Introduction
Nuclear receptors (NRs) are members of a large
superfamily of transcription factors in metazoans, which are
involved in complex biological processes and are major
pharmacological targets (1–3). The
dysfunction of NR activities is associated with a range of
diseases, including diabetes, Alzheimer's disease, cancer and
cardiovascular disease (4). NRs
are intracellular signaling proteins that bind to specific ligands,
particularly small lipid-soluble molecules, including steroids,
thyroxine, retinoic acid and vitamin D.
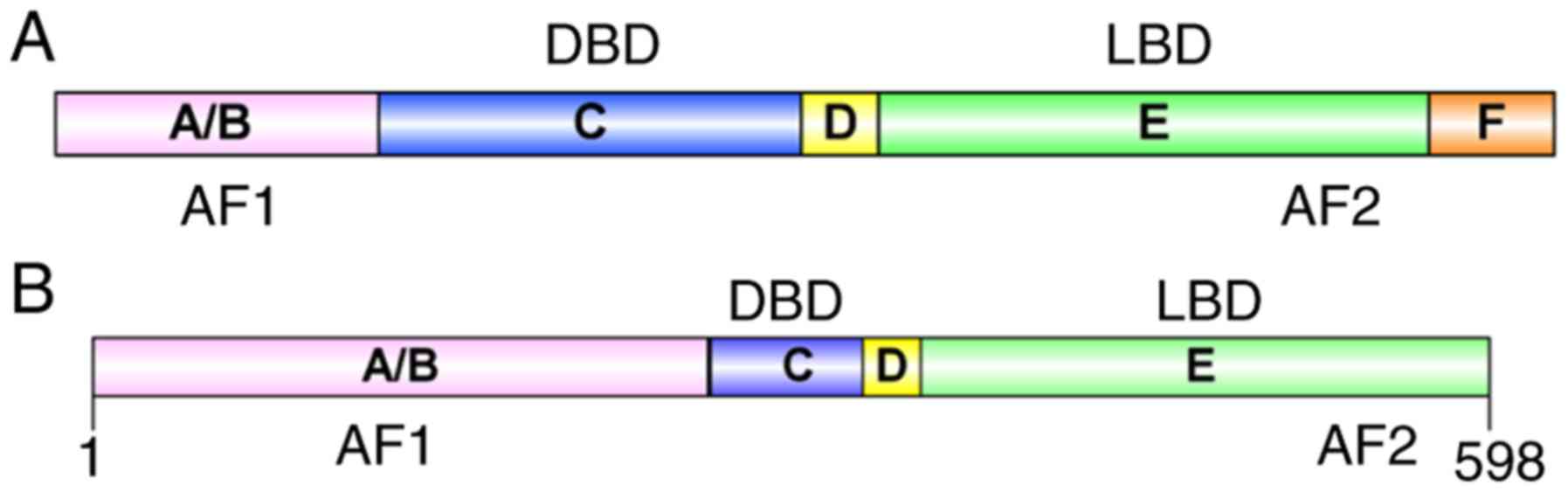
Structurally, NRs are typically divided into five
regions: A/B region, C region, D region, E region and F region
(Fig. 1A). The main domains of NRs
are the N-terminal domain, the conserved DNA-binding domain (DBD)
and the C-terminal ligand-binding domain (LBD) (5). The A/B region, which contains the
ligand-independent activation function 1 (AF-1) domain, has been
described as intrinsically disordered, and is involved in
protein-protein interactions and receptor functions (6). In the C region, the general protein
structure is similar among NRs (7,8). The
DBD within the C region includes the zinc finger transcription
factor motif, in which eight conserved cysteine residues coordinate
with two zinc ions and mediate homodimerization or
heterodimerization (2). Region D
is variable, with a flexible hinge region that connects the C and E
regions. This region often contains a nuclear localization sequence
(NLS) and can alter the structure of the protein, including
increasing the flexibility of the protein (5,9).
Region E contains a ligand-binding pocket buried in a helical
globular domain where agonists or antagonists bind. Region E is
also involved in homologous and heterologous dimerization between
receptor monomers, and contains the ligand-dependent activation
function 2 (AF-2) domain, to which cofactors and repressors bind
(5,9). The F region is only present in
certain NRs and there is marked variation in the sequence; however,
the structure and function of this region remain to be fully
elucidated.
NRs can be divided into three categories based on
the different types of ligands they bind with: Class I includes
steroid hormone receptors, including androgen receptor, estrogen
receptor and glucocorticoid receptor; class II includes non-steroid
hormone receptors, including thyroid hormone receptor, retinoic
acid receptor and vitamin D receptor; and class III includes orphan
nuclear hormone receptors, including nuclear receptor subfamily 4
group A member 1 (Nur77), nuclear receptor subfamily 0 group B
member 1 (NR0B1) and nuclear receptor subfamily 0 group B member 2
(NR0B2/SHP) (10). NRs have been
crucial in the development of targeted drugs, including BMS-564929
and ketoconazole (11). Currently,
~13% of clinically applied drugs target NRs. Therefore, NRs have
been established as useful targets for drug development (12).
Identification of Nur77
The Nur77 gene (also known as
NR4A1/NGFI-B/TR3/NAK-1/GFRP1/HMR/N10/NP10) is located at chromosome
12, NC_000012.12 (52,022,832–52,059,507) in humans. Nur77 was first
identified in 1988 by Hazel et al (13), having previously identified several
immediate early genes expressed during the G0/G1 transition in
mouse fibroblasts. The transcriptional activity of these genes is
activated following stimulation with serum or growth factors. The
nucleotide sequence of one of the cDNA clones, Nur77 (initially
termed 3CH77), was revealed to encode a member of the
ligand-binding transcription factor superfamily, which included
steroids and thyroid hormone receptors (13). Subsequently, the Nur77 rat homolog,
nerve growth factor-induced clone B (NGFI-B), was successfully
cloned from rat adrenal pheochromocytoma cells (PC-12) by Watson
and Milbrandt (14). In the same
year, testicular receptor 3, a human homologue of Nur77, was
identified by Chang et al (15) from a cDNA library of human prostate
cancer cells. An increasing number of studies have shown that this
transcription factor is present in various species, and it is
recognized as a member of NR subfamily 4 group A (16,17).
Structure, expression and localization of
Nur77
Structure of Nur77
The Nur77 protein consists of 598 amino acids and
contains A/B, C, D, and E domains (Fig. 1B) (18). Rehman et al (19) identified two protein subtypes in
mice that lack the Nur77 N-terminal domain, and the localization of
these isoforms was predicted to be predominantly outside the
nucleus. Therefore, N-terminal transactivation domain may be
required for the transport of Nur77 from the nucleus to the
cytoplasm. The DBD region recognizes the specific NGFI-B response
element (NBRE; sequence: AAAGGTCA) in target genes and regulates
the expression level of these genes (20,21).
In addition, Nur77 and retinoid X receptors (RXRs) form
heterologous dimers that can combine with the DR5 response element
(sequence: AGGTCA-NNNAA-AGGTCA, N: Any single nucleotide) to
regulate the transcription of target genes (22,23).
The LBD region of Nur77 is distinct from the typical LBD region
present in other NRs. The human Nur77 crystal structure (Protein
Data Bank-ID: 2QW4; http://www.rcsb.org/structure/2QW4) shows that the
Nur77 LBD region is blocked by hydrophobic residues (24,25).
Several NRs have been reported to have a hydrophobic-cleft
regulated by helices 3, 5 and 12, which is important for the
recruitment of co-activators or co-repressors involved in
transcriptional regulation (26).
However, it is noteworthy that, this cleft is hydrophilic in the
Nur77 protein. Additionally, partial denaturation experiments have
revealed that helix 12 is relatively flexible in Nur77 (27). Notably, Moore et al
(28) identified NR alternate-site
modulators that bind with alternative pockets of a protein, rather
than the classical LBD, which may result in a different
function.
Expression of Nur77
Nur77 has received specific interest in the
scientific community due to its role in apoptosis and cancer. The
expression level and location of Nur77 are important for protein
function. For example, in cells treated with an
n-butylidenephthalide derivative (PCH4), the expression of Nur77
was shown to be increased and the protein migrated from the nucleus
to the cytoplasm, which may inhibit the growth of malignant glioma
cell growth and induce apoptosis (29). Therefore, PCH4 may be useful as a
novel agent for the treatment of malignant glioma. Subsequently,
the overexpression of Nur77 in breast cancer cell xenografts was
reported to alter the inflammatory response and increase the risk
of metastatic disease in mice (30). Another study suggested that the
cytoplasmic expression of Nur77 can induce apoptosis in breast
cancer cells (31). Notably, one
report demonstrated that Nur77 is involved in the migration of
transforming growth factor-β-induced breast cancer cells (32); furthermore, this process is
dependent on p38α (33). Delgado
et al (34) reported that
the expression of Nur77 in ovaries was high relative to that in
other tissues, with only skeletal muscle and tracheal tissue having
higher levels of expression than the ovaries. In normal ovarian
tissues, Nur77 is predominantly localized in the nucleus, whereas
in ovarian cancer cell lines and tissues from patients with ovarian
cancer, Nur77 is present in the cytoplasm and the nucleus. In
addition, Nur77 was shown to be expressed at high levels in samples
from a subset of patients with high-grade serous ovarian cancer,
who had a poor progression-free survival rate (34), suggesting that Nur77 mediates the
growth of cancer cells. Furthermore, Zhang et al (35) demonstrated that the expression of
Nur77 was markedly reduced in the placenta of women with
preeclampsia compared with placental levels during a normal
pregnancy, and that Nur77 may also be involved in regulating
trophoblast cell motility.
Localization of Nur77
The following section considers what determines the
positioning of Nur77 in cells. In normal tissues, Nur77 is located
in the nucleus due to the NLS in the protein structure;
additionally, nuclear Nur77 acts as a carcinogenic survival factor
and promotes the growth of cancer cells. By contrast, Nur77 is a
potent death promoter when it is located in the mitochondria, where
it binds and induces conformational changes in B-cell lymphoma-2
(Bcl-2) protein, triggering the release of cytochrome c and
apoptosis (36). There are several
potential explanations for this dual function of Nur77. Firstly,
the classical hydrophobic nuclear export signal is crucial in
Nur77. RXRα acts as an active partner in transporting Nur77 from
the nucleus to the cytoplasm, however, translocation of the
RXRα/Nur77 heterodimer appears to largely depend on the cell type
and stimulus (37,38). Furthermore, studies have reported
that the phosphorylation status of Nur77 is vital in the nuclear to
cytoplasmic translocation of Nur77, which is regulated by the
mitogen-activated protein kinase (MAPK)/c-Jun N-terminal kinase and
protein kinase B (Akt) pathways (34,39,40).
During oxidative stress-induced cell death in SH-SY5Y cells, Nur77
is upregulated and is translocated from the nucleus to the cytosol
and mitochondria (41).
Additionally, agents that induce the migration of Nur77 from the
nucleus to the mitochondria effectively induce the apoptosis of
cancer cells. For example, following glutamate treatment, Nur77 is
translocated from the nucleus to the cytosol and mitochondria in
rat cerebellar granule neurons (42). Additionally, by inhibiting the
migration of Nur77 from the nucleus to the cytoplasm, 17β-estradiol
can delay 6-hydroxydopamine-induced apoptosis (43). Endogenous insulin-like growth
factor-binding protein 3 has been reported to facilitate the
phosphorylation of Nur77 and the export of Nur77 from the nucleus
to the cytoplasm, which is vital in intrinsic apoptosis (44). Furthermore, bile acids regulate the
expression and intracellular location of Nur77 to control cell
survival and death (45).
Genomic and non-genomic functions of the
orphan receptor Nur77
Genomic functions of Nur77
Nur77 is an immediate-early gene and an important
transcription factor. The protein was first identified in the
monomeric form bound to its response element NBRE. Nur77 has been
reported to form heterodimers with RXR/nuclear receptor subfamily 4
group A member 2 (Nurr1)/nuclear receptor subfamily 4 group A
member 3, and these dimers specifically bind to DR5 or NBRE
elements (46). Nur77 can also
form homodimers and bind to a novel Nur response element, which has
a palindromic structure and was identified in the regulatory region
of the pro-opiomelanocortin gene. Nur77 is responsive to
physiological stimuli in endocrine and lymphoid cells (21,46).
Furthermore, the hypothalamic corticotropin-releasing hormone and
protein kinase A rapidly increase the nuclear DNA binding activity
of Nur77 dimers, but do not affect monomers. The AF-1 domain of
Nur77 and the transcriptional mediators/intermediary factor 2, also
known as steroid receptor coactivator-2 (SRC-2) glutamine-rich
domain are also required for this DNA binding activity (47). Tripartite motif containing 28, also
known as KAP-1 or KPIP-1, acts as a transcriptional corepressor for
several transcription factors, and it acts synergistically with
SRC-2 as an important coactivator of Nur77-dependent transcription
(48). There is extensive evidence
indicating that Nur77 regulates the expression of several important
genes through interaction with specificity protein 1 (Sp1) or
specificity protein 4 (Sp4) bound to GC-rich promoters; for
example, a Nur77-Sp1-p300 DNA binding complex forms on the GC-rich
region close to the survivin promoter to inhibit the expression of
survivin, inducing an anticancer effect (49); Nur77 also regulates the expression
of β1- and β3-integrin via a Nur77/p300/Sp1 complex (50). Integrin gene promoters are GC-rich,
and α6-, α5-, and β4-integrins are regulated by
Nur77/Sp1/Sp3/Sp4/p300 complexes (51). Binding of the Nur77/Sp4 complex to
a GC-rich promoter can enhance the transcription of paired box 3
(PAX3)-forkhead box O1 (FOXO1A); furthermore, the expression of
Nur77 regulates β1-integrin which, together with PAX3-FOXO1A,
contributes to the migration of tumor cells (52). These effects may be exploited for
the development of novel antitumor drugs.
There are several downstream genes directly
regulated by Nur77, of which a number are involved in the endocrine
system, glucose metabolism, the cell cycle, apoptosis, inflammation
and other physiological processes. In skeletal muscle, Nur77
directly upregulates the gene expression of glucose transporting
protein 4 and promotes the absorption and utilization of glucose by
skeletal muscles (53,54). In addition, as a transcription
factor, Nur77 typically acts by binding to the NBRE in the promoter
regions of target genes. In the liver, Nur77 enhances
gluconeogenesis by modulating the expression of genes associated
with gluconeogenesis, including glucose-6-phosphatase, fructose
bisphosphatase 1/2 and enolase 3 (55–57).
SerpinA3 is a member of the serine protease inhibitor family and is
mainly synthesized in the liver. It is involved in inflammatory
responses and interacts with the Alzheimer's neurotoxic amyloid
peptide Aβ (58). It has been
demonstrated that Nur77 regulates SerpinA3 through an NBRE in its
promoter region, which may be associated with the role of Nur77 in
inflammatory diseases (58).
Notably, it has been reported that decanoic acid, a dietary
medium-chain fatty acid, can inhibit androgen biosynthesis in
NCI-H295R cells, and can reverse endocrine and metabolic
aberrations in a rat model of polycystic ovary syndrome; this
inhibitory effect may be achieved by the reduced recruitment of
Nur77 to the hydroxy-δ-5-steroid dehydrogenase 3β- and steroid
δ-isomerase 2 (HSD3B2) promoter, which decreases transcription of
the HSD3B2 gene and protein expression (59).
Non-genomic functions of Nur77
With the development of science and technology,
increasing evidence indicates that Nur77 can also affect the
biological functions of other proteins by non-transcriptional
functions. In the mitochondria, the Nur77 LBD binds the
anti-apoptotic protein Bcl-2, causing a conformation change that
exposes the BH3 domain of Bcl-2; this converts Bcl-2 from an
anti-apoptotic protein to a pro-apoptotic protein (36,60).
Furthermore, studies have demonstrated that p38α MAPK modulates the
Nur77-Bcl-2 apoptotic pathway (61). Another report showed that Nur77
physically interacts with p53, which leads to a decrease in
transcriptional activity by inhibiting the acetylation of p53.
Nur77 also causes the dissociation of murine double minute 2 (MDM2)
from p53, which can prevent p53 from MDM2-induced degradation,
increasing the stability of p53 (62). Nur77 can also interact with E1A
binding protein p300 and protein arginine methyltransferase 1
(PRMT1). Nur77 suppresses the acetylation of transcription factors
induced by p300 (63) and reduces
PRMT1 methyltransferase activity. Additionally, PRMT1 increases the
DNA binding and transactivation activity of Nur77 in a
non-methyltransferase manner (64). Nur77 can bind and sequester liver
kinase B1 (LKB1; also known as STK11), which is vital in governing
energy homeostasis in the nucleus; sequestering of LKB1 suppresses
the phosphorylation of adenosine 5′-monophosphate-activated protein
kinase α (AMPKα) (65). Nur77 also
binds to the tuberous sclerosis protein 1 (TSC1)/TSC2 complex to
modulate the activity of mammalian target of rapamycin complex 1
(mTORC1), which promotes TSC2 degradation through a
proteasome/ubiquitination pathway (66). In addition, the Nur77-mediated
activation of mTORC1 is associated with angiotensin II-induced
cardiac hypertrophy (66).
Inhibiting the interaction between p38α and Nur77, and inhibiting
the p38α-mediated phosphorylation of Nur77 attenuates the
lipopolysaccharide-induced hyperinflammatory response (67). The LBD of Nur77 binds to Akt2, and
the phosphorylation of Akt2 can interfere with the migration of
Nur77 to the cytoplasm and localization of Nur77 to the
mitochondria, which can result in autophagy (40,68).
Compounds that modulate the regulation of
Nur77
In recent years, progress has been made in the
treatment of cancer. In addition to the use of chemotherapy to
treat cancer, investigations now also focus on developing targeted
drugs. Targeted therapy involves the design of a therapeutic drug
to interact with a specific target. When the drug enters the body,
it interacts specifically with the carcinogenic target to inhibit
the growth of tumor cells. Compared with chemotherapy and
radiotherapy, targeted therapy can be used to attack tumor cells
with reduced effects on healthy cells. This can reduce side
effects, thus achieving improved efficacy and improving patient
quality of life. There is a demand to identify targets that can be
exploited for cancer therapy, and previous studies have shown that
Nur77 may be such a target (69,70).
However, although there are several reports of drugs that regulate
Nur77, there remains limited literature to summarize. In the
following section, the drugs reported to be are effective in
regulating Nur77 are described.
Ethyl
3,5-dihydroxy-2-(1-oxooctyl)-benzeneacetate (cytosporone B;
Csn-B)
The octaketide Csn-B (Fig. 2A), which is isolated from the
endophytic fungus Dothiorella sp. HTF3 (71), is a natural agonist of nuclear
orphan receptor Nur77 with an EC50 of 0.278 nM (72). Csn-B specifically binds to the LBD
of Nur77 and enhances Nur77-dependent transactivational activity on
target gene transcription (72).
Furthermore, Csn-B can induce apoptosis by increasing the
expression of Nur77 and promoting the translocation of Nur77 to the
cytoplasm to slow xenograft tumor growth (72). Csn-B enhances gluconeogenesis in
the mouse liver via increased activation of Nur77. Therefore, Csn-B
may be useful in the development of therapeutic agents to treat
cancer and hypoglycemia (72). Liu
et al (73) synthesized a
series of Csn-B analogues. These Csn-B-derived Nur77 agonists are a
novel group of potentially potent and effective antitumor
agents.
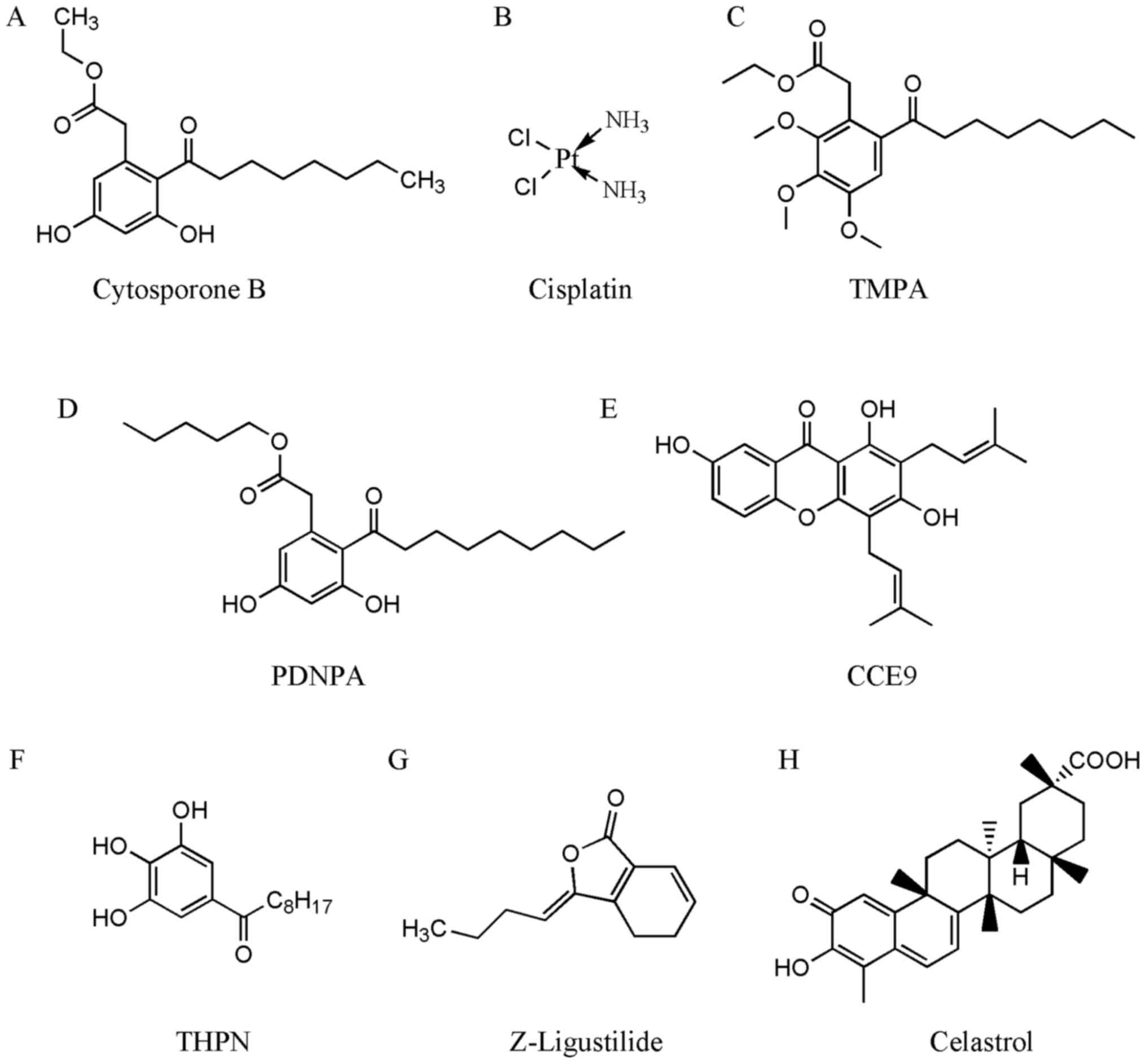 | Figure 2.Chemical structures of compounds that
interact with Nur77. Structures of (A) cytosporone B, (B)
cisplatin, (C) TMPA, (D) PDNPA, (E) CCE9, (F) THPN, (G)
Z-ligustilide, and (H) celastrol are shown. Cytosporone B, ethyl
3,5-dihydroxy-2-(1-oxooctyl)-benzeneacetate; cisplatin,
cis-diaminodichloroplatinum; TMPA, ethyl
2-[2,3,4-trimethoxy-6-(1-octanoyl)-phenyl] acetate; PDNPA, n-pentyl
2-[3,5-dihydroxy-2-(1-nonanoyl)-phenyl] acetate; CCE9,
1,3,7-trihydroxy-2,4-diprenylxanthone; THPN,
1-(3,4,5-trihydroxyphenyl)-nonan-1-one; Z-ligustilide,
3-butylidene-4,5-dihydro-1(3H)-isobenzofuranone; celastrol,
3-hydroxy-9β,13α-dimethyl-2-oxo-24,25,26-trinoroleana-1(10),3,5,7-tetraen-29-oic
acid. |
cis-diaminodichloroplatinum
(cisplatin)
Cisplatin (Fig. 2B)
is a chemotherapeutic used to treat various types of cancer.
Cisplatin effectively increases the phosphorylation of Nur77 by
activating checkpoint kinase 2 and facilitating cross-talk between
these two proteins, resulting in apoptosis and the inhibition of
intestinal tumorigenesis (74).
Additionally, xenografts that originate from the knockdown of Nur77
colon cancer cells are insensitive to cisplatin treatment.
Therefore, Nur77 is required for effective cisplatin therapy in
colon cancer (74).
Ethyl
2-[2,3,4-trimethoxy-6-(1-octanoyl)-phenyl] acetate (TMPA)
TMPA (Fig. 2C) is a
Csn-B derivative Nur77 agonist with high affinity. The interaction
of TMPA with Nur77 leads to the release and translocation of LKB1
to the cytoplasm, leading to the increased phosphorylation of AMPKα
(65). Furthermore, TMPA has been
shown to decrease blood glucose and attenuate insulin resistance in
type II db/db and high-fat diet mice, which was not observed in
Nur77 gene-knockout mice (65).
These findings indicate that TMPA may have use as a drug for the
treatment of metabolic disorders via its effect on Nur77 (65).
N-pentyl
2-[3,5-dihydroxy-2-(1-nonanoyl)-phenyl] acetate (PDNPA)
PDNPA (Fig. 2D)
targets the LBD of Nur77 and inhibits the interaction between Nur77
and p38α, which can eliminate the p38α-mediated phosphorylation of
Nur77 (67). The p38α-mediated
phosphorylation of Nur77 impedes the inhibition of nuclear
factor-κB (NF-κB) by Nur77. As reported by Li et al
(67), Nur77 directly binds NF-κB
p65 to inhibit its interactions with the κB response elements in
DNA, which inhibits the transcriptional activity of NF-κB.
Additionally, the reduced p38α-Nur77 interaction by PDNPA may
alleviate LPS-induced hyperinflammatory responses mediated by Nur77
(67). This finding suggests that
modulating p38α/Nur77-regulated functions may be a novel
therapeutic strategy to treat hyperinflammatory responses.
1,3,7-trihydroxy-2,4-diprenylxanthone
(CCE9)
CCE9 (Fig. 2E), a
xanthone compound, is isolated from the Chinese medicinal plant
Cratoxylum formosum ssp. pruniflorum (75). CCE9 positively regulates the
Nur77/Bcl-2-dependent apoptotic pathway. A previous report
suggested that CCE9 can promote the expression of Nur77 and
phosphorylation of Bcl-2 in a p38α MAPK-dependent manner, leading
to the cytoplasmic localization and mitochondrial targeting of
Nur77 and the Nur77/Bcl-2 interaction (61). The CCE9-induced induction of
apoptosis depends on the activation of p38α MAPK, followed by the
phosphorylation of Bcl-2 and binding of Bcl-2 with Nur77.
Additionally, interference of the expression of p38α MAPK inhibits
the interaction between Bcl-2 and Nur77 (61), and the phosphorylation of Bcl-2 by
p38α MAPK at Ser87 and Thr56 is required for the interaction with
Nur77 (61). This mechanism of the
Nur77-Bcl-2 apoptotic pathway may provide a novel strategy for the
development of diverse chemotherapeutic agents.
1-(3,4,5-trihydroxyphenyl)-nonan-1-one
(THPN)
THPN (Fig. 2F),
another Csn-B derivative and Nur77-specific ligand, induces
autophagy (76). In melanoma, THPN
stimulation was found to increase Nur77 cytoplasmic localization
and facilitate its binding with NIP3-like protein X (Nix), a
mitochondrial outer membrane protein. The THPN-induced Nix-Nur77
interaction may be the first step in mitochondrial depolarization
and autophagic cell death (76).
THPN-induced Nur77 transport to the mitochondrial inner membrane
may be another critical event in autophagic cell death.
Furthermore, THPN inhibits melanoma growth via autophagy, not via
the induction of apoptosis (76).
This suggests that Nur77 is involved in the regulation of autophagy
in melanoma, which may be exploited to develop anticancer
therapeutics.
3-Butylidene-4,5-dihydro-1(3H)-isobenzofuranone (Z-ligustilide;
Z-LIG)
Z-LIG (Fig. 2G) is
a phthalide compound and accounts for >50% of the volatile oil
in Radix Angelica Sinensis (77), which is used to treat gynecological
disorders in traditional Chinese medicine and has been reported to
have an inhibitory effect on tumors in various types of human
cancer, including colon cancer (78) and breast cancer (79). It is well established that
autophagy can have a dual role; it can either promote apoptosis or
have pro-survival effects, depending on the DNA damage present in
cancer cells. For example, autophagy delays apoptotic death in
tamoxifen-resistant MCF-7 (MCF-7TR5) breast cancer cells
(79,80). Notably, Z-LIG has been reported to
inhibit autophagic flux by inhibiting autophagosome-lysosome
fusion; therefore, Z-LIG may be a novel autophagy inhibitor that
can induce cell death of tamoxifen-resistant breast cancer cells.
Z-LIG has been shown to enhance the efficacy of tamoxifen therapy
(79); following Nur77 knockdown,
the ability of Z-LIG to sensitize tamoxifen-induced cell death was
reduced (79). This indicates that
Nur77 may be critical in Z-LIG-mediated DNA damage and in the
restoration tamoxifen sensitivity in MCF-7TR5 cells
(79). Together these findings
indicate that the effects of Z-LIG and Nur77 may be useful for
obtaining novel insights into the mechanisms that mediate tamoxifen
resistance in breast cancer.
3-hydroxy-9β,13α-dimethyl-2-oxo-24,25,26-trinoroleana-1(10),3,5,7-tetraen-29-oic
acid (celastrol)
Celastrol (Fig. 2H)
is one of the traditional medicinal compounds that may have the
potential to be developed as a modern therapeutic agent. It has
been reported that celastrol has potent anti-inflammatory
activities within several inflammatory diseases and in obesity
(81,82); however, the effect of celastrol on
inflammatory mechanisms remains to be fully elucidated. It has been
reported that celastrol binds Nur77 to inhibit inflammation and
induce autophagy in a Nur77-dependent manner. Celastrol induces
translocation of Nur77 from the nucleus to the mitochondria, where
it is ubiquitinated by tumor necrosis factor receptor-associated
factor 2, a scaffold protein, and E3 ubiquitin ligase; these are
important for inflammatory signaling (83). The ubiquitinated Nur77 binds to
p62/sequestosome 1, which results in sensitivity to autophagy under
inflammatory conditions.
As inflammatory mitochondrial dysfunction causes
various diseases and types of cancer, these pathways may provide
directions for the investigation of cancer and development of novel
therapies. Celastrol is a promising drug within this class of
therapeutics that acts by targeting Nur77 (83).
Bisindole methane (DIM) compounds
Indole compounds are bioactive components present in
cruciferous plants. In relatively low pH environments, indoles can
be converted into various polymers, of which DIM is the most
important class. Increasing studies have reported that DIM and its
derivatives can specifically bind to and activate multiple nuclear
receptors, including Nur77 and Nurr1, to regulate signaling
pathways. 1,1-Bis (3-indolyl)-1-(p-substituted phenyl) methane
(C-DIM) compounds have significant antitumor activity and low
toxicity in cancer cells. Treating athymic nude bearing
A549-derived metastases with two C-DIM analogs, DIM-C-pPhOCH3
(C-DIM-5) and DIM-C-pPhOH (C-DIM-8), resulted in significant
regression of lung tumors. Furthermore, the effects of the two
analogues were different; C-DIM-5 inhibited the cell cycle from the
G0/G1 phase to the S phase, and C-DIM-8 induced A549 cell apoptosis
(84). Additionally, C-DIM-5 and
C-DIM-8 have been identified as prototypical activators and
antagonists of Nur77, respectively (49,85).
C-DIM-5 has been used as a prototype activator for Nur77 in a
transactivation assay using the GAL4-Nur77/GAL4-reactive element
reporter assay system. However, subsequent investigations with
human GAL4-Nur77 showed minimal transactivation of C-DIM-5
(84,86). C-DIM-8 inhibits the activation of
Nur77 in pancreatic cancer and lung cancer cells, resulting in the
inhibition of cell growth and induction of apoptosis, with results
similar to those observed following RNA interference-induced Nur77
(49,87). In addition, C-DIM-5 inhibits the
growth and induces apoptosis of UC-5 and KU7 bladder cancer cells
(88). The high-affinity
interaction between C-DIM-8 and the analogous compounds in the
ligand-binding pocket of Nur77 reduces the growth of colon cancer
cells, induces cell apoptosis and decreases the expression of
survivin and other Sp1-regulated genes (87). Furthermore, treatment of
pancreatic, colon and breast cancer cells with C-DIM-8 or
DIM-C-pPhCO2Me resulted in an effect similar to the knockdown of
Nur77, with reduced the expression of β1-integrin, migration and
adhesion. A high expression of β1-integrin is a poor prognostic
factor for patients with colon and pancreatic cancer, and this
protein is important in cell migration and invasion (50,89,90).
Additionally, treatment of rhabdomyosarcoma cells, and of ACHN and
786-O renal cell lines with C-DIM-8 and DIM-C-pPhCO2Me was shown to
inhibit tumor cell growth and induce apoptosis (91,92).
In summary, diindolylmethane analogues bind Nur77 and are Nur77
antagonists in various types of cancer cells, which may provide
novel avenues for anticancer treatment.
Conclusions and perspectives
Nur77 is a relatively well-researched orphan nuclear
receptor, and the majority of current studies focus on identifying
ligands that regulate the function of Nur77 under pathological
conditions. The literature suggests that the orphan nuclear
receptor Nur77 is important in cancer, including in colorectal
cancer (93,94), androgen-induced bladder cancer
(95) and lung cancer (96). In general, although the endogenous
ligand for the Nur77 receptor has not been identified, studies have
identified structurally diverse compounds that bind to and activate
or inactivate Nur77, induce the transfer of Nur77 from the nucleus
to the cytoplasm, and can regulate associated diseases. These
compounds include Csn-B, cisplatin, celastrol, Z-LIG and DIM
compounds. Cisplatin is already an antitumor chemotherapeutic drug.
Cytosporone B is currently being assessed for anticancer activates;
however, its detailed biological effects and potential side effects
require investigation. In addition, whether there are other factors
that influence the expression and location of Nur77, the mechanism
of action of Nur77 as a potential regulator of inflammation, and
the specific association between Nur77 and apoptosis and autophagy
remain to be fully elucidated. Furthermore, the continued
investigation of compounds that can modulate Nur77 is required,
followed by the development and performing of clinical trials using
these compounds to evaluate their potential for therapeutic use. In
conclusion, the orphan nuclear receptor Nur77 is a promising
research target for investigating cancer mechanisms and prospective
therapies. It is important to understand the structure, function,
expression and localization of Nur77, and to screen and develop
compounds that act as agonists or antagonists and may have clinical
applications.
Acknowledgements
Not applicable.
Funding
This study was supported by grants from the National
Natural Science Foundation (grant no. 31500616) and the Natural
Science Foundation of Fujian Province (grant no. 2017J01445).
Availability of data and materials
Not applicable.
Authors' contributions
LW and LC designed and supervised the project and
wrote the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mangelsdorf DJ, Thummel C, Beato M,
Herrlich P, Schütz G, Umesono K, Blumberg B, Kastner P, Mark M,
Chambon P and Evans RM: The nuclear receptor superfamily: The
second decade. Cell. 83:835–839. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Margolis RN and Christakos S: The nuclear
receptor superfamily of steroid hormones and vitamin D gene
regulation. An update. Ann N Y Acad Sci. 1192:208–214. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Robinson-Rechavi M, Garcia Escriva H and
Laudet V: The nuclear receptor superfamily. J Cell Sci.
116:585–586. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Giguere V: Structure and function of the
nuclear receptor superfamily for steroid, thyroid hormone and
retinoic acid. Genet Eng (N Y). 12:183–200. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Brelivet Y, Rochel N and Moras D:
Structural analysis of nuclear receptors: From isolated domains to
integral proteins. Mol Cell Endocrinol. 348:466–473. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
McEwan IJ: The Nuclear receptor
superfamily at thirty. Methods Mol Biol. 1443:3–9. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Germain P, Staels B, Dacquet C, Spedding M
and Laudet V: Overview of nomenclature of nuclear receptors.
Pharmacol Rev. 58:685–704. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rastinejad F, Huang P, Chandra V and
Khorasanizadeh S: Understanding nuclear receptor form and function
using structural biology. J Mol Endocrinol. 51:T1–T21. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chandra V, Huang P, Hamuro Y, Raghuram S,
Wang Y, Burris TP and Rastinejad F: Structure of the intact
PPAR-gamma-RXR-nuclear receptor complex on DNA. Nature.
456:350–356. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Olefsky JM: Nuclear receptor minireview
series. J Biol Chem. 276:36863–36864. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen T: Nuclear receptor drug discovery.
Curr Opin Chem Biol. 12:418–426. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen T: Overcoming drug resistance by
regulating nuclear receptors. Adv Drug Deliv Rev. 62:1257–1264.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hazel TG, Nathans D and Lau LF: A gene
inducible by serum growth factors encodes a member of the steroid
and thyroid hormone receptor superfamily. Proc Natl Acad Sci USA.
85:8444–8448. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Watson MA and Milbrandt J: The NGFI-B
gene, a transcriptionally inducible member of the steroid receptor
gene superfamily: Genomic structure and expression in rat brain
after seizure induction. Mol Cell Biol. 9:4213–4219. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chang C, Kokontis J, Liao SS and Chang Y:
Isolation and characterization of human TR3 receptor: A member of
steroid receptor superfamily. J Steroid Biochem. 34:391–395. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hwang DS, Lee BY, Kim HS, Lee MC, Kyung
DH, Om AS, Rhee JS and Lee JS: Genome-wide identification of
nuclear receptor (NR) superfamily genes in the copepod Tigriopus
japonicus. BMC Genomics. 15:9932014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sharma Y, Chilamakuri CS, Bakke M and
Lenhard B: Computational characterization of modes of
transcriptional regulation of nuclear receptor genes. PLoS One.
9:e888802014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kurakula K, Koenis DS, van Tiel CM and de
Vries CJ: NR4A nuclear receptors are orphans but not lonesome.
Biochim Biophys Acta. 1843:2543–2555. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rehman SU, Sarwar T, Husain MA, Ishqi HM
and Tabish M: Identification of two novel isoforms of mouse NUR77
lacking N-terminal domains. IUBMB Life. 69:106–114. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wilson TE, Fahrner TJ and Milbrandt J: The
orphan receptors NGFI-B and steroidogenic factor 1 establish
monomer binding as a third paradigm of nuclear receptor-DNA
interaction. Mol Cell Biol. 13:5794–5804. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Maira M, Martens C, Philips A and Drouin
J: Heterodimerization between members of the Nur subfamily of
orphan nuclear receptors as a novel mechanism for gene activation.
Mol Cell Biol. 19:7549–7557. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Forman BM, Goode E, Chen J, Oro AE,
Bradley DJ, Perlmann T, Noonan DJ, Burka LT, McMorris T, Lamph WW,
et al: Identification of a nuclear receptor that is activated by
farnesol metabolites. Cell. 81:687–693. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Perlmann T and Jansson L: A novel pathway
for vitamin A signaling mediated by RXR heterodimerization with
NGFI-B and NURR1. Genes Dev. 9:769–782. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Flaig R, Greschik H, Peluso-Iltis C and
Moras D: Structural basis for the cell-specific activities of the
NGFI-B and the Nurr1 ligand-binding domain. J Biol Chem.
280:19250–19258. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Michiels P, Atkins K, Ludwig C, Whittaker
S, van Dongen M and Günther U: Assignment of the orphan nuclear
receptor Nurr1 by NMR. Biomol NMR Assign. 4:101–105. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wansa KD, Harris JM and Muscat GE: The
activation function-1 domain of Nur77/NR4A1 mediates
trans-activation, cell specificity, and coactivator recruitment. J
Biol Chem. 277:33001–33011. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lanig H, Reisen F, Whitley D, Schneider G,
Banting L and Clark T: In silico adoption of an orphan nuclear
receptor NR4A1. PLoS One. 10:e01352462015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Moore TW, Mayne CG and Katzenellenbogen
JA: Minireview: Not picking pockets: Nuclear receptor
alternate-site modulators (NRAMs). Mol Endocrinol. 24:683–695.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chang LF, Lin PC, Ho LI, Liu PY, Wu WC,
Chiang IP, Chang HW, Lin SZ, Harn YC, Harn HJ and Chiou TW:
Overexpression of the orphan receptor Nur77 and its translocation
induced by PCH4 may inhibit malignant glioma cell growth and induce
cell apoptosis. J Surg Oncol. 103:442–450. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Holmes WF, Soprano DR and Soprano KJ:
Early events in the induction of apoptosis in ovarian carcinoma
cells by CD437: Activation of the p38 MAP kinase signal pathway.
Oncogene. 22:6377–6386. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Niu G, Lu L, Gan J, Zhang D, Liu J and
Huang G: Dual roles of orphan nuclear receptor TR3/Nur77/NGFI-B in
mediating cell survival and apoptosis. Int Rev Cell Mol Biol.
313:219–258. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou F, Drabsch Y, Dekker TJ, de Vinuesa
AG, Li Y, Hawinkels LJ, Sheppard KA, Goumans MJ, Luwor RB, de Vries
CJ, et al: Nuclear receptor NR4A1 promotes breast cancer invasion
and metastasis by activating TGF-β signalling. Nat Commun.
5:33882014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hedrick E and Safe S: Transforming growth
factor β/NR4A1-inducible breast cancer cell migration and
epithelial-to-mesenchymal transition is p38α (Mitogen-Activated
Protein Kinase 14) dependent. Mol Cell Biol. 37:pii: e00306. –17.
2017. View Article : Google Scholar
|
|
34
|
Delgado E, Boisen MM, Laskey R, Chen R,
Song C, Sallit J, Yochum ZA, Andersen CL, Sikora MJ, Wagner J, et
al: High expression of orphan nuclear receptor NR4A1 in a subset of
ovarian tumors with worse outcome. Gynecol Oncol. 141:348–356.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang X, Yan G, Diao Z, Sun H and Hu Y:
NUR77 inhibits the expression of TIMP2 and increases the migration
and invasion of HTR-8/SVneo cells induced by CYR61. Placenta.
33:561–567. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lin B, Kolluri SK, Lin F, Liu W, Han YH,
Cao X, Dawson MI, Reed JC and Zhang XK: Conversion of Bcl-2 from
protector to killer by interaction with nuclear orphan receptor
Nur77/TR3. Cell. 116:527–540. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cao X, Liu W, Lin F, Li H, Kolluri SK, Lin
B, Han YH, Dawson MI and Zhang XK: Retinoid X receptor regulates
Nur77/TR3-dependent apoptosis [corrected] by modulating its nuclear
export and mitochondrial targeting. Mol Cell Biol. 24:9705–9725.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang XK: Targeting Nur77 translocation.
Expert Opin Ther Targets. 11:69–79. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Han YH, Cao X, Lin B, Lin F, Kolluri SK,
Stebbins J, Reed JC, Dawson MI and Zhang XK: Regulation of Nur77
nuclear export by c-Jun N-terminal kinase and Akt. Oncogene.
25:2974–2986. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen HZ, Zhao BX, Zhao WX, Li L, Zhang B
and Wu Q: Akt phosphorylates the TR3 orphan receptor and blocks its
targeting to the mitochondria. Carcinogenesis. 29:2078–2088. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
No H, Bang Y, Lim J, Kim SS, Choi HS and
Choi HJ: Involvement of induction and mitochondrial targeting of
orphan nuclear receptor Nur77 in 6-OHDA-induced SH-SY5Y cell death.
Neurochem Int. 56:620–626. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Debernard Boldingh KA, Mathisen GH and
Paulsen RE: Differences in NGFI-B, Nurr1, and NOR-1 expression and
nucleocytoplasmic translocation in glutamate-treated neurons.
Neurochem Int. 61:79–88. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Renaud J, Chiasson K, Bournival J,
Rouillard C and Martinoli MG: 17β-estradiol delays 6-OHDA-induced
apoptosis by acting on Nur77 translocation from the nucleus to the
cytoplasm. Neurotox Res. 25:124–134. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Agostini-Dreyer A, Jetzt AE, Stires H and
Cohick WS: Endogenous IGFBP-3 mediates intrinsic apoptosis through
modulation of Nur77 phosphorylation and nuclear export.
Endocrinology. 156:4141–4151. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hu Y, Chau T, Liu HX, Liao D, Keane R, Nie
Y, Yang H and Wan YJ: Bile acids regulate nuclear receptor (Nur77)
expression and intracellular location to control proliferation and
apoptosis. Mol Cancer Res. 13:281–292. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Drouin J, Maira M and Philips A: Novel
mechanism of action for Nur77 and antagonism by glucocorticoids: A
convergent mechanism for CRH activation and glucocorticoid
repression of POMC gene transcription. J Steroid Biochem Mol Biol.
65:59–63. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Maira M, Martens C, Batsché E, Gauthier Y
and Drouin J: Dimer-specific potentiation of NGFI-B (Nur77)
transcriptional activity by the protein kinase A pathway and
AF-1-dependent coactivator recruitment. Mol Cell Biol. 23:763–776.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Rambaud J, Desroches J, Balsalobre A and
Drouin J: TIF1beta/KAP-1 is a coactivator of the orphan nuclear
receptor NGFI-B/Nur77. J Biol Chem. 284:14147–14156. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Lee SO, Abdelrahim M, Yoon K,
Chintharlapalli S, Papineni S, Kim K, Wang H and Safe S:
Inactivation of the orphan nuclear receptor TR3/Nur77 inhibits
pancreatic cancer cell and tumor growth. Cancer Res. 70:6824–6836.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hedrick E, Lee SO, Doddapaneni R, Singh M
and Safe S: NR4A1 antagonists inhibit β1-integrin-dependent breast
cancer cell migration. Mol Cell Biol. 36:1383–1394. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hedrick E, Li X and Safe S: Penfluridol
represses integrin expression in breast cancer through induction of
reactive oxygen species and downregulation of Sp transcription
factors. Mol Cancer Ther. 16:205–216. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Lacey A, Rodrigues-Hoffman A and Safe S:
PAX3-FOXO1A expression in rhabdomyosarcoma is driven by the
targetable nuclear receptor NR4A1. Cancer Res. 77:732–741. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Chao LC, Zhang Z, Pei L, Saito T, Tontonoz
P and Pilch PF: Nur77 coordinately regulates expression of genes
linked to glucose metabolism in skeletal muscle. Mol Endocrinol.
21:2152–2163. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kanzleiter T, Preston E, Wilks D, Ho B,
Benrick A, Reznick J, Heilbronn LK, Turner N and Cooney GJ:
Overexpression of the orphan receptor Nur77 alters glucose
metabolism in rat muscle cells and rat muscle in vivo.
Diabetologia. 53:1174–1183. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Pei L, Waki H, Vaitheesvaran B, Wilpitz
DC, Kurland IJ and Tontonoz P: NR4A orphan nuclear receptors are
transcriptional regulators of hepatic glucose metabolism. Nat Med.
12:1048–1055. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
56
|
Pols TW, Ottenhoff R, Vos M, Levels JH,
Quax PH, Meijers JC, Pannekoek H, Groen AK and de Vries CJ: Nur77
modulates hepatic lipid metabolism through suppression of SREBP1c
activity. Biochem Biophys Res Commun. 366:910–916. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhao Y and Bruemmer D: NR4A orphan nuclear
receptors: Transcriptional regulators of gene expression in
metabolism and vascular biology. Arterioscler Thromb Vasc Biol.
30:1535–1541. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhao Y, Liu Y and Zheng D: Alpha
1-antichymotrypsin/SerpinA3 is a novel target of orphan nuclear
receptor Nur77. FEBS J. 275:1025–1038. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lee BH, Indran IR, Tan HM, Li Y, Zhang Z,
Li J and Yong EL: A dietary medium-chain fatty acid, decanoic acid,
inhibits recruitment of Nur77 to the HSD3B2 promoter in vitro and
reverses endocrine and metabolic abnormalities in a rat model of
polycystic ovary syndrome. Endocrinology. 157:382–394. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Kolluri SK, Zhu X, Zhou X, Lin B, Chen Y,
Sun K, Tian X, Town J, Cao X, Lin F, et al: A short Nur77-derived
peptide converts Bcl-2 from a protector to a killer. Cancer Cell.
14:285–298. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Liu J, Wang GH, Duan YH, Dai Y, Bao Y, Hu
M, Zhou YQ, Li M, Jiang F, Zhou H, et al: Modulation of the
Nur77-Bcl-2 apoptotic pathway by p38alpha MAPK. Oncotarget.
8:69731–69745. 2017.PubMed/NCBI
|
|
62
|
Zhao BX, Chen HZ, Lei NZ, Li GD, Zhao WX,
Zhan YY, Liu B, Lin SC and Wu Q: p53 mediates the negative
regulation of MDM2 by orphan receptor TR3. EMBO J. 25:5703–5715.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Li GD, Fang JX, Chen HZ, Luo J, Zheng ZH,
Shen YM and Wu Q: Negative regulation of transcription coactivator
p300 by orphan receptor TR3. Nucleic Acids Res. 35:7348–7359. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Lei NZ, Zhang XY, Chen HZ, Wang Y, Zhan
YY, Zheng ZH, Shen YM and Wu Q: A feedback regulatory loop between
methyltransferase PRMT1 and orphan receptor TR3. Nucleic Acids Res.
37:832–848. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Zhan YY, Chen Y, Zhang Q, Zhuang JJ, Tian
M, Chen HZ, Zhang LR, Zhang HK, He JP, Wang WJ, et al: The orphan
nuclear receptor Nur77 regulates LKB1 localization and activates
AMPK. Nat Chem Biol. 8:897–904. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Wang RH, He JP, Su ML, Luo J, Xu M, Du XD,
Chen HZ, Wang WJ, Wang Y, Zhang N, et al: The orphan receptor TR3
participates in angiotensin II-induced cardiac hypertrophy by
controlling mTOR signalling. EMBO Mol Med. 5:137–148. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Li L, Liu Y, Chen HZ, Li FW, Wu JF, Zhang
HK, He JP, Xing YZ, Chen Y, Wang WJ, et al: Impeding the
interaction between Nur77 and p38 reduces LPS-induced inflammation.
Nat Chem Biol. 11:339–346. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Wang WJ, Wang Y, Hou PP, Li FW, Zhou B,
Chen HZ, Bian XL, Cai QX, Xing YZ, He JP, et al: Induction of
autophagic death in cancer cells by agonizing TR3 and attenuating
Akt2 activity. Chem Biol. 22:1040–1051. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
To SK, Zeng JZ and Wong AS: Nur77: A
potential therapeutic target in cancer. Expert Opin Ther Targets.
16:573–585. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Zeng Y, Ye X, Liao D, Huang S, Mao H, Zhao
D and Zeng H: Orphan nuclear receptor TR3/Nur77 is a specific
therapeutic target for hepatic cancers. J Clin Exp Oncol. 6:pii:
184. 2017. View Article : Google Scholar
|
|
71
|
Brady SF, Wagenaar MM, Singh MP, Janso JE
and Clardy J: The cytosporones, new octaketide antibiotics isolated
from an endophytic fungus. Org Lett. 2:4043–4046. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhan Y, Du X, Chen H, Liu J, Zhao B, Huang
D, Li G, Xu Q, Zhang M, Weimer BC, et al: Cytosporone B is an
agonist for nuclear orphan receptor Nur77. Nat Chem Biol.
4:548–556. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Liu JJ, Zeng HN, Zhang LR, Zhan YY, Chen
Y, Wang Y, Wang J, Xiang SH, Liu WJ, Wang WJ, et al: A unique
pharmacophore for activation of the nuclear orphan receptor Nur77
in vivo and in vitro. Cancer Res. 70:3628–3637. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Yao LM, He JP, Chen HZ, Wang Y, Wang WJ,
Wu R, Yu CD and Wu Q: Orphan receptor TR3 participates in
cisplatin-induced apoptosis via Chk2 phosphorylation to repress
intestinal tumorigenesis. Carcinogenesis. 33:301–311. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Duan YH, Dai Y, Wang GH, Zhang X, Chen HF,
Chen JB, Yao XS and Zhang XK: Bioactive xanthones from the stems of
Cratoxylum formosum ssp. pruniflorum. J Nat Prod. 73:1283–1287.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Wang WJ, Wang Y, Chen HZ, Xing YZ, Li FW,
Zhang Q, Zhou B, Zhang HK, Zhang J, Bian XL, et al: Orphan nuclear
receptor TR3 acts in autophagic cell death via mitochondrial
signaling pathway. Nat Chem Biol. 10:133–140. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Zeng Q, Jia YW, Xu PL, Xiao MW, Liu YM,
Peng SL and Liao X: Quick and selective extraction of Z-ligustilide
from Angelica sinensis using magnetic multiwalled carbon
nanotubes. J Sep Sci. 38:4269–4275. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Kan WL, Cho CH, Rudd JA and Lin G: Study
of the anti-proliferative effects and synergy of phthalides from
Angelica sinensis on colon cancer cells. J Ethnopharmacol.
120:36–43. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Qi H, Jiang Z, Wang C, Yang Y, Li L, He H
and Yu Z: Sensitization of tamoxifen-resistant breast cancer cells
by Z-ligustilide through inhibiting autophagy and accumulating DNA
damages. Oncotarget. 8:29300–29317. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Abedin MJ, Wang D, McDonnell MA, Lehmann U
and Kelekar A: Autophagy delays apoptotic death in breast cancer
cells following DNA damage. Cell Death Differ. 14:500–510. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Greenhill C: Celastrol identified as a
leptin sensitizer and potential novel treatment for obesity. Nat
Rev Endocrinol. 11:4442015. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Liu J, Lee J, Hernandez Salazar MA,
Mazitschek R and Ozcan U: Treatment of obesity with celastrol.
Cell. 161:999–1011. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Hu M, Luo Q, Alitongbieke G, Chong S, Xu
C, Xie L, Chen X, Zhang D, Zhou Y, Wang Z, et al: Celastrol-induced
Nur77 interaction with TRAF2 alleviates inflammation by promoting
mitochondrial ubiquitination and autophagy. Mol Cell. 66:141–153,
e6. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Andey T, Patel A, Jackson T, Safe S and
Singh M: 1,1-Bis (3′-indolyl)-1-(p-substitutedphenyl)methane
compounds inhibit lung cancer cell and tumor growth in a metastasis
model. Eur J Pharm Sci. 500:227–241. 2013. View Article : Google Scholar
|
|
85
|
Yoon K, Lee SO, Cho SD, Kim K, Khan S and
Safe S: Activation of nuclear TR3 (NR4A1) by a diindolylmethane
analog induces apoptosis and proapoptotic genes in pancreatic
cancer cells and tumors. Carcinogenesis. 32:836–842. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Cho SD, Yoon K, Chintharlapalli S,
Abdelrahim M, Lei P, Hamilton S, Khan S, Ramaiah SK and Safe S:
Nur77 agonists induce proapoptotic genes and responses in colon
cancer cells through nuclear receptor-dependent and nuclear
receptor-independent pathways. Cancer Res. 67:674–683. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Lee SO, Li X, Hedrick E, Jin UH, Tjalkens
RB, Backos DS, Li L, Zhang Y, Wu Q and Safe S: Diindolylmethane
analogs bind NR4A1 and are NR4A1 antagonists in colon cancer cells.
Mol Endocrinol. 28:1729–1739. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Cho SD, Lee SO, Chintharlapalli S,
Abdelrahim M, Khan S, Yoon K, Kamat AM and Safe S: Activation of
nerve growth factor-induced B alpha by methylene-substituted
diindolylmethanes in bladder cancer cells induces apoptosis and
inhibits tumor growth. Mol Pharmacol. 77:396–404. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Hedrick E, Lee SO and Safe S: The nuclear
orphan receptor NR4A1 regulates β1-integrin expression in
pancreatic and colon cancer cells and can be targeted by NR4A1
antagonists. Mol Carcinog. 56:2066–2075. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Hedrick E, Lee SO, Doddapaneni R, Singh M
and Safe S: Nuclear receptor 4A1 as a drug target for breast cancer
chemotherapy. Endocr Relat Cancer. 22:831–840. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Lacey A, Hedrick E, Li X, Patel K,
Doddapaneni R, Singh M and Safe S: Nuclear receptor 4A1 (NR4A1) as
a drug target for treating rhabdomyosarcoma (RMS). Oncotarget.
7:31257–31269. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Hedrick E, Lee SO, Kim G, Abdelrahim M,
Jin UH, Safe S and Abudayyeh A: Nuclear receptor 4A1 (NR4A1) as a
drug target for renal cell adenocarcinoma. PLoS One.
10:e01283082015. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Wang JR, Gan WJ, Li XM, Zhao YY, Li Y, Lu
XX, Li JM and Wu H: Orphan nuclear receptor Nur77 promotes
colorectal cancer invasion and metastasis by regulating MMP-9 and
E-cadherin. Carcinogenesis. 35:2474–2484. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
To SK, Zeng WJ, Zeng JZ and Wong AS:
Hypoxia triggers a Nur77-β-catenin feed-forward loop to promote the
invasive growth of colon cancer cells. Br J Cancer. 110:935–945.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Wu J, Liu J, Jia R and Song H: Nur77
inhibits androgen-induced bladder cancer growth. Cancer Invest.
31:654–660. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Wohlkoenig C, Leithner K, Olschewski A,
Olschewski H and Hrzenjak A: TR3 is involved in hypoxia-induced
apoptosis resistance in lung cancer cells downstream of HIF-1α.
Lung Cancer. 111:15–22. 2017. View Article : Google Scholar : PubMed/NCBI
|