF, MQ, XQ and JO contributed
to study design, data analysis and interpretation. All authors
reviewed and approved the manuscript.
Ethics approval and consent to
participate
The experimental protocol was approved by the Ethics
Committee of Xinhua Hospital. Written informed consent was obtained
from the parents of the patient.
Patient consent for publication
Written informed consent was obtained for
publication.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Fishman SJ and Mulliken JB: Hemangiomas
and vascular malformations of infancy and childhood. Pediatr Clin
North Am. 40:1177–1200. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jacobs AH and Walton RG: The incidence of
birthmarks in the neonate. Pediatrics. 58:218–222. 1976.PubMed/NCBI
|
|
3
|
Amir J, Metzker A, Krikler R and Reisner
SH: Strawberry hemangioma in preterm infants. Pediatr Dermatol.
3:331–332. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Holland KE and Drolet BA: Infantile
hemangioma. Pediatr Clin North Am. 57:1069–1083. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Grzesik P and Wu JK: Current perspectives
on the optimal management of infantile hemangioma. Pediatric Health
Med Ther. 8:107–116. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Baraniskin A, Birkenkamp-Demtroder K,
Maghnouj A, Zöllner H, Munding J, Klein-Scory S, Reinacher-Schick
A, Schwarte-Waldhoff I, Schmiegel W and Hahn SA: MiR-30a-5p
suppresses tumor growth in colon carcinoma by targeting DTL.
Carcinogenesis. 33:732–739. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Reinhart BJ, Slack FJ, Basson M,
Pasquinelli AE, Bettinger JC, Rougvie AE, Horvitz HR and Ruvkun G:
The 21-nucleotide let-7 RNA regulates developmental timing in
Caenorhabditis elegans. Nature. 403:901–906. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Engels BM and Hutvagner G: Principles and
effects of microRNA-mediated post-transcriptional gene regulation.
Oncogene. 25:6163–6169. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen CZ, Li L, Lodish HF and Bartel DP:
MicroRNAs modulate hematopoietic lineage differentiation. Science.
303:83–86. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Calin GA, Ferracin M, Cimmino A, Di Leva
G, Shimizu M, Wojcik SE, Iorio MV, Visone R, Sever NI, Fabbri M, et
al: A MicroRNA signature associated with prognosis and progression
in chronic lymphocytic leukemia. N Engl J Med. 353:1793–1801. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li D, Li P, Guo Z, Wang H and Pan W:
Downregulation of miR-382 by propranolol inhibits the progression
of infantile hemangioma via the PTEN-mediated AKT/mTOR pathway. Int
J Mol Med. 39:757–763. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bertoni N, Pereira LM, Severino FE, Moura
R, Yoshida WB and Reis PP: Integrative meta-analysis identifies
microRNA-regulated networks in infantile hemangioma. BMC Med Genet.
17:42016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Venneti S, Boateng LA, Friedman JR,
Baldwin DA, Tobias JW, Judkins AR, Mourelatos Z and Lal P: MiRNA-9
and MiRNA-200a distinguish hemangioblastomas from metastatic clear
cell renal cell carcinomas in the CNS. Brain Pathol. 22:522–529.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang C, Huang J, Ma P and Yu G:
microRNA-143 acts as a suppressor of hemangioma growth by targeting
Bcl-2. Gene. 628:211–217. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chamorro-Jorganes A, Araldi E, Penalva LO,
Sandhu D, Fernández-Hernando C and Suárez Y: MicroRNA-16 and
microRNA-424 regulate cell-autonomous angiogenic functions in
endothelial cells via targeting vascular endothelial growth factor
receptor-2 and fibroblast growth factor receptor-1. Arterioscler
Thromb Vasc Biol. 31:2595–2606. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nakashima T, Jinnin M, Etoh T, Fukushima
S, Masuguchi S, Maruo K, Inoue Y, Ishihara T and Ihn H:
Down-regulation of mir-424 contributes to the abnormal angiogenesis
via MEK1 and cyclin E1 in senile hemangioma: Its implications to
therapy. PLoS One. 5:e143342010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Olsson AK, Dimberg A, Kreuger J and
Claesson-Welsh L: VEGF receptor signalling-in control of vascular
function. Nat Rev Mol Cell Biol. 7:359–371. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hwang HW and Mendell JT: MicroRNAs in cell
proliferation, cell death, and tumorigenesis. Br J Cancer.
94:776–780. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cho WC: MicroRNAs: Potential biomarkers
for cancer diagnosis, prognosis and targets for therapy. Int J
Biochem Cell Biol. 42:1273–1281. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cho WC: MicroRNAs in cancer-from research
to therapy. Biochim Biophys Acta. 1805:209–217. 2010.PubMed/NCBI
|
|
23
|
Long XH, Mao JH, Peng AF, Zhou Y, Huang SH
and Liu ZL: Tumor suppressive microRNA-424 inhibits osteosarcoma
cell migration and invasion via targeting fatty acid synthase. Exp
Ther Med. 5:1048–1052. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yu L, Ding GF, He C, Sun L, Jiang Y and
Zhu L: MicroRNA-424 is down-regulated in hepatocellular carcinoma
and suppresses cell migration and invasion through c-Myb. PLoS One.
9:e916612014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pallasch CP, Patz M, Park YJ, Hagist S,
Eggle D, Claus R, Debey-Pascher S, Schulz A, Frenzel LP, Claasen J,
et al: miRNA deregulation by epigenetic silencing disrupts
suppression of the oncogene PLAG1 in chronic lymphocytic leukemia.
Blood. 114:3255–3264. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Faraoni I, Laterza S, Ardiri D, Ciardi C,
Fazi F and Lo-Coco F: MiR-424 and miR-155 deregulated expression in
cytogenetically normal acute myeloid leukaemia: Correlation with
NPM1 and FLT3 mutation status. J Hematol Oncol. 5:262012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu J, Li Y, Wang F, Wang X, Cheng B, Ye F,
Xie X, Zhou C and Lu W: Suppressed miR-424 expression via
upregulation of target gene Chk1 contributes to the progression of
cervical cancer. Oncogene. 32:976–987. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang L, Dai J, Li F, Cheng H, Yan D and
Ruan Q: The expression and function of miR-424 in infantile skin
hemangioma and its mechanism. Sci Rep. 7:118462017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Folkman J: Role of angiogenesis in tumor
growth and metastasis. Semin Oncol. 29 6 Suppl 16:S15–S18. 2002.
View Article : Google Scholar
|
|
30
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Folkman J: Anti-angiogenesis: New concept
for therapy of solid tumors. Ann Surg. 175:409–416. 1972.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Samant RS and Shevde LA: Recent advances
in anti-angiogenic therapy of cancer. Oncotarget. 2:122–134. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bose D, Meric-Bernstam F, Hofstetter W,
Reardon DA, Flaherty KT and Ellis LM: Vascular endothelial growth
factor targeted therapy in the perioperative setting: Implications
for patient care. Lancet Oncol. 11:373–382. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ellis LM and Hicklin DJ: VEGF-targeted
therapy: Mechanisms of anti-tumour activity. Nat Rev Cancer.
8:579–591. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jiang BH, Zheng JZ, Aoki M and Vogt PK:
Phosphatidylinositol 3-kinase signaling mediates angiogenesis and
expression of vascular endothelial growth factor in endothelial
cells. Proc Natl Acad Sci USA. 97:pp. 1749–1753. 2000; View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Herbst RS: Review of epidermal growth
factor receptor biology. Int J Radiat Oncol Biol Phys. 59 2
Suppl:S21–S26. 2004. View Article : Google Scholar
|