Introduction
Cervical cancer, the second most predominant cancer
in females, is the fourth most fatal cancer in women worldwide
(1). Human papillomavirus (HPV)
has been demonstrated to cause the initiation and progression of
~99% of cervical tumors (2).
Conventional treatment of cervical cancer, including surgery,
chemotherapy and radiotherapy, may significantly increase chances
of survival (3,4). In total, ~90% of novel cases
occurring in developing countries may be due to the lack of
nationwide screening programs and HPV vaccines (3,4).
However, the pathogenesis and exact molecular mechanisms underlying
the progression of cervical cancer require further investigation.
Therefore, the identification of alterations to crucial genes
associated with cervical cancer is urgently required.
Long non-coding RNAs (lncRNAs), a class of
endogenous non-coding RNAs, are >200 nucleotides in length and
lack protein-coding capacity (5,6).
Accumulating evidence has suggested that lncRNAs serve a vital
regulatory role in numerous physiological and pathological
processes, including cell proliferation, apoptosis, migration,
invasion, differentiation and angiogenesis (5–8).
Aberrant expression of lncRNAs is involved in the progression of a
variety of diseases, including cervical cancer (5,9,10).
For example, downregulated XLOC_010588 expression is an independent
poor prognostic factor and may serve as a predictor of poor
prognosis for patients with cervical cancer (11). Knockdown of XLOC_010588 inhibits
cervical cancer cell growth by decreasing the expression of
proto-oncogene c-Myc (11).
Overexpression of the lncRNA HOTAIR promoted cervical cancer cell
proliferation, migration and invasion and was associated with lymph
node metastasis and shorter overall survival (12). lncRNA MEG3 expression level is
significantly lower in cervical cancer tissues, and downregulation
of MEG3 expression is correlated with the Fédération Internationale
de Gynécologie et d'Obstétrique (FIGO) stage and lymph node
metastasis in cervical cancer patients (10). lncRNA DLX6-AS1 expression is
significantly higher in lung adenocarcinoma tissues and high
DLX6-AS1 expression is correlated with higher histological
differentiation and advanced Tumor, Node and Metastasis (TNM)
stages (13). Furthermore,
DLX6-AS1 modulates cell growth and tumorigenesis via regulation of
the micro RNA (miRNA/miR)-26a/PTEN axis in renal cell carcinoma
(14). In addition, DLX6-AS1
silencing inhibits the proliferation, migration and invasion of
hepatocellular carcinoma cells via the miR-203a/matrix
metalloprotease (MMP)-2 pathway (15). However, the expression and
biological functions of DLX6-AS1 in cervical cancer remain largely
unknown.
In the present study, it was observed that DLX6-AS1
expression was significantly increased in cervical cancer tissues
and cell lines. Furthermore, DLX6-AS1 knockdown impaired cell
proliferation and induced cell apoptosis in vitro. The
interaction between DLX6-AS1 and miR-199a was additionally examined
to reveal the underlying mechanism of DLX6-AS1 in cervical
cancer.
Materials and methods
Tissue samples
A total of 78 paired adjacent noncancerous specimens
and cervical cancer samples were collected from patients who had
undergone surgical resection at the Department of Gynecology of the
Women's Hospital of Zhejiang University (Hangzhou, China) between
May 2010 and December 2013. Patients (age range, 37–62 years, and
mean 52.4 years) were diagnosed with cervical cancer by
histological pathology. Tumor grades and stages were adjusted to
comply with the new 2009 classification (16). None of the patients had received
preoperative chemotherapy or radiotherapy. All patients enrolled in
the present study were pathologically confirmed as having cervical
cancer, and written informed consent was obtained from each
patient. The present study was approved by the Ethics Committee of
Zhejiang University (license no. 2016-01AH). All samples were
frozen at the time of surgery in liquid nitrogen and stored at
−80°C until use.
Cell lines
Human immortalized cervical epithelial cell lines
(NC104) and human cervical cancer cell lines (CaSki, ME-180, C-33A,
SiHa and HeLa) were purchased from the Cell Bank of the Shanghai
Institutes for Biological Sciences, Chinese Academy of Sciences
(Shanghai, China). All cell lines were cultured in Dulbecco's
modified Eagle's medium (DMEM; Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (FBS; Invitrogen; Thermo Fisher Scientific, Inc.),
penicillin (100 U/ml), and streptomycin (100 mg/ml) in a humidified
incubator at 37°C in 5% CO2.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA from tissues and cells was isolated using
Trizol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. RNA was reverse
transcribed to complementary DNA (cDNA) using PrimeScript™ 1st
Strand cDNA Synthesis kit (Takara Biotechnology Co., Ltd., Dalian,
China) under conditions of 30°C for 10 min, 42°C for 30 min, 95°C
for 5 sec, and 4°C for 10 min. RT-qPCR was performed using Syber
Green PCR mastermix (Applied Biosystems; Thermo Fisher Scientific,
Inc.) on an ABI7500 system (Applied Biosystems; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. The
sequences of the primers were as follows: lncRNA DLX6-AS1,
5′-AGTTTCTCTCTAGATTGCCTT-3′ (forward) and
5′-ATTGACATGTTAGTGCCCTT-3′ (reverse); and GAPDH,
5′-GCACCGTCAAGGCTGAGAAC-3′ (forward) and 5′-TGGTGAAGACGCCAGTGGA-3′
(reverse). PCR was performed under the following parameters:
Pre-denaturation cycle of 2 min at 94°C, 40 cycles of 95°C for 15
sec, 60°C for 30 sec, and 72°C for 2 min, with a final extension at
72°C for 5 min. GAPDH was used as a loading control. The relative
expressions of DLX6-AS1 were normalized to GAPDH and calculated
according to 2−∆∆Cq method (17).
Cell transfection
Specific small interfering (si)RNA oligonucleotides
(Shanghai GenePharma Co., Ltd., Shanghai, China) targeting DLX6-AS1
and negative control siRNAs were obtained from Shanghai GenePharma
Co., Ltd. miR-199a mimics, miR-199a inhibitor and its negative
control were additionally purchased from Shanghai GenePharma Co.,
Ltd. For transfection, a total of 6×105 cells were
seeded into 100 mm culture dishes and transfected with the
respective RNA oligonucleotides at a final concentration of 50 nm,
using Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturers' protocol.
Cell viability and proliferation
assay
Cells transfected with a corresponding
oligonucleotide were seeded into 96-well plates at a density of
3×104 cells/well. A volume of 10 µl Cell Counting Kit-8
(CCK-8) solution (Dojindo Molecular Technologies, Inc., Kumamoto,
Japan) was added to each well at 24, 48, 72 and 96 h of incubation,
followed by an incubation of 2 h. The absorbance at 450 nm was
measured using a SpectraMaxM3 microplate reader (Molecular Devices,
LLC, Sunnyvale, CA, USA).
Colony formation assay
The cells were seeded in each well of a 6-well
culture plate at a density of 600 cells/well and cultured in DMEM
supplemented with 10% FBS for 2 weeks with the growth media
replaced every third day. The colonies (containing ≥50 cells) were
washed with PBS, fixed with methanol for 15 min at room temperature
and stained with 1% crystal violet (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) for 15 min at room temperature. Cell number was
calculated under an inverted light microscope (TS100; Nikon
Corporation, Tokyo, Japan).
Cell apoptosis analyses
Cell apoptosis was assessed with Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) staining
(BD Biosciences, Franklin Lakes, NJ, USA) according to the
manufacturer's protocol. Cells were harvested and washed twice with
PBS, and 5 µl PI and 5 µl FITC were added and incubated at room
temperature in the dark for 15 min. Cell samples were analyzed
using a flow cytometer (FACScan; BD Biosciences) with CellQuest Pro
Software (version 5.1, BD Biosciences).
Reporter vectors construction and
luciferase assays
To identify miRNAs that bind DLX6-AS1, predictions
were made with starBase v2.0 (starbase.sysu.edu.cn/mirLncRNA.php). The sequence
containing the mir-199a binding sites plus 500 base pairs at 5′-
and 3′-flanking regions in DLX6-AS1 was amplified by PCR using
PrimeSTAR® Max DNA Polymerase (Takara Biotechnology Co.,
Ltd.) and cloned into Psi-CHECK-2 vectors (Promega Corporation,
Madison, WI, USA). The sequence of primers as follows: (forward)
5′-GGCCTAGTCAACCAAGGATG-3′ and (reverse)
5′-GGTCCCCTTGCTGAAGATTAC-3′. The corresponding mutants were created
by mutating the mir-199a seed-region binding site using
QuikChange® Multi Site-Directed Mutagenesis kit (Agilent
Technologies, Inc., Santa Clara, CA, USA). A total of 1,000
cervical cancer cells were plated in a 96-well plate (Corning
Incorporated, Corning, NY, USA) and co-transfected with 100 ng
wild-type Psi-CHECK-2-DLX6-AS1 (or DLX6-AS1 mutant) reporter
plasmid and 20 ng miR-199a mimics or mimic controls using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). The luciferase activities were measured using
the Dual-Luciferase Reporter Assay System and normalized to Renilla
luciferase activities (Promega Corporation) after 36 h, according
to the manufacturer's protocol.
Statistical analysis
Data are expressed as the mean ± standard deviation,
and statistical analysis was performed using GraphPad Prism
(version 5.01; GraphPad Software, Inc., La Jolla, CA, USA). These
experiments were performed in triplicate. Differences between
groups were tested using paired and unpaired Student's t-test, and
one-way analysis of variance followed by Tukey's post hoc test were
performed to assess the difference between more than two groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
lncRNA DLX6-AS1 expression is
upregulated in cervical cancer tissues and cells
Altered DLX6-AS1 expression has been identified in a
number of types of tumors; however, whether aberrant DLX6-AS1
expression is involved in cervical cancer development remains
unknown. Expression levels of DLX6-AS1 were investigated in the 78
cervical cancer tissue samples and matched non-tumor adjacent
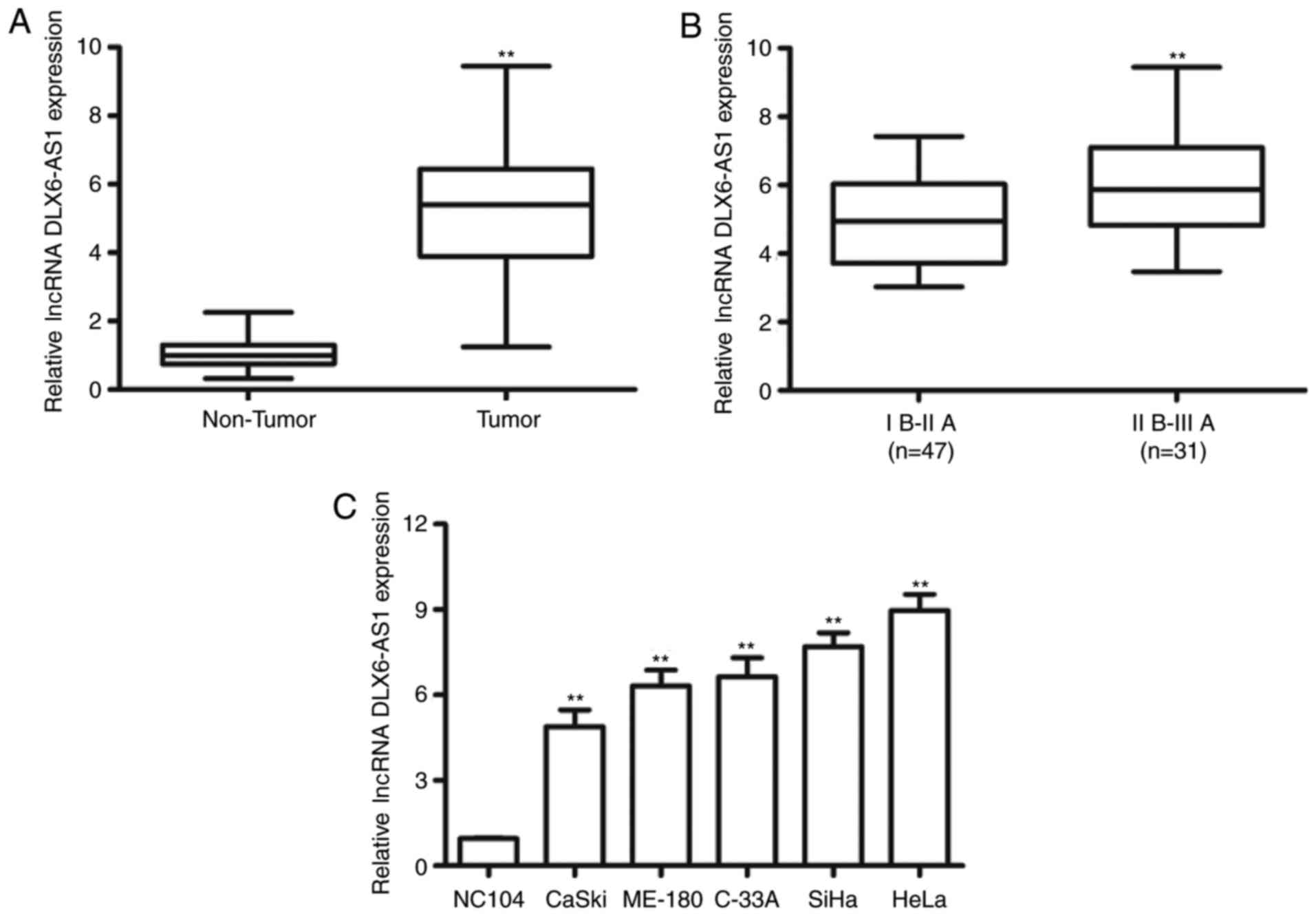
tissue samples using RT-qPCR. As demonstrated in Fig. 1A, DLX6-AS1 was significantly
overexpressed in cervical cancer tissue samples compared with the
non-tumor tissue samples from the same patient (P<0.01).
Furthermore, DLX6-AS1 expression was significantly increased in
patients at advanced FIGO stages (II B-III A phase) compared with
those at earlier clinical stages (I B-II A phase; P<0.01;
Fig. 1B). In addition, the
expression of DLX6-AS1 in five human cervical cancer cell lines
(CaSki, ME-180, C-33A, SiHa and HeLa) and immortalized cervical
epithelial cell lines (NC104) was examined. The expression of
DLX6-AS1 was significantly upregulated in cervical cancer cells
compared with the NC104 cells (P<0.01; Fig. 1C). These results suggested that
DLX6-AS1 may serve as an oncogene involved in the tumorigenesis and
progression of cervical cancer.
lncRNA DLX6-AS1 affects cervical
cancer cell proliferation
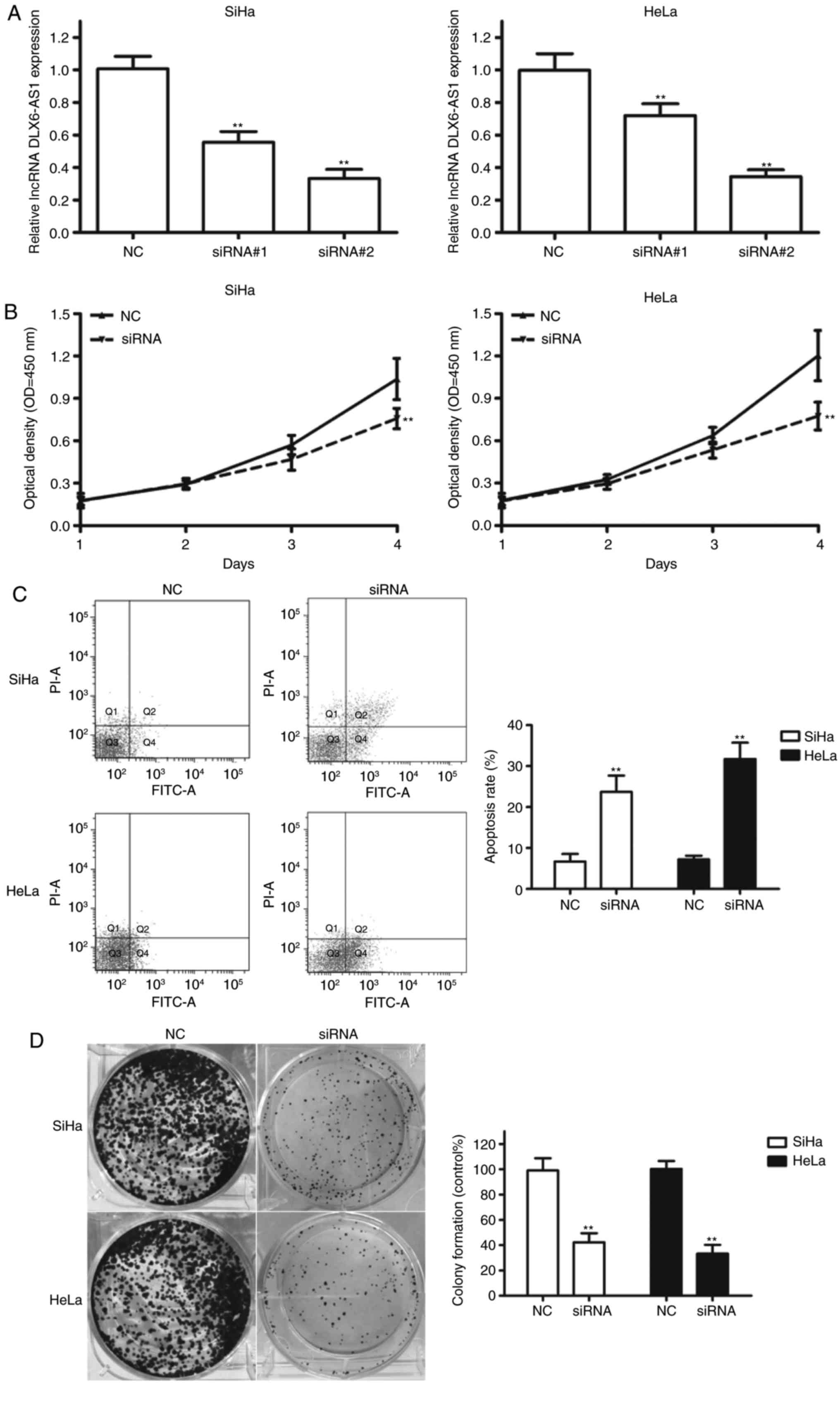
To investigate the biological role of DLX6-AS1 in
the progression of cervical cancer, DLX6-AS1 was knocked down in
SiHa and HeLa cells with high endogenous DLX6-AS1 expression by
transfecting with specific DLX6-AS1 siRNAs. As demonstrated in
Fig. 2A, transfection with siRNA#1
and siRNA#2 significantly decreased the DLX6-AS1 expression levels
in SiHa and HeLa cells compared with the negative control (NC;
P<0.01). siRNA#2 was more effective at inhibiting DLX6-AS1
expression; therefore, this construct was termed ‘siRNA’ and used
for all subsequent experiments. A CCK-8 assay demonstrated that
DLX6-AS1 silencing in SiHa and HeLa cell lines resulted in a
decrease in proliferative capability compared with the NC (Fig. 2B). Subsequently, apoptosis was
assessed using flow cytometry, and the results demonstrated that
silencing DLX6-AS1 led to a significant increase in the apoptotic
rate of the two cell lines compared with the NC (Fig. 2C; P<0.01). In addition,
colony-formation assays revealed a significant decrease in colony
formation following DLX6-AS1 knockdown in the cervical cancer cell
lines (Fig. 2D; P<0.01).
Therefore, it may be concluded that DLX6-AS1 mediates cervical
cancer cell proliferation by regulating cell apoptosis.
Reciprocal repression exists between
lncRNA DLX6-AS1 and miR-199a in cervical cancer cells
Increasing evidence has demonstrated that lncRNAs
may serve as competitive endogenous RNAs (ceRNAs) to regulate the
target mRNAs of miRNAs (6). A
number of miRNAs were predicted to target DLX6-AS1 (data not
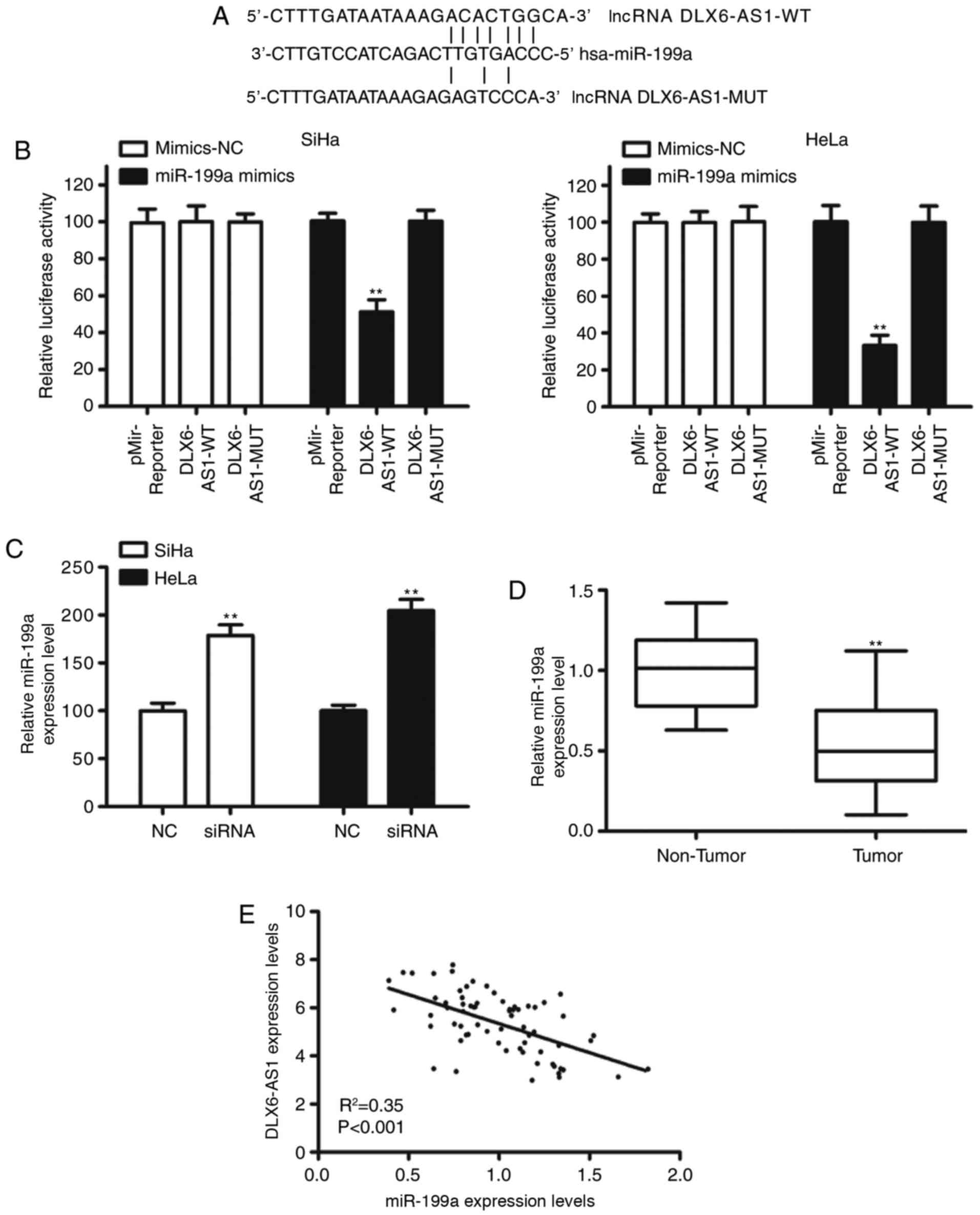
shown); however, miR-199a was selected as miR-199a is reported to
serve as a tumor suppressor (16–18).
The binding sites of DLX6-AS1 and miR-199a are presented in
Fig. 3A. Dual reporter luciferase
assays revealed that co-transfection of miR-199a mimics and
DLX6-AS1-wild-type (WT) luciferase reporter constructs
significantly reduced the luciferase activity of DLX6-AS1-WT in
SiHa and HeLa cell lines (P<0.01; Fig. 3B). However, this inhibition was
eliminated by the introduction of nucleotide alterations to the
predicted seed-binding sequences of miR-199a (Fig. 3B). Furthermore, knockdown
DLX6-AS1increased miR-199a expression in cervical cancer cells
(Fig. 3C). The expression of
miR-199a in clinical samples was assessed and it was observed that
the expression of miR-199a in cervical cancer tissues was
significantly decreased compared with adjacent normal tissue
(P<0.01; Fig. 3D). Spearman
correlation analysis demonstrated a significant inverse correlation
between miR-199a expression and DLX6-AS1 expression (Fig. 3E). These results suggested that
DLX6-AS1 may serve as an oncogene by medicating miR-199a expression
in cervical cancer.
DLX6-AS1 enhances cervical cancer cell
proliferation and migration by inhibiting miR-199a expression
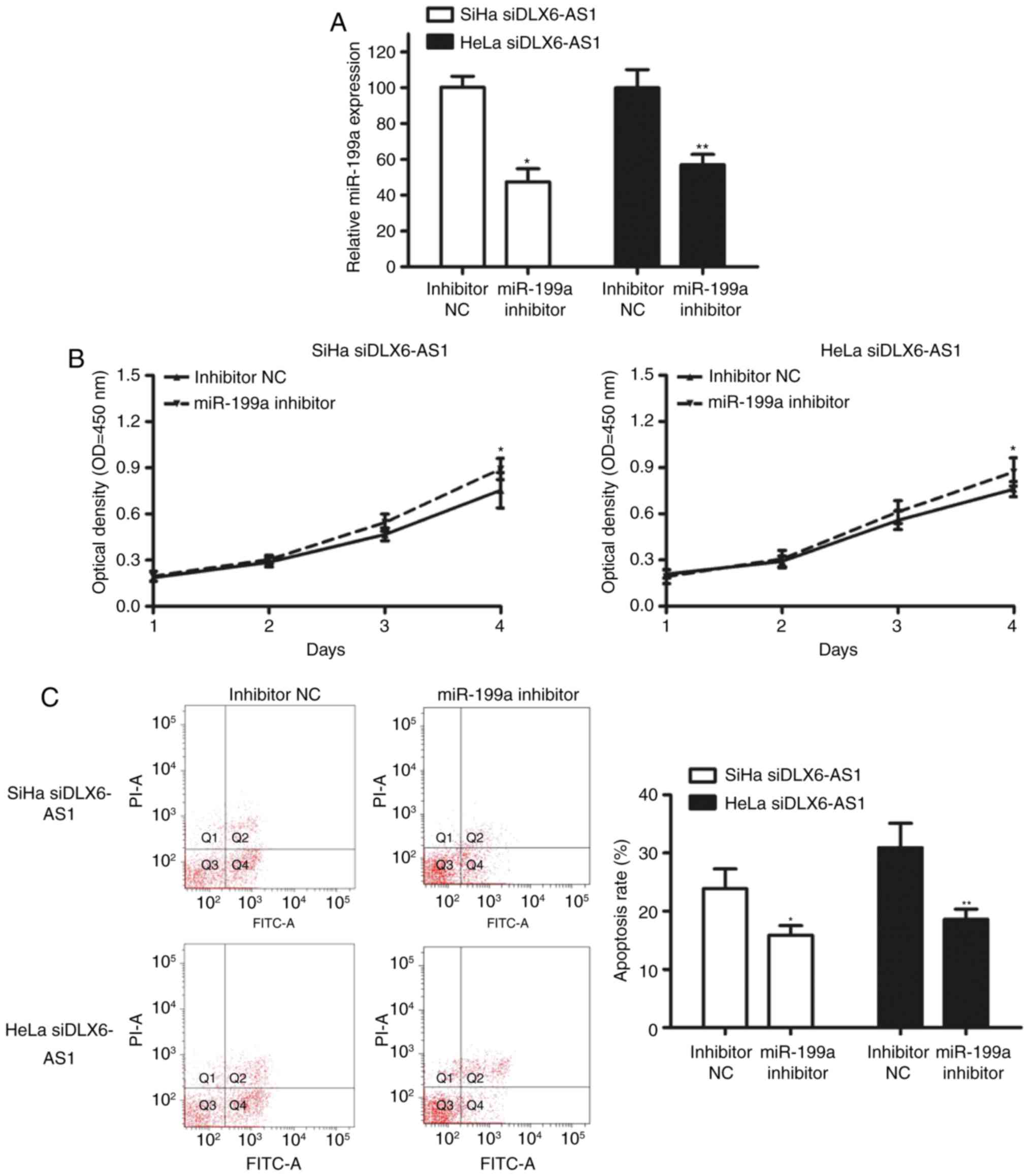
To determine whether DLX6-AS1 mediates decreased
miR-199a-induced inhibition of proliferation and cell apoptosis
in vitro, miR-199a was inhibited in DLX6-AS1 knockdown cells
by transfection with miR-199a inhibitors, which resulted in a
significant decrease of miR-199a in SiHa and HeLa cells (Fig. 4A). As hypothesized, miR-199a
silencing led to a marked higher cell proliferation rate (Fig. 4B) and a significantly lower
apoptosis rate (Fig. 4C) compared
with the control cells, which indicated that DLX6-AS1 partly
functions by downregulating miR-199a.
Discussion
An increased understanding of the important role of
non-coding RNA in the initiation and progression of multiple cancer
types has revealed an exciting avenue for the development of novel
cancer therapies (5,9,19).
Until now, the molecular mechanism and effect of lncRNAs were
largely unknown in cervical cancer tumorigenesis and progression.
In the present study, the expression and molecular mechanisms of
DLX6-AS1 in cervical cancer were investigated. High expression of
DLX6-AS1 was observed in cervical cancer tissues and cells, and
expression increased with the FIGO stage. Furthermore, knockdown of
DLX6-AS1 using siRNA inhibited cell proliferation and induced
apoptosis in vitro. In addition, the present study provided
evidence that DLX6-AS1 exerted oncogene functions by downregulating
miR-199a, a well-known tumor suppressor.
A previous study has suggested that DLX6-AS1 acts as
an oncogene in lung adenocarcinoma and that high DLX6-AS1
expression levels are significantly associated with histological
differentiation and TNM stage (13). It has been recently demonstrated
that DLX6-AS1 is upregulated in renal cell carcinoma and that
DLX6-AS1 promotes renal cell carcinoma cell growth and
tumorigenesis by functioning as a ceRNA to sponge miR-26a (14). Furthermore, DLX6-AS1 promotes cell
proliferation, migration and invasion by regulating the
miR-203a/MMP-2 pathway (15).
Consistent with these results, DLX6-AS1 was additionally identified
in cervical cancer tissues and cell lines, and the expression of
DLX6-AS1 was positively associated with FIGO stage. DLX6-AS1
knockdown decreased the proliferation of cervical cancer cells by
inducing cell apoptosis.
A previous study suggested that lncRNAs serve as
ceRNAs to sponge miRNAs and regulate the biological function of
cancer cells (19). To elucidate
the mechanism of DLX6-AS1 in cervical cancer, bioinformatics
analysis was performed to predict miRNAs that bind DLX6-AS1.
Notably, it was confirmed that expression levels of miR-199a were
downregulated in cervical cancer tissues. Furthermore, it was
observed that the expression level of DLX6-AS1 was negatively
correlated with the expression level of miR-199a in cervical cancer
samples. Knockdown of miR-199a abrogated the inhibition of cell
growth and apoptosis caused by DLX6-AS1, which was consistent with
previous studies of cancer cells (14,15,20,21),
suggesting that DLX6-AS1 may promote cell proliferation by serving
as a ceRNA to sponge miR-199a in cervical cancer.
In conclusion, the present results demonstrated that
DLX6-AS1 expression was higher in cervical cancer, and high
DLX6-AS1 expression was associated with FIGO stage in patients with
cervical cancer. Furthermore, knockdown of DLX6-AS1 significantly
induced cell apoptosis and inhibited cell growth and as an inverse
correlation was identified between miR-199a and DLX6-AS1
expression, so knockdown of DLX6-AS1 results in an increased
miR-199a expression. These results indicate that DLX6-AS1 is
important for cervical cancer progression and suggest that it may
be used as a potential therapeutic target for cervical cancer
treatment.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Zhejiang
Natural Science Youth Fund (grant no. LQ13H040001).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
XW and YL contributed to design, wrote, and revised
the manuscript. JL collected and classified the human BC tissue
samples. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Zhejiang University (license no. 2016-01AH) and
written informed consent was obtained from each patient.
Patient consent for publication
Written informed consent was obtained from all human
subjects.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Muñoz N, Bosch FX, de Sanjosé S, Herrero
R, Castellsagué X, Shah KV, Snijders PJ and Meijer CJ:
International Agency for Research on Cancer Multicenter Cervical
Cancer Study Group: Epidemiologic classification of human
papillomavirus types associated with cervical cancer. N Engl J Med.
348:518–527. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Crosbie EJ, Einstein MH, Franceschi S and
Kitchener HC: Human papillomavirus and cervical cancer. Lancet.
382:889–899. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gravitt PE and Rositch AF: HPV
self-testing and cervical cancer screening coverage. Lancet Oncol.
15:128–129. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huarte M: The emerging role of lncRNAs in
cancer. Nat Med. 21:1253–1261. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lu X, Zhou C, Li R, Deng Y, Zhao L and
Zhai W: Long noncoding RNA AFAP1-AS1 promoted tumor growth and
invasion in cholangiocarcinoma. Cell Physiol Biochem. 42:222–230.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fatica A and Bozzoni I: Long non-coding
RNAs: New players in cell differentiation and development. Nat Rev
Genet. 15:7–21. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hosseini ES, Meryet-Figuiere M,
Sabzalipoor H, Kashani HH, Nikzad H and Asemi Z: Dysregulated
expression of long noncoding RNAs in gynecologic cancers. Mol
Cancer. 16:1072017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang J, Lin Z, Gao Y and Yao T:
Downregulation of long noncoding RNA MEG3 is associated with poor
prognosis and promoter hypermethylation in cervical cancer. J Exp
Clin Cancer Res. 36:52017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liao LM, Sun XY, Liu AW, Wu JB, Cheng XL,
Lin JX, Zheng M and Huang L: Low expression of long noncoding
XLOC_010588 indicates a poor prognosis and promotes proliferation
through upregulation of c-Myc in cervical cancer. Gynecol Oncol.
133:616–623. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sun J, Chu H, Ji J, Huo G, Song Q and
Zhang X: Long non-coding RNA HOTAIR modulates HLA-G expression by
absorbing miR-148a in human cervical cancer. Int J Oncol.
49:943–952. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li J, Li P, Zhao W, Yang R, Chen S, Bai Y,
Dun S, Chen X, Du Y, Wang Y, et al: Expression of long non-coding
RNA DLX6-AS1 in lung adenocarcinoma. Cancer Cell Int. 15:482015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zeng X, Hu Z, Ke X, Tang H, Wu B, Wei X
and Liu Z: Long noncoding RNA DLX6-AS1 promotes renal cell
carcinoma progression via miR-26a/PTEN axis. Cell Cycle.
16:2212–2219. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang L, He X, Jin T, Gang L and Jin Z:
Long non-coding RNA DLX6-AS1 aggravates hepatocellular carcinoma
carcinogenesis by modulating miR-203a/MMP-2 pathway. Biomed
Pharmacother. 96:884–891. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
FIGO Committee on Gynecologic Oncology.
Current FIGO staging for cancer of the vagina, fallopian tube,
ovary, and gestational trophoblastic neoplasia. Int J Gynaecol
Obstet. 105:3–4. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hou J, Lin L, Zhou W, Wang Z, Ding G, Dong
Q, Qin L, Wu X, Zheng Y, Yang Y, et al: Identification of miRNomes
in human liver and hepatocellular carcinoma reveals miR-199a/b-3p
as therapeutic target for hepatocellular carcinoma. Cancer Cell.
19:232–243. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ghosh A, Dasgupta D, Ghosh A, Roychoudhury
S, Kumar D, Gorain M, Butti R, Datta S, Agarwal S, Gupta S, et al:
MiRNA199a-3p suppresses tumor growth, migration, invasion and
angiogenesis in hepatocellular carcinoma by targeting VEGFA VEGFR1,
VEGFR2, HGF and MMP2. Cell Death Dis. 8:e27062017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Celià-Terrassa T, Liu DD, Choudhury A,
Hang X, Wei Y, Zamalloa J, Alfaro-Aco R, Chakrabarti R, Jiang YZ,
Koh BI, et al: Normal and cancerous mammary stem cells evade
interferon-induced constraint through the miR-199a-LCOR axis. Nat
Cell Biol. 19:711–723. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The Rosetta Stone of a hidden RNA
language? Cell. 146:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|