Introduction
Previous studies indicated that epidermal growth
factor receptor (EGFR) as a cell membrane growth factor receptor
has a crucial role in controlling the key cellular transduction
pathways in cancerous cells (1,2). The
AKT serine/threonine kinase 1 (AKT)/glycogen synthase kinase 3β
(GSK3β) and the extracellular signal-regulated kinase
(ERK)/lysine-tRNA ligase MSK1 (MSK1) signaling pathways are the two
important downstream signaling pathways of EGFR, which have
essential roles in tumor development. Previous studies confirmed
this report (3–5). Previous studies indicated that ERK,
MSKs, AKT and GSK3β are all critical signal transducers of
oncogenic signals in various human tumors (6–8). For
example, AKT may be activated through phosphorylation in Ser473 and
Thr308 and active AKT may subsequently phosphorylate GSK3β (Ser9)
(9). Previous studies revealed
that selective GSK3 inhibitors and GSK3β small interfering RNA
attenuated tetrandrine-induced G1 type GSK3β that may lead to G1
arrest and apoptosis via cyclin D1 downregulation (10,11).
Overexpression of cyclin D1 may promote the development of
malignant tumors, such as parathyroid adenoma, breast cancer, colon
cancer, lymphoma, melanoma and prostate cancer (12–16).
Therefore, the present study investigated whether a natural
compound may alter the EGF/ERK/MSK1 and EGF/AKT/GSK3β signaling
pathways to prevent EGF-induced JB6 C141 transformation and
proliferation.
Aloe emodin is one of the primary components in Rhei
Rhizoma. It may be found in aloe and the roots of Rheum, with a
molecular formula is C15H10O5
(11). Previous studies revealed
that aloe emodin has anti-tumor, anti-bacterial, cell-restoring and
laxative function (17). A
previous study reported that aloe emodin may selectively inhibit
human neuroectodermal tumor cell growth and had little acute or
chronic toxicity in animal model (18). It inhibited proliferation and
anchorage-independent growth of PC3 cells (8). The present study determined that aloe
emodin was able to inhibit proliferation and transformation of the
EGF-induced JB6 C141 cells. Protein content analysis and
vitro kinase western blot analysis revealed that aloe-emodin
inhibited the ERK/MSK1 and AKT/GSK3β signaling pathways,
specifically expression levels of MSK1 and phosphorylation-AKT
(Ser473) were markedly inhibited. Phosphorylation of cyclin D1
significantly decreased and large proportion of JB6 C141 cells was
arrested at the G1/S phase. Therefore, these findings demonstrated
that the EGF/ERK/MSK1 and EGF/AKT/GSK3β signaling pathways may
contribute to the suppression proliferation and transformation of
EGF-induced JB6 C141 cells, whereas aloe emodin suppressed
EGF-induced neoplastic cell transformation and proliferation.
Therefore, aloe emodin may be a useful chemoprevention drug for
cancer treatment.
Materials and methods
Materials
Aloe emodin with purity >95% and other chemical
reagents, including Tris, NaCl and SDS, were purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany) for molecular
biology and buffer preparation. Antibodies for western blot
analysis were purchased from Cell Signaling Technology, Inc.
(Beverly, MA, USA), Santa Cruz Biotechnology Inc. (Dallas, TX, USA)
or Upstate Biotechnology (EMD Millipore, Billerica, MA, USA).
Cell culture
JB6 C141 cells were purchased from American Type
Culture Collection (ATCC; Manassas, VA, USA). JB6 C141 cells were
propagated in F-12K medium (Cellgro; Corning, Inc., Corning, NY,
USA), containing 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin
(Cellgro; Corning, Inc.), in a 37°C humidified incubator with 5%
CO2. JB6 C141 cells were cytogenetically tested and
authenticated prior being frozen. Enough frozen vials were
available to ensure that all cell-based experiments could be
conducted. Every vial of frozen cells was thawed and maintained in
culturing for a maximum of 8 weeks.
EGF or TPA-induced cell
transformation
Cells (8×103/ml) were exposed to EGF
(0.1–10 ng/ml) or TPA (2–20 ng/ml) in 1 ml of 0.33% basal medium
(Sigma-Aldrich; Merck KGaA) containing 10% FBS. The cultures were
maintained in a 37°C, 5% CO2 incubator for 10 days (EGF)
or 3–4 weeks (TPA), and the cell colonies were scored using a
fluorescence microscope (Olympus, Tokyo, Japan) and the Image-Pro
PLUS version 4.5 computer software program (Media Cybernetics,
Rockville, MD, USA).
Toxicity of aloe emodin
The cytotoxic activity of aloe emodin was detected
following exponential cell growth in cell culture medium, which
contains 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin
(Cellgro; Corning, Inc.) over 48 h under the conditions listed
above. The cells were seeded in 96-well plates for 12 h prior
treatment. Monolayer cells were plated with a density of
1×104 cells/well. Aloe emodin (purity >95%) was added
to the final experimental concentration and cells were counted
after 48 h using the trypan blue exclusion assay. All of the
experiments were conducted in triplicate.
MTS assay
JB6 C141 cells (1×103 cells/well) were
seeded into 96-well plates in 100 µl F-12K medium supplemented with
10% FBS and incubated in a 5% CO2 incubator with 37°C.
Following culturing for 12 h with 2.5, 5, 10, 15 µM aloe emodin
were added to every well. After incubation with 37°C for an
additional 24, 48, 72 or 96 h, 20 µl CellTiter96 Aqueous One
solution (Promega, Madison, WI, USA) was added to every well. Then
cells were cultivated for another 2 h at 37°C. Absorbance was
quantified at 490 and 690 nm.
Western blotting
For western blot analysis, cells (2×106)
were cultured in a 10 cm dish for 24 h. The cells were then treated
with 0, 2.5, 5, 10, 15 µM aloe emodin for 24 h. The proteins were
extracted using a lysis buffer, which contains 50 mM Tris (pH 7.4),
150 mM NaCl, 1% NP-40, 0.1% SDS and was purchased from
Sigma-Aldrich (Merck KGaA) and the concentration was determined
using the folin-phenol method. A total of 30 µg lysate protein per
lane was subjected to 10% SDS-polyacrylamide gel electrophoresis.
Following separation, bands were transferred to polyvinylidende
difluoride membranes and incubated overnight at 4°C with anti-p-ERK
(MK12; cat. no. 610030; 1:1,000) and T-ERK (cat. no. 610235,
1:5,000) (both from BD Biosciences San Jose, CA, USA), p-MSK1
(Ser360; cat. no. 9594; 1:1,000; Cell Signaling Technology, Inc.,
Danvers, MA, USA), T-MSK1 (cat. no. NB120-2562; 1:1,000; Novus
Biologicals, LLC, Littleton, CO, USA), p-PDK1 (Ser241; cat. no.
3438S; 1:1,000), T-PDK1 (cat. no. 3062; 1:1,000), p-AKT (cat. no.
9614; 1:1,000), T-AKT (cat. no. 9272; 1:1,000), p-GSK3 (cat. no.
9327; 1:200), T-GSK3 (cat. no. 7265; 1:100) antibodies and β-actin
control antibody (cat. no. 8457; 1:1,000) (all from Cell Signaling
Technology, Inc.). Following hybridization with a horseradish
peroxidase-conjugated secondary antibody (1:2,000; cat. no. Cor:
SE206; Beijing Solarbio Science and Technology Co., Ltd., Beijing,
China) for 30 min at 50°C, polyvinylidende difluoride membranes
were visualized by an enhanced chemiluminescence Detection kit
(Amersham Biosciences; GE Healthcare, Chicago, IL, USA).
Anchorage-independent cell growth
Cells were exposed to 0, 2.5, 5, 10, 15 µM aloe
emodin in 1 ml of 0.33% basal medium (Sigma-Aldrich; Merck KGaA)
Eagle's agar containing 10% FBS. The medium was in a 5%
CO2 incubator at 37°C for 14 days and the cell colonies
were counted under a fluorescence microscope (Olympus) using
Image-Pro Plus version 6.0 (Media Cybernetics, Inc.).
Cell cycle analysis
Cell cycle was analyzed with the Cell Cycle and
Apoptosis Analysis kit (Beyotime Institute of Biotechnology,
Shanghai, China) following 5 min propidium iodide staining in 37°C
with a FACSCalibur flow cytometer (Becton-Dickinson, San Jose, CA,
USA).
Transfection and luciferase assay
The human cyclin D1 promoter reporter plasmid (1745
CD1 LUC) containing the full-length cyclin D1 gene promoter was
provided by RG Pestell (Albert Einstein College of Medicine, Bronx,
NY, USA). Transient transfection was performed by using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). Briefly, 6.5×105 cells/well were
transfected with 1.5 µg β-galactosidase gene (TTP0042 of Thermus
thermophilus HB27) for normalization. JB6 C141 cells were
cotransfected with the F-12K medium (Cellgro; Corning, Inc.,
Corning, NY, USA). A protease inhibitor cocktail (Sigma-Aldrich;
Merck KGaA) was added to F-12K medium containing FBS 3 h
post-transfection. Cells were incubated for 24 h, lysed with a
reporter lysis buffer (Promega) and collected for luciferase assays
and P-cyclin D1 activity using a dual-luciferase assay kit
(Promega).
Statistical analysis
All quantitative data are expressed as the mean ±
standard deviation. The one-way analysis of variance and
Student-Newman-Keuls q test was used for statistical analysis with
Statistic Package for Social Science (SPSS 21.0; IBM Corp., Armonk,
NY, USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Cytotoxic activity of aloe emodin
Previous studies determined that aloe emodin may
selectively inhibit human tumor cell transformation and growth
(19,20). High concentration of aloe emodin
may lead to cell death; therefore, it is necessary to identify an
appropriate dose to inhibit the tumor cell transformation without
cytotoxicity. In the present study, the potential cytotoxicity of
aloe-emodin was evaluated on exponentially growing cells at two
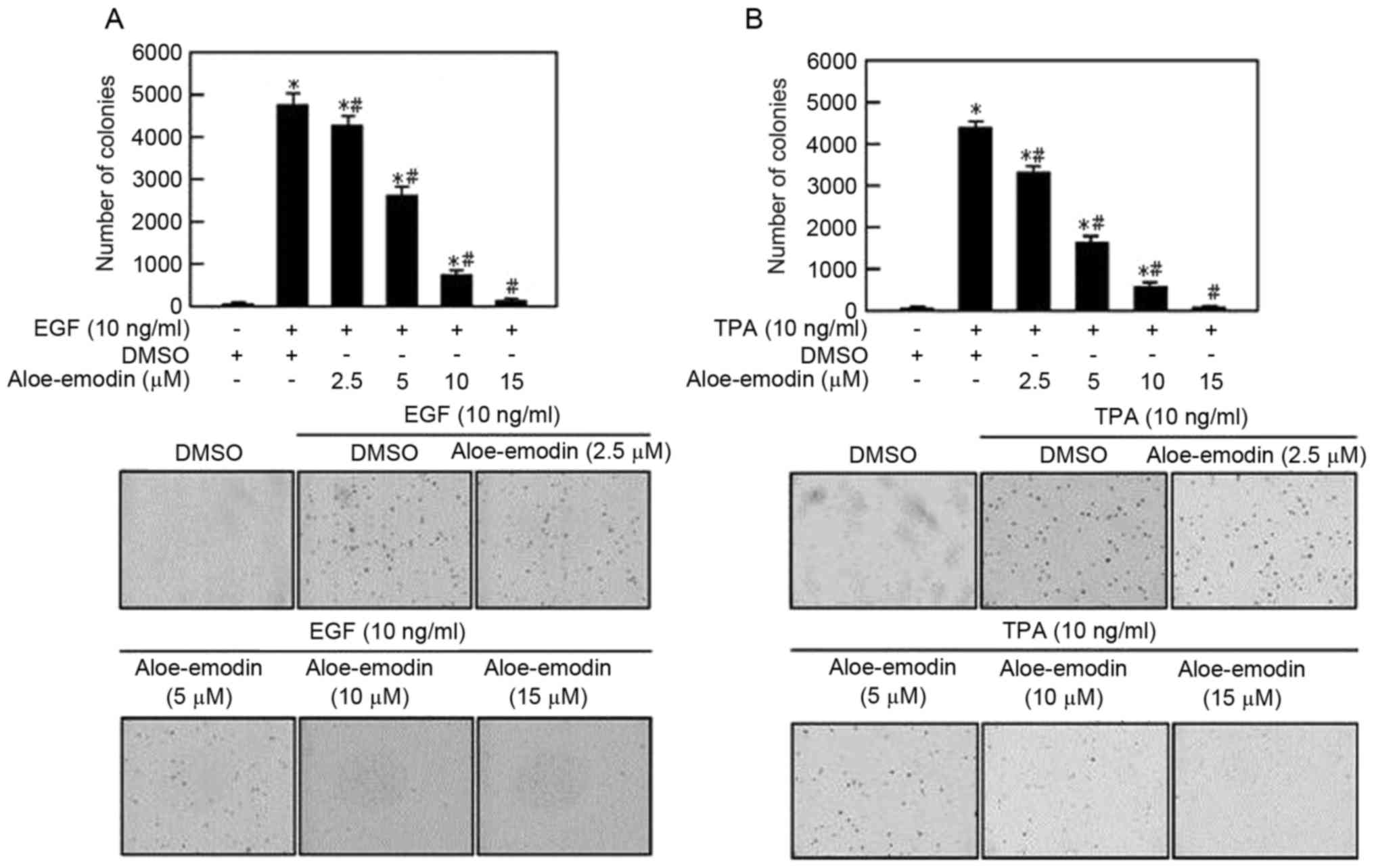
periods (24 and 48 h). As is demonstrated in Fig. 1, aloe-emodin exhbitis a specific
dose-dependent cytotoxic effect on EGF- and tissue plasminogen
activator (TPA)-induced JB6 C141 cells.
Aloe emodin inhibits EGF and TPA
induced JB6 C141 transformation
Cell transformation is a key event of tumorigenesis
(21). Aloe emodin exhibited
anti-tumor effects against PC3 androgen refractory prostate cells
(8). In order to verify whether
aloe emodin inhibited cell transformation, the present study
examined the effect of aloe emodin on anchorage-independent growth
of EGF-induced JB6 C141 cells. Aloe emodin-treated cells had an
impaired anchorage-independent growth capability, leading to a
dose-dependent reduction in colony formation (Fig. 1A) and TPA-induced JB6 C141 cells
were also observed to have reduced colony formation (Fig. 1B).
Aloe emodin inhibits EGF induced MSK
and AKT activation
In order to identify the mechanism and molecular
target of aloe emodin, the present study treated EGF-induced JB6
C141 cells with various aloe emodin quantities for 24 h. The data
indicated that EGF-induced phosphorylation of ERK was not affected
by aloe emodin treatment. However, EGF-induced phosphorylation of
MSK1 was inhibited by aloe emodin in a dose-dependent manner
(Fig. 2A). In addition, aloe
emodin could inhibit the phosphorylation of PDK1 and GSK3β (Ser9),
which were demonstrated to be upstream and downstream of AKT.
Phosphorylated-AKT (Ser473) was inhibited by aloe emodin in a
dose-dependent manner. These findings suggested that aloe emodin
may inhibit AKT alone or downstream components as opposed to the
upstream regulators of EGF-induced PDK1-AKT-GSK3β pathway (Fig. 2B).
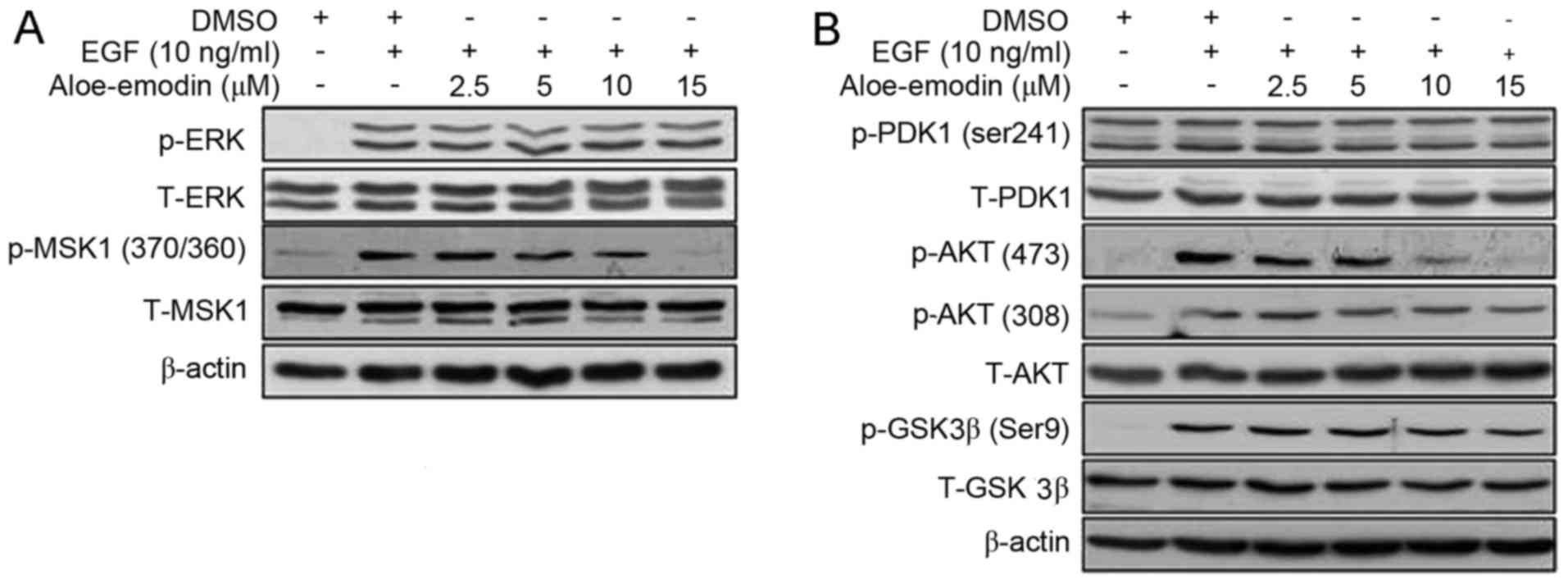 | Figure 2.Aloe emodin inhibited EGF-induced MSK1
and AKT activation. EGF-induced JB6 C141 cells were treated for 24
h with the indicated dose of aloe emodin. The levels of (A) p- and
T-ERK, p- and T-MSK1, (B) p- and T-PDK1, p- and T-AKT and p- and
T-GSK3β proteins were visualized by western blotting with specific
primary and horseradish peroxidase-conjugated secondary antibodies.
Every experiment was repeated three times. EGF, epidermal growth
factor; p, phosphorylated; T, total; MSK1, lysine-tRNA ligase MSK1;
AKT, AKT serine/threonine kinase 1; ERK, extracellular-signal
regulated kinase; PDK, pyruvate dehydrogenase kinase 1; GSK3β,
glycogen synthase kinase 3β. |
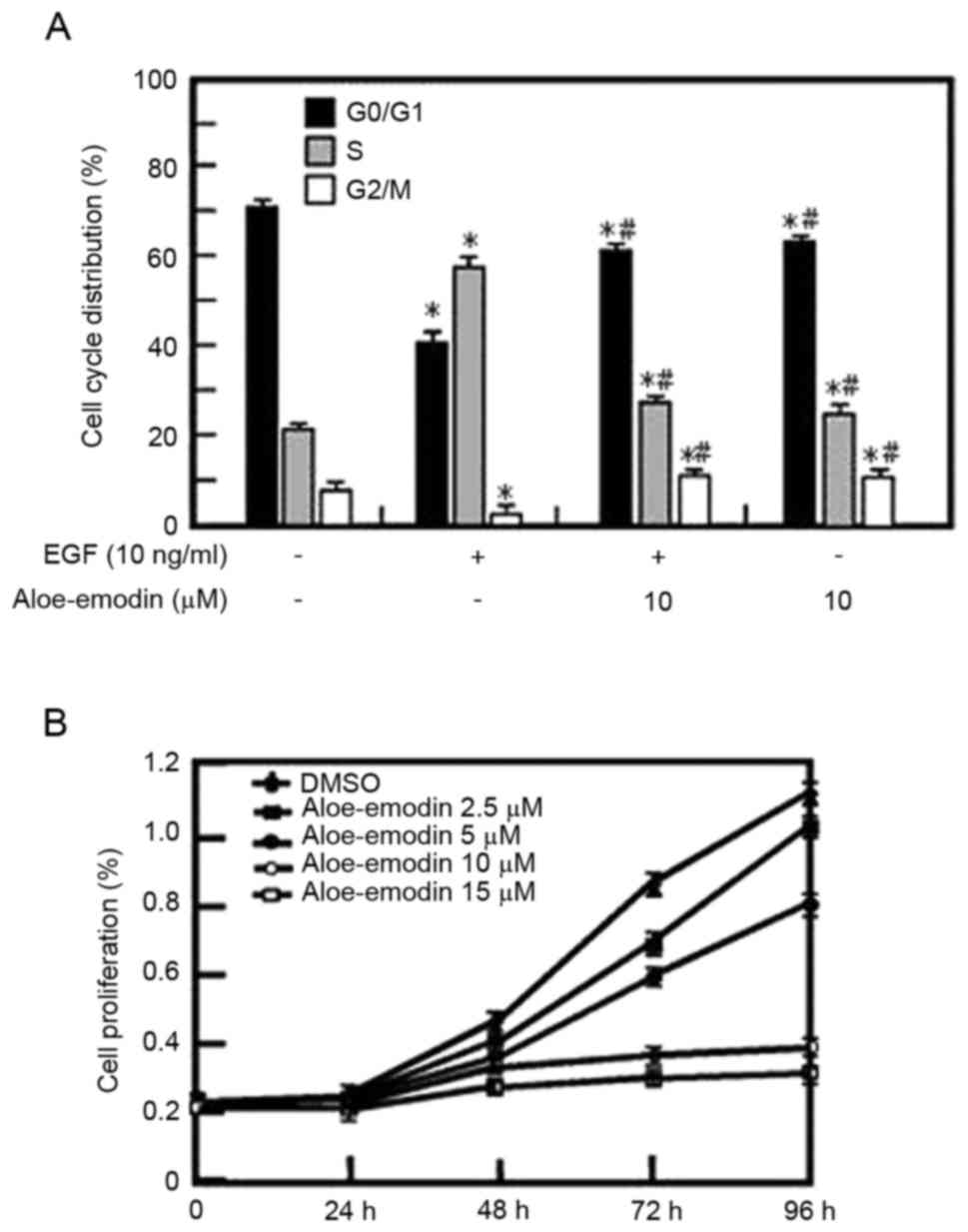
Aloe emodin inhibits EGF-induced JB6
C141 cells proliferation in dose-dependent manner and leads to cell
cycle arrest at the G1 phase
The proliferation and transformation abilities of
tumor cells are associated with the cell cycle; therefore, the
present study used cell cycle analysis. The cell cycle distribution
of the EGF-induced JB6 C141 cells was analyzed (Fig. 3A). It was revealed that the
proliferation of JB6 C141 cells was reduced dose-dependently by
aloe emodin treatment (Fig. 3B)
and the effect may be associated with its inhibition of the G1/S
cell cycle transition (Fig. 3A).
Additionally, the present study indicated that aloe emodin
inhibited EGF-induced cell transformation in a dose-dependent
manner. These findings demonstrated that aloe emodin suppressed the
activity of MSK1 and AKT in the ERK/MSK1 and AKT/GSK3β signaling
pathways leading to in the suppression of EGF-induced cell
proliferation and transformation.
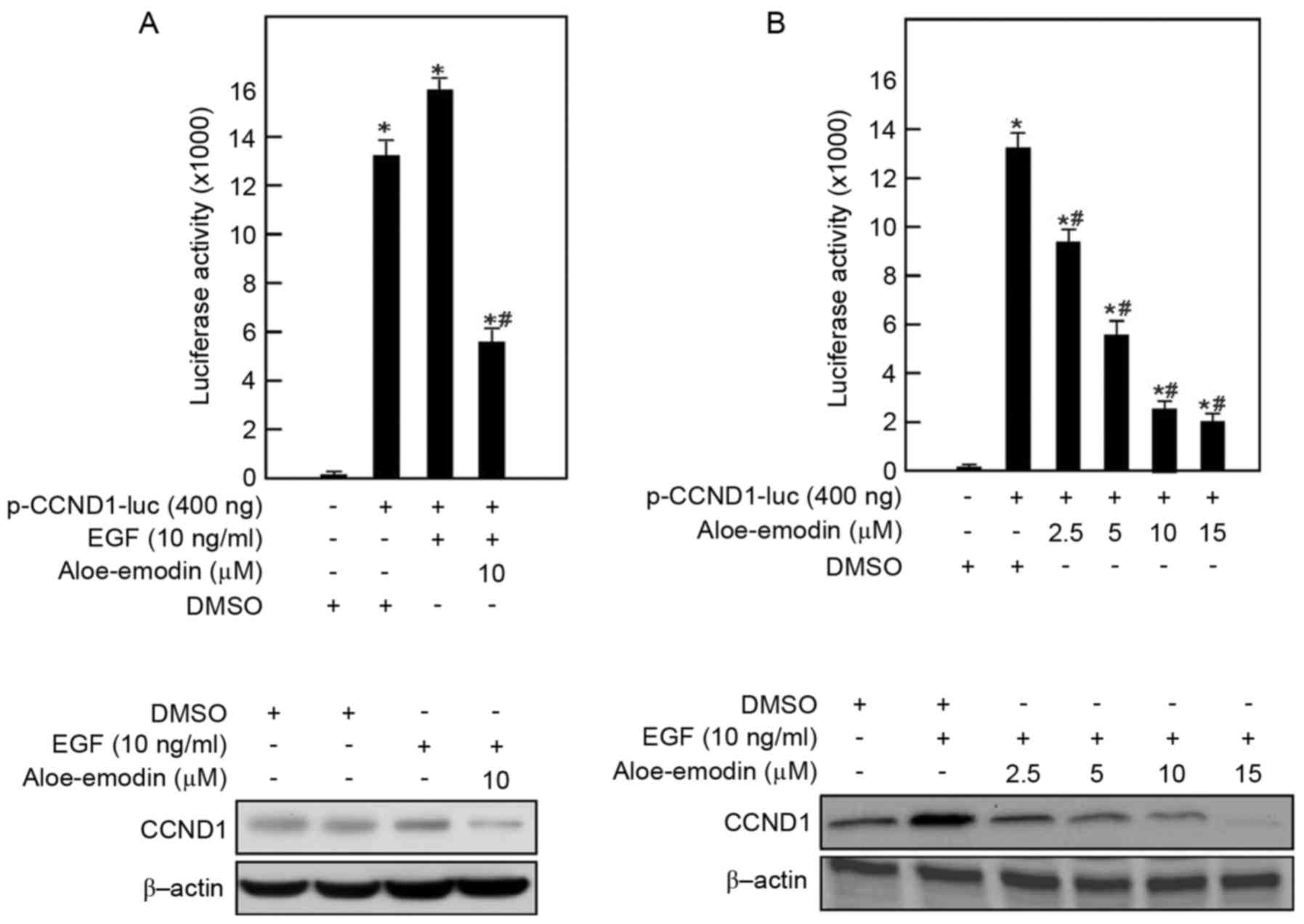
Aloe emodin downregulates the
transcriptional activity of cyclin D1 in a dose-dependent
manner
Cyclin D1 has an important role in the G1/G0 phase
of the cell cycle. The present study determined that aloe emodin
may lead to cell cycle arrest at the G1 phase. In order to
determine whether aloe emodin modulated the EGF-inducing
transcriptional activity of cyclin D1, reporter gene assay was
conducted with EGF-inducing JB6 C141 cells (Fig. 4). In this assay, EGF-induced JB6
C141 cells were transiently transfected with plasmids encoding
luciferase-driven by a promoter containing cyclin D1 sites, and
treated with aloe emodin at 2.5–15 µmol/l. The present study
revealed that aloe emodin reduced the cyclin D1 transcriptional
activity in EGF-induced JB6 C141 cells in a dose-dependent manner.
After 24 h of treatment, 10 µmol/l aloe emodin reduced cyclin D1
transcriptional activity to 25% of that in the DMSO-treated cells.
The protein expression level of cyclin D1 was also inhibited in a
dose-dependent manner.
Discussion
Aloe emodin is one of primary components of Rhei
Rhizoma and has been used for the treatment of various inflammatory
diseases in traditional Chinese medicine. Previous studies have
reported that aloe emodin may exhibit antitumor effects, it may be
able to selectively inhibit the growth and transformation of human
tumor cells (18,22). However, these findings were
primarily focused on the effect aloe emodin had on cell
proliferation as opposed to the influence of cell transformation.
Therefore, the present study used JB6 C141 cells (a common cell
transformation model) and treated EGF-induced JB6 C141 cells with
different doses of aloe emodin.
The current study determined that aloe emodin may
inhibit the EGF-induced or TPA-induced JB6 C141 cell proliferation
and anchorage-independent growth (Figs. 1 and 3B). Additionally, it inhibited the cell
proliferation and transformation in a dose-dependent manner.
Subsequently the present study determined that when the JB6 C141
cells were stimulated with tumor promoters, EGF or TPA, the
expression levels of phosphorylated MSK1 and AKT was significantly
reduced in a dose-dependent manner following aloe emodin treatment
(Fig. 2). The current findings
indicated that aloe emodin inhibited cell growth and
differentiation by interrupting the ERK/MSK1 and PDK1/AKT/GSK3β
signaling pathways. Therefore, ERK/MSK1 and PDK1/AKT/GSK3β
signaling pathways may have critical roles in EGF-induced JB6 C141
cell transformation and proliferation. Previous studies revealed
that the ERK/MSK1 signaling pathway participated in cell migration
and invasion (23), whereas the
PDK1/AKT/GSK3β signaling pathway was associated with cell
proliferation (24).
In addition, the present study identified that the
phosphorylated-AKT (Ser473) and GSK3β (Ser9) were inhibited by aloe
emodin treatment in a dose-dependent manner (Fig. 2). Therefore, the present study
hypothesized that a reduction of the phosphorylation-AKT (Ser473)
may downregulate the activation of GSK3β (Ser9).
Furthermore, proliferation was inhibited by 10
µmol/l aloe emodin when compared with the control group. The effect
was associated with the impairment of the G0/G1 cell cycle
transition which was associated with the PDK1/AKT/GSK3β signaling
pathway (25), particularly the
cyclin D1 protein (26) (Fig. 3A). The luciferase activity assays
revealed that cyclin D1 transcription was suppressed by aloe emodin
treatment in EGF-induced JB6 C141 cells (Fig. 4). The proliferation of EGF-induced
JB6 C141 cells was reduced in a dose-dependent manner by aloe
emodin treatment (Fig. 3B). A
previous study concluded that the reduction of cyclin D1 may arrest
the cell cycle in the G2/M phase (16). However, the present study revealed
that lower concentrations of aloe emodin had the same effect.
Therefore aloe emodin treatment may lead to G0/G1 cell cycle arrest
when used in small doses.
Subsequently, the present study focused on the
inhibitory mechanism of aloe emodin. Aloe emodin treatment may lead
to in the dephosphorylation-AKT (Ser473), inhibition of GSK3β
(Ser9), and effectively limit cyclin D1 degradation. A previous
study indicated that the AKT/GSK3β signaling pathway had an
important role in regulating the expression of cyclin D1 (27). However, the downstream regulators
of GSK3β activating cyclin D1 remain to be elucidated.
In conclusion, the present study demonstrated that
aloe emodin effectively suppressed transformation and proliferation
in a small dose by inhibiting AKT and MSK1 activity. Aloe emodin
was able to inhibit the transcriptional activity of cyclin D1 by
suppressing the EGF/AKT/GSK3β signaling pathway. These findings
revealed that aloe emodin may be a potential natural
chemopreventive compound for human tumors.
References
|
1
|
Yewale C, Baradia D, Vhora I, Patil S and
Misra A: Epidermal growth factor receptor targeting in cancer: A
review of trends and strategies. Biomaterials. 34:8690–8707. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mitsudomi T and Yatabe Y: Epidermal growth
factor receptor in relation to tumor development: EGFR gene and
cancer. Febs J. 277:301–308. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Drobic B, Espino PS and Davie JR: Mitogen-
and stress-activated protein kinase 1 activity and histone h3
phosphorylation in oncogene-transformed mouse fibroblasts. Cancer
Res. 64:9076–9079. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vermeulen L, Vanden Berghe W, Beck IM,
Bosscher K and Haegeman G: The versatile role of MSKs in
transcriptional regulation. Trends Biochem Sci. 34:311–318. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Seshacharyulu P, Ponnusamy MP, Haridas D,
Jain M, Ganti AK and Batra SK: Targeting the EGFR signaling pathway
in cancer therapy. Expert Opin Ther Targets. 16:15–31. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chang S, Iversen L, Kragballe K, Arthur JS
and Johansen C: Mice lacking MSK1 and MSK2 show reduced skin tumor
development in a two-stage chemical carcinogenesis model. Cancer
Invest. 29:240–245. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pérez-Cadahía B, Drobic B, Espino PS, He
S, Mandal S, Healy S and Davie JR: Role of MSK1 in the malignant
phenotype of Ras-transformed mouse fibroblasts. J Biol Chem.
286:42–49. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu K, Park C, Li S, Lee KW, Liu H, He L,
Soung NK, Ahn JS, Bode AM, Dong Z, et al: Aloe emodin suppresses
prostate cancer by targeting the mTOR complex 2. Carcinogenesis.
33:1406–1411. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen XL, Ren KH, He HW and Shao RG:
Involvement of PI3K/AKT/GSK3beta pathway in tetrandrine-induced G1
arrest and apoptosis. Cancer Biol Ther. 7:1073–1078. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vermeulen K, Van Bockstaele DR and
Berneman ZN: The cell cycle: A review of regulation, deregulation
and therapeutic targets in cancer. Cell Prolif. 36:131–149. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen MJ, Cheng AC, Lee MF and Hsu YC:
Simvastatin induces G1 arrest by up-regulating GSK3beta and
down-regulating CDK4/cyclin D1 and CDK2/cyclin E1 in human primary
colorectal cancer cells. J Cell Physiol. Aug 18–2017.doi:
10.1002/jcp.26156 (Epub ahead of print).
|
|
12
|
Sutherland RL and Musgrove EA: Cyclin D1
and mammary carcinoma: New insights from transgenic mouse models.
Breast Cancer Res. 4:14–17. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pecere T, Sarinella F, Salata C, Gatto B,
Bet A, Dalla Vecchia F, Diaspro A, Carli M, Palumbo M and Palù G:
Involvement of p53 in specific anti-neuroectodermal tumor activity
of aloe emodin. Int J Cancer. 10:836–847. 2003. View Article : Google Scholar
|
|
14
|
Knudsen KE, Diehl JA, Haiman CA and
Knudsen ES: Cyclin D1: polymorphism, aberrant splicing and cancer
risk. Oncogene. 25:1620–1628. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hao L and ElShamy WM: BRCA1-IRIS activates
cyclin D1 expression in breast cancer cells by downregulating the
JNK phosphatase DUSP3/VHR. Int J Cancer. 121:39–46. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Deharvengt SJ, Gunn JR, Pickett SB and
Korc M: Intratumoral delivery of shRNA targeting cyclin D1
attenuates pancreatic cancer growth. Cancer Gene Ther. 17:325–333.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen R, Zhang J, Hu Y, Wang S, Chen M and
Wang Y: Potential antineoplastic effects of Aloe-emodin: A
comprehensive review. Am J Chin Med. 42:275–288. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Woo SW, Nan JX, Lee SH, Park EJ, Zhao YZ
and Sohn DH: Aloe emodin suppresses myofibroblastic differentiation
of rat hepatic stellate cells in primary culture. Pharmacol
Toxicol. 90:193–198. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chiu TH, Lai WW, Hsia TC, Yang JS, Lai TY,
Wu PP, Ma CY, Yeh CC, Ho CC, Lu HF, et al: Aloe-emodin induces cell
death through S-phase arrest and caspase-dependent pathways in
human tongue squamous cancer SCC-4 cells. Anticancer Res.
29:4503–4511. 2009.PubMed/NCBI
|
|
20
|
Huang PH, Huang CY, Chen MC, Lee YT, Yue
CH, Wang HY and Lin H: Emodin and aloe-emodin suppress breast
cancer cell proliferation through ER α inhibition. Evid Based
Complement Alternat Med. 2013:3761232013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Guo J, Xiao B, Zhang S, Liu D, Liao Y and
Sun Q: Growth inhibitory effects of gastric cancer cells with an
increase in S phase and alkaline phosphatase activity repression by
aloe emodin. Cancer Biol Ther. 6:85–88. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kang SC, Lee CM, Choung ES, Bak JP, Bae
JJ, Yoo HS, Kwak JH and Zee OP: Anti-proliferative effects of
estrogen receptor-modulating compounds isolated from Rheum
palmatum. Arch Pharm Res. 31:722–726. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lee CJ, Lee MH, Yoo SM, Choi KI, Song JH,
Jang JH, Oh SR, Ryu HW, Lee HS, Surh YJ and Cho YY: Magnolin
inhibits cell migration and invasion by targeting the ERKs/RSK2
signaling pathway. BMC Cancer. 15:5762013. View Article : Google Scholar
|
|
24
|
Zhang Q, Yan HB, Wang J, Cui SJ, Wang XQ,
Jiang YH, Feng L, Yang PY and Liu F: Chromatin remodeling gene
AT-rich interactive domain-containing protein 1A suppresses gastric
cancer cell proliferation by targeting PIK3CA and PDK1. Oncotarget.
7:46127–46141. 2016.PubMed/NCBI
|
|
25
|
Vadlakonda L, Pasupuleti M and Pallu R:
Role of PI3K-AKT-mTOR and Wnt signaling pathways in transition of
G1-S phase of cell cycle in cancer cells. Front Oncol. 3:852013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hinz M, Krappmann D, Eichten A, Heder A,
Scheidereit C and Strauss M: NF-kappaB function in growth control:
Regulation of cyclin D1 expression and G0/G1-to-S-phase transition.
Mol Cell Biol. 19:2690–2698. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lai SS, Zhao DD, Cao P, Lu K, Luo OY, Chen
WB, Liu J, Jiang EZ, Yu ZH, Lee G, et al: PP2Acα positively
regulates the termination of liverregeneration in mice through the
AKT/GSK3β/Cyclin D1pathway. J Hepatol. 64:352–360. 2012. View Article : Google Scholar
|