Introduction
Hepatocellular carcinoma (HCC) is the most common
type of liver cancer and is a principal health problem worldwide
(1). The majority of HCC cases are
associated with viral infection; hepatitis B virus (HBV) is the
principal cause of HCC in Asia (2–4). HBV
is epidemiologically associated with the development of liver
cirrhosis (5). HCC frequently
develops in the background of cirrhosis following chronic HBV
infection (6,7).
Bioinformatics analysis combined with microarray
technology has provided a novel method for comprehensively studying
the alterations in expression associated with disease at the
molecular level. Critical tumor-associated genes and cellular
signaling pathways have been identified in recent decades using
these methods. Therefore, examining the differentially expressed
genes (DEGs) between HCC and liver cirrhosis tissues is helpful for
identifying the key genes and pathways leading to tumor
progression.
However, previous studies have focused primarily on
progression from liver cirrhosis to HCC without regard to the
etiology of cirrhosis (8,9). Furthermore, analysis regarding the
associations between gene expression and patient survival is
lacking. At present, there is insufficient knowledge regarding the
molecular mechanisms for HCC transformation from liver cirrhosis in
patients with HBV infection.
In the present study, expression profiles from HCC
and liver cirrhosis tissue samples in a Gene Expression Omnibus
(GEO) dataset were compared to identify DEGs. Gene Ontology (GO),
Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment
and protein-protein interaction (PPI) network analyses were
performed using the DEGs. The prognostic value of the hub genes
from the PPI network was validated using survival and expression
data from The Cancer Genome Atlas (TCGA) database. The results of
the present study may help to identify the genes involved in the
progression of HBV-positive liver cirrhosis to HCC, and provide
information regarding biomarkers for verification and development
in further studies.
Materials and methods
Microarray data
The GSE17548 gene expression profile dataset, based
on the GPL570 platform [(HG-U133_Plus_2) Affymetrix Human Genome
U133 Plus 2.0 Array; Affymetrix; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA] and deposited by Yildiz et al (8), was downloaded from the GEO database
(www.ncbi.nlm.nih.gov/geo/). In total, 21
HBV-positive samples, including 11 liver cirrhosis and 10 HCC
tissues, were analyzed.
Identification of DEGs
The raw expression data were preprocessed using the
robust multiarray average method (10) in the affy package (11) in Bioconductor. Following
preprocessing, HCC tissue samples were compared with liver
cirrhosis tissues using an unpaired t-test in the Bioconductor
limma package (version 3.10.3; bioconductor.org/biocLite.R). An adjusted P<0.01
and |log fold change| >1 were set as thresholds for the
identification of DEGs.
GO term and KEGG pathway enrichment
analysis of the DEGs
Subsequent to obtaining the DEGs, GO term and KEGG
pathway enrichment analyses were performed using the Database for
Annotation, Visualization and Integrated Discovery (DAVID;
http://david.ncifcrf.gov/). GO (12) analysis determines the potential
functions of a list of genes, including the associated biological
process (BP), molecular function (MF) and cellular component (CC)
terms. KEGG (http://www.genome/ad.jp/kegg/) is a database of
genomes, biological pathways, associated diseases and possible drug
targets. P<0.05 was considered to indicate a statistically
significant difference for the GO and KEGG enrichment analyses.
PPI network construction
A PPI network of DEGs was constructed using the
Search Tool for the Retrieval of Interacting Genes (http://string.embl.de/) database and Cytoscape
software (version 3.6.0; www.cytoscape.org). As a large number of DEGs was
being analyzed, an interaction confidence score threshold of
>0.90 was set to avoid uncertain PPIs. PPI networks were
visualized with Cytoscape software. Modules from the PPI network
were screened using the Molecular Complex Detection (MCODE) plugin
(13) with the following criteria:
Degree cutoff, 2; node score cutoff, 0.2; k-core, 2; and max depth,
100. P<0.05 was used as a threshold value. The top 10 genes, as
ranked by degree, were considered the hub genes.
Survival validation in TCGA
The mRNA-sequencing expression profiles and clinical
data from HCC samples (14) were
downloaded from TCGA (https://cancergenome.nih.gov/). For the analysis of
hub genes identified in the PPI network, patients with cancer were
divided into different groups (high and low) based on the median
expression of each hub gene. To investigate the association of each
gene with overall survival, survival outcomes between the high and
low gene expression groups were compared using the Kaplan-Meier
method and log-rank test in the hash package in Bioconductor
(https://CRAN.R-project.org/package=hash). P<0.05
was considered to indicate a statistically significant difference
in survival.
Results
Data preprocessing and identification
of DEGs
Following background correction and normalization, a
total of 20,460 genes were annotated in the 21 HCC and liver
cirrhosis expression profiles (Fig.
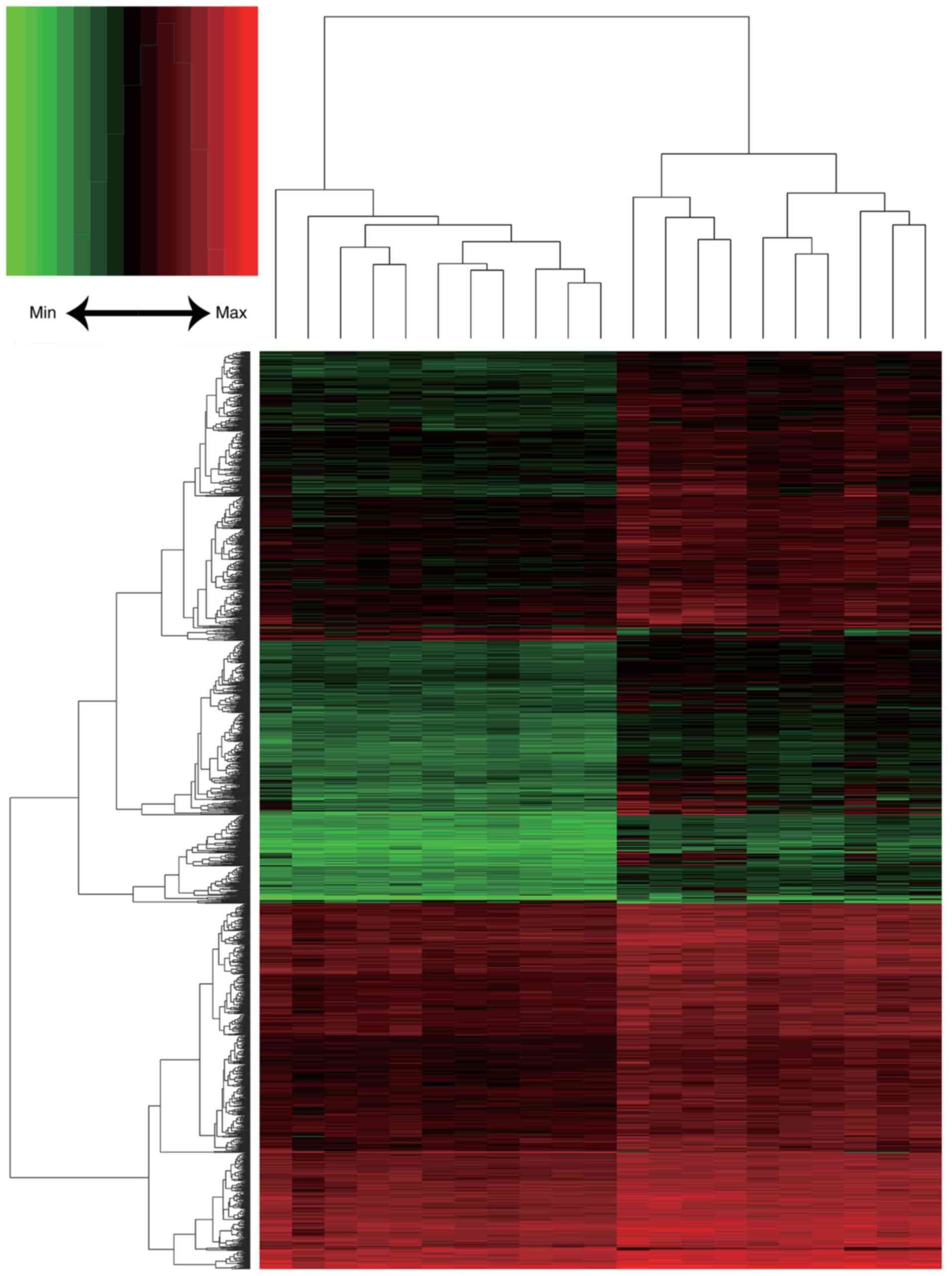
1). In total, 1,845 DEGs were identified, including 1,803
upregulated and 42 downregulated genes. A heat map of DEG
expression is presented in Fig.
2.
GO term and KEGG pathway enrichment
analysis
Using the DAVID tool, the GO term and KEGG pathway
enrichment of the identified DEGs was analyzed. GO BP analysis
suggested that the DEGs were primarily associated with the ‘cell
division’, ‘mitotic nuclear division’, ‘DNA replication’, ‘sister
chromatid cohesion’, ‘DNA replication initiation’, ‘G1/S
transition of mitotic cell cycle’, ‘telomere maintenance via
recombination’, ‘DNA synthesis involved in DNA repair’, ‘chromosome
segregation’ and ‘G2/M transition of mitotic cell cycle’
terms (Table I).
 | Table I.Gene Ontology term enrichment
analysis of the differentially expressed genes (top 10 terms
selected according to P-value). |
Table I.
Gene Ontology term enrichment
analysis of the differentially expressed genes (top 10 terms
selected according to P-value).
| A, Biological
process |
|---|
|
|---|
| Term | Description | Count | P-value |
|---|
| GO:0051301 | Cell division | 96 |
1.40×10−26 |
| GO:0007067 | Mitotic nuclear
division | 70 |
2.80×10−20 |
| GO:0006260 | DNA
replication | 52 |
9.60×10−19 |
| GO:0007062 | Sister chromatid
cohesion | 37 |
1.69×10−14 |
| GO:0006270 | DNA replication
initiation | 18 |
5.43×10−11 |
| GO:0000082 | G1/S
transition of mitotic cell cycle | 32 |
7.00×10−11 |
| GO:0000722 | Telomere
maintenance via recombination | 16 |
7.09×10−9 |
| GO:0000731 | DNA synthesis
involved in DNA repair | 15 |
2.63×10−7 |
| GO:0007059 | Chromosome
segregation | 21 |
2.99×10−7 |
| GO:0000086 | G2/M
transition of mitotic cell cycle | 31 |
5.38×10−7 |
|
| B, Cellular
component |
|
| Term |
Description | Count | P-value |
|
| GO:0005654 | Nucleoplasm | 354 |
2.06×10−21 |
| GO:0005634 | Nucleus | 565 |
6.15×10−15 |
| GO:0005737 | Cytoplasm | 523 |
2.26×10−10 |
| GO:0000777 | Condensed
chromosome kinetochore | 26 |
9.69×10−9 |
| GO:0000775 | Chromosome,
centromeric region | 20 |
4.06×10−8 |
| GO:0005829 | Cytosol | 343 |
5.61×10−8 |
| GO:0000776 | Kinetochore | 23 |
2.22×10−7 |
| GO:0030496 | Midbody | 30 |
2.56×10−7 |
| GO:0005813 | Centrosome | 65 |
6.05×10−7 |
| GO:0005819 | Spindle | 26 |
8.38×10−6 |
|
| C, Molecular
function |
|
| Term |
Description | Count | P-value |
|
| GO:0005515 | Protein
binding | 842 |
2.67×10−11 |
| GO:0019901 | Protein kinase
binding | 60 |
1.10×10−6 |
| GO:0003677 | DNA binding | 180 |
1.19×10−4 |
| GO:0005524 | ATP binding | 163 |
1.33×10−4 |
| GO:0004674 | Protein
serine/threonine kinase activity | 50 |
9.04×10−4 |
| GO:0004842 | Ubiquitin-protein
transferase activity | 45 |
9.53×10−4 |
| GO:0003697 | Single-stranded DNA
binding | 18 | 0.001422 |
| GO:0016874 | Ligase
activity | 38 | 0.001512 |
| GO:0043130 | Ubiquitin
binding | 16 | 0.00157 |
| GO:0042802 | Identical protein
binding | 85 | 0.002182 |
In the GO CC category, the ‘nucleoplasm’, ‘nucleus’,
‘cytoplasm’, ‘condensed chromosome kinetochore’, ‘chromosome,
centromeric region’, ‘cytosol’, ‘kinetochore’, ‘midbody’,
‘centrosome’ and ‘spindle’ terms were enriched in the DEGs
(Table I).
In the GO MF category, the ‘protein binding’,
‘protein kinase binding’, ‘DNA binding’, ‘ATP binding’, ‘protein
serine/threonine kinase activity’, ‘ubiquitin-protein transferase
activity’, ‘single-stranded DNA binding’, ‘ligase activity’,
‘ubiquitin binding’ and ‘identical protein binding’ terms were
enriched (Table I).
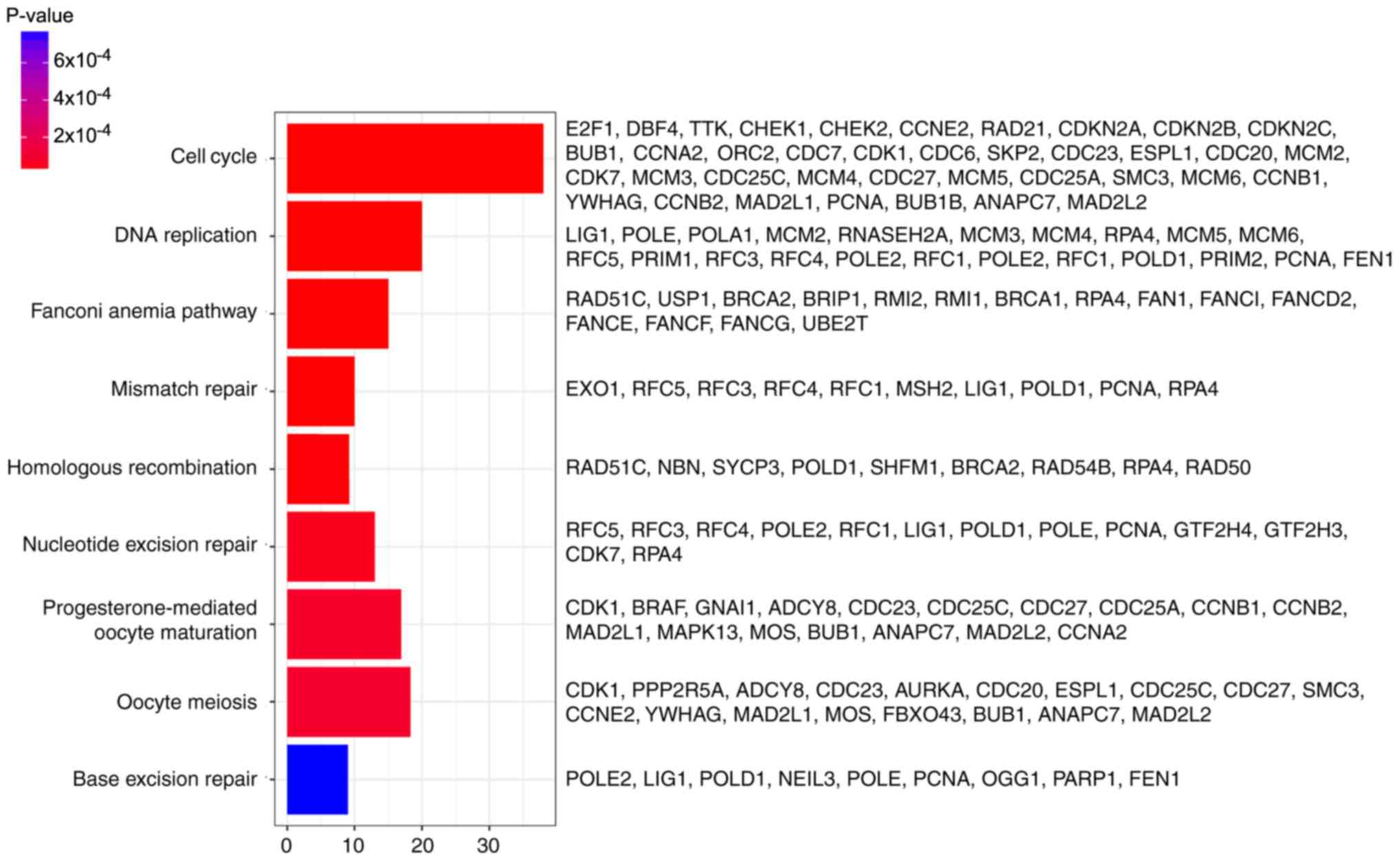
In addition, DEGs were primarily associated with the
‘cell cycle’, ‘DNA replication’, ‘fanconi anemia pathway’,
‘mismatch repair’, ‘homologous recombination’, ‘nucleotide excision
repair’, ‘progesterone-mediated oocyte maturation’, ‘oocyte
meiosis’ and ‘base excision repair’ pathways, determined by KEGG
analysis (Fig. 3).
PPI network and module selection
The PPI network consisted of 685 nodes and 4,603
edges (Fig. 4A). According to the
degree for each gene, the top 10 hub genes were cyclin dependent
kinase 1 (CDK1), cell division cycle 20, cyclin B1 (CCNB1), cyclin
B2 (CCNB2), mitotic arrest deficient 2 like 1 (MAD2L1), aurora
kinase B, BUB1 mitotic checkpoint serine/threonine kinase (BUB1),
cyclin A2 (CCNA2), centromere protein E and kinesin family member
2C (Table II). Using the MCODE
Cytoscape plugin, four significant modules were identified
(Fig. 4B-E). An enrichment
analysis of the genes involved in the top significant modules was
performed. The results identified that the DEGs in modules were
principally related to ubiquitin mediated proteolysis, cell cycle,
oocyte meiosis, progesterone-mediated oocyte maturation, HTLV-I
infection, p53 signaling pathway and Epstein-Barr virus
infection.
| Table II.Core identified differentially
expressed genes and their corresponding degrees. |
Table II.
Core identified differentially
expressed genes and their corresponding degrees.
| Gene | Degree |
|---|
| CDK1 | 129 |
| CDC20 | 101 |
| CCNB1 | 93 |
| CCNB2 | 81 |
| MAD2L1 | 77 |
| AURKB | 76 |
| BUB1 | 68 |
| CCNA2 | 67 |
| CENPE | 65 |
| KIF2C | 64 |
Validation with TCGA data
Information regarding 57,000 genes from 424 patients
was included in the TCGA mRNA-sequencing expression data. Patients
were sorted into high or low expression groups according to the
median expression value. As CDK1, CCNB1, CCNB2, MAD2L1, BUB1 and
CCNA2 were enriched in the ‘cell cycle’ and the
‘progesterone-mediated oocyte maturation’ pathways, the
significance of these six hub genes in prognosis with TCGA data was
subsequently validated with the Kaplan-Meier method. It was
identified that the high expression of CDK1, CCNB1, CCNB2, MAD2L1,
BUB1 and CCNA2 was associated with a decreased overall survival for
patients with HCC (P=0.00116, 0.00021, 0.04889, 0.00222, 0.00015
and 0.00647, respectively; Fig.
5).
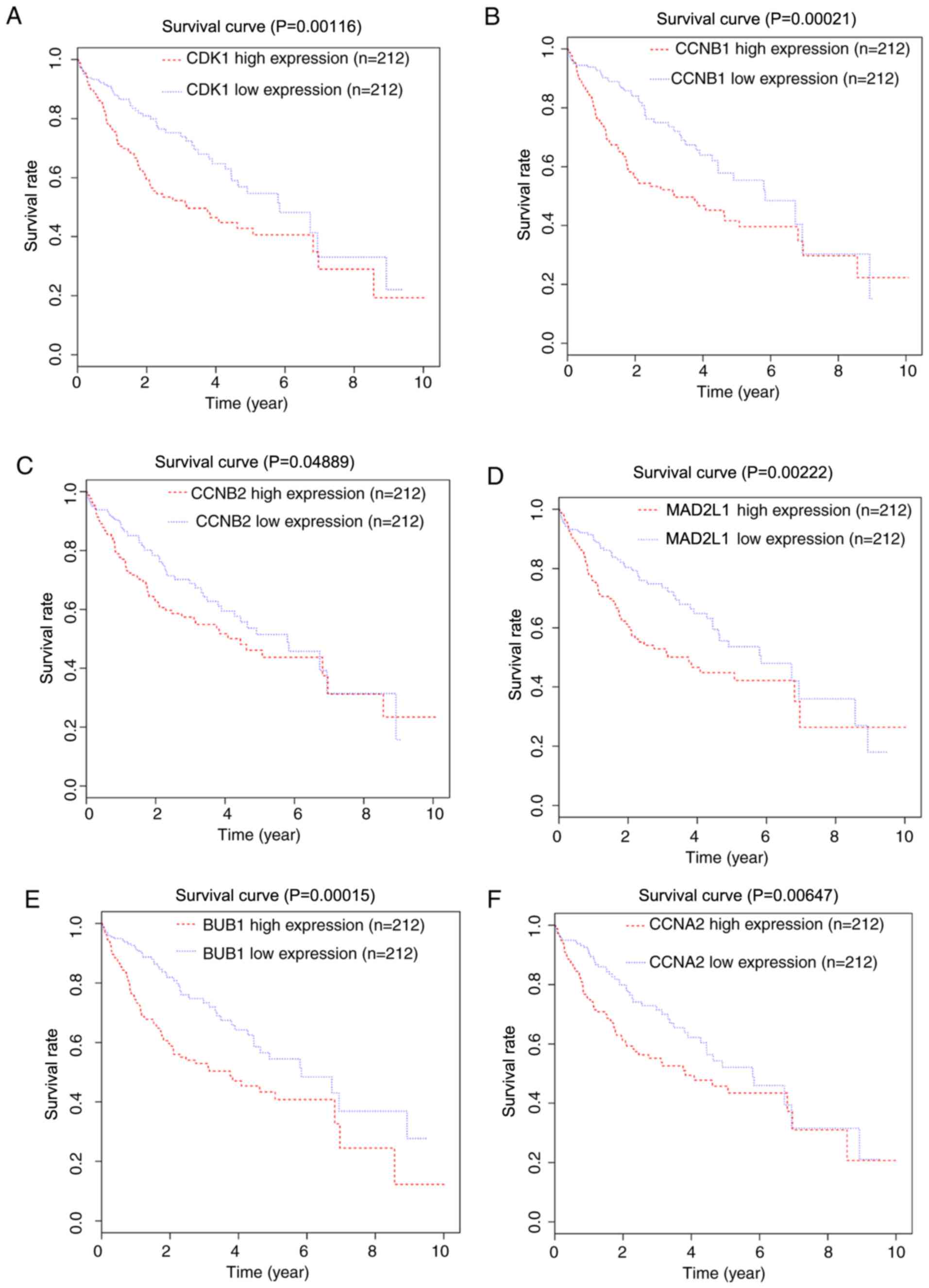 | Figure 5.Kaplan-Meier survival curves.
Kaplan-Meier survival curves for patients stratified according to
the median expression of (A) CDK1, (B) CCNB1, (C) CCNB2, (D)
MAD2L1, (E) BUB1 and (F) CCNA2. CDK1, cyclin dependent kinase 1;
CCNB1, cyclin B1; CCNB2, cyclin B2; MAD2L1, mitotic arrest
deficient 2 like 1; BUB1, BUB1 mitotic checkpoint serine/threonine
kinase; CCNA2, cyclin A2. |
Discussion
Different from the previous studies conducted by
Yildiz et al (8) and He
et al (9), which analyzed
the key genes and pathways for HCC developing from liver cirrhosis
regardless of etiology, the present study was conducted
specifically with HBV-positive samples. Therefore, drugs targeting
these identified key genes and pathways may be considered for HCC
therapy.
The hub genes CDK1, CCNB1, CCNB2, MAD2L1, BUB1 and
CCNA2 may be suitable for the diagnosis and treatment of
HBV-associated HCC. Garnier et al (15) identified that CDK1 may enhance
liver regeneration by promoting rapid and efficient hepatocyte
proliferation in a rat model, which could promote mutations in the
β-catenin gene, potentially promoting the progression to HCC
(16). Zhang et al
(17) suggested that miR-582-5p
inhibits the proliferation of HCC cells by targeting CDK1. However,
in a comparative study of HCC and cirrhosis, Masaki et al
(18) observed that the expression
of CDK1 did not differ between HCC and the adjacent cirrhotic
tissues in all the tested patients with hepatitis C virus-RNA.
Therefore, alterations of the role of CDK1 in HCC, depending on
different viral types, require further evaluation. Wang et
al (19) demonstrated that the
increased expression of CCNB1 and CCNB2 was associated with the
restoration of the young regenerating liver phenotype. Another
previous study identified that CCNB1 was associated with
G2/M phase cell cycle arrest-induced growth inhibition,
suggesting that this gene may be suitable as an anti-cancer agent
in future therapy (20). Sze et
al (21) suggested that
mitotic arrest deficient (MAD)1 interacted with MAD2, leading to
its retention in the cytoplasm, which may oppose mitotic checkpoint
control in hepatocarcinogenesis. Similarly, Sze et al
(22) suggested that MAD2
deficiency may cause a mitotic checkpoint defect in hepatoma cells.
Wills et al (23) suggested
that BUB1, a loss-of-heterozygosity driver gene, may be implicated
in the germline causes of genetic instability in polycystic liver
disease. Furthermore, Wang et al (24) identified that BUB1B activated
parathyroid hormone like hormone to promote the G-protein-coupled
receptor-induced loss of cell adhesion. CCNA functions in the S and
G2-M phases of the cell cycle, and CCNA2 overexpression
is associated with carcinogenesis in the liver (25–27).
Masaki et al (18)
suggested that the activation of CCNA expression may serve an
essential role in the transformation of cirrhosis to HCC. Wang
et al (28) additionally
identified that viral insertion was able to contribute to
tumorigenesis in HBV-associated HCC by disrupting the control of
the CCNA gene. Furthermore, Nault et al (29) identified that adeno-associated
virus type 2 was associated with oncogenic insertional mutagenesis
in human HCC, including in CCNA2. Yang et al (30) observed that bile acid-activated
farnesoid X receptor stimulated the miR-22 silencing of CCNA2,
exerting a protective effect in HCC cells.
It is noteworthy that CDK1, CCNB1, CCNB2 and MAD2L1
were enriched in the ‘p53 signaling pathway’. Cellular tumor
antigen p53 (p53), discovered in 1979, suppresses cell growth and
oncogenic transformation (31).
The disruption of p53 protein expression is an important factor
underlying tumorigenesis in humans. Reportedly, ≤50% of all
malignant tumors exhibit p53 mutations (32). Furthermore, the p53 response
pathway is frequently defective in HCC (33), and p53 status is associated with
the prognosis of patients with HCC (34). This is consistent with the poor
prognosis associated with the high expression of CDK1, CCNB1, CCNB2
and MAD2L1 in the present analysis.
The present study demonstrated that dysregulation of
the cell cycle and progesterone-mediated oocyte maturation pathways
was closely associated with the development and progression of HCC.
It was established that the dysregulation of the cell cycle pathway
is an initiating event in cancer (35). Cell cycle checkpoints ensure normal
genetic stability, the dysregulation of which may cause deviant
cell proliferation leading to tumorigenic mutations (36). Wong et al (37) identified that an abnormal cell
cycle is crucial in the development of cancer, subsequent to
analyzing cancer samples from early to late stage. Furthermore, the
dysfunction of cell cycle regulators and checkpoint mediators in
HCC was discussed in previous studies (38–40).
Dituri et al (39)
demonstrated that alterations in the expression of cell cycle
checkpoint-associated proteins frequently occur during HCC
development. Yang et al (41) identified that short hairpin RNAs
were able to suppress HBV-associated HCC cell proliferation through
an effect on the cell cycle pathway. Previous studies concerning
the function of the progesterone-mediated oocyte maturation pathway
in liver cancer development are limited. In agreement with the
study conducted by Jin et al (42), the progesterone-mediated oocyte
maturation pathway was significantly enriched in the DEGs in the
present study. In addition, Duckworth et al (43) identified that protein kinase A
negatively regulates cell division cycle 25C during oocyte
maturation. Further studies investigating the function of the
progesterone-mediated oocyte maturation pathway in liver cancer are
required.
In summary, the present study provided a
comprehensive analysis of the DEGs between HCC and cirrhotic liver
tissue samples through bioinformatics analysis to identify the
potential genes and pathways involved in the transformation of
HBV-positive liver cirrhosis to HCC. The identified genes and
pathways may be suitable for use as therapeutic targets or novel
biomarkers for prognosis. However, further studies are required to
confirm these results and determine the underlying mechanisms.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and
Technology Planning Project of Guangdong (Guangdong, China; grant
no. 2014A020212532).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
QC, JX and PW conceived and designed the study. QC,
JX, WL, LS and TH analyzed the data. TH performed literature
searches. QC, JX and TH wrote the paper. QC, WL, LS, TH and PW
reviewed and edited the manuscript. All authors read and approved
the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen QF, Jia ZY, Yang ZQ, Fan WL and Shi
HB: Transarterial chemoembolization monotherapy versus combined
transarterial chemoembolization-microwave ablation therapy for
hepatocellular carcinoma tumors ≤5 cm: A propensity analysis at a
single center. Cardiovasc Intervent Radiol. 40:1748–1755. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
El-Serag HB: Epidemiology of viral
hepatitis and hepatocellular carcinoma. Gastroenterology.
142:1264–1273, e1. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Levrero M and Zucman-Rossi J: Mechanisms
of HBV-induced hepatocellular carcinoma. J Hepatol. 64 1
Suppl:S84–S101. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Parkin DM: The global health burden of
infection-associated cancers in the year 2002. Int J Cancer.
118:3030–3044. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhou HY, Luo Y, Chen WD and Gong GZ:
Hepatitis B virus mutation may play a role in hepatocellular
carcinoma recurrence: A systematic review and meta-regression
analysis. J Gastroenterol Hepatol. 30:977–983. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Intaraprasong P, Siramolpiwat S and
Vilaichone RK: Advances in management of hepatocellular carcinoma.
Asian Pac J Cancer Prev. 17:3697–3703. 2016.PubMed/NCBI
|
|
7
|
Sherman M: Hepatocellular carcinoma:
Epidemiology, surveillance, and diagnosis. Semin Liver Dis.
30:3–16. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yildiz G, Arslan-Ergul A, Bagislar S, Konu
O, Yuzugullu H, Gursoy-Yuzugullu O, Ozturk N, Ozen C, Ozdag H,
Erdal E, et al: Genome-wide transcriptional reorganization
associated with senescence-to-immortality switch during human
hepatocellular carcinogenesis. PLoS One. 8:e640162013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
He B, Yin J, Gong S, Gu J, Xiao J, Shi W,
Ding W and He Y: Bioinformatics analysis of key genes and pathways
for hepatocellular carcinoma transformed from cirrhosis. Medicine
(Baltimore). 96:e69382017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Irizarry RA, Hobbs B, Collin F,
Beazer-Barclay YD, Antonellis KJ, Scherf U and Speed TP:
Exploration, normalization, and summaries of high density
oligonucleotide array probe level data. Biostatistics. 4:249–264.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gautier L, Cope L, Bolstad BM and Irizarry
RA: Affy-analysis of Affymetrix GeneChip data at the probe level.
Bioinformatics. 20:307–315. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The Gene
Ontology Consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bader GD and Hogue CW: An automated method
for finding molecular complexes in large protein interaction
networks. BMC Bioinformatics. 4:22003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu Y, Qiu P and Ji Y: TCGA-assembler:
Open-source software for retrieving and processing TCGA data. Nat
Methods. 11:599–600. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Garnier D, Loyer P, Ribault C,
Guguen-Guillouzo C and Corlu A: Cyclin-dependent kinase 1 plays a
critical role in DNA replication control during rat liver
regeneration. Hepatology. 50:1946–1956. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nhieu JT, Renard CA, Wei Y, Cherqui D,
Zafrani ES and Buendia MA: Nuclear accumulation of mutated
beta-catenin in hepatocellular carcinoma is associated with
increased cell proliferation. Am J Pathol. 155:703–710. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang Y, Huang W, Ran Y, Xiong Y, Zhong Z,
Fan X, Wang Z and Ye Q: miR-582-5p inhibits proliferation of
hepatocellular carcinoma by targeting CDK1 and AKT3. Tumour Biol.
36:8309–8316. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Masaki T, Shiratori Y, Rengifo W, Igarashi
K, Yamagata M, Kurokohchi K, Uchida N, Miyauchi Y, Yoshiji H,
Watanabe S, et al: Cyclins and cyclin-dependent kinases:
Comparative study of hepatocellular carcinoma versus cirrhosis.
Hepatology. 37:534–543. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang X, Quail E, Hung NJ, Tan Y, Ye H and
Costa RH: Increased levels of forkhead box M1B transcription factor
in transgenic mouse hepatocytes prevent age-related proliferation
defects in regenerating liver. Proc Natl Acad Sci USA.
98:11468–11473. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang G, Chen H, Huang M, Wang N, Zhang J,
Zhang Y, Bai G, Fong WF, Yang M and Yao X: Methyl protodioscin
induces G2/M cell cycle arrest and apoptosis in HepG2 liver cancer
cells. Cancer Lett. 241:102–109. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sze KM, Ching YP, Jin DY and Ng IO: Role
of a novel splice variant of mitotic arrest deficient 1 (MAD1),
MAD1beta, in mitotic checkpoint control in liver cancer. Cancer
Res. 68:9194–9201. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sze KM, Ching YP, Jin DY and Ng IO:
Association of MAD2 expression with mitotic checkpoint competence
in hepatoma cells. J Biomed Sci. 11:920–927. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wills ES, Cnossen WR, Veltman JA,
Woestenenk R, Steehouwer M, Salomon J, Morsche Te RH, Huch M,
Hehir-Kwa JY, Banning MJ, et al: Chromosomal abnormalities in
hepatic cysts point to novel polycystic liver disease genes. Eur J
Hum Genet. 24:1707–1714. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang L, Huang J, Jiang M, Lin H, Qi L and
Diao H: Activated PTHLH coupling feedback phosphoinositide to
G-protein receptor signal-induced cell adhesion network in human
hepatocellular carcinoma by systems-theoretic analysis.
ScientificWorldJournal. 2012:4289792012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rinaudo Spiewak JA and Thorgeirsson SS:
Detection of a tyrosine-phosphorylated form of cyclin A during
liver regeneration. Cell Growth Differ. 8:301–309. 1997.PubMed/NCBI
|
|
26
|
Kim DH, Park SE, Kim M, Ji YI, Kang MY,
Jung EH, Ko E, Kim Y, Kim S, Shim YM and Park J: A functional
single nucleotide polymorphism at the promoter region of cyclin A2
is associated with increased risk of colon, liver, and lung
cancers. Cancer. 117:4080–4091. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Simile MM, De Miglio MR, Muroni MR, Frau
M, Asara G, Serra S, Muntoni MD, Seddaiu MA, Daino L, Feo F and
Pascale RM: Down-regulation of c-myc and Cyclin D1 genes by
antisense oligodeoxy nucleotides inhibits the expression of E2F1
and in vitro growth of HepG2 and Morris 5123 liver cancer cells.
Carcinogenesis. 25:333–341. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang J, Chenivesse X, Henglein B and
Brechot C: Hepatitis B virus integration in a cyclin A gene in a
hepatocellular carcinoma. Nature. 343:555–557. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nault JC, Datta S, Imbeaud S, Franconi A,
Mallet M, Couchy G, Letouzé E, Pilati C, Verret B, Blanc JF, et al:
Recurrent AAV2-related insertional mutagenesis in human
hepatocellular carcinomas. Nat Genet. 47:1187–1193. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang F, Hu Y, Liu HX and Wan YJ:
MiR-22-silenced cyclin A expression in colon and liver cancer cells
is regulated by bile acid receptor. J Biol Chem. 290:6507–6515.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Olson DC and Levine AJ: The properties of
p53 proteins selected for the loss of suppression of
transformation. Cell Growth Differ. 5:61–71. 1994.PubMed/NCBI
|
|
32
|
Vaughan C, Pearsall I, Yeudall A, Deb SP
and Deb S: p53: Its mutations and their impact on transcription.
Subcell Biochem. 85:71–90. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hussain SP, Schwank J, Staib F, Wang XW
and Harris CC: TP53 mutations and hepatocellular carcinoma:
Insights into the etiology and pathogenesis of liver cancer.
Oncogene. 26:2166–2176. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Olivier M, Hollstein M and Hainaut P: TP53
mutations in human cancers: Origins, consequences, and clinical
use. Cold Spring Harb Perspect Biol. 2:a0010082010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wong YH, Chen RH and Chen BS: Core and
specific network markers of carcinogenesis from multiple cancer
samples. J Theor Biol. 362:17–34. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Elledge SJ: Cell cycle checkpoints:
Preventing an identity crisis. Science. 274:1664–1672. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wong YH, Li CW and Chen BS: Evolution of
network biomarkers from early to late stage bladder cancer samples.
Biomed Res Int. 2014:1590782014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cheung KF, Zhao J, Hao Y, Li X, Lowe AW,
Cheng AS, Sung JJ and Yu J: CITED2 is a novel direct effector of
peroxisome proliferator-activated receptor γ in suppressing
hepatocellular carcinoma cell growth. Cancer. 119:1217–1226. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dituri F, Mazzocca A, Lupo L, Edling CE,
Azzariti A, Antonaci S, Falasca M and Giannelli G: PI3K class IB
controls the cell cycle checkpoint promoting cell proliferation in
hepatocellular carcinoma. Int J Cancer. 130:2505–2513. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Furuta M, Kozaki K, Tanimoto K, Tanaka S,
Arii S, Shimamura T, Niida A, Miyano S and Inazawa J: The
tumor-suppressive miR-497-195 cluster targets multiple cell-cycle
regulators in hepatocellular carcinoma. PLoS One. 8:e601552013.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yang Y, Zheng B, Han Q, Zhang C, Tian Z
and Zhang J: Targeting blockage of STAT3 inhibits hepatitis B
virus-related hepatocellular carcinoma. Cancer Biol. 17:449–456.
2016. View Article : Google Scholar
|
|
42
|
Jin B, Wang W, Du G, Huang GZ, Han LT,
Tang ZY, Fan DG, Li J and Zhang SZ: Identifying hub genes and
dysregulated pathways in hepatocellular carcinoma. Eur Rev Med
Pharmacol Sci. 19:592–601. 2015.PubMed/NCBI
|
|
43
|
Duckworth BC, Weaver JS and Ruderman JV:
G2 arrest in Xenopus oocytes depends on phosphorylation of cdc25 by
protein kinase A. Proc Natl Acad Sci USA. 99:16794–16799. 2002.
View Article : Google Scholar : PubMed/NCBI
|