Introduction
Approximately 20–25% of filtered Na and Cl load is
reabsorbed by the thick ascending limb (TAL) of the kidney, and
thus it serves a key role in regulating urine concentration
(1,2). Transepithelial Na and Cl transport
occurs via a two-step process: Na and Cl enter the cells through
type II Na-K-Cl cotransporters (NKCC2) in the apical membrane and
then leave the cells via the basolateral membrane through Na pumps
and Cl channels (3). During this
process, the activity of NKCC2 can be regulated by apical and
basolateral K channels. Indirectly affected by basolateral K
channels through the generation of the cell membrane potential and
driving Cl diffusion, NKCC2 activity is maintained by apical K
channels through K recycling (4,5). Gu
et al (6) demonstrated that
several types of K channels are present in the basolateral membrane
of TAL, with the 50 pS K channel being the primary type, which is
regulated by many factors (7).
However, the regulatory mechanism remains unclear.
Tumor necrosis factor (TNF) is produced by various
epithelial cells, including those of the TAL, and is one of the
main inflammatory cytokines with a great variety of actions in
regulating inflammatory, immune and stress responses, as well as
host defense, and cellular apoptosis and necrosis (8–10).
Wei et al (11)
demonstrated that TNF stimulates the apical 70 pS K channels and
regulates NKCC2 activity by activating protein tyrosine phosphatase
(PTP) in the TAL. As aforementioned, apical and basolateral K
channels can regulate NKCC2 activity, thus, it may also be possible
that TNF modulates NKCC2 activity by affecting basolateral K
channel activity. Therefore, the present study aimed to determine
the acute effect of TNF on basolateral 50 pS K channels and explore
the regulatory mechanism of TNF.
Materials and methods
Reagents
TNF, H89, herbimycin A, arachidonyl trifluoromethyl
ketone (AACOCF3), phenylarsine oxide (PAO), polylysine and
collagenase were purchased from Sigma-Aldrich; Merck KGaA
(Darmstadt, Germany). Antibodies against phospho (p)-PTP, PTP and
actin were obtained from Affinity Biotechnology (Exeter, UK).
TNF treatment
Following patching and recording the 50 pS potassium
channel current for 2–3 min, TNF (10 nM) was added to the bath and
the 50 pS potassium channel current recorded for 3–5 min.
Inhibitor treatment
After recording 50 pS potassium channel current for
2~3 min, the inhibitor (5 µM H89/5 µM herbimycin A/5 µM AACOCF3/1
µM PAO) was added to the bath. Following recording for 3–5 min,
then the TNF was added to observe the change of 50 pS potassium
channel current.
Preparation of the TALs
Male and female pathogen-free Sprague-Dawley rats
(n=80; male:female 1:1; 4–5 weeks old: weight, 50–60 g; Animal
Facility of The Second Affiliated Hospital of Harbin Medical
University, Harbin, China) were housed at 20–25°C, 50–65% relative
humidity and a 12-h light/dark cycle with free access to normal rat
chow and tap water. Rats were sacrificed by cervical dislocation
and the kidneys were removed immediately. The kidney was cut into
1-mm thick slices with a blade, and then incubated at 37°C in a
HEPES buffer solution containing 10 mM HEPES, 140 mM NaCl, 5 mM
KCl, 1.5 mM MgCl2 and 1.8 mM CaCl2 (pH 7.4)
with collagenase type 1A (1 mg/ml) for 40–60 min. The
collagenase-treated TALs were isolated under a dissecting
microscope and placed on a 5×5 mm cover glass coated with
polylysine. The cover glass with TALs was transferred to a chamber
filled with HEPES buffer solution and mounted on an inverted
microscope (Leica Microsystems GmbH, Wetzlar, Germany). The present
study was approved by the Medical Ethics Committee of Jiamusi
University (Heilongjiang, China).
Patch-clamp technique
The patch-clamp electrodes were pulled with a P-97
electrode-puller, and filled with a pipette solution containing 10
mM HEPES, 140 mM KCl and 1.8 mM MgCl2 (pH 7.4). The
channel currents recorded by an Axon 700B patch-clamp amplifier,
were low-pass filtered at 0.5 kHz and digitized with an Axon
interface (Digidata 1400A). Data were analyzed using the pClamp
10.0 software system (Axon Instruments; Molecular Devices, LLC,
Sunnyvale, CA, USA). The channel activity was expressed as a
product of channel open probability (Po) and
channel number (N), and calculated as follows from data
samples of 90-sec duration in the steady state:
NPo =∑
(1t1+2t2+…iti),
where ti is the fractional open time spent at
each of the observed current levels.
Western blot analysis
Total protein was extracted from the TAL tissues
using RIPA lysis buffer (Beyotime Institute of Biotechnology,
Haimen, China). Once protein concentrations were determined using
the BCA method, 40 µl of protein extract was mixed with 10 µl of 5X
SDS-PAGE loading buffer and boiled for 5 min. Following separation
by electrophoresis at 50 µg protein/lane on a 10% SDS-PAGE gel, 50
µg of medullary TAL protein was transferred onto a nitrocellulose
membrane. The membranes were then placed in blocking solution
containing 5% nonfat dry milk in Tris-buffered saline-0.05% Tween
(TBS-T) and blocked for 1 h at room temperature. The membranes were
incubated with the rabbit anti-rat primary antibody (1:1,000;
AF3412; Affinity Biosciences, Cincinnati, OH, USA) and β-actin
(1:5,000; AF7018; Affinity Biosciences) for 12 h at 4°C.
Subsequently, the membranes were washed four times with TBS-T and
incubated with goat anti-rabbit secondary antibody (1:5,000;
ZB-2301; OriGene Technologies, Inc., Beijing, China) for 1 h at
room temperature. Finally, the membranes were washed again with
TBS-T four times and the protein bands were detected with enhanced
chemiluminescence (Pierce; Thermo Fisher Scientific, Inc., Waltham,
MA, USA). The protein bands were scanned and quantified to analyze
the protein expression using ImageJ software (version 1.45s;
National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
All data are presented as the mean ± standard error
of the mean. The Student's t-test was used to determine the
differences between two groups and analysis of variance followed by
Student-Newman-Keuls post hoc test were used to determine the
differences among multiple groups. SPSS software version 19.0 (IBM
Corp., Armonk, NY, USA) was used and P<0.05 was considered to
indicate a statistically significant difference.
Results
Effect of TNF on 50 pS K channel
activity
Previous studies have demonstrated that many factors
regulate basolateral 50 pS K channels; for example, adenosine
stimulating and calcium inhibiting the K channel (6,12,13).
The aim of the present study was to investigate the regulation of
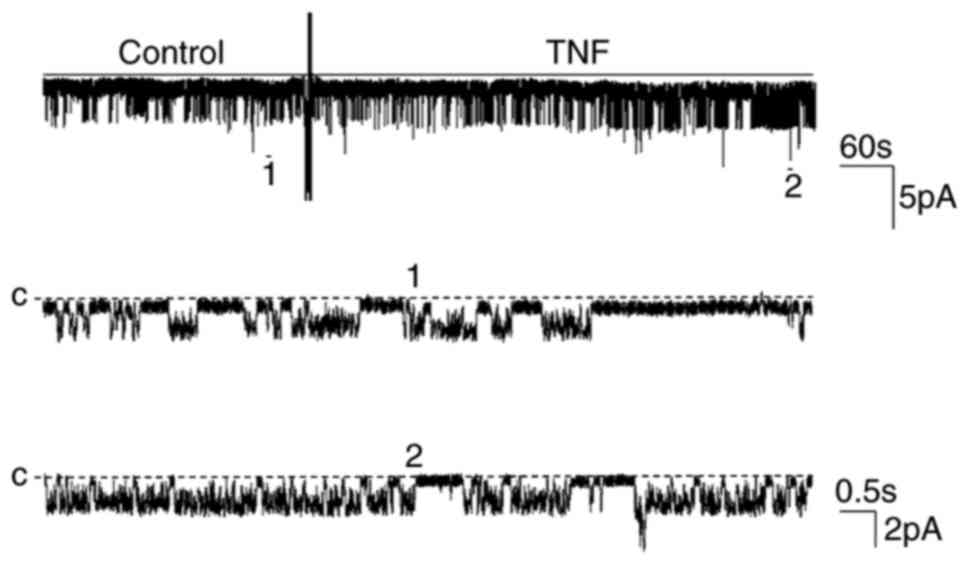
TNF on 50 pS K channels. The addition of 10 nM TNF to the bath
increased the channel activity (NPo) from
0.28±0.07 to 0.55±0.12 in a cell-attached patch (n=5; P<0.01;
Fig. 1). The stimulatory effect of
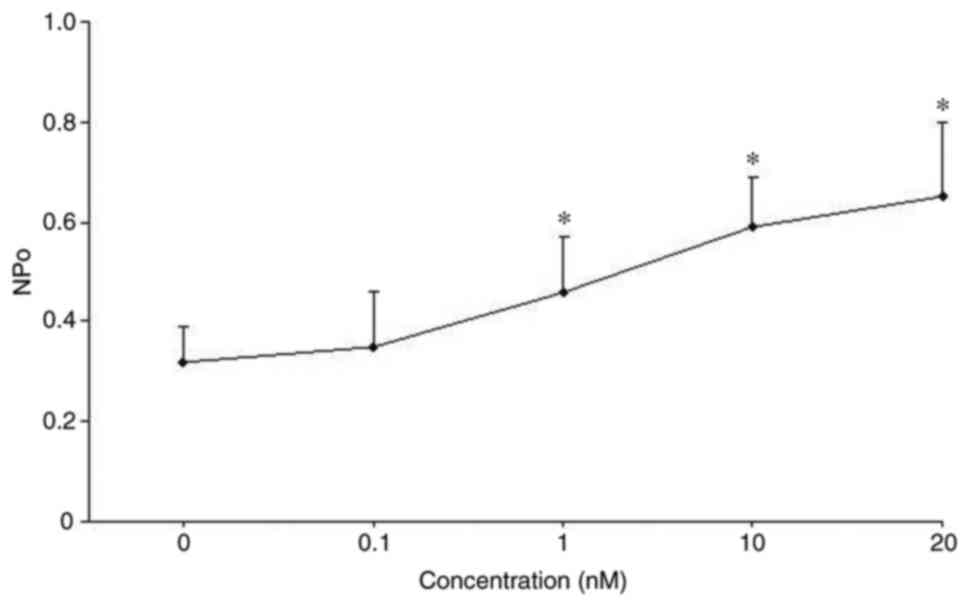
TNF on 50 pS K channel activity was concentration-dependent. The
dose-response curve presented in Fig.
2 demonstrated that 50 pS K channel activity increased with
rising concentrations of TNF from 1 to 20 nM (n=5; P<0.05).
Role of protein kinase A (PKA) in the
stimulatory effect of TNF
Once the stimulatory effect of TNF on the
basolateral 50 pS K channels had been demonstrated, the present
study investigated the mechanism underlying the stimulatory effect
of TNF. Previous studies have reported that the activity of 50 pS K
channels is increased by stimulating PKA or inhibiting
phospholipase A2 (PLA2) (6,12).
Therefore, the role of PKA in the stimulatory effect of TNF on 50
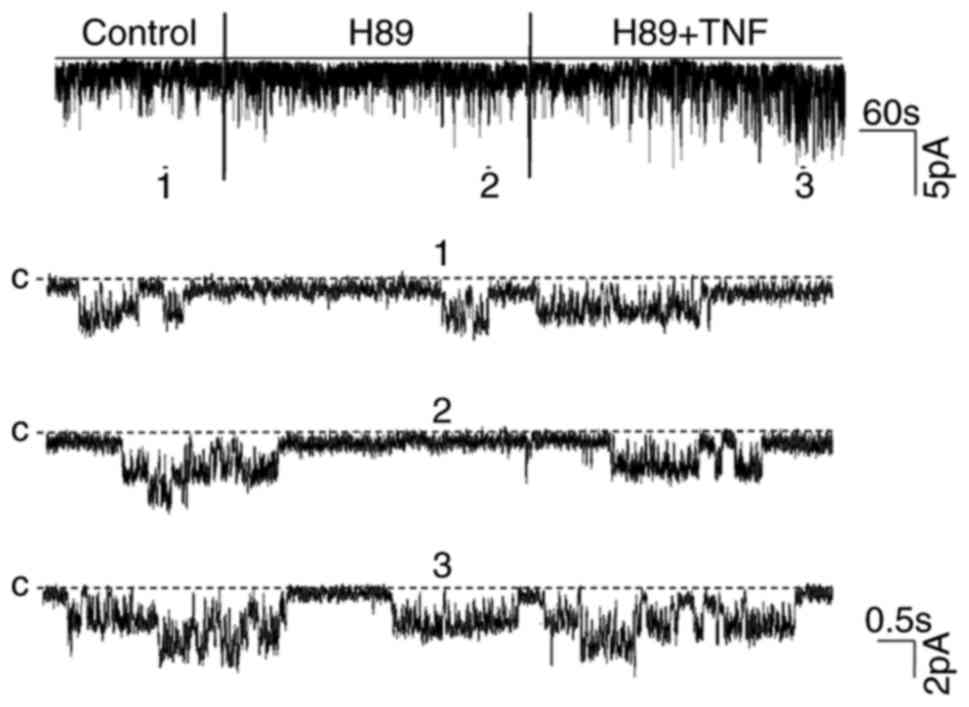
pS K channels was determined. Notably, no significant differences
were observed in regard to the channel activity nor the stimulatory
effect of TNF on the channel following the inhibition of PKA with
H89, an inhibitor of PKA (Fig. 3).
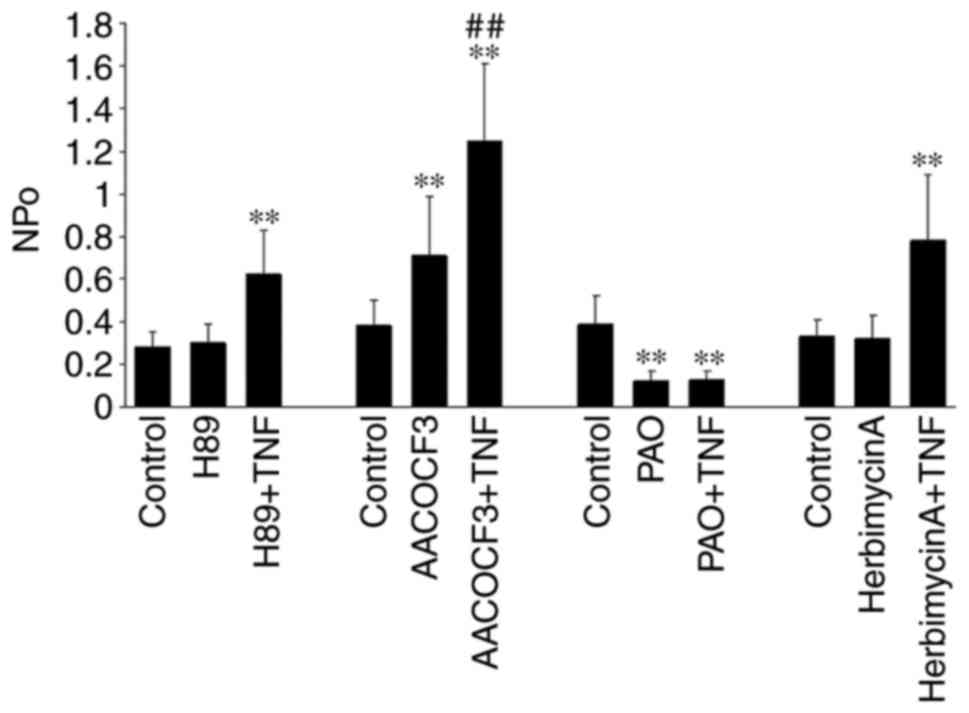
The results summarized in Fig. 4
revealed that H89 did not alter the channel activity
(NPo, 0.29±0.08 to 0.3±0.09), but did
significantly increase channel activity following the application
of TNF (0.62±0.13; n=5; P<0.01). This suggested that the
PKA-dependent pathway did not mediate the stimulatory effect of TNF
on the 50 pS K channel.
Role of PLA2 in the
stimulatory effect of TNF
Next, the present study determined the role of
AACOCF3 (5 µM), an inhibitor of PLA2, in the stimulatory
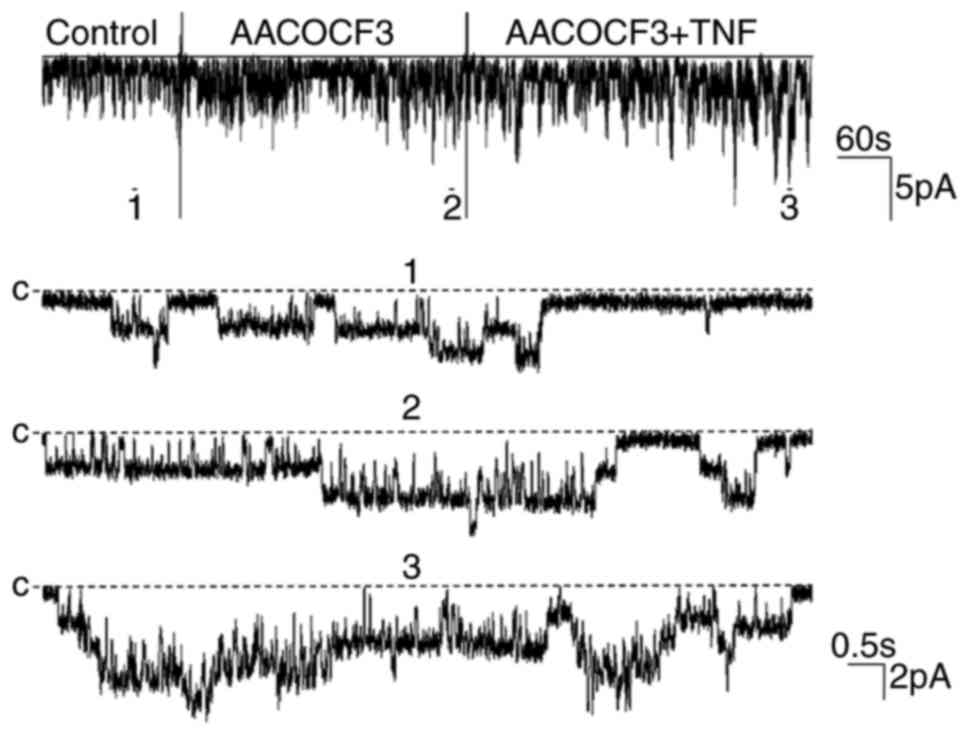
effect of TNF on the 50 pS K channels (Fig. 5). The results of the present study
were in agreement with those of previous reports (12,13),
as AACOCF3 significantly stimulated 50 pS K channel activity and
increased the NPo from 0.38±0.09 to 0.71±0.21
(n=5; P<0.01). As shown in Fig.
4, the addition of TNF increased the NPo from
0.71±0.21 to 1.25±0.26 (n=5; P<0.01) following the inhibition of
PLA2 with AACOCF3, indicating that the effect of TNF on
the K channel was not mediated by the PLA2-dependent
pathway.
Role of PTP in the stimulatory effect
of TNF
It has been reported previously that PTP can be
increased by stimulating TNF receptors (14). Thus, it is necessary to explore the
role of PTP in mediating the stimulatory effect of TNF on 50 pS K
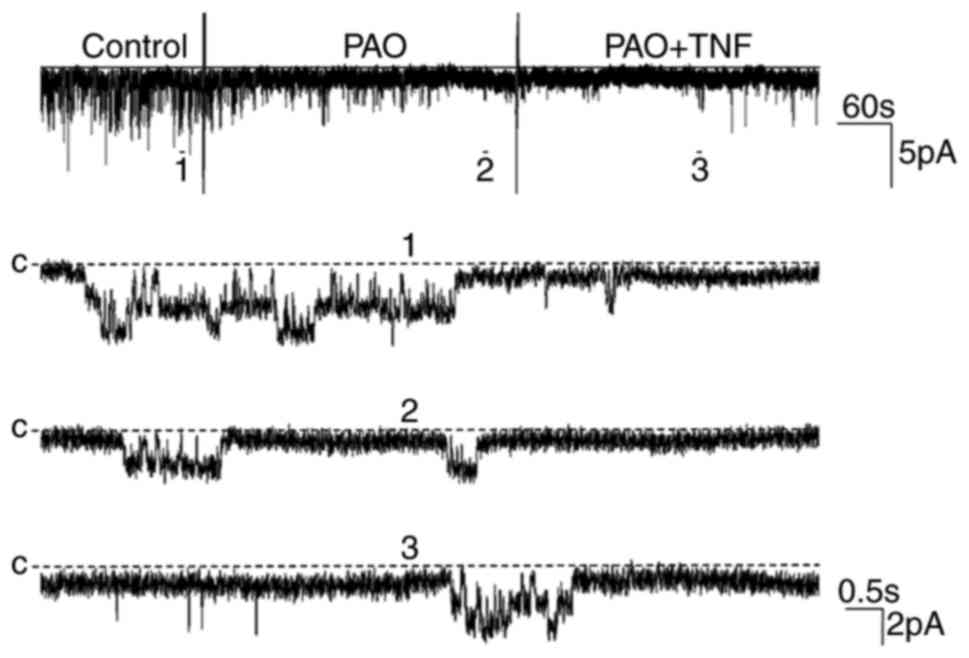
channels. The effect of TNF on channel activity was examined in the
presence of PAO (1 µM), an inhibitor of PTP. The results indicated
that PAO markedly decreased channel activity and eliminated the
stimulatory effect of TNF (Fig.
6). The statistical results in Fig. 4 revealed that the
NPo of the 50 pS K channel was decreased
following PAO treatment from 0.39±0.13 to 0.12±0.05 (n=5;
P<0.01) and the application of TNF did not increase the
NPo of the channel (0.13±0.04; n=5; P>0.05),
suggesting that the stimulatory effect of TNF may be derived from
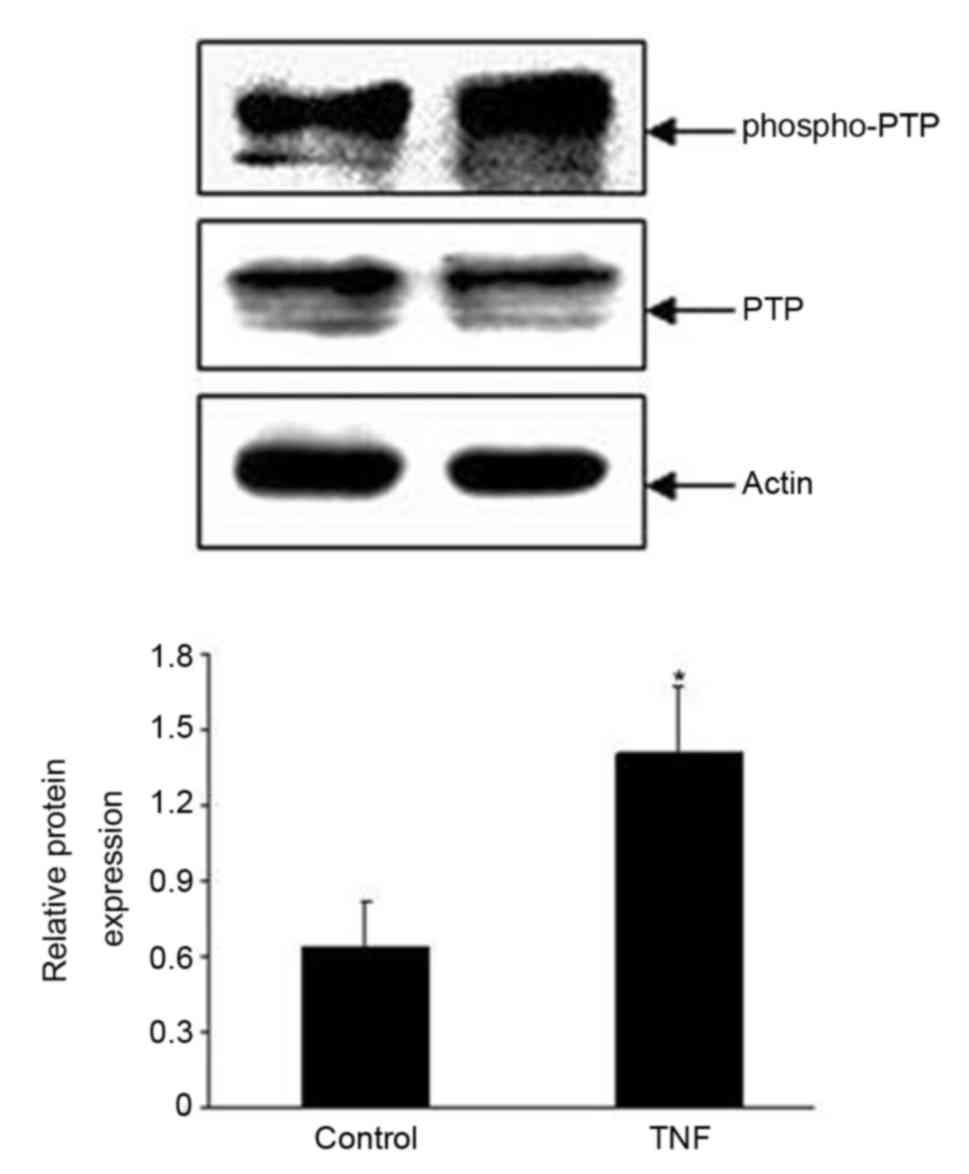
PTP stimulation. In order to further confirm the effect of PTP, a
primary antibody directed against phosphorylated PTP at tyrosine
residue 580 (p-PTP) was used to examine whether the application of
TNF (10 nM) for 5 min in the TAL affected the phosphorylation level
of PTP. Fig. 7 presents a
representative western blot image revealing that TNF treatment
significantly increased the phosphorylation level of PTP at
tyrosine residue 580 by 55±10% (n=3; P<0.01). These results
indicated that PTP activity stimulation may have increased the
TNF-induced stimulatory effect on 50 pS K channels.
Role of protein tyrosine kinase (PTK)
in the stimulatory effect of TNF
The aforementioned results suggested that the
stimulatory effect of TNF on the 50 pS K channels may have resulted
from stimulation of PTP and then enhancement of tyrosine
dephosphorylation. However, PTP tyrosine phosphorylation is
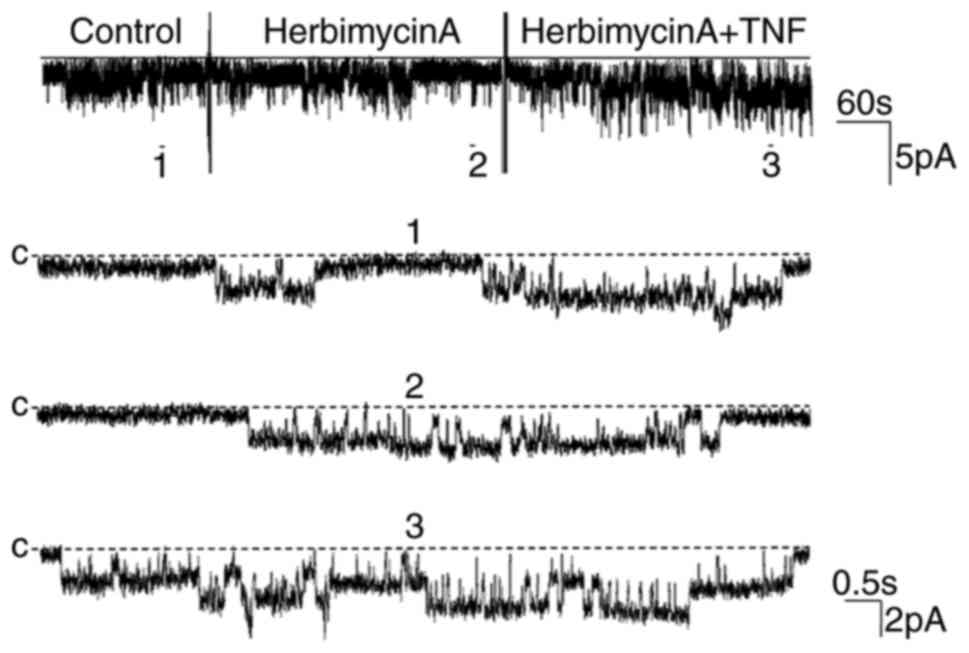
determined by PTK as well as PTP (15). Therefore, the present study further
investigated whether the PTK pathway mediated the stimulatory
effect of TNF on the 50 pS K channel. Firstly, the changes in
channel activity following treatment with Herbimycin A (5 µM), an
inhibitor of PTK, were observed (Fig.
8). Then, the role of PTK in the stimulatory effect of TNF on
the channel was also determined. The statistical results revealed
that Herbimycin A did not significantly alter the channel activity
nor the stimulatory effect of TNF. The NPo value
was 0.33±0.08 prior to the addition of Herbimycin A, and 0.32±0.10
following treatment (n=5; P>0.05; Fig. 4). Furthermore, the addition of TNF
significantly increased the channel activity
(NPo, 0.78±0.20; n=5; P<0.01; Fig. 4). These results indicated that the
stimulatory effect of TNF was not mediated by the PTK-dependent
pathway. The histogram of statistical results is shown in Fig. 4.
Discussion
In the present study, an important observation was
demonstrated: The acute application of TNF stimulated the
basolateral 50 pS K channel in the TAL through a PTP-dependent
pathway. This was supported by two lines of evidence: i) The
stimulatory effect of TNF on the channel was absent in the presence
of the PTP inhibitor; and ii) the phosphorylation level of PTP at
Tyr580 in the TAL was enhanced following TNF treatment. Therefore,
it was hypothesized that TNF may activate PTP and enhance tyrosine
dephosphorylation, thereby increasing basolateral 50 pS K channel
activity in the TAL.
Usually, TNF production is low or absent prior to
cellular stimulation; however, it increases significantly when
exposed to inflammation, infection and injury. Thus, it is possible
that TNF may be associated with the occurrence of inflammation,
cell differentiation and cell death (16). It has been reported that TNF
regulates several pathophysiological events associated with renal
inflammatory diseases (17,18),
for instance, it contributes to lipopolysaccharide-induced
glomerular endothelial injury (19). In addition, recent studies have
suggested that TNF can be produced by physiologically relevant
stimulations. It has been demonstrated that stimulation of the
Ca2+-sensing receptor (CaR) and hypertonic NaCl intake
may increase TNF generation in TAL cells, and thereby regulate NaCl
transport (20–23).
Wang et al (20) demonstrated that stimulation of the
CaR increases TNF production via TAL cell incubation with high
Ca2+ for >3 h. Furthermore, TNF increases
cyclooxygenase (COX)-2 expression and prostaglandin E2
(PGE2) production in TAL cells following CaR
stimulation. It has been reported previously that inhibition of
COX-2 in the TAL blocks TNF-mediated inhibition of Rubidium-86
(86Rb) uptake, an in vitro indicator of
natriuresis (20). In addition,
PGE2 has been demonstrated to inhibit the apical K
channel (24) and basolateral K
channel in the TAL (25),
decreasing NaCl reabsorption in that segment. Therefore, TNF
production induced by CaR stimulation may be a renal mechanism for
regulating water and salt excretion.
Hao et al (23) demonstrated that hypertonic NaCl
intake for 3 days increases renal TNF production via a pathway
involving NKCC2 in the TAL. Furthermore, it has been revealed that
the protein expression and activity of NKCC2 increases in
TNF-deficient mice (26),
indicating that TNF inhibits NKCC2 as part of a negative feedback
regulator. Thus, TNF may inhibit NaCl reabsorption in the TAL as an
endogenous inhibitor of NKCC2, subsequently leading to polyuria and
natriuresis (27,28).
The present study demonstrated that acute
application of TNF increased the activity of the basolateral 50 pS
K channel. As the basolateral K channels determine the driving
force for Cl diffusion across the basolateral membrane, a
TNF-induced increase in channel activity should be associated with
an increase in NaCl transport in the TAL. This conclusion is
inconsistent with the inhibitory effect of TNF on NaCl transport in
the TAL. Such a discrepancy may partly be accounted for by the
temporal factor. TNF reduced 86Rb uptake and NKCC
expression following h or days, whereas stimulation of channel
activity by TNF occurred in a few min. The effects of TNF on
channel activity may be time-dependent and biphasic: The acute
effect of TNF is to stimulate, whereas the chronic effect of TNF is
to inhibit NaCl transport in the TAL. This phenomenon is similar to
the delayed suppressive and acute stimulatory effect of
interferon-γ on 40 pS K+ channel activity in cultured
human proximal tubule cells (29,30).
In addition, the results of the present study are consistent with
the report by Wei et al (11), which demonstrated that TNF
stimulated apical 70 pS K+ channel activity,
consequently increasing NaCl transport in the TAL.
The significance of TNF in modulating renal tubular
K+ channels in the TAL is not well understood. However,
many reports have demonstrated that changes in the activity of
K+ channels are involved in cell injury. For instance,
Nietsch et al (31)
reported that exposure to TNF for 3–5 min increased the opening
times of K+ channels and blockers of K+
channels attenuated the progression to cell death by TNF at 4 h
following TNF exposure in a rat liver cell line. Their observation
indicates that channel opening is an early event in the
TNF-mediated pathway leading to liver cell death (31). In addition, many studies have
revealed that the occurrence of certain diseases, including
diabetic nephropathy, is associated with an increase in TNF
expression (32,33). Therefore, further studies should
focus on exploring whether activation of the basolateral K channels
induced by TNF is an early event of cell injury in the TAL.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Chinese
National Nature Science Foundation (grant no. 31400994), National
Undergraduate Innovation Project (grant no. 201610222030) and North
Medicine and Functional Food Characteristic Construction
Project.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
ZY, GL and ZX performed the experiments. MX and ZT
acquired and analyzed data. MM and LY analyzed data and drafted the
manuscript. SY and WQ designed the project and revised the
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Medical Ethics
Committee of Jiamusi University (Heilongjiang, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Greger R: Ion transport mechanisms in
thick ascending limb of Henle's loop of mammalian nephron. Physiol
Rev. 65:760–797. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hebert SC: Roles of Na-K-2Cl and Na-Cl
cotransporters and ROMK potassium channels in urinary concentrating
mechanism. Am J Physiol. 275:F325–F327. 1998.PubMed/NCBI
|
|
3
|
Vitzthum H, Castrop H, Meier-Meitinger M,
Riegger GA, Kurtz A, Krämer BK and Wolf K: Nephron specific
regulation of chloride channel CLC-K2 mRNA in the rat. Kidney Int.
61:547–554. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Simon DB, Karet FE, Rodriguez-Soriano J,
Hamdan JH, DiPietro A, Trachtman H, Sanjad SA and Lifton RP:
Genetic heterogeneity of Bartter's syndrome revealed by mutations
in the K+ channel, ROMK. Nat Genet. 14:152–156. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hebert SC, Desir G, Giebisch G and Wang W:
Molecular diversity and regulation of renal potassium channels.
Physiol Rev. 85:319–371. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gu R, Wang J, Zhang Y, Li W, Xu Y, Shan H,
Wang WH and Yang B: Adenosine stimulates the basolateral 50 pS K
channels in the thick ascending limb of the rat kidney. Am J
Physiol Renal Physiol. 293:F299–F305. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Paulais M, Lourdel S and Teulon J:
Properties of an inwardly rectifying K(+) channel in the
basolateral membrane of mouse TAL. Am J Physiol Renal Physiol.
282:F866–F876. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Foster D, Parrish-Novak J, Fox B and Xu W:
Cytokinereceptor pairing: Accelerating discovery of cytokine
function. Nat Rev Drug Discov. 3:160–170. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hehlgans T and Pfeffer K: The intriguing
biology of the tumour necrosis factor/tumour necrosis factor
receptor superfamily: Players, rules and the games. Immunology.
115:1–20. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tansey MG and Szymkowski DE: The TNF
superfamily in 2009: New pathways, new indications, and new drugs.
Drug Discovery Today. 14:1082–1088. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wei Y, Babilonia E, Pedraza PL, Ferreri NR
and Wang WH: Acute application of TNF stimulates apical 70-pS K+
channels in the thick ascending limb of rat kidney. Am J Physiol
Renal Physiol. 285:F491–F497. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang M, Sui H, Li W, Wang J, Liu Y, Gu L,
Wang WH and Gu R: Stimulation of A(2a) adenosine receptor abolishes
the inhibitory effect of arachidonic acid on the basolateral 50-pS
K channel in the thick ascending limb. Am J Physiol Renal Physiol.
300:F906–F913. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kong S, Zhang C, Li W, Wang L, Luan H,
Wang W and Gu R: Stimulation of Ca2+-sensing receptor inhibits the
basolateral 50-pS K channels in the thick ascending limb of rat
kidney. Biochim Biophys Acta. 1823:273–281. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sasaki CY and Patek PQ: Transformation is
associated with an increase in sensitivity to TNF-mediated lysis as
a result of an increase in TNF-induced protein tyrosine phosphatase
activity. Int J Cancer. 81:141–147. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gu RM, Wei Y, Falck JR, Krishna UM and
Wang WH: Effects of protein tyrosine kinase and protein tyrosine
phosphatase on apical K(+) channels in the TAL. Am J Physiol Cell
Physiol. 281:C1188–C1195. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
MacEwan DJ: TNF receptor subtype
signaling: Differences and cellular consequences. Cell Signal.
14:477–492. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Klahr S and Morrissey JJ: The role of
vasoactive compounds, growth factors and cytokines in the
progression of renal disease. Kidney Int Suppl. 75:S7–S14. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cunningham PN, Dyanov HM, Park P, Wang J,
Newell KA and Quigg RJ: Acute renal failure in endotoxemia is
caused by TNF acting directly on TNF receptor-1 in kidney. J
Immunol. 168:5817–5823. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xu C, Chang A, Hack BK, Eadon MT, Seth LA
and Cunningham PN: TNF-mediated damage to glomerular endothelium is
an important determinant of acute kidney injury in sepsis. Kidney
Int. 85:72–81. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang D, Pedraza PL, Abdullah HI, McGiff JC
and Ferreri NR: Calcium-sensing receptor-mediated TNF production in
medullary thick ascending limb cells. Am J Physiol Renal Physiol.
283:F963–F970. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Abdullah HI, Pedraza PL, Hao S, Rodland
KD, McGiff JC and Ferreri NR: NFAT regulates calcium-sensing
receptor-mediated TNF production. Am J Physiol Renal Physiol.
290:F1110–F1117. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Abdullah HI, Pedraza PL, McGiff JC and
Ferreri NR: CaR activation increases TNF production by mTAL cells
via a Gi-dependent mechanism. Am J Physiol Renal Physiol.
294:F345–F354. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hao S, Bellner L and Ferreri NR: NKCC2A
and NFAT5 regulate renal TNF production induced by hypertonic NaCl
intake. Am J Physiol Renal Physiol. 304:F533–F542. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu HJ, Wei Y, Ferreri NR, Nasjletti A and
Wang WH: Vasopressin and PGE(2) regulate the apical 70 pS K(+)
channel in the thick ascending limb of rat kidney. Am J Physiol
Cell Physiol. 278:C905–C913. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gu R, Jin Y, Zhai Y, Yang L, Zhang C, Li
W, Wang L, Kong S, Zhang Y, Yang B and Wang WH: PGE2 inhibits
basolateral 50 pS potassium channels in the thick ascending limb of
the rat kidney. Kidney Int. 74:478–485. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Battula S, Hao S, Pedraza PL, Stier CT and
Ferreri NR: Tumor necrosis factor-alpha is an endogenous inhibitor
of the Na+-K+-2Cl- cotransporter (NKCC2) isoform A in the thick
ascending limb. Am J Physiol Renal Physiol. 301:F94–F100. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
van Lanschot JJ, Mealy K, Jacobs DO, Evans
DA and Wilmore DW: Splenectomy attenuates the inappropriate
diuresis associated with tumor necrosis factor administration. Surg
Gynecol Obstet. 172:293–297. 1991.PubMed/NCBI
|
|
28
|
Shahid M, Francis J and Majid DS: Tumor
necrosis factor-alpha induces renal vasoconstriction as well as
natriuresis in mice. Am J Physiol Renal Physiol. 295:F1836–F1844.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nakamura K, Komagiri Y, Kojo T and
Kubokawa M: Effects of cytokines on activity of an inwardly
rectifying K+ channel in cultured human proximal tubule
cells. J Iwate Med Assoc. 59:375–385. 2007.
|
|
30
|
Nakamura K, Komagiri Y, Kojo T and
Kubokawa M: Delayed and acute effects of interferon-gamma on
activity of an inwardly rectifying K+ channel in cultured human
proximal tubule cells. Am J Physiol Renal Physiol. 296:F46–F53.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nietsch HH, Roe MW, Fiekers JF, Moore AL
and Lidofsky SD: Activation of potassium and chloride channels by
tumor necrosis factor. Role in liver cell death. J Biol Chem.
275:20556–20561. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chung CH, Fan J, Lee EY, Kang JS, Lee SJ,
Pyagay PE, Khoury CC, Yeo TK, Khayat MF, Wang A and Chen S: Effects
of tumor necrosis factor-α on podocyte expression of monocyte
chemoattractant protein-1 and in diabetic nephropathy. Nephron
Extra. 5:1–18. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kalantarinia K, Awad AS and Siragy HM:
Urinary and renal interstitial concentrations of TNF-alpha increase
prior to the rise in albuminuria in diabetic rats. Kidney Int.
64:1208–1213. 2003. View Article : Google Scholar : PubMed/NCBI
|