Introduction
Preeclampsia (PE) is characterized by hypertension
and proteinuria. The condition increases the risk for maternal and
perinatal morbidity and mortality, with worldwide incidence of
approximately 4–6% (1). Despite
extensive research, the etiology and pathogenesis of PE are not
completely understood. Previous research has shown that that
placental tissue but not the fetus is necessary for the development
of PE. The condition is cured upon delivery of the placenta, which
underscores the indispensable role of this tissue in the
pathophysiology of PE (2).
The consensus reached at a recent workshop sponsored
by the National Institute of Child Health and Human
Development/National Institutes of Health is that preeclampsia is a
multifactorial disease whose pathogenesis is not solely vascular,
genetic, immunologic, or environmental but a complex combination of
factors (3). In PE, excessive
activation of peripheral blood leukocytes is associated with
exaggerated innate and adaptive immune responses, which may
interfere with normal progression of a pregnancy (4).
Regulatory T cells (Tregs) are a specialized
subpopulation of T cells that act as potent suppressors of
inflammation to prevent autoimmunity and graft rejection (5). Tregs prevent activation of maternal T
cells against fetal cells; this protection of the fetus from the
maternal immune system has been widely reported in both mice and
humans (6–8). A reduction in maternal Treg
populations could prevent immunological tolerance of the fetus and
has been associated with obstetrical complications, such as
miscarriage, PE and preterm labor (9–14).
Tregs comprise Th1 and Th2 cells. Tregs expressing
forkhead box p3 (Foxp3) help to maintain immune tolerance and
homeostasis. At the steady-state level, transforming growth
factor-β 1 (TGF-β1) produced in the immune system induces
Foxp3+ regulatory T cells and maintains self-tolerance
(15). However, in the
inflammatory insult, by the activated innate immune system IL-6
will suppress the generation of TGF-β1-induced Treg cells and
induce a pro-inflammatory Th17 cell response (15). CD4+CD25+
Tregs play a critical role in maternal tolerance in mice and in
humans (6,9,16).
CD4+CD25+ Tregs play a critical role in PE.
This immunological disturbance seems to result from overactivation
of the maternal inflammatory response, which involves
overproduction of pro-inflammatory cytokines such as interferon-γ
(IFN-γ), interleukin (IL)-1β, IL-6 and tumor necrosis factor-α
(TNF-α) (14–16) and lower concentrations of
regulatory cytokine IL-10 (17,18).
Th17 cells are a lineage of CD4+ T cells that
differentiate in the presence of IL-6 and TGF-β. These cells
express transcription factor retinoic acid-related orphan receptor
C (RORc) and produce cytokine IL-17 (19).
Previous researchers found a decrease in Tregs and
an increase in Th17-profile cells in the peripheral blood of women
with PE compared to normal pregnant women. This imbalance may
underlie activation of the inflammatory response in women with PE
(20,21).
Tregs are also considered to play a crucial role in
the implantation of embryos and the maintenance of maternal immune
tolerance to the fetus (22).
Zhang et al demonstrated that the percentage of
CD4+CD25+CD127− cells among the
CD4+ T cells was significantly lower in patients with
severe PE compared to healthy pregnant women (23). Numerous researchers have sought to
establish a protocol for identifying Tregs. Intracellular Foxp3
staining cannot be used to isolate the subpopulation of Tregs for
functional tests, as it requires that cells be fixed and
permeabilized. Published reports have shown that simultaneous high
expression of CD25 and low expression of CD127 correspond with
increased expression of intracellular factor Foxp3. This profile
allows researchers to identify T cells with suppressor activity
(24,25).
A correlation between
CD4+CD25+CD127− cells and
CD4+CD25+Foxp3+ cells has been
confirmed in healthy individuals (26). No significant difference was found
between the number of CD25+CD127− cells among
the CD4+ T cells with and without permeabilization in
the same NHDs (27).
MicroRNAs (miRNAs) are a major class of
tissue-specific regulators of gene expression (28). Numerous studies have shown that the
miRNAs expressed primarily during pregnancy are clustered in
chromosomal regions, suggesting control by the same promoters, and
overlap in target genes (29).
Many miRNAs have been shown to tightly control placental
development, which is a complex physiological process that involves
numerous cell types (30). miRNAs
are exported from human placental syncytiotrophoblasts into the
maternal circulation via exosomes, where they regulate trophoblasT
cell invasion, placental immune activation and platelet aggregation
(31,32).
Alterations in placental miRNA expression have been
associated with in utero exposure and adverse pregnancy outcomes
(33–36). Zhao et al (37) reported significantly higher levels
of miR-210 in CD4+ T cells of patients with psoriasis,
compared with controls. The same authors confirmed Foxp3 as a
target gene regulated by miR-210. After overexpression of
miRNA-210, CD4+ T cells from healthy controls showed
decreased expression of Foxp3 and impaired Treg immune suppressor
function.
The present study aimed to explore the profile of Th
cytokines and Treg/Th17 cells in healthy non-pregnant, normal
pregnant and preeclamptic women. The aim of the present study was
to characterize the evolution of Treg subsets (defined as
CD4+CD25+,
CD4+CD25+CD127− T cells) in
peripheral blood in patients with PE. To investigate the mechanisms
underlying Treg/Th17 activity in patients with PE, we also measured
mRNA expression of Foxp3, retinoic acid-related orphan receptor C
(RORc) and miR-210 in placenta from patients with PE.
Materials and methods
A total of 66 women (age, 24–38 years) were included
in this study: 29 patients with late-onset preeclampsia (≥36 weeks
of gestation), 27 pregnant women with normal uncomplicated
pregnancies (≥36 weeks of gestation) and 10 healthy non-pregnant
women. Serum levels of cytokines (IL-6, IL-10, IL-17 and TGF-β1)
were detected using enzyme-linked immunosorbent assay (ELISA).
Levels of CD4+CD25+CD127low/−
Tregs in peripheral blood were evaluated by flow cytometry (FCM).
The study included 29 patients with PE with placenta as the
experimental group and 27 women with normal pregnancy placenta as a
control group. Reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) was performed to detect mRNA expression of
maternal placenta retinoic acid-related orphan receptor C (RORc),
Foxp3 and miR-210. Expression of inflammation-related protein Foxp3
was evaluated by western blot analysis. Samples from the same
patient were used to quantify the levels of Tregs, FOX-3 and
miR-210.
Study patients
Study participants were enrolled at the Department
of Obstetrics and Gynecology, Shenzhen Longhua District Central
Hospital (Shenzhen, China). Exclusion criteria were multifetal
gestation, hypertension, diabetes, autoimmune or vascular disease,
infection (HIV, syphilis or hepatitis B or C) and renal
disorder.
Pregnant women were excluded if they had signs of
infection. None of the pregnant women were in active labor, and
none had rupture of the amniotic membrane. Serum samples were
collected at the time of delivery via Cesarean section. Placental
tissue was collected during Cesarean section for testing Foxp3,
RORc mRNA and miR-210 expression. We also recruited sequential
healthy non-pregnant women who were attending routine annual
well-woman examinations. None of these women had received hormonal
contraception, which may influence peripheral blood lymphocytes
(38).
The clinical diagnosis of PE was based on the
development of hypertension [blood pressure (BP) ≥140×90 mmHg] and
proteinuria (≥300 mg/24 h) during week 20 of gestation or later
(39). Late onset PE (≥34 weeks)
was defined as those patients who displayed gestational
hypertension and proteinuria without a history of hypertension.
All women included in the study were recruited
during the period from July 2016 to March 2017. The Ethics
Committee of the Shenzhen Longhua District Central Hospital
approved the study protocol. All patients provided written informed
consent prior to the participation in the study.
Measurement of cytokine levels in
serum and placenta
Whole blood was collected in sterile EDTA tubes and
centrifuged at 3,000 × g for 15 min to separate the plasma. Plasma
samples were stored at −80°C for subsequent analysis. Cytokine
concentrations in plasma were determined by enzyme-linked
immunosorbent assay (ELISA). All ELISA kits were purchased from
Mlbio Co., Ltd. (Shanghai, China), including: IL-6 (cat. no.
m19028583), IL-17 (cat. no. m19028599), IL-10 (cat. no. m19027436)
and TGF-β1 (cat. no. m16025653). Assay sensitivity limits for IL-6,
IL-10, TGF-β1 and IL-17 (Th17) were 1.5, 1.25, 7.5 and 10 pg/ml,
respectively. Absorbance was measured at 450 nm.
Flow cytometry
EDTA anticoagulated whole blood samples were
collected and processed within 24 h. Briefly, peripheral blood
mononuclear cells (PBMCs) were obtained with standard
Ficoll-Hypaque technique. PBMCs (5×105) were stained
with the following antibodies: Fluorescein
isothiocyanate-conjugated CD127 (cat. no. 560549; BD Pharmingen; BD
Biosciences, San Jose, CA, USA); phycoerythrin-conjugated CD25
(cat. no. 557138; BD Pharmingen; BD Biosciences) and
allophycocyanin-conjugated CD4 (cat. no. 551980; BD Pharmingen; BD
Biosciences) at 4°C in dark for 30 min. Following washing with FACs
buffer (PBS with 2% fetal bovine serum (FBS, cat. no. 10099141;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) and 1 mM EDTA),
cells were analyzed using a BD FACSAria II cell sorter with BD
FACSDiva software (Version 6.1; BD Biosciences); a subpopulation of
CD4+CD25+CD127− cells was
identified.
mRNA expression of Foxp3 and RORc
Placentas from a total of 56 women were included in
this study. The women consented to the collection of samples at the
time of birth. A full-thickness placental biopsy was obtained after
delivery, avoiding the periphery and areas of obvious infarction.
Samples obtained were flash frozen in liquid nitrogen and stored at
−80°C prior to analysis. RT-qPCR was performed to measure mRNA
expression of ROR, Foxp3 and miR-210 in maternal placenta.
mRNA expression levels of transcription factors
associated with Treg/Th17 in placenta were measured by RT-qPCR.
miRNA was extracted from 30–50 mg fresh tissue with the Qiagen
miRNeasy Mini kit (cat. no. 217004; Qiagen GmbH, Hilden, Germany).
Total RNA was extracted from 30–50 mg fresh tissue using
TRIzol® Reagent (cat. no. 15596026; Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) according to the
manufacturer's instructions. cDNA was synthesized with PrimeScript
RT Master Mix (Takara Bio, Inc., Shiga, Japan), as described in the
manufacturer's instructions. qPCR was performed with PowerUp SYBR
Green Master Mix (cat. no. a25780; Thermo Fisher Scientific, Inc.).
Table II shows all PCR primer
pairs. qPCR was carried out in duplicate wells using a LightCycler
Real-Time PCR System (Roche Diagnostics, Indianapolis, IN, USA).
Quantitation cycle (Cq) values for target genes and the
housekeeping gene GAPDH were calculated. Levels of mRNA and miRNA
expression were normalized to GAPDH and U6 respectively, and
relative expression of indicated genes in PE were compared to that
in normal pregnant women using the 2−∆∆Cq method
(40).
 | Table II.Primer sequences used for
RT-qPCR. |
Table II.
Primer sequences used for
RT-qPCR.
| Gene | qPCR primer
sequence (5′→3′) |
|---|
| Foxp3 | F:
GTGGCCCGGATGTGAGAAG |
|
| R:
GGAGCCCTTGTCGGATGATG |
| RORc | F:
GAAGTGGTGCTGGTTAGGATGTG |
|
| R:
GCCACCGTATTTGCCTTCAA |
| GAPDH | F:
GCACCGTCAA-GGCTGAGAAC |
|
| R:
TGGTGAAGACGCCAGTGGA |
| U6 | F:
TGCGGGTGCTCGCTTCGGCAGC |
|
| R:
CCAGTGCAGGGTCCGAGGTAT |
| miR-210 | F:
CGGCGGTCTGTGCGTGTGACAGC |
|
| R:
CCAGTGCAGGGTCCGAGGTAT |
|
| Gene | RT primer
sequence (5′→3′) |
|
| U6 RT |
GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAAAATATGGAAC |
| miR-210 RT |
GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTCAGCC |
Western blotting of Foxp3
For protein extraction, 30–50 mg placentas were
homogenized, washed with PBS, and lysed for 30 min in ice-cold
radioimmunoprecipitation assay lysis buffer supplemented with 10 mM
NaF, 1 mM Na3VO4, 1 mM phenylmethylsulfonyl
fluoride and protease inhibitor cocktail (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany). Protein concentrations were determined
by Pierce BCA Protein Assay kit, according to the manufacturer's
instructions (cat. no. 23225; Thermo Fisher Scientific, Inc.).
Proteins samples (~30 µg) were separated by 10% SDS-PAGE and
electrotransferred to polyvinylidene fluoride (PVDF) membranes (EMD
Millipore, Billerica, MA, USA). PVDF membranes were blocked using
5% non-fat dried milk dissolved in TBS (20 mM Tris-HCl, 150 mM
NaCl, pH 7.6) supplemented with 0.1% (v/v) Tween-20 (TBST) at room
temperature for 1 h. Following washing with TBST, the PVDF
membranes were incubated with 1:500 diluted anti-FOXP3 antibody
(cat. no. ab4728; Abcam, Cambridge, MA, USA) overnight at 4°C.
Membranes were washed with TBST and subsequently incubated with
horseradish peroxidase (HRP)-conjugated goat anti-rabbit antibody
(1:5,000; cat. no. ab205718; Abcam) for 1 h at room temperature;
protein bands were visualized with the Enhanced Chemiluminescence
(ECL) detection reagents (Millipore). Membranes were then stripped
and reblotted with 1:5,000 diluted anti-β-actin antibody (cat. no.
A5316; Sigma-Aldrich; Merck KGaA) at room temperature for 1 h.
Following TBST washing, blots were incubated with HRP-conjugated
goat anti-mouse secondary antibody (1:5,000; cat. no. ab205719;
Abcam) for 1 h at room temperature; the blots were visualized with
the ECL detection reagents. Protein expression levels were
calculated as the relative band density to that of β-actin using
ImageJ software (National Institute of Health, Bethesda, MD,
USA).
Statistical analysis
Continuous variables are presented as mean ±
standard deviation. For continuous variables, inter-group
differences were compared with Student's t-test (for 2 groups) or
one-way analysis of variance (ANOVA; for ≥3 groups). For
significant differences in ANOVA, multiple comparisons were
conducted with the Dunnett's t-test. For categorical variables,
inter-group differences were compared with the χ2 test
or the Fisher's exact probability test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Patient characteristics
The clinical characteristics of the study
participants are presented in Table
I. The percentage of primiparas was similar in all study
groups. Maternal age was significantly higher among preeclamptic
patients compared with normal pregnant women. Gestational age was
significantly lower in preeclamptic patients than in normal
pregnant women. Systolic and diastolic blood pressures were
significantly higher in both preeclamptic groups as compared to
normal pregnant women. All other clinical features presented in
Table I differed significantly
among groups. Fetal growth restriction was absent in normal
pregnant women but observed in 41.4% of preeclamptic women.
| Table I.Clinicopathological characteristics
of the patients used in the present study. |
Table I.
Clinicopathological characteristics
of the patients used in the present study.
| Clinicopathological
characteristic | Patients with
preeclampsia (n=29) | Normal pregnant
women (n=27) | Healthy
non-pregnant women (n=10) | ANOVA F value;
P-value |
|---|
| Maternal age at
delivery (years) | 31±7a,b | 26±6 | 27±7 | 4.251; 0.015 |
| BMI at blood draw
(kg/m2) |
28.7±6.3a,b |
26.0±5.9b | 21.6±1.6 | 4.912; 0.001 |
| Gestational age at
delivery (weeks) | 37±2.1a | 39±2.2 | n.a. | 2.376;
<0.001 |
| Primiparous women
(%) | 17 (58.6%) | 16 (59.25%) | n.a. | 0.002; 0.961 |
| Systolic blood
pressure (mmHg) | 155±49a,b | 119±30 | 115±36 | 6.814; 0.001 |
| Diastolic blood
pressure (mmHg) | 100±36a,b | 77±26 | 74±33 | 4.593; 0.010 |
| Fetal birth weight
(g) |
2,932±378c | 3,212±212 | n.a. | 2.563; 0.013 |
| Fetal growth
restriction (%) | 12
(41.4%)a | 0 | n.a. | <0.001 |
Cytokine levels
Regulatory T cells are well known to suppress
effector activity of T cells. We then aimed to determine cytokine
production in the patients with PE. Plasma production of IL-6,
IL-17 and TGF-β1 was significantly higher in preeclamptic patients
compared with these levels in normal pregnant women (P=0.0021,
P=0.015 and P=0.019, respectively; Fig. 1A-C). Accordingly, levels of Treg
cytokines such as IL-10 were significantly lower in preeclamptic
women than in normal pregnant women (P=0.0012; Fig. 1D). Levels of serum IL-17, TGF-β1
and IL-10 were similar in healthy non-pregnant and normal pregnant
women (P=0.085, P=0.163 and P=0.248, respectively; Fig. 1B-D). However, levels of serum IL-6
were significantly higher in healthy non-pregnant women than in
normal pregnant women (P=0.008; Fig.
1A).
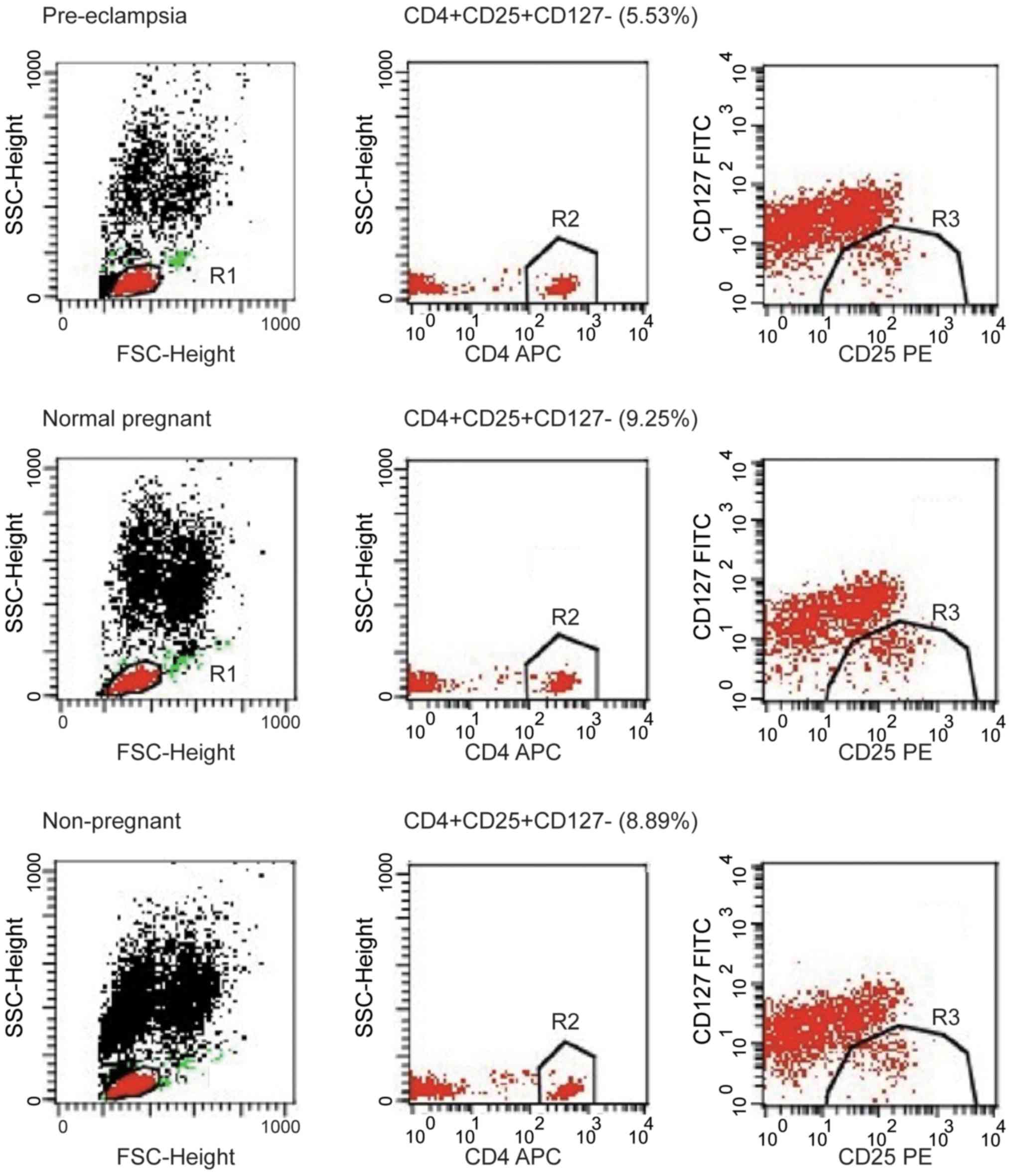
Decreased frequency of Tregs in
preeclampsia patient
Tregs are key players in successful pregnancy and
their deficiencies are implicated in complications in pregnancy
such as PE, but the results are inconsistent among studies.
Although the percentage of CD4+CD25+ cells
among the CD4+ T cells was notably higher in the normal
pregnancy group compared to the percentage in the healthy
non-pregnant women (46.27±1.11 vs. 42.71±1.05%), the difference was
not significant; the percentage of CD4+CD25+
cells among the CD4+ T cells were significantly lower in
women with PE compared with normal pregnant subjects and healthy
non-pregnant subjects (both P<0.01; Fig. 2A). The percentage of
CD4+CD25+CD127− cells among the
CD4+ T cells was analyzed in peripheral blood of
patients, and the percentage of
CD4+CD25+CD127− cells among the
CD4+ T cells was significantly lower in the peripheral
blood of patients with PE compared with normal pregnant women
(6.34±0.28 vs. 9.10±0.28%; P<0.01; Fig. 2B). The percentage of
CD4+CD25+CD127− cells among the
CD4+ T cells was significantly lower in the peripheral
blood of PE compared with healthy non-pregnant women (6.34±0.28 vs.
8.75±0.54%; P<0.01). However, the proportion of
CD4+CD25+CD127− cells among the
CD4+ T cells was similar in healthy non-pregnant and
normal pregnant women, indicating that a decrease in
CD4+CD25+CD127− cells was
associated with PE. These findings suggest that the proportion of
Tregs in peripheral blood was increased in normal pregnant subjects
but decreased in women with PE. The number of
CD4+CD25+CD127− T cell population
decreased in peripheral blood of PE patients. The image shows the
typical data of the T cell through flow cytometry (Fig. 3). The peripheral blood of patients
with PE exhibited a decline in the population of
CD4+CD25+ T cells; typical T cell data from
flow cytometry are presented in Fig.
4.
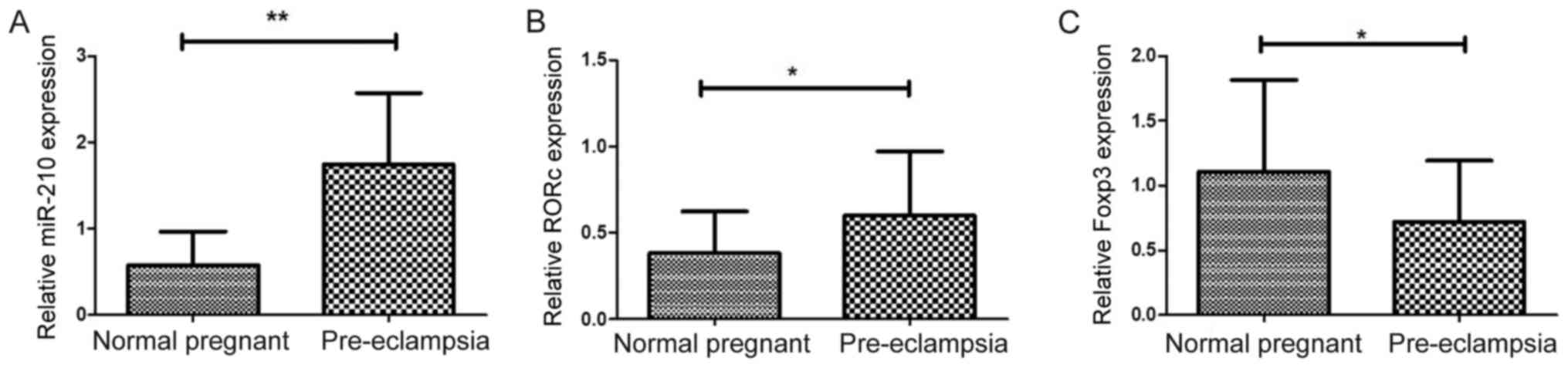
Expression of Foxp3, RORc and miR-210
in placenta
The mRNA expression levels of transcription factors
associated with Treg/Th17 cells were analyzed in placentas from
patients with PE and normal pregnant women. miR-210 was previously
reported to target Foxp3 and inhibit Treg cell function in patients
with psoriasis (37). In the
present study, significantly higher expression of miR-210 was
observed in patients with PE compared with normal pregnant women
(1.744±0.153 vs. 0.578±0.744; P<0.01; Fig. 5A). In addition, mRNA expression
levels of RORc were significantly higher in women with PE compared
with normal pregnant women (0.599±0.069 vs. 0.380±0.046; P=0.0127;
Fig. 5B). Higher expression of
RORc may promote enhanced production on inflammatory cytokines
(20). The results demonstrated
that mRNA expression of Foxp3 was significantly lower in
preeclamptic women compared with normal pregnant women (0.713±0.088
vs. 1.105±0.136; P=0.019; Fig.
5C), which was consistent with decreased Tregs in patients with
PE.
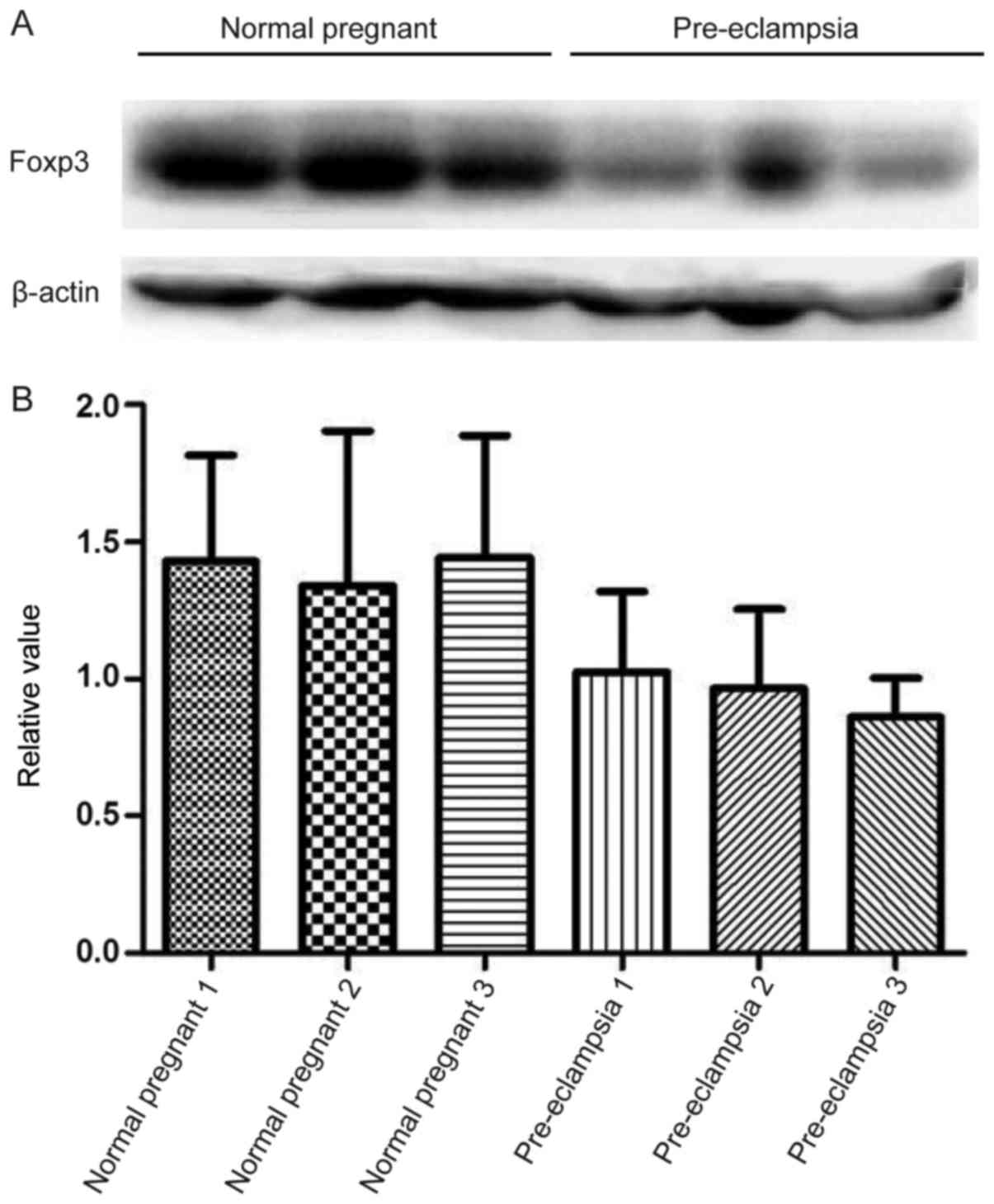
Expression of inflammation-related
protein Foxp3
Consistent with mRNA expression results, lower
protein expression levels of Foxp3 was observed in patients with PE
compared with normal pregnant subjects; however, this difference
was not indicated to be significant (Fig. 6).
Discussion
Foxp3 is a member of the fork head transcription
factor family and is exclusively expressed in
CD4+CD25+ Tregs (41). CD4+CD25+
Tregs are a subset of mature T cells that play an important role in
maintaining immune homeostasis and preventing autoimmune disease.
Patients with preeclampsia (PE) have been shown to have fewer
active CD4+CD25+ Tregs in peripheral blood
and related tissues (42).
The TGF-β1 signaling pathway is involved in the
induction of Foxp3 as well as the generation of peripheral T cells
and thymocytes. Previous research has revealed a reduced number of
CD4+CD25+Foxp3+ Tregs in the
peripheral blood of women with PE compared to normal pregnant
women. CD4+CD25+Foxp3+ Tregs
express specific anti-inflammatory cytokines such as IL-10, which
dampen an excessive effector immune response (43,44).
In the absence of any inflammatory insult, TGF-β
produced in the immune system will suppress the generation of
effector T cells and induce Foxp3+ Tregs, thus,
maintaining self-tolerance. However, upon infection or
inflammation, IL-6 produced by the activated innate immune system
will suppress the generation of TGF-β-induced Tregs and induce a
pro-inflammatory T-cell response (predominately Th17 cells)
(45). These results support the
results of recent studies showing an increase in circulating levels
of IL-17 in pregnant women with PE when compared to healthy
pregnant women and non-pregnant women (46).
The results obtained for healthy non-pregnant women
in this study are consistent with those of previous studies, which
report a higher prevalence of peripheral blood IL-17 in
preeclamptic patients (21,47,48).
TGF-β1 may act as a potent inhibitor of T/B cell proliferation and
auto-antibody production (49).
TGF-β1 also contributes to suppression of the T cell response by
regulating Foxp3+ Tregs and is essential to the
development of natural Tregs (nTregs) and induced Tregs (iTregs)
(50). nTregs are produced in the
thymus early in life, and iTregs are generated in the periphery
from naive CD4+ T cells (51). Tregs maintain peripheral tolerance
through both cell-contact-dependent and cell-contact-independent
mechanisms (52).
In the present study, we measured serum levels of
cytokines (IL-6, IL −10, IL-17 and TGF-β1) in healthy non-pregnant
women, normal pregnant women and preeclamptic patients. The results
showed that serum IL-17, IL-6 and TGF-β1 levels were significantly
higher in preeclamptic patients than in normal pregnant women.
Levels of Treg cytokines IL-10 were significantly lower in
preeclamptic women than in normotensive women.
To further investigate the mechanisms resulting in
deficient Tregs in PE, we measured mRNA expression levels of
transcription factors that play an important role in the
proliferation and differentiation of Tregs in PE. Foxp3 is known as
a specific molecular marker for Tregs and is indispensable to the
development and function of Tregs (53).
In the present study, the authors suggest that
peripheral CD4+CD25+ Tregs cannot be
accurately identified and purified based on surface expression of
CD127 (as an alternative to expression of transcription factor
Foxp3) (54). Therefore, low
expression of the CD127 molecule is not an inherent attribute of
Tregs (55).
In the present study, cytofluorometry showed that
the percentage of CD4+CD25+CD127−
Tregs among CD4+ T cells in peripheral blood was reduced
in patients with PE compared to normal pregnant women and healthy
non-pregnant women. Furthermore, the percentage of
CD4+CD25+ Tregs was significantly lower in
the peripheral blood of patients with PE compared to normal
individuals.
Such tissue-specific patterns of gene expression are
regulated by miRNAs. In this study, the mRNA expression of Foxp3
was significantly lower in placenta from patients with PE. The mRNA
expression of miR-210 showed a significant increase in PE.
Furthermore, protein levels of Foxp3 were notably decreased in
patients with PE, compared to normal pregnant subjects. These
differences in miRNA expression are important as miRNAs (non-coding
RNAs approximately 19–25 nucleotides in length) regulate nearly
30–50% of all genes by binding to target protein-encoding
mRNAs.
In conclusion, the results from our experiments
demonstrated that serum IL-10 levels are decreased in women with
PE. This effect may contribute to the development of excessive
systemic inflammation, as is characteristic of maternal PE. The
findings presented above show a decreased number of circulating
CD4+CD25+CD127− Tregs in women
with PE compared with normal pregnant women. Placenta from patients
with PE showed significantly decreased mRNA expression of Foxp3 and
significantly increased expression of miR-210. The existence of a
positive correlation between expression of miR-210 and Foxp3
protein in PE requires further experimental verification.
Additional studies are required to confirm and extend these
findings.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from The
2016 Shenzhen Science and Technology R&D Fundamental Research:
Medical and Health Free Exploration (grant no.
JCYJ20160428140315277) and The 2016 Shenzhen Municipal Health and
Family Planning Commission (grant no. 201606027).
Availability of data and materials
The data sets used and/or analysed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
JC designed the experiments and wrote the initial
draft of the manuscript. LZ, DW, YX, HG and WT conducted the
experiments and analyzed the data. CW analyzed and interpreted the
data, and drafted the manuscript.
Ethics approval and consent to
participate
All experimental research on human samples followed
the guidelines and approvals of Shenzhen Longhua District Central
Hospital Ethics Committee (Shenzhen, China). The Ethics Committee
of the Shenzhen Longhua District Central Hospital approved the
study protocol. All patients provided written informed consent
prior to the participation in the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interest.
References
|
1
|
Abalos E, Cuesta C, Grosso AL, Chou D and
Say L: Global and regional estimates of preeclampsia and eclampsia:
A systematic review. Eur J Obstet Gynecol Repro Biol. 170:1–7.
2013. View Article : Google Scholar
|
|
2
|
Moore-Maxwell CA and Robboy SJ: Placental
site trophoblastic tumor arising from antecedent molar pregnancy.
Gynecol Oncol. 92:708–712. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ilekis JV, Reddy UM and Roberts JM:
Preeclampsia-A pressing problem: An executive summary of a national
institute of child health and human development workshop. Reprod
Sci. 14:508–523. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lok CA, Jebbink J, Nieuwland R, Faas MM,
Boer K, Sturk A and Van Der Post JA: Leukocyte activation and
circulating leukocyte-derived microparticles in preeclampsia. Am J
Reprod Immunol. 61:346–359. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sakaguchi S, Sakaguchi N, Asano M, Itoh M
and Toda M: Immunologic self-tolerance maintained by activated T
cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a
single mechanism of self-tolerance causes various autoimmune
diseases. J Immunol. 155:1151–1164. 1995.PubMed/NCBI
|
|
6
|
Aluvihare VR, Kallikourdis M and Betz AG:
Regulatory T cells mediate maternal tolerance to the fetus. Nat
Immuno. 5:266–271. 2004. View
Article : Google Scholar
|
|
7
|
Xiong YH, Yuan Z and He L: Effects of
estrogen on CD4+ CD25+ regulatory T cell in
peripheral blood during pregnancy. Asian Pac J Trop Med. 6:748–752.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Heikkinen J, Mottonen M, Alanen A and
Lassila O: Phenotypic characterization of regulatory T cells in the
human decidua. Clin Exp Immunol. 136:373–378. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zenclussen AC, Gerlof K, Zenclussen ML,
Sollwedel A, Bertoja AZ, Ritter T, Kotsch K, Leber J and Volk HD:
Abnormal T-cell reactivity against paternal antigens in spontaneous
abortion: Adoptive transfer of pregnancy-induced
CD4+CD25+ T regulatory cells prevents fetal
rejection in a murine abortion model. Am J Pathol. 166:811–822.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sasaki Y, Darmochwal-Kolarz D, Suzuki D,
Sakai M, Ito M, Shima T, Shiozaki A, Rolinski J and Saito S:
Proportion of peripheral blood and decidual CD4+
CD25bright regulatory T cells in pre-eclampsia. Clin Exp
Immunol. 149:139–145. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Darmochwal-Kolarz D, Saito S, Rolinski J,
Tabarkiewicz J, Kolarz B, Leszczynska-Gorzelak B and Oleszczuk J:
Activated T lymphocytes in pre-eclampsia. Am J Reprod Immunol.
58:39–45. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nakashima A, Shima T, Inada K, Ito M and
Saito S: The balance of the immune system between T cells and NK
cells in miscarriage. Am J Reprod Immunol. 67:304–310. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Koucky M, Malickova K, Cindrova-Davies T,
Germanova A, Parizek A, Kalousova M, Hajek Z and Zima T: Low levels
of circulating T-regulatory lymphocytes and short cervical length
are associated with preterm labor. J Reprod Immunol. 106:110–117.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xiong H, Zhou C and Qi G: Proportional
changes of CD4+CD25+Foxp3+
regulatory T cells in maternal peripheral blood during pregnancy
and labor at term and preterm. Clin Invest Med. 33:E4222010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sakaguchi S, Ono M, Setoguchi R, Yagi H,
Hori S, Fehervari Z, Shimizu J, Takahashi T and Nomura T:
Foxp3+ CD25+ CD4+ natural
regulatory T cells in dominant self-tolerance and autoimmune
disease. Immunol Rev. 212:8–27. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sasaki Y, Sakai M, Miyazaki S, Higuma S,
Shiozaki A and Saito S: Decidual and peripheral blood
CD4+CD25+ regulatory T cells in early
pregnancy subjects and spontaneous abortion cases. Mol Hum Reprod.
10:347–353. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cristofalo R, Bannwart-Castro CF,
Magalhaes CG, Borges VT, Peracoli JC, Witkin SS and Peracoli MT:
Silibinin attenuates oxidative metabolism and cytokine production
by monocytes from preeclamptic women. Free Radic Res. 47:268–275.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Peracoli JC, Bannwart-Castro CF, Romao M,
Weel IC, Ribeiro VR, Borges VT, Rudge MV, Witkin SS and Peracoli
MT: High levels of heat shock protein 70 are associated with
pro-inflammatory cytokines and may differentiate early- from
late-onset preeclampsia. J Reprod Immunol. 100:129–134. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Figueiredo AS and Schumacher A: The T
helper type 17/regulatory T cell paradigm in pregnancy. Immunology.
148:13–21. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Darmochwal-Kolarz D, Kludka-Sternik M,
Tabarkiewicz J, Kolarz B, Rolinski J, Leszczynska-Gorzelak B and
Oleszczuk J: The predominance of Th17 lymphocytes and decreased
number and function of Treg cells in preeclampsia. J Reprod
Immunol. 93:75–81. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Santner-Nanan B, Peek MJ, Khanam R,
Richarts L, Zhu E, Fazekas de St Groth B and Nanan R: Systemic
increase in the ratio between Foxp3+ and IL-17-producing
CD4+ T cells in healthy pregnancy but not in
preeclampsia. J Immunol. 183:7023–7030. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Alijotas-Reig J, Llurba E and Gris JM:
Potentiating maternal immune tolerance in pregnancy: A new
challenging role for regulatory T cells. Placenta. 35:241–248.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang Z, Liu H, Shi Y, Xu N, Wang Y, Li A
and Song W: Increased circulating Th22 cells correlated with Th17
cells in patients with severe preeclampsia. Hypertens Pregnancy.
36:100–107. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu W, Putnam AL, Xu-Yu Z, Szot GL, Lee
MR, Zhu S, Gottlieb PA, Kapranov P, Gingeras TR, Fazekas de St
Groth B, et al: CD127 expression inversely correlates with FoxP3
and suppressive function of human CD4+ T reg cells. J
Exp Med. 203:1701–1711. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Miyara M and Sakaguchi S: Human
FoxP3+CD4+ regulatory T cells: Their knowns
and unknowns. Immunol Cell Biol. 89:346–351. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Seddiki N, Santner-Nanan B, Martinson J,
Zaunders J, Sasson S, Landay A, Solomon M, Selby W, Alexander SI,
Nanan R, et al: Expression of interleukin (IL)-2 and IL-7 receptors
discriminates between human regulatory and activated T cells. J Exp
Med. 203:1693–1700. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Aerts NE, Dombrecht EJ, Ebo DG, Bridts CH,
Stevens WJ and De Clerck LS: Activated T cells complicate the
identification of regulatory T cells in rheumatoid arthritis. Cell
Immunol. 251:109–115. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huntzinger E and Izaurralde E: Gene
silencing by microRNAs: Contributions of translational repression
and mRNA decay. Nat Rev Genet. 12:99–110. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Morales-Prieto DM, Ospina-Prieto S,
Chaiwangyen W, Schoenleben M and Markert UR: Pregnancy-associated
miRNA-clusters. J Reprod Immunol. 97:51–61. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xu X, Kriegel AJ, Liu Y, Usa K, Mladinov
D, Liu H, Fang Y, Ding X and Liang M: Delayed ischemic
preconditioning contributes to renal protection by upregulation of
miR-21. Kidney Int. 82:1167–1175. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lin Z, Murtaza I, Wang K, Jiao J, Gao J
and Li PF: miR-23a functions downstream of NFATc3 to regulate
cardiac hypertrophy. Proc Natl Acad Sci USA. 106:12103–12108. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shen XC, Lu Y and Qian ZY: Effects of
crocetin on the matrix metalloproteinases in cardiac hypertrophy
induced by norepinephrine in rats. J Asian Nat Prod Res. 8:201–208.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bentwich I, Avniel A, Karov Y, Aharonov R,
Gilad S, Barad O, Barzilai A, Einat P, Einav U, Meiri E, et al:
Identification of hundreds of conserved and nonconserved human
microRNAs. Nat Genet. 37:766–770. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pineles BL, Romero R, Montenegro D, Tarca
AL, Han YM, Kim YM, Draghici S, Espinoza J, Kusanovic JP, Mittal P,
et al: Distinct subsets of microRNAs are expressed differentially
in the human placentas of patients with preeclampsia. Am J Obstet
Gynecol. 196:e261–e266. 2007. View Article : Google Scholar
|
|
35
|
Chim SS, Shing TK, Hung EC, Leung TY, Lau
TK, Chiu RW and Lo YM: Detection and characterization of placental
microRNAs in maternal plasma. Clin Chem. 54:482–490. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mouillet JF, Chu T, Hubel CA, Nelson DM,
Parks WT and Sadovsky Y: The levels of hypoxia-regulated microRNAs
in plasma of pregnant women with fetal growth restriction.
Placenta. 31:781–784. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhao M, Wang LT, Liang GP, Zhang P, Deng
XJ, Tang Q, Zhai HY, Chang CC, Su YW and Lu QJ: Up-regulation of
microRNA-210 induces immune dysfunction via targeting FOXP3 in
CD4+ T cells of psoriasis vulgaris. Clin Immunol.
150:22–30. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Auerbach L, Hafner T, Huber JC and Panzer
S: Influence of low-dose oral contraception on peripheral blood
lymphocyte subsets at particular phases of the hormonal cycle.
Fertil Steril. 78:83–89. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
No authors listed, : Report of the
national high blood pressure education program working group on
high blood pressure in pregnancy. Am J Obstet Gynecol. 183:S1–S22.
2000. View Article : Google Scholar
|
|
40
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCt method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Redman CW, Sacks GP and Sargent IL:
Preeclampsia: An excessive maternal inflammatory response to
pregnancy. Am J Obstet Gynecol. 180:499–506. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Walker MR, Kasprowicz DJ, Gersuk VH,
Benard A, Van Landeghen M, Buckner JH and Ziegler SF: Induction of
FoxP3 and acquisition of T regulatory activity by stimulated human
CD4+CD25 T cells. J Clin Invest. 112:1437–1443. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jianjun Z, Yali H, Zhiqun W, Mingming Z
and Xia Z: Imbalance of T-cell transcription factors contributes to
the Th1 type immunity predominant in pre-eclampsia. Am J Reprod
Immunol. 63:38–45. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Saito S, Nakashima A, Shima T and Ito M:
Th1/Th2/Th17 and regulatory T-cell paradigm in pregnancy. Am J
Reprod Immunol. 63:601–610. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bettelli E, Carrier Y, Gao W, Korn T,
Strom TB, Oukka M, Weiner HL and Kuchroo VK: Reciprocal
developmental pathways for the generation of pathogenic effector
TH17 and regulatory T cells. Nature. 441:235–238. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Molvarec A, Czegle I, Szijarto J and Rigo
J Jr: Increased circulating interleukin-17 levels in preeclampsia.
J Reprod Immunol. 112:53–57. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Szarka A, Rigo J Jr, Lazar L, Beko G and
Molvarec A: Circulating cytokines, chemokines and adhesion
molecules in normal pregnancy and preeclampsia determined by
multiplex suspension array. BMC Immunol. 11:592010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Toldi G, Rigo J Jr, Stenczer B, Vasarhelyi
B and Molvarec A: Increased prevalence of IL-17-producing
peripheral blood lymphocytes in pre-eclampsia. Am J Reprod Immunol.
66:223–229. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cross D and Cambier JC: Transforming
growth factor beta 1 has differential effects on B cell
proliferation and activation antigen expression. J Immunol.
144:432–439. 1990.PubMed/NCBI
|
|
50
|
Travis MA and Sheppard D: TGF-β activation
and function in immunity. Annu Rev Immunol. 32:51–82. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Bilate AM and Lafaille JJ: Induced
CD4+Foxp3+ regulatory T cells in immune
tolerance. Annu Rev Immunol. 30:733–758. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Josefowicz SZ, Lu LF and Rudensky AY:
Regulatory T cells: Mechanisms of differentiation and function.
Annu Rev Immunol. 30:531–564. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Cretney E, Kallies A and Nutt SL:
Differentiation and function of Foxp3 effector regulatory T cells.
Trends Immunol. 34:74–80. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Klein S, Kretz CC, Krammer PH and Kuhn A:
CD127low/− and FoxP3+ expression levels
characterize different regulatory T-cell populations in human
peripheral blood. J Invest Dermatol. 130:492–499. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Mazzucchelli R, Hixon JA, Spolski R, Chen
X, Li WQ, Hall VL, Willette-Brown J, Hurwitz AA, Leonard WJ and
Durum SK: Development of regulatory T cells requires IL-7Rα
stimulation by IL-7 or TSLP. Blood. 112:3283–3292. 2008. View Article : Google Scholar : PubMed/NCBI
|