Introduction
Pelvic organ prolapse (POP) is an increasingly
serious health problem that impairs quality of life with a diverse
clinical spectrum, including herniation of the uterus, bladder
and/or rectum. This condition occurs with a high prevalence in
postmenopausal women, with a lifetime risk of surgery for POP or
urinary incontinence of 11% (1).
This figure is expected to increase with the aging society.
Currently, despite the growing prevalence of POP, no
consensus exists among researchers regarding its etiology and
pathogenesis. There is no doubt, however, that it is a
multifactorial disorder associated with a genetic predisposition.
In addition, multiple additive genetic, environmental and lifestyle
factors contribute to the pathogenesis of POP. Epidemiological
studies have identified several risk factors, including age
(2), race (3,4),
parity (5,6) and obesity (5,7,8).
However, these factors fail to explain the pathogenesis of POP in
nulliparous women and to answer the question of why POP does not
develop in all women exhibiting high-risk factors.
Recently several gene arrays for POP were revealed.
Differential expression signatures were identified in 81 genes and
specific extracellular matrix (ECM)-associated genes may
participate in the pathology of POP, according to RNA-Seq data
(9). Ak et al (10) reported, for the first time, that
certain genes serve a role in the cell cycle, proliferation and
embryonic development, along with cell adhesion processes during
the development of POP, using gene chip microarrays, and a
genome-wide association study (11) identified promising single
nucleotide polymorphisms associated with POP. Recently published
genome-wide linkage analysis (12)
provided evidence for two additional loci in relation to
symptomatic POP and whole-exome sequencing identified a novel gene,
WNK1, that influences susceptibility to POP (13).
Generally, epigenetic regulation of gene
transcription occurs by three main mechanisms: DNA methylation,
histone modification and microRNA (miRNA) expression (14). DNA methylation, the most common
epigenetic mechanism, leads to changes in gene expression without
alteration of the DNA sequence. Aberrant (hyper or hypo-)
methylation is believed to be greatly influenced by environmental
risk factors. Klutke et al (15) first reported that methylation in
the promoter region may suppress lysyl oxidase (LOX) gene
expression in women with POP, but the DNA methylome of POP has
never been characterized.
Since the uterosacral ligaments (ULs) provide
primary support for the uterus and the upper vagina, it was
hypothesized that the disruption of these ligaments may lead to a
loss of support and eventually contribute to POP. In the present
study, whether there is any aberrant methylation in the ULs of
patients with POP compared to the controls was investigated.
Materials and methods
Tissue collection
Approval from the institutional review board was
obtained from the Beijing Obstetrics and Gynecology Hospital Ethics
Committee. Informed consent was obtained from all individual
participants included in the study. A total of nine postmenopausal
women, with five POP and four non-POP controls, undergoing
hysterectomy for benign conditions were included, from January 2015
to June 2017. The clinicopathological characteristics of these
patients are presented in Table I.
In order to eliminate the intermixing factors between the
experimental group and the control group, strict limits on
inclusion and exclusion criteria were set for the uterine ligament
samples. Exclusion criteria were as follows: Women with a history
of connective tissue disorders, endometriosis, prior pelvic
reconstructive surgery and cancer. Inclusion in the POP group
required uterine prolapse beyond the hymen (stage 3 or stage 4)
with/without cystocele and/or rectocele. Patient characteristics
assessed included: Age, parity, body mass index (BMI), menopause,
duration of menopausal period, history of hormone replacement
therapy (HRT), smoking history and history of hypertension. The ULs
were obtained during the procedures, providing ~1 g of tissue per
sample.
 | Table I.Clinical characteristics. |
Table I.
Clinical characteristics.
|
Characteristics | POP (n=5) | CON (n=4) | P-value |
|---|
| Age (years) |
|
|
|
|
Range |
60.60±5.23 (55–66) |
61.25±6.65 (53–68) | NS |
| BMI
(kg/m2) | 24.84±2.17 | 26.80±5.74 | NS |
| Pregnancy (n) |
|
|
|
|
Range |
3.6±1.14 (2–5) | 2.75±0.05
(2–3) | NS |
| Parity (n) | 2.00±0.00 | 1.75±0.50 | NS |
|
Patients (n) without vaginal
delivery | 0 | 0 | – |
|
Patients (n) one vaginal
delivery | 0 | 1 | – |
|
Patients (n) two vaginal
deliveries | 5 | 3 | – |
| Menopause (n) | 5 | 4 | – |
| During of
menopausal period (years) |
|
|
|
|
Range | 10.20±4.82
(5–16) | 11.00±7.79
(4–20) | NS |
| Smoking (n) | 0 | 0 | – |
| Suffering from
hypertension (n) | 1 | 2 | – |
DNA extraction
DNA was isolated using an OMEGA TISSUE DNA kit
(Omega Bio-Tek, Inc., Norcross, GA, USA), according to the
manufacturer's protocol. DNA was quantified by spectrophotometry
(NanoDrop; Thermo Fisher Scientific, Inc., Wilmington, DE, USA).
Genomic DNA (500 ng) was treated with bisulfate using an EZ DNA
Methylation Gold kit (Zymo Research Corps., Irvine, CA, USA),
according to the manufacturer's protocol.
Microarray experiments
Microarray experiment correction for multiple
testing was done in order to separate the most significant
differences from the background noise. Prior to microarray
experiments, clustering test was done between the two groups. POP
and control subjects were separated (Data not shown). The
methylation of DNA was assayed on an Illumina Infinium Methylation
EPICBeadChip (Illumina, Inc., San Diego, CA, USA) using the
Illumina HD methylation assay kit from Shanghai Biotechnology
Corporation (Shanghai, China). Each of these arrays contained
853,307 probes, covered CpG islands, RefSeq genes, ENCODE open
chromatin, ENCODE transcription factor binding sites and FANTOM5
enhancers. Bisulfite-converted DNA was analyzed on an EPICBeadChip
following the manufacturer's protocol. GenomeStudio methylation
module V.1.9.0 (Illumina, Inc.) was used to extract the image
intensities.
Statistical analysis and microarray
data analysis
Clinical characteristics were analyzed using SPSS
software version 17.0 (SPSS, Inc., Chicago, IL, USA). Statistical
significance for the t-test was determined to be at the level of
P<0.05. Values are expressed as the mean and standard
deviation.
The CpG probe signal intensities were normalized
using Subset-Quantile Within Array Normalization in the minfi
packages from R Bioconductor (16)
for background correction and subtraction. Methylation values for
individual CpG sites were obtained as β-values, calculated as the
ratio of the methylated signal intensity to the sum of the
methylated and unmethylated signals following background
subtraction. The β-values were reported as a DNA methylation score
ranging from 0 (non-methylated) to 1 (completely methylated).
Initially, probes located on the sex chromosome were excluded.
Differentially methylated CpGs were selected using an algorithm in
IMA Bioconductor. In the present study, the mean-difference β-value
(Δβ) between the two sample groups for each CpG site was assessed.
Specifically, a probe was considered to be differentially
methylated if the absolute Δβ was higher than 0.14 and the
statistical test was significant (P<0.05 was considered to
indicate a statistically significant difference). The interaction
network was generated using the Search Tool for the Retrieval of
Interacting Genes (STRING) database (string-db.org)
to analyze genes from the experiments and text mining.
Results
Clinical characteristics of all
participants
A total of 9 subjects were recorded, including five
patients with POP and 4 controls. ULs were obtained from a total of
9 hysterectomies. The overall grade of prolapse was 3–4 and the
degree of uterine prolapse was grade 3–4 beyond the hymen. Clinical
characteristics of all participants included in the study are
presented in Table I.
Differential methylation and
expression profiling in POP
DNA methylation analysis identified 3,723
differentially methylated CpG sites; 0.4% of the total sites in POP
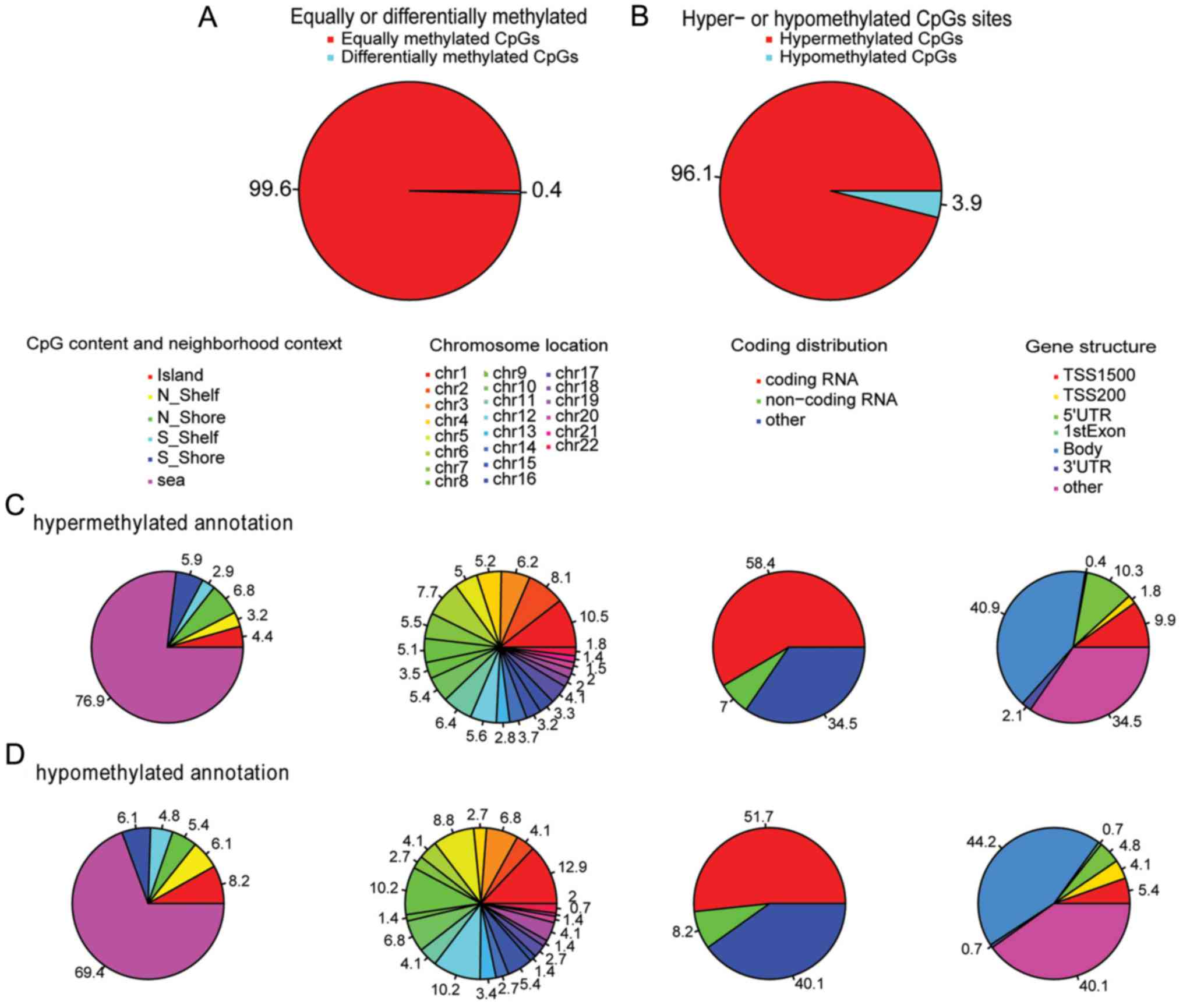
ULs were detected, compared to the controls (Fig. 1A). Surprisingly, the
hypermethylated CpG site accounted for ~96.1% (3,576), which was
markedly higher than the number of hypomethylated ones (147 CpG
sites, 3.9%, Fig. 1B). Notably, in
methylated CpG content and neighborhood context (Shore: 0–2 k
distance around the island; Shelf: 2 to 4 k distance around the
island), there was a difference between hypermethylation and
hypomethylation; hypermethylated CpG site percentages were 4.4,
3.2, 6.8, 2.9, 5.9 and 76.9% in Island, N-Shelf, N-Shore, S-Shore,
S-Shelf and Open Sea, respectively. However, hypomethylated CpG
site percentages were 8.2, 6.1, 5.4, 4.8, 6.1 and 69.4% in Island,
N-Shelf, N-Shore, S-Shore, S-Shelf and Open Sea, respectively. In
particular, more hypomethylated CpG sites were detected in Island
and N-Shelf (Fig. 1C and D).
Hypermethylated and hypomethylated CpG sites are presented in
Fig. 1C and D in chromosomal
locations. Finally, probes from the X and Y chromosomes were
excluded due to the partial homology of the X and Y chromosomes,
and the fact that the 9 samples were females. As for gene
structure, there was significant difference (TSS1500, TSS200,
5′UTR, 1st Exon, Body, 3′UTR and others) between hypermethylation
and hypomethylation; in particular, there was greater
hypermethylation residing in TSS1500 and 5′UTR compared with
hypomethylation. Detailed differences in gene structure are
presented in Fig. 1C and D.
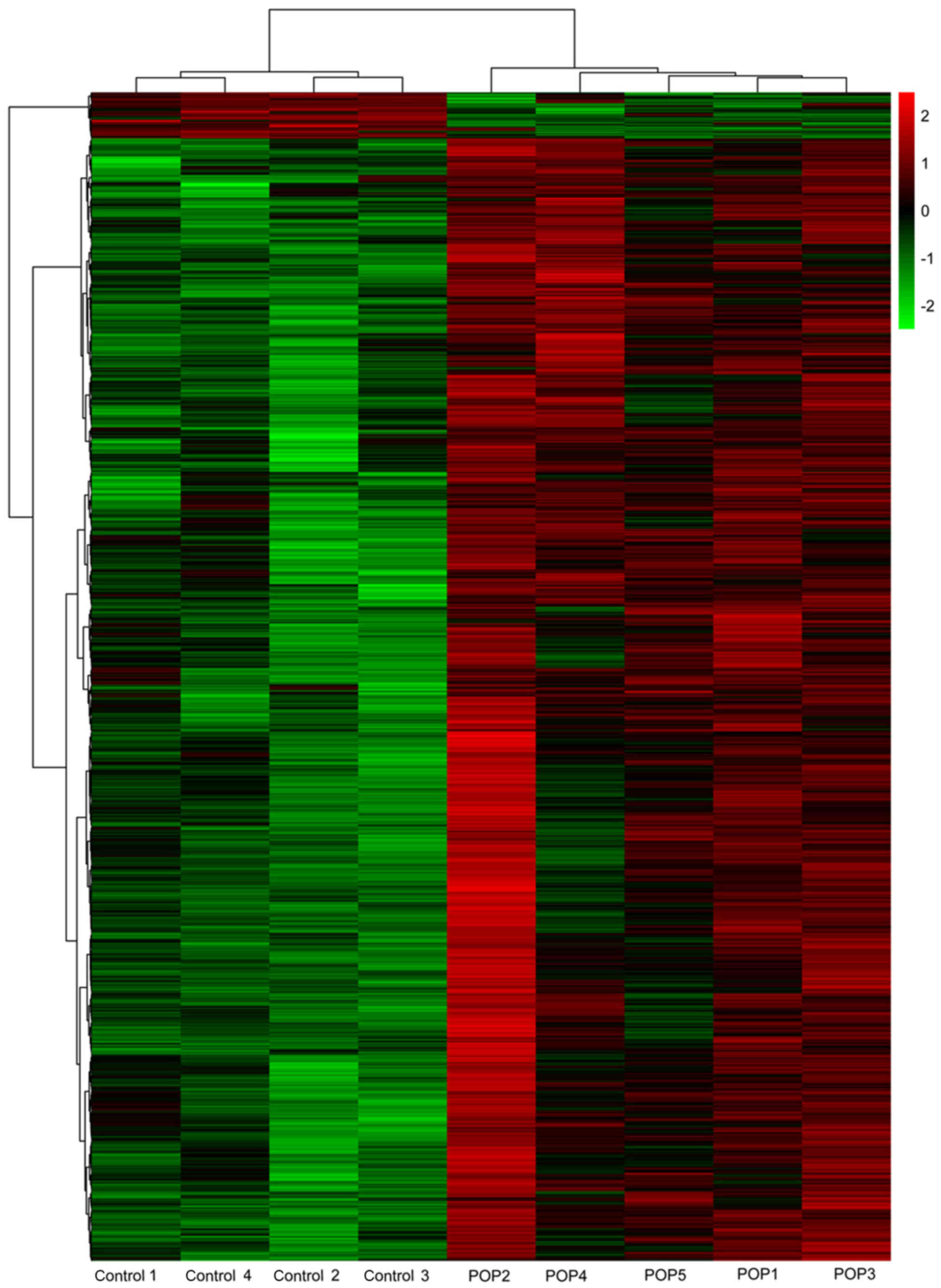
Heat maps of differential methylation are presented
in Fig. 2. Total methylation of
POP samples was markedly different from that of the controls;
however, in the POP group, samples POP4 and POP5 were more similar
to each other, and were slightly different from the other three POP
samples.
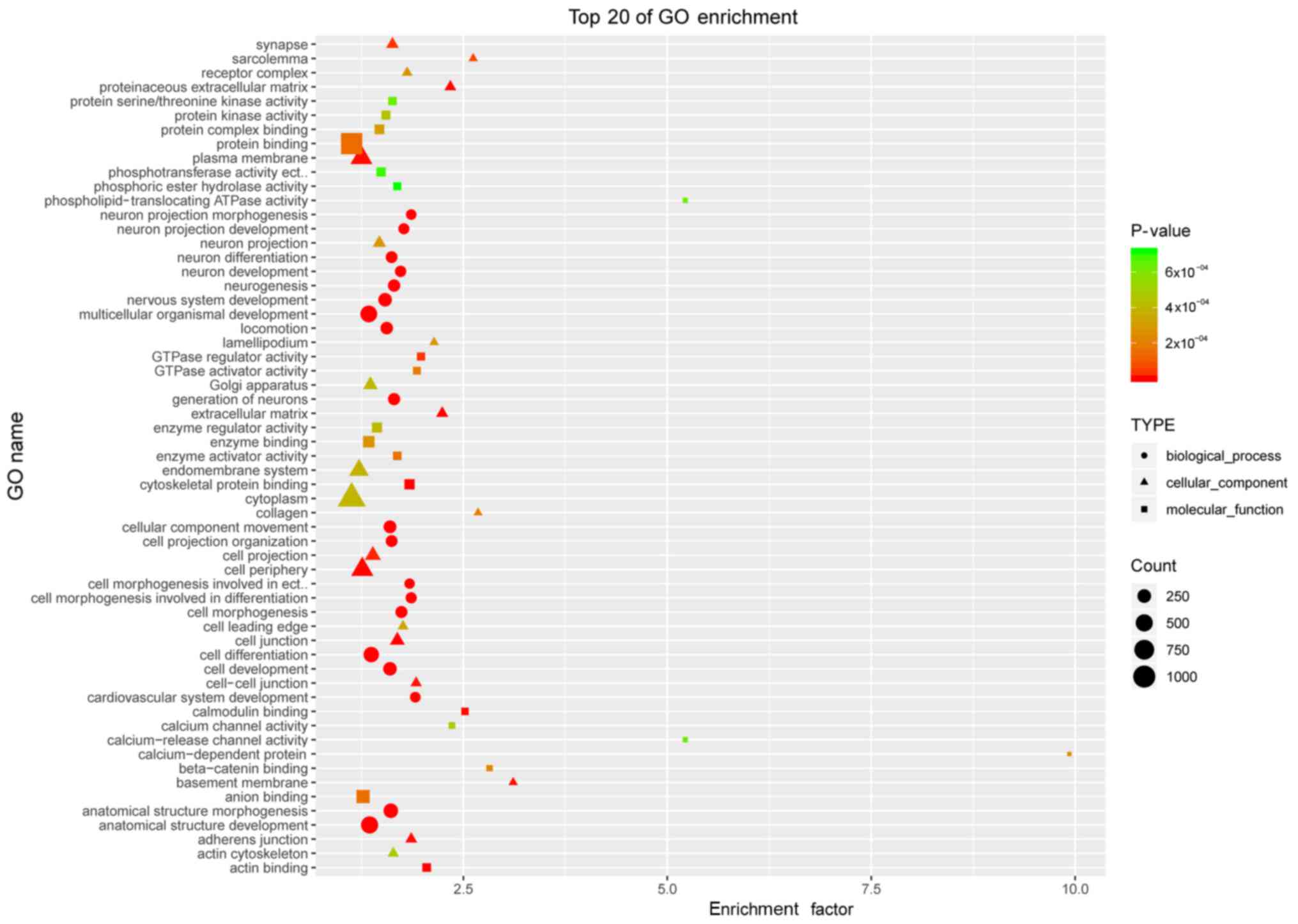
GO analysis of the differentially
methylated genes in POP
To begin defining the functional significance of the
extensive changes in DNA methylation in POP, GO analysis was used.
According to the functions of the differential genes, the top 20 in
each category were listed, respectively, in molecular function,
biological process and cellular component (Fig. 3). These differentially methylated
genes were widely associated with cell morphogenesis, the
extracellular matrix, cell junctions, protein binding and guanosine
triphosphatase (GTPase) activity.
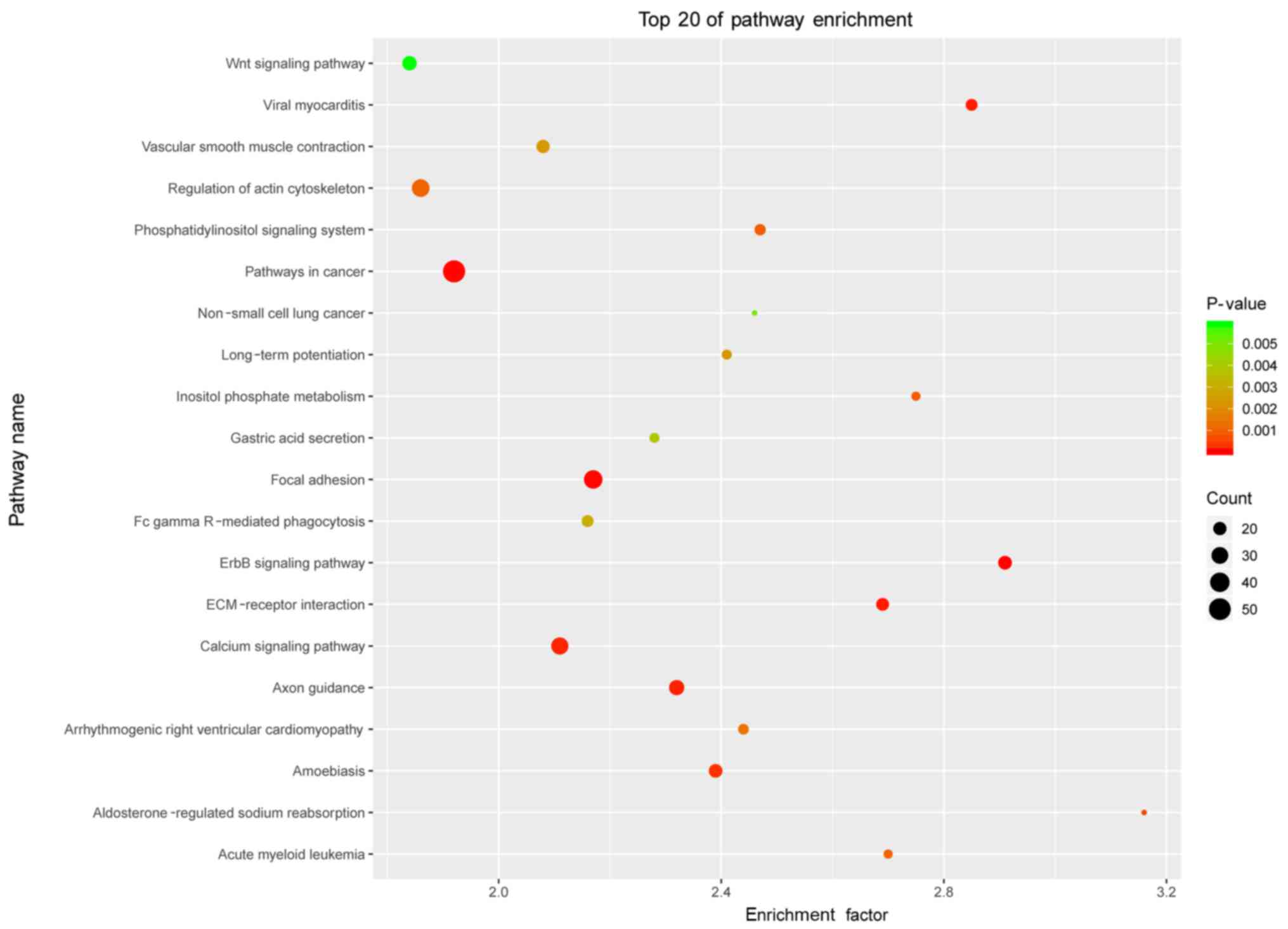
KEGG analysis of the differentially
methylated genes in POP
To further investigate key pathways associated with
these distinct genes, the interaction network of the significant
pathways associated with POP was built according to the KEGG
database. The analysis in the present study demonstrated that these
differentially methylated genes were widely involved in various
cellular pathways. There were a total of 206 pathways associated
with POP and the top 20 included ‘ErbB signaling pathway’,
‘pathways in cancer’, ‘focal adhesion’ and ‘ECM-receptor
interaction’ (presented in Fig.
4). Notably, ‘focal adhesion’ and ‘ECM-receptor interaction’
were enriched in the top 3 and top 4 of differentially methylated
profiling in the top 20 enriched pathways. The two pathways of the
differentially methylated genes are listed in Table II.
| Table II.KEGG analysis of distinct methylated
genes in pathways. KEGG analysis of distinct hepermethylated genes
and hypomethylated genes in focal adhesion and ECM-receptor
interaction pathways. |
Table II.
KEGG analysis of distinct methylated
genes in pathways. KEGG analysis of distinct hepermethylated genes
and hypomethylated genes in focal adhesion and ECM-receptor
interaction pathways.
| Pathway ID | Description | P-value | Hypermethylated
Genes | Hypomethylated
Genes |
|---|
| hsa04510 | Focal adhesion |
3.38×10−05 | COL5A1 LAMB1 ITGA9
PDK1 FYN VWF AKT3 COL1A2 VEGFC MAPK9 ZYX BCAR1 CRK PARVB PIK3R1
PIK3CB THBS2 FN1 ITGAV COL11A2 PRKCA LAMB3 SOS1 CAPN2 LAMA2 DOCK1
RASGRF1 TNR COL6A3 PTK2 MYLK3 COL4A1 ITGB5 COL5A2 FIGF | ITGA8 |
| hsa04512 | ECM-receptor
interaction |
1.18×10−04 | COL5A1 LAMB1 SDC2
ITGA9 VWF COL1A2 THBS2 FN1 ITGAV COL11A2 LAMB3 LAMA2 TNR COL6A3
COL4A1 ITGB5 CD44 COL5A2 | ITGA8 |
Network analysis in POP
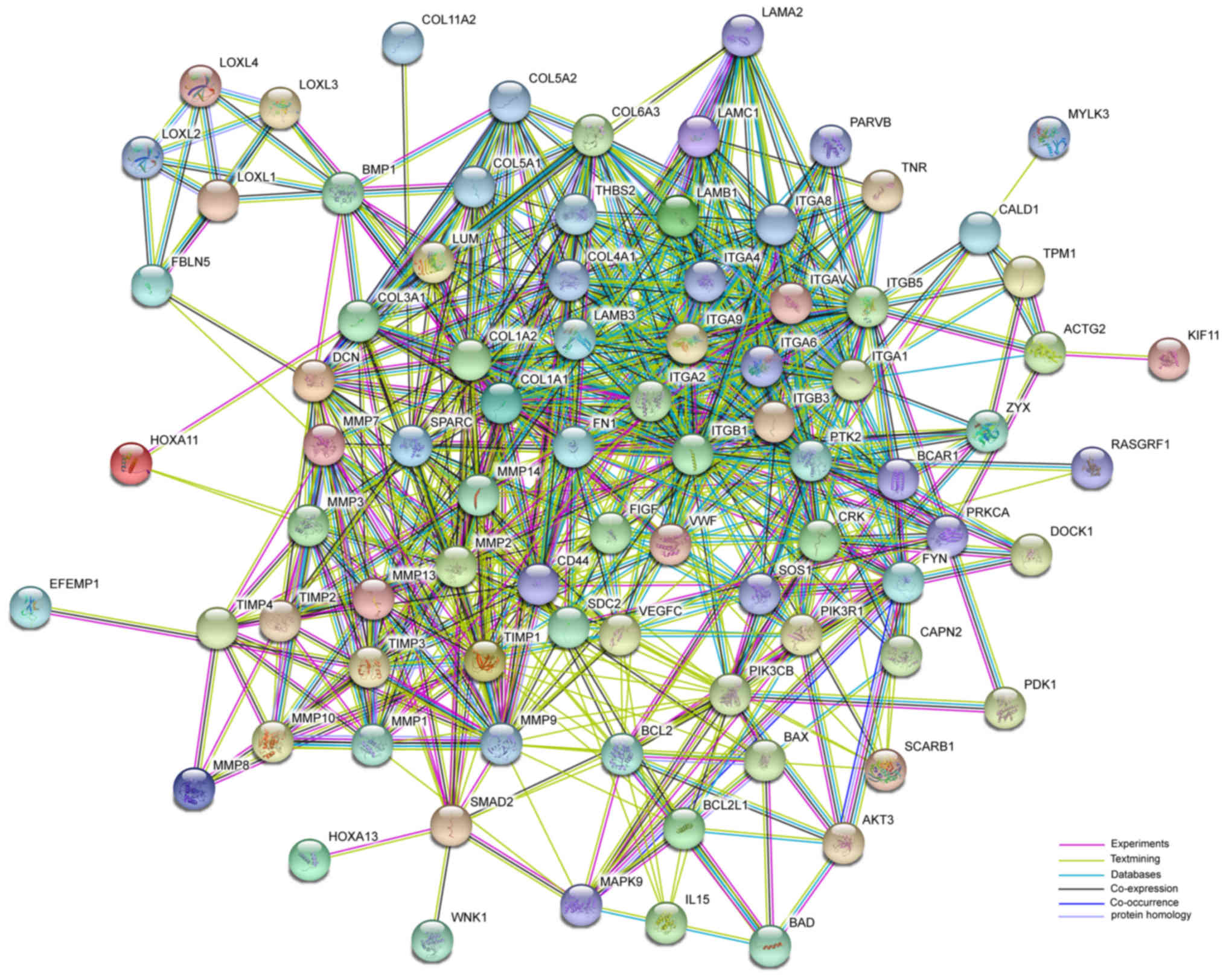
The STRING was used to provide information regarding
predicted and experimental interactions of proteins. Differentially
expressed genes were demonstrated to be enriched via methylation
profiling and literature mining in the focal adhesion and
ECM-receptor interaction pathways (Fig. 5); furthermore, the nodes with
higher degrees of interaction were considered as hub nodes. The top
5 nodes were integrin (IGTB1; degree=43), fibronectin (FN1;
degree=39), protein tyrosine kinase (PTK2; degree=38), cluster of
differentiation (CD)44 (degree=37) and integrin (IGTA1;
degree=37).
Discussion
Emerging evidence indicates that epigenetic
mechanisms may partly contribute to the pathogenesis of POP
(15). However, until now there
had been no reports on genome-wide DNA methylation profiling in
POP, except the LOX methylation. Klutke et al (15) measured promoter methylation in the
LOX gene in women with POP and found a total of 66 methylated CpG
sites in the POP group and only one methylated CpG site in the
non-prolapse control group. In the present study, it was reported
that there were 3,723 differentially methylated CpG sites, 0.4% of
the total sites in POP ULs compared with the controls in menopausal
women.
In general, increased DNA methylation means higher
levels of gene expression (17).
Over the past decades, a number of studies have revealed that a
considerable percentage of CpG site methylation varies with age
(18), giving rise to genome-wide
hypomethylation with site-specific incidences of hypermethylation.
Notably, tumors have a unique methylation pattern with high levels
of hypomethylation (19). In the
present study, the five menopausal women with POP and the four
without POP demonstrated a unique methylation pattern with low
levels of hypomethylation, which may partly be associated with
aging. The age range of the nine subjects was between 53 and 68
years old and therefore all were menopausal women. As for the
sample limitations, age-associated variations in methylation should
be further investigated in the future.
DNA methylation is a key epigenetic process involved
in the regulation of gene expression. There is no doubt, however,
that POP is a multifactorial disorder with a genetic
predisposition, determined by interactions between additive
genetic, environmental and lifestyle factors. As presented in the
heat map, hypermethylation and hypomethylation were obviously
different between POPs and controls, demonstrating their
contribution to the pathogenesis of POP. Notably, with respect to
the methylation within the POP group, POP4 and POP5 were more
similar to each other and slightly different from the other three.
Analyzing the clinical characteristics, it was demonstrated that
the duration of menopause in the two cases was ~15 years, but it
was five, six and ten years in the other three. These two POP cases
were older than the other three also; there were no differences in
parity or BMI. Therefore, age and duration of menopause may be
highly associated with methylation.
For DNA methylation profiling, gene ontology
analysis was used to separately assess molecular function,
biological processes and cellular components. In biological
processes, the enriched terms were ‘anatomical structure
morphogenesis’, ‘anatomical structure development’, ‘cell
morphogenesis’, ‘locomotion involved in differentiation’, ‘cellular
component movement’ and ‘locomotion’, which was likely due to the
pelvic organs moving from their normal anatomical position,
accounting for prolapse. In the cellular component category, the
terms were ‘proteinaceous extracellular matrix’, ‘cell junction’,
‘cell periphery’, ‘adherens junction’, ‘basement membrane’,
‘cell-cell junction’ and ‘collagen’, indicating that POP may be
associated with the ECM. In the molecular function category, the
terms were ‘cytoskeletal protein binding’, ‘actin binding’,
‘calmodulin binding’, ‘GTPase regulator activity’, ‘calcium channel
activity’, ‘calcium-release channel activity’ and ‘protein
binding’. Similarly, previous evidence has demonstrated that
several families, including collagen (20–22),
LOX (15,23–25)
and fibulin (25), have important
roles in the synthetic metabolism and pathogenesis of POP; however,
in the catabolism of the pathogenesis of POP, matrix
metalloproteinase (26–30) and tissue inhibitors of
metalloproteinase serve vital roles (27,30).
In the regulation of anabolic and catabolic processes, the homeobox
gene family is key (31,32). Notably, in the present study, it
was demonstrated that specific key members of the families
mentioned above were detected to be DNA methylated in the molecular
function, biological process and cellular component categories.
KEGG analysis of expression profiling suggested that
the ‘focal adhesion’ and ‘ECM-receptor interaction’ pathways may
potentially serve direct or indirect roles in POP; however, to
date, no studies have linked them to POP. Notably, the two pathways
contained only one hypomethylated gene, ITGA8; however, ITGAV was
hypermethylated in the two pathways. A hypomethylated gene tends to
indicate a high expression level, with hypermethylation meaning
lower expression, in accordance with Kufaishi's studies (33,34)
in vaginal cells derived from premenopausal women with and without
severe POP. Li et al (35)
demonstrated that mechanical strain could activate the
phosphoionositide 3 kinase/protein kinase B signaling pathway in
POP. Chen (36) reported that
smooth muscle cells could activate the TGFER2/ALK5/mothers against
decapentaplegic homolog 2 (Smad)2 and Smad3 signaling pathways,
which may indicate a potential approach for the management of POP.
In these two pathways certain genes, which were differentially
methylated in POP were demonstrated.
Integrins are transmembrane receptors facilitating
cell-ECM adhesion and activating signal transduction pathways. Not
only do they bind cells to the ECM, they also regulate the cell
cycle, organize the intracellular cytoskeleton and translocate
novel receptors to the cell membrane (37). Several types of integrins exist and
one cell may have multiple different types on its surface.
Integrins interact with the ECM to generate large molecular
complexes by focal adhesions. The study of Rhee et al
(38) revealed that POP is
associated with alterations in the
sphingosine-1-phosphate/Rho-kinase signaling pathway, which could
be reduced by a Rho-kinase inhibitor. Apart from ITGAV, there were
two other genes, which were hypermethylated, ITGA9 and ITGB5, which
require further investigation. In the network, ITGAV (degree=32),
ITGA9 (degree=27) and ITGA8 (degree=25) were highly connected. This
suggested that the integrin family may include hub genes in POP.
Fibronectin is the main part of the ECM and while rarely mentioned
with regard to POP, may be another important hub gene.
In the present study, there were certain
limitations. All the ULs were from menopausal women and
premenopausal women were not included. The number in each group was
relatively small. Since microarray experiments use large numbers of
variables (genes) in a small number of samples, standard power
analysis determination according to standard hypothesis testing
cannot be applied (39). Pavlidis
et al reported (39) that
8–15 subjects in each group is sufficient to obtain near-maximal
levels of power in array studies. Gene expression and functional
tests were not performed. Although this issue was not the subject
of this study, the products of these genes and their functions
should be further investigated in future studies.
In conclusion, the present study demonstrated and
elaborated upon the differences in genome-wide DNA methylation
between POP ULs and controls. Epigenetic mechanisms may partly
contribute to the pathogenesis of POP.
Acknowledgements
Not applicable.
Funding
The present study was supported by Beijing
Obstetrics & Gynecology Hospital affiliated Capital Medical
University Funds (Project no. fcyy201407).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
LZ analyzed and interpreted the data, wrote the
paper. DL helped editing the paper. LZ, PZ, AD, YH, CL and DL
collected the tissues.
Ethics approval and consent to
participate
The institutional review board was obtained from the
Beijing Obstetrics and Gynecology Hospital Ethics Committee. The
study was approved in written by Ethics Committee. Informed consent
was obtained from all individual participants included in the
study.
Patient consent for publication
Informed consent was obtained from all individual
participants included in the study.
Competing interests
The authors declare they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
POP
|
Pelvic organ prolapse
|
|
UL
|
uterosacral ligaments
|
|
ECM
|
extracellular matrix
|
|
LOX
|
lysyl oxidase gene
|
References
|
1
|
DeLancey JO: The hidden epidemic of pelvic
floor dysfunction: Achievable goals for improved prevention and
treatment. Am J Obstet Gynecol. 192:1488–1495. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nygaard I, Barber MD, Burgio KL, Kenton K,
Meikle S, Schaffer J, Spino C, Whitehead WE, Wu J and Brody DJ:
Pelvic Floor Disorders Network: Prevalence of symptomatic pelvic
floor disorders in US women. JAMA. 300:1311–1316. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Whitcomb EL, Rortveit G, Brown JS,
Creasman JM, Thom DH, Van Den Eeden SK and Subak LL: Racial
differences in pelvic organ prolapse. Obstet Gynecol.
114:1271–1277. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rortveit G, Brown JS, Thom DH, Van Den
Eeden SK, Creasman JM and Subak LL: Symptomatic pelvic organ
prolapse: Prevalence and risk factors in a population-based,
racially diverse cohort. Obstet Gynecol. 109:1396–1403. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hendrix SL, Clark A, Nygaard I, Aragaki A,
Barnabei V and McTiernan A: Pelvic organ prolapse in the Women's
Health Initiative: Gravity and gravidity. Am J Obstet Gynecol.
186:1160–1166. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Glazener C, Elders A, Macarthur C,
Lancashire RJ, Herbison P, Hagen S, Dean N, Bain C, Toozs-Hobson P,
Richardson K, et al: Childbirth and prolapse: Long-term
associations with the symptoms and objective measurement of pelvic
organ prolapse. BJOG. 120:161–168. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bradley CS, Zimmerman MB, Qi Y and Nygaard
IE: Natural history of pelvic organ prolapse in postmenopausal
women. Obstet Gynecol. 109:848–854. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kudish BI, Iglesia CB, Sokol RJ, Cochrane
B, Richter HE, Larson J, Hendrix SL and Howard BV: Effect of weight
change on natural history of pelvic organ prolapse. Obstet Gynecol.
113:81–88. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xie R, Xu Y, Fan S and Song Y:
Identification of differentially expressed genes in pelvic organ
prolapse by RNA-Seq. Med Sci Monit. 22:4218–4225. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ak H, Zeybek B, Atay S, Askar N, Akdemir A
and Aydin HH: Microarray gene expression analysis of uterosacral
ligaments in uterine prolapse. Clin Biochem. 49:1238–1242. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Allen-Brady K, Cannon-Albright L, Farnham
JM, Teerlink C, Vierhout ME, van Kempen LC, Kluivers KB and Norton
PA: Identification of six loci associated with pelvic organ
prolapse using genome-wide association analysis. Obstet Gynecol.
118:1345–1353. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Allen-Brady K, Cannon-Albright LA, Farnham
JM and Norton PA: Evidence for pelvic organ prolapse predisposition
genes on chromosomes 10 and 17. Am J Obstet Gynecol. 212:771.e1–e7.
2015. View Article : Google Scholar
|
|
13
|
Rao S, Lang J, Zhu L and Chen J: Exome
sequencing identifies a novel gene, WNK1, for susceptibility to
pelvic organ prolapse (POP). PLoS One. 10:e01194822015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sui X, Zhu J, Zhou J, Wang X, Li D, Han W,
Fang Y and Pan H: Epigenetic modifications as regulatory elements
of autophagy in cancer. Cancer Lett. 360:106–113. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Klutke J, Stanczyk FZ, Ji Q, Campeau JD
and Klutke CG: Suppression of lysyl oxidase gene expression by
methylation in pelvic organ prolapse. Int Urogynecol J. 21:869–872.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Aryee MJ, Jaffe AE, Corrada-Bravo H,
Ladd-Acosta C, Feinberg AP, Hansen KD and Irizarry RA: Minfi: A
flexible and comprehensive Bioconductor package for the analysis of
Infinium DNA methylation microarrays. Bioinformatics. 30:1363–1369.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bird AP and Wolffe AP: Methylation-induced
repression-belts, braces, and chromatin. Cell. 99:451–454. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Christensen BC, Houseman EA, Marsit CJ,
Zheng S, Wrensch MR, Wiemels JL, Nelson HH, Karagas MR, Padbury JF,
Bueno R, et al: Aging and environmental exposures alter
tissue-specific DNA methylation dependent upon CpG island context.
PLoS Genet. 5:e10006022009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rang FJ and Boonstra J: Causes and
consequences of age-related changes in DNA methylation: A Role for
ROS? Biology (Basel). 3:403–425. 2014.PubMed/NCBI
|
|
20
|
Borazjani A, Kow N, Harris S, Ridgeway B
and Damaser MS: Transcriptional regulation of connective tissue
metabolism genes in women with pelvic organ prolapse. Female Pelvic
Med Reconstr Surg. 23:44–52. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sun MJ, Cheng YS, Sun R, Cheng WL and Liu
CS: Changes in mitochondrial DNA copy number and extracellular
matrix (ECM) proteins in the uterosacral ligaments of premenopausal
women with pelvic organ prolapse. Taiwan J Obstet Gynecol. 55:9–15.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Connell KA, Guess MK, Chen H, Andikyan V,
Bercik R and Taylor HS: HOXA11 is critical for development and
maintenance of uterosacral ligaments and deficient in pelvic
prolapse. J Clin Invest. 118:1050–1055. 2008.PubMed/NCBI
|
|
23
|
Alarab M, Bortolini MA, Drutz H, Lye S and
Shynlova O: LOX family enzymes expression in vaginal tissue of
premenopausal women with severe pelvic organ prolapse. Int
Urogynecol J. 21:1397–1404. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kobak W, Lu J, Hardart A, Zhang C,
Stanczyk FZ and Felix JC: Expression of lysyl oxidase and
transforming growth factor beta2 in women with severe pelvic organ
prolapse. J Reprod Med. 50:827–831. 2005.PubMed/NCBI
|
|
25
|
Klutke J, Ji Q, Campeau J, Starcher B,
Felix JC, Stanczyk FZ and Klutke C: Decreased endopelvic fascia
elastin content in uterine prolapse. Acta Obstet Gynecol Scand.
87:111–115. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang X, Li Y, Chen J, Guo X, Guan H and Li
C: Differential expression profiling of matrix metalloproteinases
and tissue inhibitors of metalloproteinases in females with or
without pelvic organ prolapse. Mol Med Rep. 10:2004–2008. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Alarab M, Kufaishi H, Lye S, Drutz H and
Shynlova O: Expression of extracellular matrix-remodeling proteins
is altered in vaginal tissue of premenopausal women with severe
pelvic organ prolapse. Reprod Sci. 21:704–715. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Usta A, Guzin K, Kanter M, Ozgul M and
Usta CS: Expression of matrix metalloproteinase-1 in round ligament
and uterosacral ligament tissue from women with pelvic organ
prolapse. J Mol Histol. 45:275–281. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yilmaz N, Ozaksit G, Terzi YK, Yilmaz S,
Budak B, Aksakal O and Sahin FI: HOXA11 and MMP2 gene expression in
uterosacral ligaments of women with pelvic organ prolapse. J Turk
Ger Gynecol Assoc. 15:104–108. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liang CC, Huang HY and Chang SD: Gene
expression and immunoreactivity of elastolytic enzymes in the
uterosacral ligaments from women with uterine prolapse. Reprod Sci.
19:354–359. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ma Y, Guess M, Datar A, Hennessey A,
Cardenas I, Johnson J and Connell KA: Knockdown of Hoxa11 in vivo
in the uterosacral ligament and uterus of mice results in altered
collagen and matrix metalloproteinase activity. Biol Reprod.
86:1002012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Connell KA, Guess MK, Chen HW, Lynch T,
Bercik R and Taylor HS: HOXA11 promotes fibroblast proliferation
and regulates p53 in uterosacral ligaments. Reprod Sci. 16:694–700.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kufaishi H, Alarab M, Drutz H, Lye S and
Shynlova O: Comparative characterization of vaginal cells derived
from premenopausal women with and without severe pelvic organ
prolapse. Reprod Sci. 23:931–943. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kufaishi H, Alarab M, Drutz H, Lye S and
Shynlova O: Static mechanical loading influences the expression of
extracellular matrix and cell adhesion proteins in vaginal cells
derived from premenopausal women with severe pelvic organ prolapse.
Reprod Sci. 23:978–992. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li BS, Guo WJ, Hong L, Liu YD, Liu C, Hong
SS, Wu DB and Min J: Role of mechanical strain-activated PI3K/Akt
signaling pathway in pelvic organ prolapse. Mol Med Rep.
14:243–253. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen X, Kong X, Liu D, Gao P, Zhang Y, Li
P and Liu M: In vitro differentiation of endometrial regenerative
cells into smooth muscle cells: Alpha potential approach for the
management of pelvic organ prolapse. Int J Mol Med. 38:95–104.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Giancotti FG and Ruoslahti E: Integrin
signaling. Science. 285:1028–1032. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rhee SH, Zhang P, Hunter K, Mama ST,
Caraballo R, Holzberg AS, Seftel RH, Seftel AD, Echols KT and
DiSanto ME: Pelvic organ prolapse is associated with alteration of
sphingosine-1-phosphate/Rho-kinase signalling pathway in human
vaginal wall. J Obstet Gynaecol. 35:726–732. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pavlidis P, Li Q and Noble WS: The effect
of replication on gene expression microarray experiments.
Bioinformatics. 19:1620–1627. 2003. View Article : Google Scholar : PubMed/NCBI
|