Introduction
Intervertebral disc degeneration (IDD) is considered
to be closely associated with low back pain-associated diseases,
including lumbar disc herniation, lumbar spinal stenosis, and
degenerative lumbar spondylolisthesis (1,2). Low
back pain is one of the common symptoms of IDD, which is a leading
cause of disability with a high economic burden worldwide (3). Stimulation of the inflammatory
response, mechanical stress and biochemical effectors have been
implicated as active components in a number of events and processes
associated with IDD (4,5). However, the specific pathological
mechanisms and effective treatment methods remain unknown (6).
Intervertebral discs are composed of the nucleus
pulposus (NP), annulus fibrosus and cartilaginous endplates. NP
cells have been proposed to serve a crucial role in the
physiological function of intervertebral discs via maintaining
homeostasis among various components of the extracellular matrix
(ECM) (7). When the homeostasis of
the catabolic and anabolic activities within the ECM, which include
collagen I, collagen II and proteoglycans, are disrupted, IDD may
occur (8,9). The process of ECM breakdown is caused
and promoted by matrix metalloproteinases (MMPs) and a disintegrin
and metalloproteinase with thrombospondin motifs (ADAMTS) (10,11).
Among these molecules, MMP-3, MMP-9, MMP-13, and ADAMTS-4 and
ADAMTS-5 are specific typical representatives of degradative matrix
components, which have been identified to be highly expressed in
IDD tissues and are involved in normal cell turnover and
pathological degradation of the major structural components of
intervertebral discs (12,13). The activities of MMPs may be
controlled by tissue inhibitors of metalloproteinases (TIMPs),
which also participate in the maintenance of ECM homeostasis. Among
TIMPs, TIMP-1 is closely associated with MMP-3 and MMP-13, and
downregulates their activities (14).
Maintaining the stability of viable NP cell numbers
is also considered to serve a pivotal role in maintaining the
normal function of the NP. Apoptosis, or type I programmed cell
death, is an important part of normal cell growth cycle. However,
excessive NP cell apoptosis, induced by various pathogenic factors,
often causes a decrease in the number of viable cells in the NP,
which is associated with IDD (15). Interleukin (IL)-1β, a key
proinflammatory cytokine, is an important mediator that causes
uncontrolled NP cells apoptosis and degeneration of the ECM
(16). As a result, suppression of
IL-1β-mediated apoptosis and degeneration of the ECM may be a
potential strategy to alleviate IDD development.
The nuclear factor kappa-light-chain-enhancer of
activated B cells (NF-κB) signaling pathway serves as a crucial
mediator of inflammatory responses and induces the expression of
various pro-inflammatory genes, including those encoding cytokines
and chemokines (17). In addition,
the NF-κB pathway becomes active when responding to a number of
different types of stimuli, including oxidative, mechanical,
genotoxic and chemical factors, which finally results in the
nuclear accumulation of NF-κB transcription factors, consequently
affecting the expression of target genes (18). Previous studies have suggested that
the NF-κB pathway may markedly increase the production of specific
MMPs and ADAMTS to compromise the normal ECM by degrading it in
animal NP cells (19). Similar to
the NF-κB pathway, the toll-like receptor (TLR)-4/myeloid
differentiation primary response protein MyD88 (MyD88)/NF-κB
signaling pathway was identified to be critical in molecular
modulation, and has also been demonstrated to be involved in the
inflammatory response (20–24).
Andrographolide (AG), a type of diterpenoid
extracted from Andrographis paniculata, has been used for
patients suffering from infectious diseases of different etiologies
(25). It has been well documented
that AG possesses potent anti-inflammatory capabilities in
conditions associated with inflammation, including certain
bacterial or viral infections, tumors and other chronic diseases
(24,26–28).
It has been reported that AG is a potential therapeutic agent for
IL-1β-induced cartilage degeneration and the production of
inflammatory factors involved in the pathogenesis of osteoarthritis
(29). Additionally, the study of
tumor and immune-associated diseases also revealed that the
therapeutic effects of AG are closely associated with the
TLR4/MyD88/NF-κB signaling pathway (20,24).
In addition, it has been well established that abnormal apoptosis
may be modulated by AG treatment in certain pathological states
(30,31). All of these data indicate that AG
may be a therapeutic agent for treating and preventing inflammatory
diseases; however, the effects in IDD have not been well
explored.
Therefore, on the basis of these data, we
hypothesized that AG may have protective properties against the
degeneration of NP cells via inhibiting the TLR4/MyD88/NF-κB
signaling pathway. In the present study, human NP cells were used
to mimic the model of IDD in vitro, and the
anti-inflammatory effect and underlying mechanism of AG on the
IL-1β-induced degeneration of NP cells was investigated.
Materials and methods
Reagents and antibodies
AG was purchased from Sigma-Aldrich; Merck KGaA
(Darmstadt, Germany). Dulbecco's modified Eagle's medium (DMEM) and
fetal bovine serum (FBS) were purchased from Thermo Fisher
Scientific, Inc., (Waltham, MA, USA). Anti-collagen (cat. no.
MA1-37493; 1:200), anti-aggrecan (cat. no. MA3-16888; 1:200) and
anti-TRL-4 (cat. no. 48-2300; 1:50) were purchased from Thermo
Fisher Scientific, Inc. (Waltham, MA, USA). Anti-MyD88 (cat. no.
4283; 1:500), anti-transcription factor p65 (p65; cat. no. 8242;
1:100), anti-phosphorylated (p)-p65 (cat. no. 3033; 1:500),
anti-B-cell lymphoma 2 (Bcl-2; cat. no. 3498; 1:1,000),
anti-Bcl-2-associated X protein (Bax; cat. no. 5023; 1:1,000),
anti-cleaved caspase 3 (cat. no. 9661; 1:1,000), and goat
anti-rabbit (cat. no. 7074; 1:1,000) and anti-mouse (cat. no. 7076;
1:1,000) IgG-horseradish peroxidase secondary antibodies were
purchased from Cell Signaling Technology, Inc., (Danvers, MA, USA).
β-actin antibodies (cat. no. sc-81178; 1:200) were purchased from
Santa Cruz Biotechnology (Dallas, TX, USA). An enhanced
chemiluminescence (ECL) kit was purchased from Bio-Rad
Laboratories, Inc., (Hercules, CA, USA). All of the other reagents
were purchased from Sigma-Aldrich; Merck KGaA unless specified
otherwise.
NP cell isolation and culture
All the tissues from human discs were collected as
surgical waste from patients undergoing spinal surgical procedures
(Fig. 1A). Young patients
diagnosed with lumbar disc herniation were considered ideal
candidates. In addition, mild or absent degeneration of the lumbar
intervertebral discs in MRI were required. The present study was
approved by the Ethics Committee of the Second Affiliated Hospital
and Yuying Children's Hospital of Wenzhou Medical University
(Wenzhou, China), and informed consent for the collection of
tissues was acquired from patients or relatives. The Pfirrmann
grading system was used to evaluate the degenerative conditions of
the discs (32). Subsequent to
collection of the NP tissues, the NP cells were isolated using
0.25% trypsin and 0.2% type II collagenase for ~3 h at 37°C and
used within the first 3 passages. Then, the NP cells were cultured
in DMEM supplemented with 10% FBS and antibiotics (1%
penicillin/streptomycin) at 37°C in a humidified atmosphere of 5%
CO2. During passage, no significant changes in the
morphology of cells was identified between the primary cells
(passage 0) and later passage cells (passage 2).
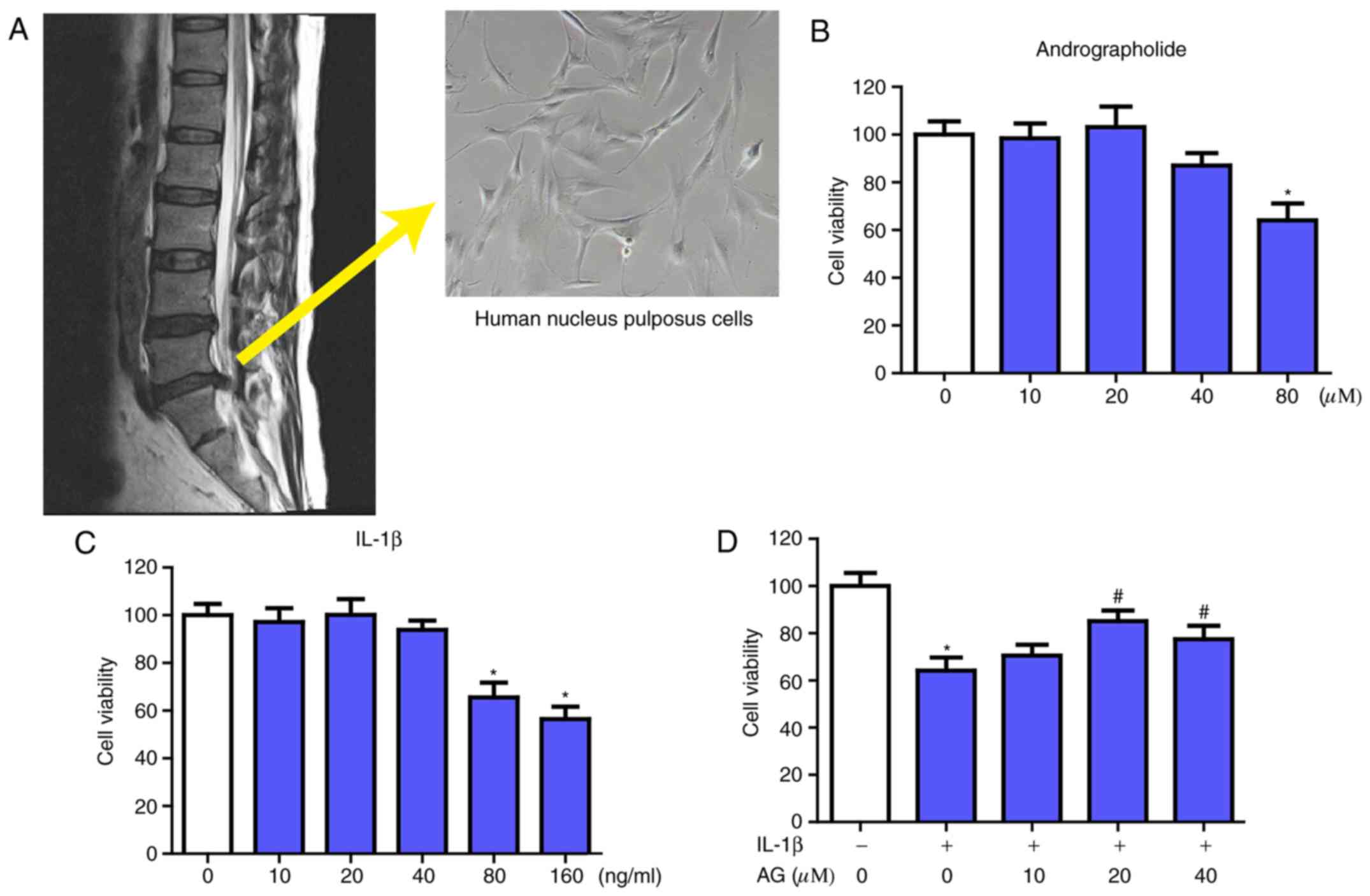 | Figure 1.Acquisition and morphology of NP
cells and the effect of AG and IL-1β on NP cells viability. (A) The
NP cells were collected from patients undergoing spinal surgical
procedures, and the morphology of NP cells (magnification, ×400)
was investigated. The NP cells obtained were then cultured with
increasing concentrations of (B) andrographolide (0, 10, 20, 40 and
80 µM) and (C) IL-1β (0, 10, 20, 40, 80 and 160 ng/ml) for 24 h,
followed by the Cell Counting kit-8 analysis for cell viability.
(D) NP cells were pretreated for 2 h with various concentrations of
andrographolide (0, 10, 20 and 40 µM) and then stimulated with
IL-1β (160 ng/ml) for 24 h, followed by Cell Counting kit-8
analysis of cell viability. The values are presented as the mean ±
standard deviation of 3 independent experiments. *P<0.05 vs.
control group; #P<0.05 vs. IL-1β only group (0 µM
AG). NP, nucleus pulposus; IL-1β, interleukin 1β; AG,
andrographolide. |
Cell viability assays
The viability of the NP cells was assessed by the
Cell Counting kit-8 (CCK-8) assay (Dojindo Laboratories, Kumamoto,
Japan). For this assay, the cells were seeded at a density of 5,000
cells/well in 96-well plates with 6 replicate wells. DMEM was used
to dilute the CCK-8 reagent 10-fold (10:100 µl) prior to addition
to each well. Following incubation at 37°C for 3 h, the absorbance
of each well was then measured at 450 nm by a microplate reader.
The optical density at 450 nm is proportional to the degree of cell
viability.
RNA isolation and reverse
transcription quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted by a TRIzol®
reagent (Thermo Fisher Scientific, Inc.) according to
manufacturer's protocol. RNA was reverse transcribed into cDNA
using a PrimeScript RT Reagent kit (Takara Biotechnology Co., Ltd.,
Dalian, China), and RT-qPCR was conducted using SYBR Green Supermix
(QPK-212; Toyobo Life Science, Osaka, Japan) and a Light Cycler480
system (Roche Diagnostics GmbH, Mannheim, Germany). The reverse
transcription reaction was conducted under the following
conditions: 37°C for 15 min and 85°C for 5 sec, then held at 4°C.
To amplify genomic DNA, qPCR was performed under the following
conditions: 95°C for 5 min, followed by 40 cycles of 95°C for 10
sec, 60°C for 10 sec, and 72°C for 10 sec. The cycle threshold (Cq)
values were collected and normalized to the level of the
housekeeping gene GAPDH. The relative expression levels were
analyzed using the 2−ΔΔCq method (33). The RT-qPCR was performed with the
following specific primers: MMP-3 forward,
5′-CAAGGAGGCAGGCAAGACAGC-3′ and reverse,
5′-GCCACGCACAGCAACAGTAGG-3′; MMP-9 forward,
5′-CTTTGAGTCCGGTGGACGAT-3′ and reverse, 5′-TCGCCAGTACTTCCCATCCT-3′;
MMP-13 forward, 5′-TGCTTCCTGATGACGATGTAC-3′ and reverse,
5′-TCCTCGGAGACTGGTAATGG-3′; TMIP-1 forward,
5′-TGGCTTCTGGCATCCTGTTGTTG-3′ and reverse,
5′-CGCTGGTATAAGGTGGTCTGGTTG-3′; ADAMTS-4 forward,
5′-GGATTACAGGTGTGAGCCACCA-3′ and reverse,
5′-GGATGCAACCACATCTGTCTGA-3′; ADAMTS-5 forward,
5′-GCAGTATGACAAGTGCGGAGT-3′ and reverse,
5′-CAGGGCTAAATAGGCAGTGAA-3′.
Western blot analysis
The western blot analysis protocol was performed as
described previously (34).
Briefly, NP cells were homogenized in ice-cold lysis buffer
containing 50 mM Tris-HCl pH 8.0, 150 mM NaCl, 1% NP-40, 0.5%
deoxycholate, 0.1% SDS, 10 mM
Na2P2O7, 10 mM NaF, 1 mg/ml
aprotinin, 10 mg/ml leupeptin, 1 mM sodium vanadate and 1 mM PMSF.
The cells homogenates were incubated for 15 min at 4°C and
centrifuged at 11,792 × g for 15 min at 4°C. The protein
concentration of each sample was determined using the Bicinchoninic
Acid method. The total protein (60 µg) was loaded onto SDS-PAGE
(8–10%) and transferred to polyvinylidene difluoride (PVDF)
membranes (Bio-Rad Laboratories, Inc.). Then, the 5% non-fat milk
dissolved in TBS was used to block the PVDF membranes for 2 h at
room temperature and the membranes were incubated overnight at 4°C
with the aforementioned primary antibodies. Following washing with
TBS 3 times for 5 min, the PVDF membranes were incubated at room
temperature with horseradish peroxidase-conjugated secondary
antibodies to detect the primary antibodies. Signals were
visualized using the ChemiDocTM XRS + Imaging System
(Bio-Rad Laboratories, Inc.,). Densitometric quantification of the
membranes was performed using Image J version 1.4 (National
Institutes of Health, Bethesda, MD, USA). Experiments were repeated
3 times.
Immunofluorescence
The NP cells grown on 14×14 mm microscopic glass
were washed with ice-cold PBS and fixed with ice-cold 4%
paraformaldehyde for 30 min at 4°C. Following washing with ice-cold
PBS, the cells were blocked in 5% bovine serum albumin
(Sigma-Aldrich; Merck KGaA) at room temperature for 1 h. Then, the
NP cells were incubated with primary antibodies (against
collagen-II, 1:200; aggrecan, 1:200; and p65, 1:100) at 4°C
overnight. Following washing with PBS, the cells were incubated
with Alexa Fluor 488-conjugated anti-IgG or Texas red-conjugated
anti-IgG secondary antibodies for 1 h at 37°C to detect the primary
antibodies. The nuclei of cells were stained at room temperature
with DAPI (10 µg/ml) for 5 min and finally washed in PBS and sealed
with a coverslip. All images were captured on a Nikon ECLIPSE Ti-U
fluorescence microscope (Nikon Corporation, Tokyo, Japan;
magnification, ×400).
Statistical analysis
The experiments were repeated at least 3 times. The
results were analyzed using SPSS software, version 22.0 (IBM Corp.,
Armonk, NY, USA) and presented as the mean ± standard error of the
mean from 3 independent experiments. Statistical significance was
examined using one-way analysis of variance and Dunnett's post-hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Effects of AG on the viability of NP
cells with or without IL-1β treatment
The source and morphology of NP cells are presented
in the Fig. 1A. Firstly, the
potential cytotoxicity of AG in NP cells was evaluated using a
CCK-8 assay. Increasing concentrations of AG (0, 10, 20, 40 and 80
µM) and IL-1β (0, 10, 20, 40, 80 and 160 ng/ml) were used to
culture the NP cells for 24 h, followed by the CCK-8 assay. As
indicated in Fig. 1B, AG did not
exhibit significant cytotoxic effects on human NP cells at the
concentrations of 0–40 µM. When treated with IL-1β only, it
demonstrated marked cytotoxic effects in NP cells at the
concentrations of 80 and 160 ng/ml, while there was no significant
change at concentrations of 0–40 ng/ml (Fig. 1C). The viability of the NP cells
following treatment with IL-1β combined with various concentrations
of AG (0–40 µM) was also measured by the CCK-8 assay. The results
indicated that AG mitigated the cytotoxic effects of IL-1β
treatment in a dose-dependent manner, and the concentration of AG
that exhibited the most marked effect was 20 µM (Fig. 1D). Therefore, these results
indicated that AG had no cytotoxic effects, but did exhibit
protective effects on NP cells at concentrations of 20 µM in the
assay prior to IL-1β treatment. Therefore, 20 µM AG was used in the
subsequent experiments.
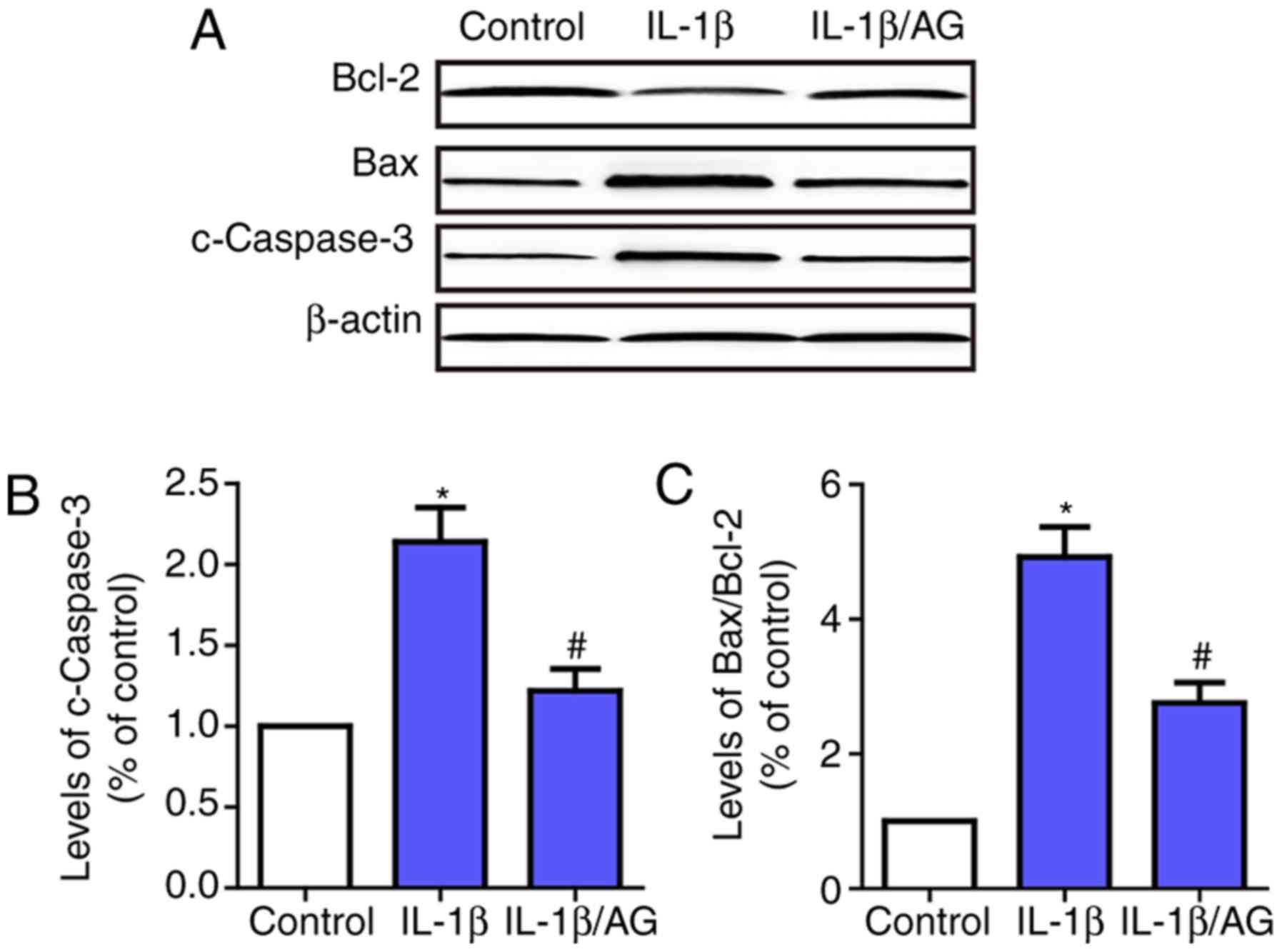
AG attenuates IL-1β induced apoptosis
of NP cells
Subsequently, the effect of AG on IL-1β-induced
apoptosis of NP cells was examined by investigating the changes in
apoptosis-associated enzymes, including caspase-3, Bax and Bcl-2.
As indicated in Fig. 2, the
results suggested that the level of caspase-3 and the Bax/Bcl2
ratio were increased significantly following IL-1β stimulation
compared with the control group. Conversely, AG markedly inhibited
IL-1β-induced expression of caspase-3 and Bax. These results
demonstrated that AG attenuated IL-1β-induced apoptosis of NP
cells.
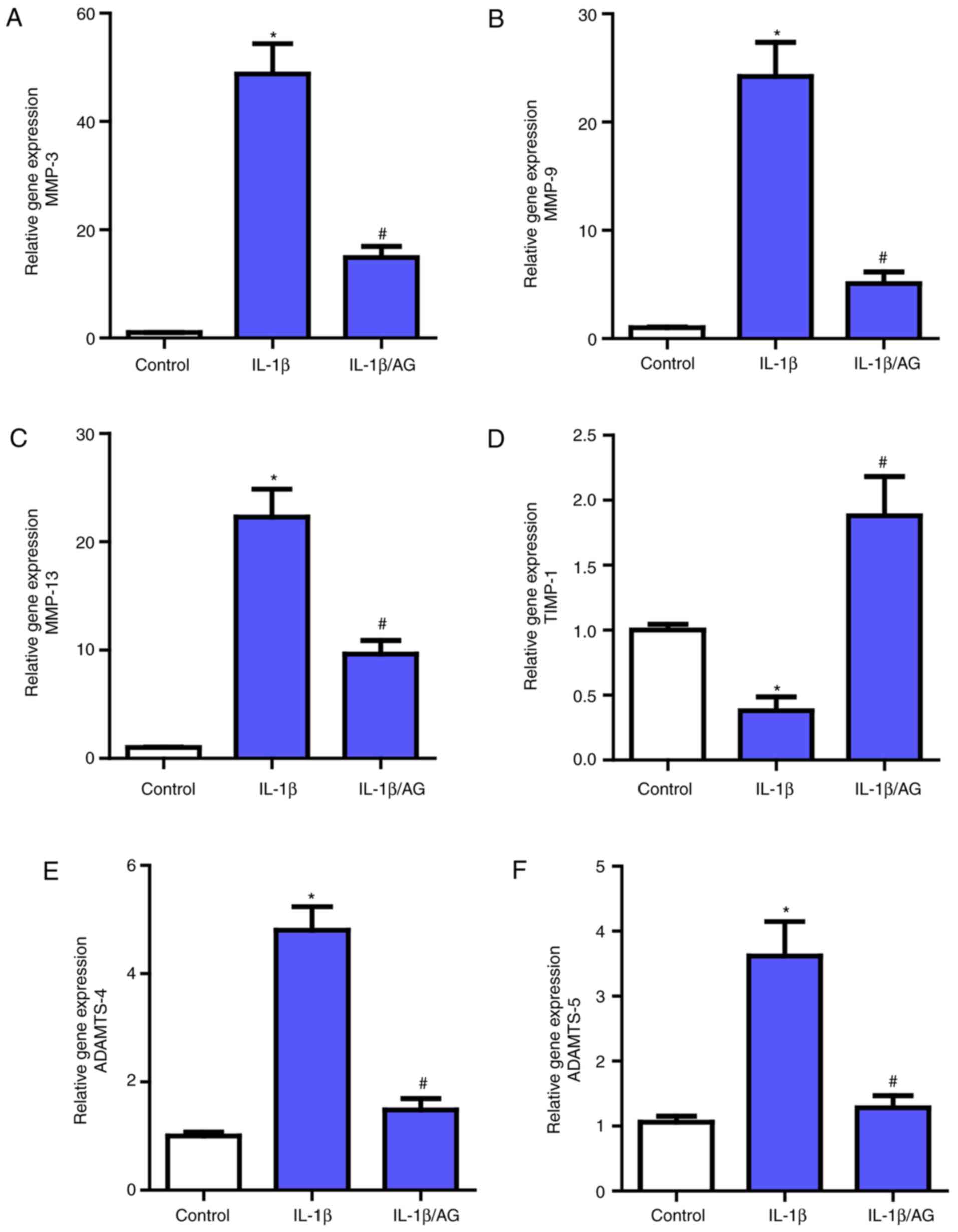
AG suppresses IL-1β induced MMPs and
ADAMTS expression in NP cells
Then, the effect of AG on MMPs (MMP3, MMP-9, MMP-13)
and ADAMTS (ADAMTS-4 and ADAMTS-5) expression in NP cells was
measured and analyzed by RT-qPCR. As demonstrated in Fig. 3, a marked upregulation of the mRNA
expression of these MMPs and ADAMTS following IL-1β stimulation
compared with the control group was observed, which was consistent
with the previously mentioned association between these catabolic
enzymes and NP cell ECM. However, following treatment with AG,
there was a marked inhibition of the effect of IL-1β on these
enzymes, as demonstrated by RT-qPCR. Additionally, as the
endogenous inhibitors of MMPs, the opposite result was observed for
TIMP-1 levels compared with MMPs. These results indicated that AG
may suppress the expression levels of the catabolic enzymes.
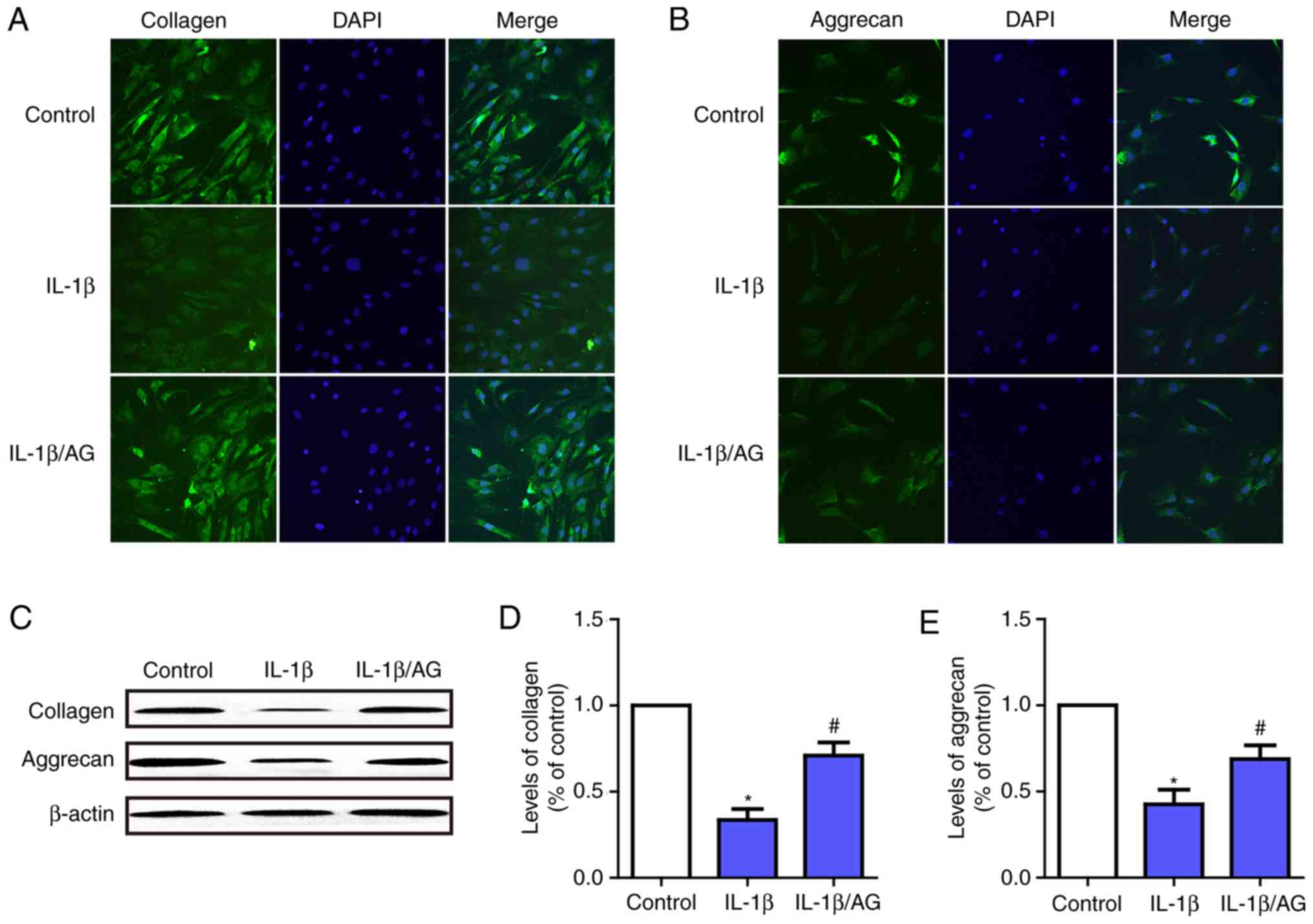
AG mitigates IL-1β-induced NP cells
ECM degradation
To investigate the function of AG on IL-1β-induced
ECM (aggrecan and collagen-II) degradation in NP cells,
immunofluorescence staining and western blot analysis were used. As
presented in Fig. 4A and B,
immunofluorescence staining revealed that IL-1β stimulation
resulted in a significant downregulation of aggrecan and
collagen-II at the mRNA and protein levels. Conversely, this
downregulation was markedly attenuated by AG. Fig. 4C-E demonstrate the results of the
western blot analysis, which were consistent with the results of
the immunofluorescence assay. These results suggested that AG was
likely to serve a role in protecting intervertebral discs by
attenuating ECM degeneration in NP cells.
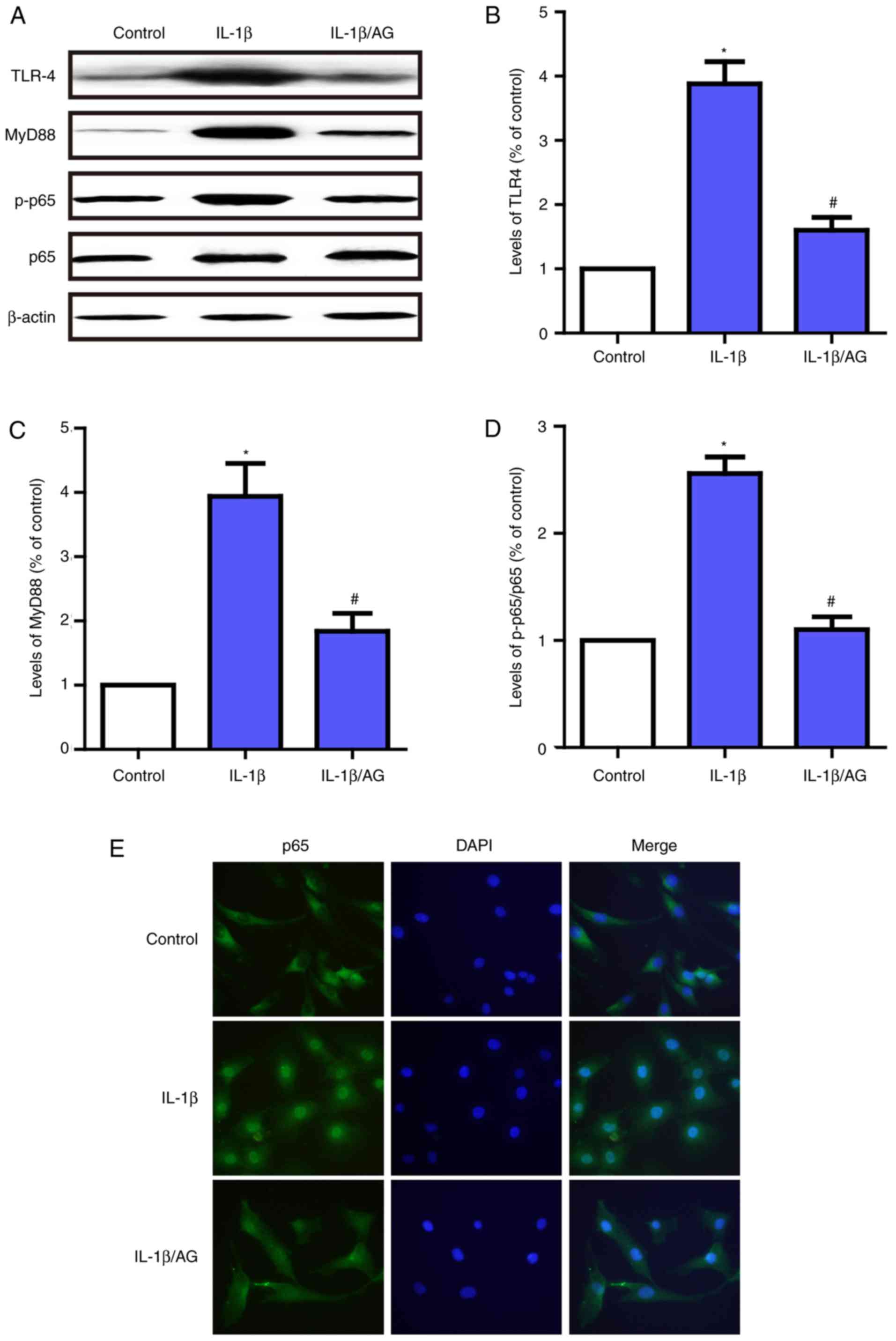
AG protects against NP cell
degradation via the TLR4/MyD88/NF-κB signaling pathway
In order to additionally examine the protective
mechanism of AG in NP cell degeneration, immunofluorescent staining
and western blot analysis were used to detect the effect of AG on
IL-1β-induced TLR4/MyD88/NF-κB activation. As demonstrated in
Fig. 5A-D, the levels of TLR-4 and
MyD88, and the phosphorylation of p65 were significantly
upregulated following stimulation with IL-1β. By contrast, AG
inhibited the effect on IL-1β-induced activation of the
TLR4/MyD88/NF-κB signaling pathway. In addition, the
immunofluorescence results suggested that p65 was primarily
distributed in the cytoplasm of unstimulated NP cells and exhibited
marked localization in the nucleus upon IL-1β stimulation; however,
AG significantly inhibited this translocation induced by IL-1β
(Fig. 5E). These results suggest
that AG has the potential to suppress the expression of TLR-4 and
MyD88, and inhibit NF-κB activation in NP cells. The therapeutic
effects of AG may be exerted via the TLR4/MyD88/NF-κB signaling
pathway.
Discussion
At present, the primary clinical therapy for low
back pain-associated disease is symptomatic, or surgical treatment
to relieve symptoms, and there is no efficacious medication to
mitigate the degenerative progress of the discs. A number of
studies have verified the presence of lumbar IDD as an important
factor for the promotion of the onset and development of low back
pain-associated disease (1,6). It
has been established that the degeneration of ECM and the apoptosis
of NP cells serve an important role in the basic pathological
mechanisms of IDD. Due to the vital role of IDD in low back pain,
therapeutic approaches that may postpone or even reverse the IDD
process are urgently required. AG, an important component of A.
paniculata, has been confirmed to exhibit potential
anti-inflammatory and protective effects in infections, tumors and
other diseases (24,26). In addition, it has been also
suggested that AG possesses therapeutic properties in IL-1β-induced
osteoarthritis (29), and has the
ability to attenuate abnormal apoptosis under pathological
conditions (30,31). In the present study, the primary
results indicated that AG treatment effectively mitigated the
IL-1β-induced inflammatory and apoptosis effects on the
degeneration of human NP cells. The primary mechanism of function
is the suppression of the expression of certain MMPs and ADAMTS,
and attenuation of the degradation of aggrecan and collagen-II.
Additionally, the inflammatory cytokine-mediated changes in
apoptosis-associated proteins were modulated by AG. The present
study also indicated that the molecular mechanisms of AG treatment
may be partly associated with the TLR4/MyD88/NF-κB signaling
pathway.
It has been established that IDD is primarily
associated with the inflammatory response (35), and inflammatory cytokines have been
suggested to have a vital role in the pathological processes of
osteoarthritis and IDD. It has been well documented that the
expression of inflammatory mediators in degraded tissue is
increased compared with normal tissues (36,37).
IL-1β is a key inflammatory cytokine, which may significantly
induce the occurrence of IDD (38). Concomitantly, NP cell degradation
may also be mitigated by the inhibition of IL-1β (39). Therefore, in the present study, the
pathophysiology of IDD was mimicked by stimulation with IL-1β.
Apoptosis, also termed type I programmed cell death,
is involved in a number of physiological processes. However,
uncontrolled apoptosis may be detrimental and serve a causative
factor in certain diseases, including IDD (15). Abnormal mechanical stress,
oxidative stress, cytokines and nitric oxide may all induce
apoptosis of intervertebral disc cells, in particular NP cells
(40,41). Adequate, viable NP cells form the
physiological basis of intervertebral discs; conversely, the
excessive apoptosis of NP cells will lead to the development of IDD
(16). The data from the present
study suggested that IL-1β accelerated the apoptosis of NP cells,
but this effect was significantly inhibited by AG, which indicated
that AG may mitigate IL-1β-induced NP cell degeneration by
ameliorating the increased levels of apoptosis.
The normal physiological function of intervertebral
discs is also considered to depend on the homeostasis between ECM
catabolic and anabolic activities, and disturbance of this balance
will result in the occurrence of degeneration (8). The ECM of NP cells is primarily
composed of aggrecan and collagen-II, and it performs essential
functions in intervertebral discs, including absorption of
nutrition, maintenance of disc height and withstanding mechanical
load (8,42). In the present study, following
treatment with IL-1β, the levels of aggrecan and collagen-II
decreased significantly; this decrease was suppressed by AG. From a
molecular perspective, the MMPs and ADAMTS families are two major
groups of catabolic enzymes involved in ECM regulation. MMPs have
been demonstrated as vital proteases in the progression of the
irreversible breakdown of collagen-II and proteoglycans, with a
high expression in degenerative disc tissues. It was also suggested
that ADAMTS-4 and ADAMTS-5 are 2 primary proteolytic enzymes
responsible for the cleavage of aggrecan. Furthermore, certain
studies provided evidence that inhibiting the expression and
activity of MMPs and ADAMTS may prevent the progression of IDD
(43). For example, Zhongyi et
al (19) identified that
IL-1β-dependent gene upregulation of MMP-3, MMP-9, MMP-13, ADAMTS-4
and ADAMTS-5 was significantly decreased by NF-κB inhibition.
Therefore, the inhibition of MMPs and ADAMTS may be considered
potential therapeutic targets for treating IDD. In addition, TIMPs,
the endogenous inhibitors of MMPs, also participate in the
maintenance of ECM homeostasis. Among TIMPs, TIMP-1 is closely
associated with MMP-3 and MMP-13, and downregulates their
activities (14). Therefore,
TIMP-1 is also considered to have an inhibitory effect on IDD. In
the present study, it was identified that IL-1β upregulated the
expression of MMP-3, MMP-9, MMP-13, ADAMTS-4, and ADAMTS-5;
concomitantly, TIMP-1 exhibited a marked decrease. However, AG
treatment significantly suppressed these IL-1β-induced changes in
the ECM and metabolic enzymes in NP cells. These changes suggested
that AG inhibited IL-1β-induced NP cells degeneration via
decreasing the level of ECM degeneration and suppressing the
expression these catabolic enzymes.
The NF-κB signaling pathway is known for its crucial
regulation in a series of catabolic processes active in response to
inflammation, stress, and cellular damage (17,19).
For example, following stimulation with IL-1β, the inactive NF-κB
combined with the inhibitory protein NF-κB inhibitor α may be
activated and released, subsequently translocated from the
cytoplasm into the nucleus, and finally activate the transcription
of its target genes, including MMPs (44). It has been demonstrated that the
activation of the NF-κB signaling pathway contributes to ECM
degradation by increasing the activity of matrix-degrading enzymes
in the NP cells (19). Therefore,
the targeted inhibition of NF-κB may be a critical therapeutic
target for IDD. Additionally, The p65 binding site has also been
identified to be in the promoter regions of several MMP genes
(45). Therefore, in the present
study, it was determined whether the anti-inflammatory effects of
AG against ECM degradation functioned through NF-κB signaling
pathways by investigating the changes in p65 and nuclear
translocation. Notably, the IL-1β-induced phosphorylation of p65
and nuclear translocation were significantly inhibited by AG. These
results were consistent with Peng et al (46), who identified that AG markedly
decreased the p65 phosphorylation level following ovalbumin
stimulation. The TLR4/MyD88 signaling pathway is also a pivotal
pathway involved in inflammation response (20,21),
which is considered to function in conjunction with NF-κB signaling
pathway (22–24). The TLRs are a family of receptor
proteins used by the innate immune system in mammals; activation of
TLRs is involved in the production of a number of proinflammatory
cytokines. MyD88 is a signal adaptor molecule with roles in
signaling via the TLRs, including TLR4 (47). The activation of the TLR4/MyD88
pathway is considered as an activating factor for the NF-κB
signaling pathway (23,24). The results of the present study
demonstrated that the IL-1β-mediated upregulation of TLR4 and MyD88
was inhibited by AG treatment, which was consistent with the
changes of p65 observed. Taken together, these data suggest that
the inhibition of the IL-1β-induced inflammatory response by AG may
be partly associated with TLR4/MyD88/NF-κB signaling pathway. It
should be also noted that additional studies, which reconfirm this
mechanism by using gene knockout mice, are expected to clarify this
issue.
In conclusion, the data from the present study
revealed that AG may alleviate IL-1β-induced human NP cells
apoptosis. Furthermore, AG may also attenuate IL-1β-induced
degeneration of the ECM, and the expression of MMPs and ADAMTS via
inhibiting the TLR4/MyD88/NF-κB signaling pathway. Therefore, AG
may be a potential agent for IDD prevention and treatment. However,
the exact mechanism of AG-based regulation of inflammation in NP
cells remains unclear, and additional studies are required.
Acknowledgements
The authors would like to thank the Laboratory of
Orthopedics and Scientific Research Center of Second Affiliated
Hospital of Wenzhou Medical University (Zhejiang, China).
Funding
The present study was supported by Zhejiang Province
Medical Science and Technology Project (grant no. 2017171281) and
the Wenzhou Bureau of Science and Technology Project (grant no.
Y20160136).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LZ and SS conceived and designed the experiments.
LZ, QC and JY performed the experiments and analyzed the data. HW
prepared and assessed the figures. SS provided guidance for
experiments. LZ was primarily responsible for the preparation of
the manuscript. SS contributed to revision of the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Second Affiliated Hospital and Yuying Children's
Hospital of Wenzhou Medical University (Zhejiang, China) and the
patients provided written informed consent.
Patient consent for publication
The patients provided written informed consent.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
IDD
|
intervertebral disc degeneration
|
|
NP
|
nucleus pulposus
|
|
ECM
|
extracellular matrix
|
|
IL-1β
|
interleukin-1β
|
|
MMP
|
matrix metalloproteinase
|
|
ADAMTS
|
a disintegrin and metalloproteinase
with thrombospondin motifs
|
|
TLRs
|
toll-like receptors
|
|
MyD88
|
myeloid differentiation primary
response protein MyD88
|
|
NF-κB
|
nuclear factor
kappa-light-chain-enhancer of activated B cells
|
|
TIMPs
|
tissue inhibitors of
metalloproteinases
|
|
AG
|
andrographolide
|
References
|
1
|
Luoma K, Riihimäki H, Luukkonen R,
Raininko R, Viikari-Juntura E and Lamminen A: Low back pain in
relation to lumbar disc degeneration. Spine (Phila Pa 1976).
25:487–492. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Livshits G, Popham M, Malkin I, Sambrook
PN, Macgregor AJ, Spector T and Williams FM: Lumbar disc
degeneration and genetic factors are the main risk factors for low
back pain in women: The UK Twin Spine Study. Ann Rheum Dis.
70:1740–1745. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Martin BI, Deyo RA, Mirza SK, Turner JA,
Comstock BA, Hollingworth W and Sullivan SD: Expenditures and
health status among adults with back and neck problems. JAMA.
299:656–664. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lotz JC and Ulrich JA: Innervation,
inflammation, and hypermobility may characterize pathologic disc
degeneration: Review of animal model data. J Bone Joint Surg Am. 88
Suppl 2:76–82. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Vo N, Niedernhofer LJ, Nasto LA, Jacobs L,
Robbins PD, Kang J and Evans CH: An overview of underlying causes
and animal models for the study of age-related degenerative
disorders of the spine and synovial joints. J Orthop Res.
31:831–837. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Froud R, Patterson S, Eldridge S, Seale C,
Pincus T, Rajendran D, Fossum C and Underwood M: A systematic
review and meta-synthesis of the impact of low back pain on
people's lives. BMC Musculoskel Dis. 15:502014. View Article : Google Scholar
|
|
7
|
Vergroesen PP, Kingma I, Emanuel KS,
Hoogendoorn RJ, Welting TJ, van Royen BJ, van Dieën JH and Smit TH:
Mechanics and biology in intervertebral disc degeneration: A
vicious circle. Osteoarthritis Cartilage. 23:1057–1070. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kepler CK, Ponnappan RK, Tannoury CA,
Risbud MV and Anderson DG: The molecular basis of intervertebral
disc degeneration. Spine J. 13:318–330. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hangai M, Kaneoka K, Kuno S, Hinotsu S,
Sakane M, Mamizuka N, Sakai S and Ochiai N: Factors associated with
lumbar intervertebral disc degeneration in the elderly. Spine J.
8:732–740. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pockert AJ, Richardson SM, Le Maitre CL,
Lyon M, Deakin JA, Buttle DJ, Freemont AJ and Hoyland JA: Modified
expression of the ADAMTS enzymes and tissue inhibitor of
metalloproteinases 3 during human intervertebral disc degeneration.
Arthritis Rheum. 60:482–491. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bachmeier BE, Nerlich A, Mittermaier N,
Weiler C, Lumenta C, Wuertz K and Boos N: Matrix metalloproteinase
expression levels suggest distinct enzyme roles during lumbar disc
herniation and degeneration. Eur Spine J. 18:1573–1586. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Roberts S, Caterson B, Menage J, Evans EH,
Jaffray DC and Eisenstein SM: Matrix metalloproteinases and
aggrecanase: Their role in disorders of the human intervertebral
disc. Spine (Phila Pa 1976). 25:3005–3013. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Le Maitre CL, Freemont AJ and Hoyland JA:
Localization of degradative enzymes and their inhibitors in the
degenerate human intervertebral disc. J Pathol. 204:47–54. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang L, Wu Y, Tan Y, Fei X, Deng Y, Cao H,
Chen B, Wang H, Magdalou J and Chen L: Cytotoxic effects of the
quinolone levofloxacin on rabbit meniscus cells. J Appl Toxicol.
34:870–877. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhou GQ, Yang F, Leung VVL and Cheung KMC:
Molecular and cellular biology of the intervertebral disc and the
use of animal models. Curr Orthopaed. 22:267–273. 2008. View Article : Google Scholar
|
|
16
|
Yang SD, Yang DL, Sun YP, Wang BL, Ma L,
Feng SQ and Ding WY: 17β-estradiol protects against apoptosis
induced by interleukin-1β in rat nucleus pulposus cells by
down-regulating MMP-3 and MMP-13. Apoptosis. 20:348–357. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Baker RG, Hayden MS and Ghosh S: NF-κB,
inflammation, and metabolic disease. Cell Metab. 13:11–22. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wuertz K, Vo N, Kletsas D and Boos N:
Inflammatory and catabolic signalling in intervertebral discs: The
roles of NF-κB and MAP kinases. Eur Cells Mater. 23:103–119;
discussion 119–119. 2012.
|
|
19
|
Zhongyi S, Sai Z, Chao L and Jiwei T:
Effects of nuclear factor kappa B signaling pathway in human
intervertebral disc degeneration. Spine (Phila Pa 1976).
40:224–232. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mulla MJ, Brosens JJ, Chamley LW, Giles I,
Pericleous C, Rahman A, Joyce SK, Panda B, Paidas MJ and Abrahams
VM: Antiphospholipid antibodies induce a pro-inflammatory response
in first trimester trophoblast via the TLR4/MyD88 pathway. Am J
Reprod Immunol. 62:96–111. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kyo F, Futani H, Matsui K, Terada M,
Adachi K, Nagata K, Sano H, Tateishi H, Tsutsui H and Nakanishi K:
Endogenous interleukin-6, but not tumor necrosis factor alpha,
contributes to the development of toll-like receptor 4/myeloid
differentiation factor 88-mediated acute arthritis in mice.
Arthritis Rheum. 52:2530–2540. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Biragyn A, Coscia M, Nagashima K, Sanford
M, Young HA and Olkhanud P: Murine beta-defensin 2 promotes
TLR-4/MyD88-mediated and NF-kappaB-dependent atypical death of APCs
via activation of TNFR2. J Leukoc Biol. 83:998–1008. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vijayan V, Khandelwal M, Manglani K, Gupta
S and Surolia A: Methionine down-regulates TLR4/MyD88/NF-κB
signalling in osteoclast precursors to reduce bone loss during
osteoporosis. Br J Pharmacol. 171:107–121. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang QQ, Ding Y, Lei Y, Qi CL, He XD, Lan
T, Li JC, Gong P, Yang X, Geng JG and Wang LJ: Andrographolide
suppress tumor growth by inhibiting TLR4/NF-κB signaling activation
in insulinoma. Int J Biol Sci. 10:404–414. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sheeja K and Kuttan G: Activation of
cytotoxic T lymphocyte responses and attenuation of tumor growth in
vivo by Andrographis paniculata extract and andrographolide.
Immunopharmacol Immunotoxicol. 29:81–93. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wintachai P, Kaur P, Lee RC, Ramphan S,
Kuadkitkan A, Wikan N, Ubol S, Roytrakul S, Chu JJ and Smith DR:
Activity of andrographolide against chikungunya virus infection.
Sci Rep. 5:141792015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chiou WF, Chen CF and Lin JJ: Mechanisms
of suppression of inducible nitric oxide synthase (iNOS) expression
in RAW 264.7 cells by andrographolide. Br J Pharmacol.
129:1553–1560. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lu WJ, Lin KH, Hsu MJ, Chou DS, Hsiao G
and Sheu JR: Suppression of NF-κB signaling by andrographolide with
a novel mechanism in human platelets: Regulatory roles of the p38
MAPK-hydroxyl radical-ERK2 cascade. Biochem Pharmacol. 84:914–924.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ding QH, Ji XW, Cheng Y, Yu YQ, Qi YY and
Wang XH: Inhibition of matrix metalloproteinases and inducible
nitric oxide synthase by andrographolide in human osteoarthritic
chondrocytes. Mod Rheumatol. 23:1124–1132. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Burgos RA, Seguel K, Perez M, Meneses A,
Ortega M, Guarda MI, Loaiza A and Hancke JL: Andrographolide
inhibits IFN-gamma and IL-2 cytokine production and protects
against cell apoptosis. Planta Med. 71:429–434. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang J, Zhu D, Wang Y and Ju Y:
Andrographolide attenuates LPS-induced cardiac malfunctions through
inhibition of IκB phosphorylation and apoptosis in mice. Cell
Physiol Biochem. 37:1619–1628. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pfirrmann CW, Metzdorf A, Zanetti M,
Hodler J and Boos N: Magnetic resonance classification of lumbar
intervertebral disc degeneration. Spine (Phila Pa 1976).
26:1873–1878. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim J, Choi Y, Ahn M, Jung K and Shin T:
Olfactory dysfunction in autoimmune central nervous system
neuroinflammation. Mol Neurobiol. 55:8499–8508. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Risbud MV and Shapiro IM: Role of
cytokines in intervertebral disc degeneration: Pain and disc
content. Nat Rev Rheumatol. 10:44–56. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hoyland JA, Le Maitre C and Freemont AJ:
Investigation of the role of IL-1 and TNF in matrix degradation in
the intervertebral disc. Rheumatology (Oxford). 47:809–814. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Smith LJ, Chiaro JA, Nerurkar NL, Cortes
DH, Horava SD, Hebela NM, Mauck RL, Dodge GR and Elliott DM:
Nucleus pulposus cells synthesize a functional extracellular matrix
and respond to inflammatory cytokine challenge following long-term
agarose culture. Eur Cells Mater. 22:291–301. 2011. View Article : Google Scholar
|
|
38
|
Le Maitre CL, Freemont AJ and Hoyland JA:
The role of interleukin-1 in the pathogenesis of human
intervertebral disc degeneration. Arthritis Res Ther. 7:R732–R745.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lee JM, Song JY, Baek M, Jung HY, Kang H,
Han IB, Kwon YD and Shin DE: Interleukin-1β induces angiogenesis
and innervation in human intervertebral disc degeneration. J Orthop
Res. 29:265–269. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang CC, Cui GP, Hu JG, Xiao YZ, Zhou XS,
Shao C, Lin Q and Zhou JS: Effects of adenoviral vector expressing
hIGF-1 on apoptosis in nucleus pulposus cells in vitro. Int J Mol
Med. 33:401–405. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yang D, Wang D, Shimer A, Shen FH, Li X
and Yang X: Glutathione protects human nucleus pulposus cells from
cell apoptosis and inhibition of matrix synthesis. Connect Tissue
Res. 55:132–139. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Feng H, Danfelter M, Strömqvist B and
Heinegård D: Extracellular matrix in disc degeneration. J Bone
Joint Surg Am. 88 Suppl 2:25–29. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Vo NV, Hartman RA, Yurube T, Jacobs LJ,
Sowa GA and Kang JD: Expression and regulation of
metalloproteinases and their inhibitors in intervertebral disc
aging and degeneration. Spine J. 13:331–341. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yang JP, Hori M, Sanda T and Okamoto T:
Identification of a novel inhibitor of nuclear factor-kappaB,
RelA-associated inhibitor. J Biol Chem. 274:15662–15670. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Frank S, Peters MA, Wehmeyer C, Strietholt
S, Koers-Wunrau C, Bertrand J, Heitzmann M, Hillmann A, Sherwood J,
Seyfert C, et al: Regulation of matrixmetalloproteinase-3 and
matrixmetalloproteinase-13 by SUMO-2/3 through the transcription
factor NF-κB. Ann Rheum Dis. 72:1874–1881. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Peng S, Gao J, Liu W, Jiang C, Yang X, Sun
Y, Guo W and Xu Q: Andrographolide ameliorates OVA-induced lung
injury in mice by suppressing ROS-mediated NF-κB signaling and
NLRP3 inflammasome activation. Oncotarget. 7:80262–80274. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yamamoto M, Sato S, Hemmi H, Hoshino K,
Kaisho T, Sanjo H, Takeuchi O, Sugiyama M, Okabe M, Takeda K and
Akira S: Role of adaptor TRIF in the MyD88-independent toll-like
receptor signaling pathway. Science. 301:640–643. 2003. View Article : Google Scholar : PubMed/NCBI
|