Introduction
Type 2 diabetes mellitus (T2DM) is characterized by
loss of β-cell mass, dysfunction of β-cells and increased β-cell
apoptosis (1). Islet pathology in
T2DM is characterized by the accumulation of extracellular islet
amyloid deposits derived from islet amyloid polypeptide (IAPP),
which is also known as amylin (2).
IAPP, a 4-kDa peptide hormone composed of 37 amino acids, is
synthesized and co-secreted from pancreatic β-cells along with
insulin. Although extracellular islet amyloid is relatively inert,
the intracellular membrane-permeant toxic IAPP oligomers that form
within β-cells in T2DM are thought to induce β-cell dysfunction and
apoptosis (3,4). A previous study investigating the
mechanisms involved in the human IAPP (hIAPP)-induced onset of the
multidimensional pathogenic response in β-cells suggested that
impaired autophagy, endoplasmic reticulum (ER) stress, reactive
oxygen species activation, membrane disruption and
receptor-mediated signal transduction cascades may serve a role in
this process (5).
Glucagon-like peptide-1 (GLP-1) is a
gastrointestinal hormone primarily secreted by L cells in the
intestine in response to food intake. GLP-1 has been shown to
ameliorate hyperglycemia by augmenting β-cell insulin secretion and
by imposing a low risk of induced hypoglycemia (6). An increasing number of studies have
revealed that GLP-1 exhibits beneficial pleiotropic effects in
β-cells, including attenuating oxidative stress (7), the inflammatory response (8) and unbalanced autophagy (9), and enhancing cellular proliferation
and survival (10).
Amylin and structurally related calcitonin
gene-related peptide (CGRP) restrain the stimulatory effect of
GLP-1 (7–36) amide (11). GLP-1 protects the brain from
β-amyloid peptide toxicity (12,13).
Furthermore, Park et al (14) revealed that a GLP-1 receptor
agonist may preserve β-cells by restoring impaired pro-hIAPP
processing and reducing hIAPP aggregation. However, further
investigation into the exendin-4-mediated protective effects on
hIAPP-induced damage in pancreatic β-cells is still required
(15,16). In addition, it is also thought that
complex interactions may exist between GLP-1 and IAPP.
Autophagy is an intracellular process characterized
by dynamic rearrangement of subcellular membranes that sequester
cytoplasm, protein aggregates, pathogens and organelles for
delivery to vacuoles and lysosomes, where the components are
degraded and recycled (17).
Autophagy is important for maintaining protein quality, which is
crucial for diverse biological processes (17,18).
Several studies have revealed that human IAPP induces β-cell mass
loss by targeting autophagy (19,20);
however, it is not known whether hIAPP-induced islet β-cell
toxicity attenuation by GLP-1 is associated with alterations in
autophagy.
The aim of the present study was to elucidate the
hIAPP secretion patterns and insulin stimulatory effects of
exendin-4 in MIN6 cells and to determine whether a GLP-1 receptor
agonist has anti-autophagic and anti-apoptotic effects on hIAPP
overexpression in MIN6 cells.
Materials and methods
Cell culture and transfection
MIN-6 cells, purchased from China Center for Type
Culture Collection (Wuhan, China) were cultured in Dulbecco's
modified Eagle's medium containing 25 mM glucose (Thermo Fisher
Scientific, Inc., Waltham, MA, USA) with 10% fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml penicillin
(Beyotime Institute of Biotechnology, Shanghai, China) and 100
mg/ml streptomycin-sulfate (Beyotime Institute of Biotechnology,
Jiangsu, China) at 37°C and 100% humidified air containing 5%
CO2 until 80–90% confluence was reached.
Cell transfection was performed using Lipofectamine
3000™ (Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. Briefly, cells were seeded at
1×106 cells/well in a 6-well plate with 2 ml of growth
medium without antibiotics at 37°C. Once 70% confluence was
reached, MIN-6 cells were divided into three groups according to
transfection: hIAPP, blank plasmid and control group without
plasmid. In hIAPP or blank plasmid group, cells were transfected
with 2.5 µg of hIAPP plasmid or empty plasmid DNA (Beyotime
Institute of Biotechnology, Jiangsu, China) and 7.5 µl of
Lipofectamine® 3000 (Beyotime Institute of
Biotechnology, Jiangsu, China), respectively. In the control group,
cells were culture in growth medium without antibiotics. After 8 h
at 37°C, the medium was changed to growth medium containing
antibiotics. At 24 h post-transfection, MIN-6 cells were further
cultured at 37°C in 6-well plates until 70% confluence was reached.
Then, 100 nM Exendin4 was added to the culture medium for 24 h.
Untransfected MIN-6 cells treated with rapamycin only were used as
positive controls.
Determination of insulin and IAPP
levels by enzyme-linked immunosorbent assay (ELISA)
Insulin and amylin levels in MIN-6 cells
supernatants were determined using a mouse ELISA kit (cat. nos.
BPE20353 and BPE20356) purchased from Shanghai Langton
Biotechnology Co. Ltd. (Shanghai, China) according to the
manufacturer's instructions. Absorbance at 450 nm was measured
using an ELX800 Universal Microplate Reader (BioTek Instruments,
Inc., Winooski, VT, USA). A standard curve was obtained using
purified proteins supplied with the ELISA kit.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from cells using TRIzol
(Invitrogen; Thermo Fisher Scientific, Inc.). cDNA was reverse
transcribed with a SYBR Premix Ex Taq II RT kit from Takara Bio,
Inc. (Otsu, Japan). The RT reaction was conducted under the
following conditions: 37°C for 15 min and 85°C for 5 sec; then held
at 4°C. The qPCR was performed on a CFX96 instrument (Bio-Rad
Laboratories, Inc.) with SYBR Premix Ex Taq (Takara Bio, Inc.,
Otsu, Japan). The primers used to amplify insulin, IAPP and β-actin
genes, were synthesized by Invitrogen (Thermo Fisher Scientific,
Inc.): Mouse insulin sense, 5′-TGTTGGTGCACTTCCTACCC-3′ and
antisense, 5′-ACACACCAGGTAGAGAGCCT-3′; mouse IAPP sense,
5′-AGATGGACAAACGGAAGTGC-3′ and antisense 5′-TTGGTTGGTGGGAGGACTG-3′;
and mouse β-actin sense 5′-GGGAAATCGTGCGTGAC-3′ and antisense
5′-AGGCTGGAAAAGAGCCT-3′. qPCR was conducted under the following
conditions: 95°C for 5 min, followed by 40 cycles of 95°C for 5
sec, 60°C for 30 sec and 72°C for 10 sec. The melting procedure was
as follows: 30 sec at 95°C, followed by 40 cycles of 15 sec at 95°C
and 60 sec at 60°C. Gene expression was quantified using the
2−∆∆Cq method (21);
the relative amount of each gene was normalized to that of β-actin,
and the result is expressed as a ratio of the relative amount.
Methyl thiazolyl tetrazolium (MTT)
assay
MIN6 cells were seeded (4×103/well) in
96-well plates and incubated at 37°C for 24 h. The cells were then
cultured with 25, 50 or 100 nM exendin-4 or an equal volume of DMSO
for 24 h and subjected to an MTT assay. An MTT (0.5 mg/ml) solution
prepared in phosphate-buffered saline was added to each well, and
the plates were then incubated at 37°C in a 5%
CO2-humidified atmosphere for 4 h. Subsequently, the
cells were treated with 150 µl DMSO, and the optical density was
detected with an ELX800 Universal Microplate Reader (BioTek
Instruments, Inc.) at a wavelength of 630 nm. Viability was defined
as the ratio (expressed as a percentage) of the absorbance of
exendin-4-treated cells to that of DMSO-treated cells.
Western blot analysis
Cells were harvested and lysed for protein isolation
with total protein extraction reagent (cat. no. AR0103; Boster
Biological Technology, Pleasanton, CA, USA). Protein concentration
was determined with the bicinchoninic acid protein assay. Protein
samples (50 µg/lane) were separated by 12% SDS-PAGE, then
electrotransferred onto polyvinylidene fluoride membranes (Bio-Rad
Laboratories, Inc.). Following preincubation in blocking buffer [5%
non-fat milk in Tris-buffered saline containing 0.05% Tween-20
(TBS-T)] for 1.5 h at room temperature, the membranes were
incubated with primary antibodies against sequestosome 1/p62
(1:1,000; cat. no. 8025, Cell Signaling Technology, Inc., Danvers,
MA, USA), light chain 3 (LC3, 1:1,000; cat. no. 3868; Cell
Signaling Technology, Inc.), IAPP (1:1,000; cat. no. PA5-29713;
Thermo Fisher Scientific, Inc.), cleaved caspase-3 (1:1,000; cat.
no. 9664; Cell Signaling Technology, Inc.) or β-actin (1:5,000;
cat. no. BS6007M; Bioworld Technology, Inc., St. Louis Park, MN,
USA) for 12 h at 4°C. The membranes were then washed in TBS-T and
incubated with horseradish peroxidase-conjugated secondary
antibodies (1:5,000; cat. nos. IgGA0208 and IgGA0216; Beyotime
Institute of Biotechnology) for 1–2 h at room temperature.
Reactions were visualized with enhanced chemiluminescence reagents
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). Densitometric
analysis was performed with by ImageJ version 1.46r (National
Institutes of Health, Bethesda, MD, USA).
Statistical analyses
Data are presented as the mean ± standard error of
the mean. Statistical significance was assessed by one-way analysis
of variance with a post-hoc Student-Newman-Keuls test for multiple
comparisons using GraphPad Pro software (GraphPad Software, Inc.,
La Jolla, CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
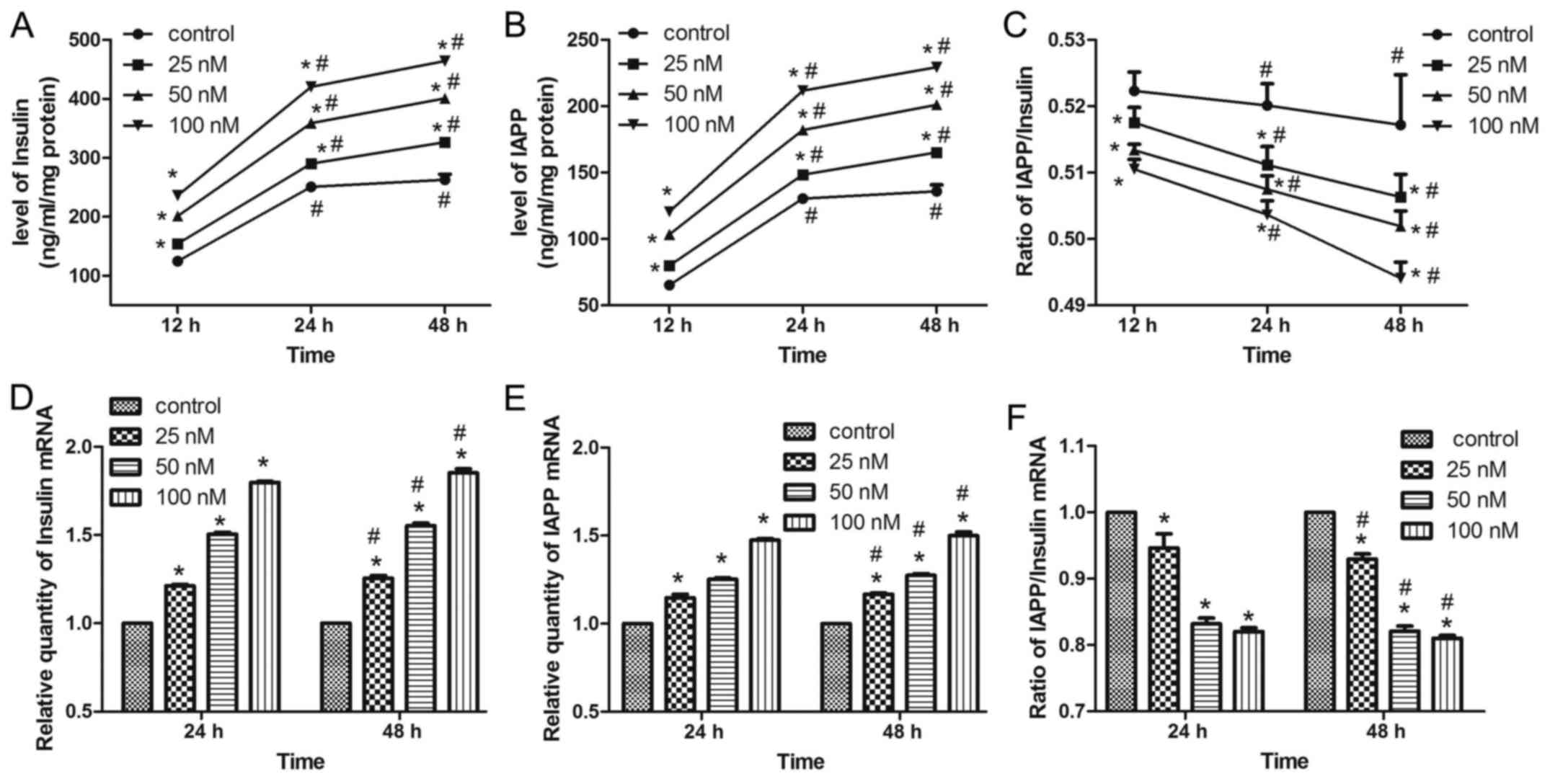
Insulin/IAPP secretion patterns of
MIN6 cells following exendin-4 pretreatment
As exendin-4 stimulates insulin secretion (22), alterations in the levels and
secretion patterns of insulin and IAPP in MIN6 cells were
investigated following treatment with the GLP-1 receptor agonist,
exendin-4. Insulin and IAPP secretion increased significantly in
time- and dose-dependent manners (Fig.
1A and B). As the increase in the level of insulin secretion
was greater than that of IAPP, the IAPP/insulin ratio decreased
with longer treatment times and higher concentrations of exendin-4
(Fig. 1C). In addition, the levels
of IAPP mRNA expression were significantly increased in a
concentration- and time-dependent manner; however, the IAPP/insulin
mRNA ratio decreased (Fig.
1D-F).
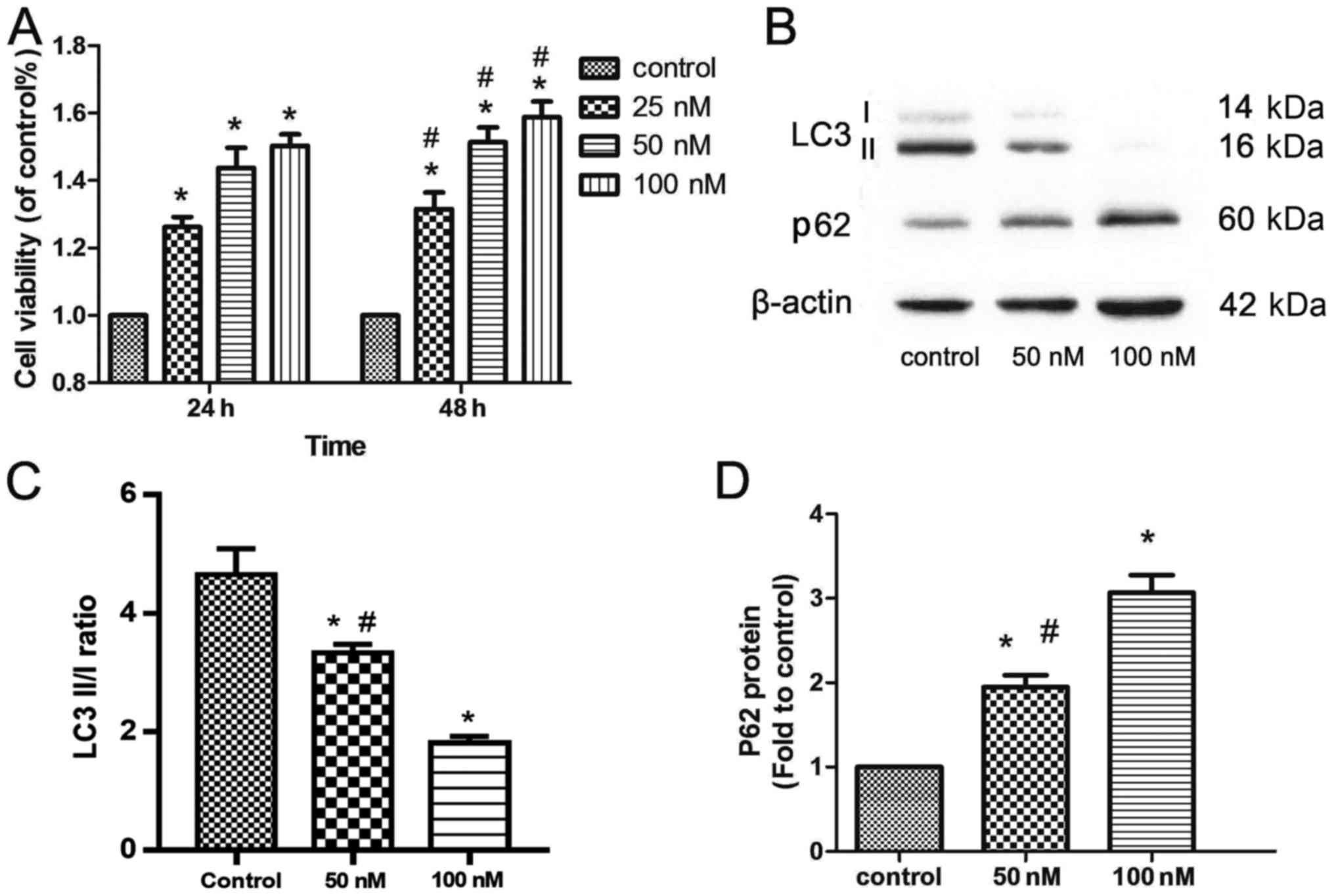
Cell viability
MTT assays were performed to examine the activities
of different exendin-4 concentrations (25, 50 or 100 nM) in MIN6
cells. As shown in Fig. 2A, cell
viability increased significantly in a time- and dose-dependent
manner (Fig. 2A).
Exendin-4 reduces autophagy in MIN6
cells
The impact of β-cell autophagy has been proposed to
be beneficial (23) and
detrimental (24) to cell
survival. To further characterize the positive effects of exendin-4
pretreatment on MIN6 cells, the present study assessed changes in
the protein levels the autophagy markers, LC3II/I and p62 (Fig. 2B). Exendin-4 pretreatment reduced
the LC3II/I ratio, which was accompanied by an increase in the
level of p62 (Fig. 2B-D).
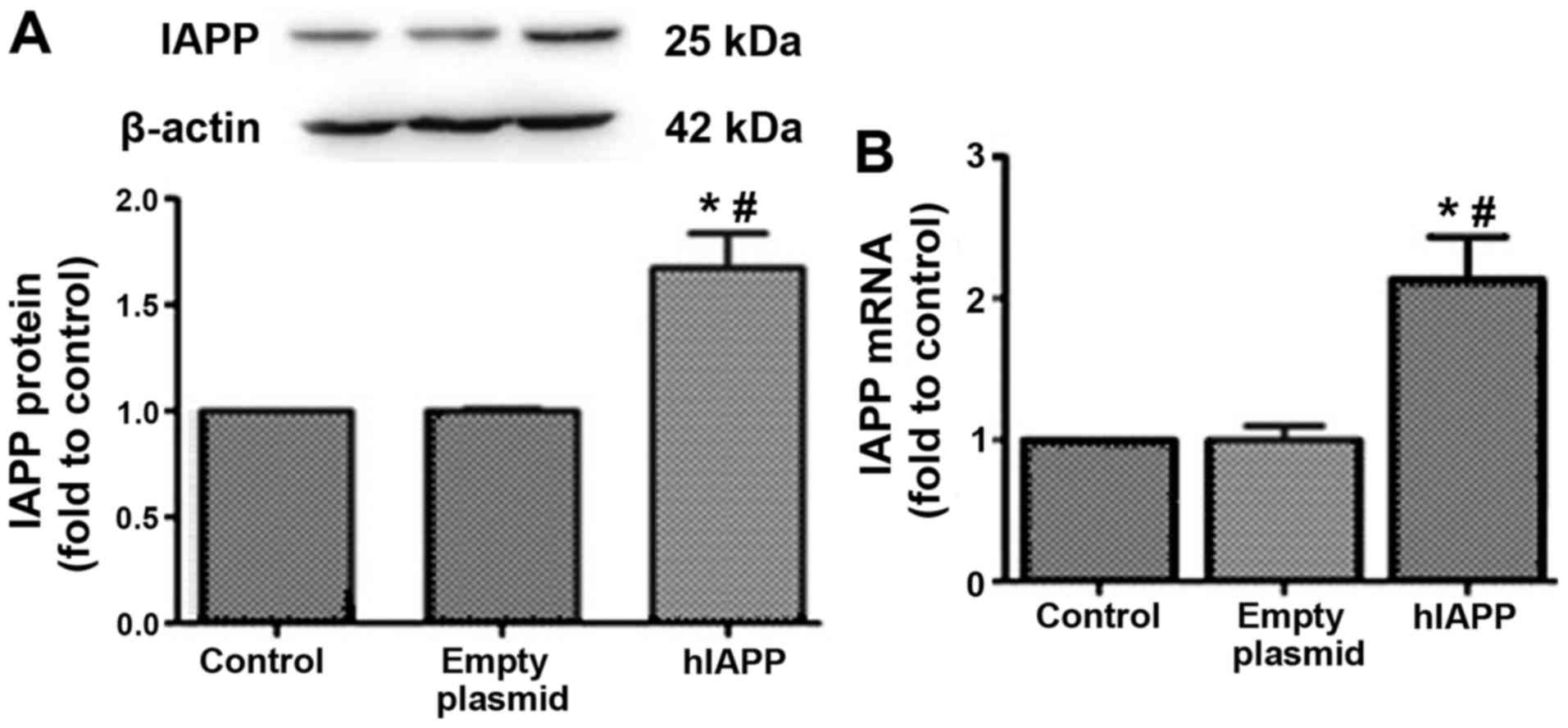
hIAPP-overexpression MIN6 cell
model
Exendin-4 has been reported to inhibit hIAPP-induced
insulinoma (INS-1E) cell death (15). To investigate the interactions
between GLP-1 and IAPP, a hIAPP-overexpression MIN6 cell model was
established (Fig. 3). The protein
and mRNA expression levels of hIAPP were significantly upregulated
following transfection with the hIAPP-overexpression plasmid into
MIN6 cells (Fig. 3).
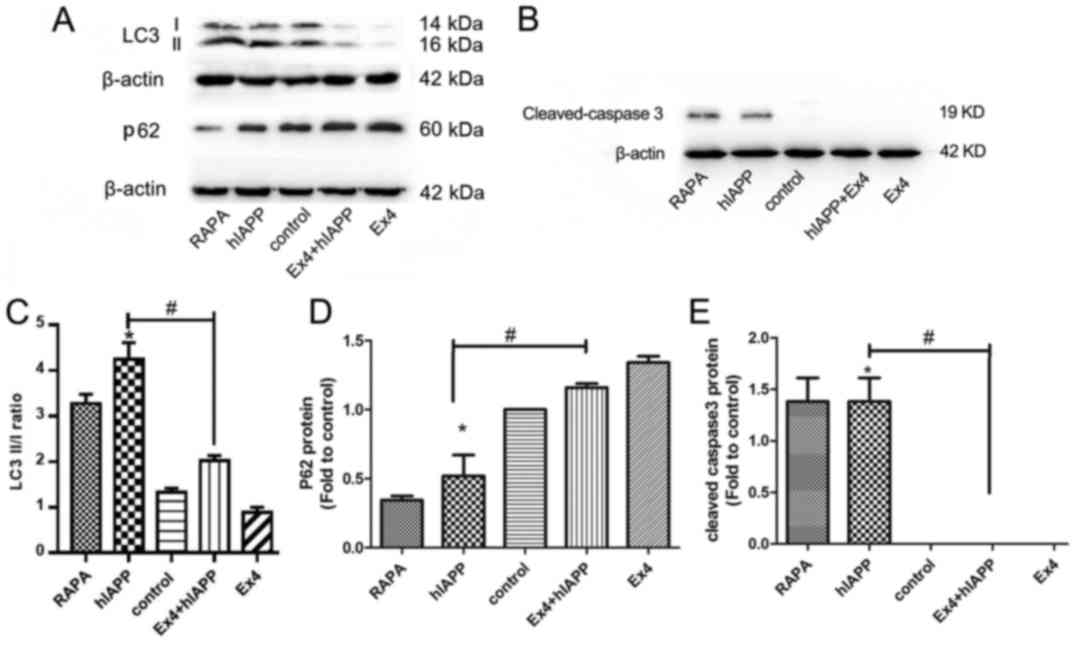
Exendin-4 protects MIN6 cells against
hIAPP-induced autophagy and apoptosis
As shown in Fig. 4A, C
and D, there was a significant increase in the LC3II/I ratio
and decrease in p62 expression in MIN6 cells overexpressing hIAPP;
these results were comparable to those achieved with treatment with
rapamycin, an autophagy inducer. However, autophagy activation was
significantly attenuated by exendin-4 pretreatment. Cleaved
caspase-3, the active form of caspase-3, was assessed to confirm
apoptosis. Cleaved caspase-3 levels were significantly increased in
hIAPP-overexpressing MIN6 cells when compared with control cells.
However, the increased level of cleaved caspase-3 was reversed by
exendin-4 pretreatment. These results indicated that hIAPP induced
autophagy and apoptosis, whereas exendin-4 attenuated these effects
(Fig. 4B and E).
Discussion
It is now widely accepted that β-cell failure,
including decreased β-cell mass and insulin secretion, leads to
T2DM. GLP-1 receptor agonist therapies have shown promising effects
in terms of positively affecting β-cell function and increasing
β-cell mass (20,22) however, the underlying mechanisms
remain unclear. In the present study, exendin-4 pretreatment
improved the insulin and IAPP secretion pattern and cell viability
in a time- and dose-dependent manner. The results support a novel
model in which hIAPP induces autophagy activation, leading to MIN6
cell apoptosis. Pretreatment with exendin-4 reversed hIAPP-induced
apoptosis and inhibited autophagy.
As IAPP is co-secreted with insulin, insulin
resistance causes IAPP overexpression, which may contribute to
amyloidogenesis. Xiao et al (25), demonstrated that short-term
exposure to high IAPP concentrations inhibited
glibenclamide-induced K (adenosine triphosphate) channel closure
and decreased calcium influx, which may ultimately lead to reduced
insulin secretion in INS-1 cells. Insulin has been shown to prevent
IAPP aggregation (26). In
addition, a change in the IAPP/insulin ratio, rather than an
increase in IAPP levels, is vital for amyloid formation (27). Circulating levels of GLP-1 directly
stimulate β-cell insulin secretion through protein kinase
C-dependent transient receptor potential cation channel subfamily M
member 4 (TRPM4) and TRPM5 activation (28). However, little is known about the
effects of GLP-1 receptor agonists on stimulating endogenously
released IAPP instead of insulin. By examining the
exendin-4-induced secretion pattern of IAPP and insulin in MIN6
cells incubated with 25 mM glucose, the present study revealed that
exendin-4 increased the protein and mRNA levels of insulin, as well
as those of IAPP, in a dose- and time-dependent manner. In
addition, the IAPP/insulin ratio decreased significantly with
increasing concentrations and time. These results indicated that
islet insulin release may be more sensitive than IAPP release is to
exendin-4 stimulation. According to the present results, beneficial
IAPP/insulin secretion patterns in MIN6 cells can be enhanced by
high exendin-4 concentrations and long incubation times.
Autophagy is a crucial regulator of pancreatic
β-cell homeostasis (29), and
impaired autophagic machinery may lead to β-cell dysfunction and
ultimately T2DM (30). The present
study revealed that there was inhibition of autophagy activation
with increasing concentrations of exendin-4 (50 and 100 nM) in MIN6
cells, and this inhibitory effect was also observed in β-cell
viability in a dose-dependent manner. Consistent with the present
results, a previous study reported that exendin-4 protects against
tacrolimus-induced pancreatic islet injury by regulating autophagy
clearance (10). Zhao et al
(31) also revealed that
liraglutide exerts a protective effect in the presence of high
glucose by inhibiting autophagy in HK-2 cells and in the kidneys of
diabetic rats. Nonetheless, another previous study proposed that by
activating autophagy, liraglutide exerts a protective effect on
glucolipotoxicity and lipotoxicity in INS-1E β-cells (32). These findings suggest that the
underlying mechanism of autophagic dysfunction under different
conditions may determine the impact of GLP-1 receptor agonists.
Shigihara et al (33), reported that increased insulin
resistance may enhance the toxic potential of hIAPP and ultimately
β-cell failure. Therefore, the present study evaluated the
pathogenic role of hIAPP on autophagy in an hIAPP-overexpression
MIN6 cell model as well as the possible reversion capacity of
exendin-4 pretreatment. The results indicated that exendin-4
pretreatment attenuated the hIAPP-induced increase in the LC3II/I
ratio and decrease in p62 expression, both of which are indicators
of autophagy. Previous studies have demonstrated that GLP-1 exerts
protective effects by inhibiting excessive autophagy (10,34,35).
All of these studies, taken together with the present results,
suggest that exendin-4 may exhibit protective effects by
attenuating hIAPP-induced autophagy in MIN6 cells.
GLP-1 receptor agonists have been shown in many
paradigms to enhance β-cell survival by decreasing apoptosis
(36–38). For example, an in vitro
study reported that GLP-1 receptor agonists protect β-cells from
injury-induced apoptosis, including cytokine-, ER stress- and
glucolipotoxicity-induced apoptosis (6). The present results indicated that
exendin-4 alleviated hIAPP-induced apoptosis to levels comparable
with those of the control group. These results are consistent with
our previous findings in clonal insulinoma cells (INS-1E cell line)
(15). Whether autophagy is the
driver of cell death or a pro-survival process in response to
certain stress conditions remains controversial. Although autophagy
was initially described as a cytoprotective response under nutrient
deprivation, a series of studies have suggested that autophagy
serves a vital role in promoting cell death, including apoptosis
(39–41). Future studies are required to
define the precise mechanisms underlying the effects of exendin-4
on autophagy and apoptosis.
Targeting autophagy and apoptosis inhibition
represents a novel therapeutic strategy against β-cell failure in
diabetic patients. However, the majority of traditional
anti-diabetic agents are unable to induce any direct
anti-autophagic or anti-apoptotic benefits in dysfunctional β-cells
during diabetes progression. Multiple mechanisms are involved in
the pathogenesis of hIAPP-induced β-cell failure, such as autophagy
and apoptosis (42,43). The results of the present study
further support the therapeutic potential of exendin-4 in treating
diabetic β-cell failure by attenuating hIAPP-induced autophagy and
apoptosis. With anti-autophagic and anti-apoptotic properties,
exendin-4 may gain more attention for β-cell failure treatment.
In conclusion, the present study revealed that a
beneficial IAPP/insulin secretion pattern was achieved in response
to exendin-4 pretreatment in MIN6 cells, and this was accompanied
by autophagy inhibition. hIAPP overexpression enhanced autophagy
and apoptosis in MIN6 cells, and these processes were attenuated by
exendin-4 pretreatment. Although more research is required to
examine the underlying protective mechanism of GLP-1 receptor
agonists on β-cells, the results highlight the potential
therapeutic application of GLP-1 receptor agonists in the treatment
of T2DM.
Acknowledgements
Not applicable.
Funding
This study was supported by the Clinical Medicine
Research Fund of the Chinese Medical Association (grant no.
13020110396) and Zhejiang Provincial Natural Science Foundation of
China (grant no. LY16H070006).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
XC, TH and YS performed the experiments. XG and FS
designed the study. WL and FS contributed essential reagents and/or
tools. TH, YS, LW and WL analyzed and interpreted the data. XG and
XC wrote the paper.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Butler AE, Janson J, Bonner-Weir S, Ritzel
R, Rizza RA and Butler PC: Beta-cell deficit and increased
beta-cell apoptosis in humans with type 2 diabetes. Diabetes.
52:102–110. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Haataja L, Gurlo T, Huang CJ and Butler
PC: Islet amyloid in type 2 diabetes and the toxic oligomer
hypothesis. Endocr Rev. 29:303–316. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gurlo T, Ryazantsev S, Huang CJ, Yeh MW,
Reber HA, Hines OJ, O'Brien TD, Glabe CG and Butler PC: Evidence
for proteotoxicity in beta cells in type 2 diabetes: Toxic islet
amyloid polypeptide oligomers form intracellularly in the secretory
pathway. Am J Pathol. 176:861–869. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fernández MS: Human IAPP amyloidogenic
properties and pancreatic β-cell death. Cell Calcium. 56:416–427.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Akter R, Cao P, Noor H, Ridgway Z, Tu LH,
Wang H, Wong AG, Zhang X, Abedini A, Schmidt AM and Raleigh DP:
Islet amyloid polypeptide: Structure, function, and
pathophysiology. J Diabetes Res. 2016:27982692016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lavine JA and Attie AD: Gastrointestinal
hormones and the regulation of β-cell mass. Ann N Y Acad Sci.
1212:41–58. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lotfy M, Singh J, Rashed H, Tariq S,
Zilahi E and Adeghate E: Mechanism of the beneficial and protective
effects of exenatide in diabetic rats. J Endocrinol. 220:291–304.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang C, Yuan L and Cao S: Endogenous
GLP-1 as a key self-defense molecule against lipotoxicity in
pancreatic islets. Int J Mol Med. 36:173–185. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lim SW, Jin L, Jin J and Yang CW: Effect
of exendin-4 on autophagy clearance in beta cell of rats with
tacrolimus-induced diabetes mellitus. Sci Rep. 6:299212016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lamont BJ and Andrikopoulos S: Hope and
fear for new classes of type 2 diabetes drugs: Is there preclinical
evidence that incretin-based therapies alter pancreatic morphology?
J Endocrinol. 221:T43–T61. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Göke R, McGregor GP and Göke B: Amylin
alters the biological action of the incretin hormone
GLP-1(7–36)amide. Life Sci. 53:1367–1372. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bak AM, Egefjord L, Gejl M, Steffensen C,
Stecher CW, Smidt K, Brock B and Rungby J: Targeting amyloid-beta
by glucagon-like peptide-1 (GLP-1) in Alzheimer's disease and
diabetes. Expert Opin Ther Targets. 15:1153–1162. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Qin Z, Sun Z, Huang J, Hu Y, Wu Z and Mei
B: Mutated recombinant human glucagon-like peptide-1 protects
SH-SY5Y cells from apoptosis induced by amyloid-beta peptide
(1–42). Neurosci Lett. 444:217–221. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Park YJ, Ao Z, Kieffer TJ, Chen H,
Safikhan N, Thompson DM, Meloche M, Warnock GL and Marzban L: The
glucagon-like peptide-1 receptor agonist exenatide restores
impaired pro-islet amyloid polypeptide processing in cultured human
islets: Implications in type 2 diabetes and islet transplantation.
Diabetologia. 56:508–519. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fan R, Li X, Gu X, Chan JC and Xu G:
Exendin-4 protects pancreatic beta cells from human islet amyloid
polypeptide-induced cell damage: Potential involvement of AKT and
mitochondria biogenesis. Diabetes Obes Metab. 12:815–824. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Aston-Mourney K, Hull RL, Zraika S,
Udayasankar J, Subramanian SL and Kahn SE: Exendin-4 increases
islet amyloid deposition but offsets the resultant beta cell
toxicity in human islet amyloid polypeptide transgenic mouse
islets. Diabetologia. 54:1756–1765. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Levine B and Klionsky DJ: Development by
self-digestion: Molecular mechanisms and biological functions of
autophagy. Dev Cell. 6:463–477. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nakai A, Yamaguchi O, Takeda T, Higuchi Y,
Hikoso S, Taniike M, Omiya S, Mizote I, Matsumura Y, Asahi M, et
al: The role of autophagy in cardiomyocytes in the basal state and
in response to hemodynamic stress. Nat Med. 13:619–624. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rivera JF, Costes S, Gurlo T, Glabe CG and
Butler PC: Autophagy defends pancreatic β cells from human islet
amyloid polypeptide-induced toxicity. J Clin Invest. 124:3489–3500.
2014. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Morita S, Sakagashira S, Shimajiri Y,
Eberhardt NL, Kondo T, Kondo T and Sanke T: Autophagy protects
against human islet amyloid polypeptide-associated apoptosis. J
Diabetes Investig. 2:48–55. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Asmar M and Holst JJ: Glucagon-like
peptide 1 and glucose-dependent insulinotropic polypeptide: New
advances. Curr Opin Endocrinol Diabetes Obes. 17:57–62. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bachar-Wikstrom E, Wikstrom JD, Ariav Y,
Tirosh B, Kaiser N, Cerasi E and Leibowitz G: Stimulation of
autophagy improves endoplasmic reticulum stress-induced diabetes.
Diabetes. 62:1227–1237. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mir SU, George NM, Zahoor L, Harms R,
Guinn Z and Sarvetnick NE: Inhibition of autophagic turnover in
β-cells by fatty acids and glucose leads to apoptotic cell death. J
Biol Chem. 290:6071–6085. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xiao J, Li X, Zhao X, He B, Shang X, Han
L, Wu G, Ding X and Zhu T: The effects and mechanism of islet
amyloid polypeptide on insulin secretion in INS-1 cells stimulated
by glibenclamide. Zhonghua Nei Ke Za Zhi. 54:214–218.
2015.PubMed/NCBI
|
|
26
|
Westermark P, Andersson A and Westermark
GT: Islet amyloid polypeptide, islet amyloid, and diabetes
mellitus. Physiol Rev. 91:795–826. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Krizhanovskii C, Fred RG, Oskarsson ME,
Westermark GT and Welsh N: Addition of exogenous sodium palmitate
increases the IAPP/insulin mRNA ratio via GPR40 in human EndoC-βH1
cells. Ups J Med Sci. 122:149–159. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shigeto M, Ramracheya R, Tarasov AI, Cha
CY, Chibalina MV, Hastoy B, Philippaert K, Reinbothe T, Rorsman N,
Salehi A, et al: GLP-1 stimulates insulin secretion by
PKC-dependent TRPM4 and TRPM5 activation. J Clin Invest.
125:4714–4728. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sheng Q, Xiao X, Prasadan K, Chen C, Ming
Y, Fusco J, Gangopadhyay NN, Ricks D and Gittes GK: Autophagy
protects pancreatic beta cell mass and function in the setting of a
high-fat and high-glucose diet. Sci Rep. 7:163482017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fujitani Y, Kawamori R and Watada H: The
role of autophagy in pancreatic beta-cell and diabetes. Autophagy.
5:280–282. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhao X, Liu G, Shen H, Gao B, Li X, Fu J,
Zhou J and Ji Q: Liraglutide inhibits autophagy and apoptosis
induced by high glucose through GLP-1R in renal tubular epithelial
cells. Int J Mol Med. 35:684–692. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang J, Wu J, Wu H, Liu X, Chen Y, Wu J,
Hu C and Zou D: Liraglutide protects pancreatic β-cells against
free fatty acids in vitro and affects glucolipid metabolism in
apolipoprotein E-/- mice by activating autophagy. Mol Med Rep.
12:4210–4218. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shigihara N, Fukunaka A, Hara A, Komiya K,
Honda A, Uchida T, Abe H, Toyofuku Y, Tamaki M, Ogihara T, et al:
Human IAPP-induced pancreatic β cell toxicity and its regulation by
autophagy. J Clin Invest. 124:3634–3644. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Guo H, Wang B, Li H, Ling L, Niu J and Gu
Y: Glucagon-like peptide-1 analog prevents obesity-related
glomerulopathy by inhibiting excessive autophagy in podocytes. Am J
Physiol Renal Physiol. 314:F181–F189. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cai X, Li J, Wang M, She M, Tang Y, Li J,
Li H and Hui H: GLP-1 treatment improves diabetic retinopathy by
alleviating autophagy through GLP-1R-ERK1/2-HDAC6 signaling
pathway. Int J Med Sci. 14:1203–1212. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ferdaoussi M, Abdelli S, Yang JY, Cornu M,
Niederhauser G, Favre D, Widmann C, Regazzi R, Thorens B, Waeber G
and Abderrahmani A: Exendin-4 protects beta-cells from
interleukin-1 beta-induced apoptosis by interfering with the c-Jun
NH2-terminal kinase pathway. Diabetes. 57:1205–1215. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kim MH, Kim EH, Jung HS, Yang D, Park EY
and Jun HS: EX4 stabilizes and activates Nrf2 via PKCδ,
contributing to the prevention of oxidative stress-induced
pancreatic beta cell damage. Toxicol Appl Pharmacol. 315:60–69.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zeng Z, Yu R, Zuo F, Zhang B, Ma H and
Chen S: Recombinant Lactococcus lactis expressing bioactive
exendin-4 to promote insulin secretion and beta-cell proliferation
in vitro. Appl Microbiol Biotechnol. 101:7177–7186. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Petersen M, Hofius D and Andersen SU:
Signaling unmasked: Autophagy and catalase promote programmed cell
death. Autophagy. 10:520–521. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhu J, Wang KZ and Chu CT: After the
banquet: Mitochondrial biogenesis, mitophagy, and cell survival.
Autophagy. 9:1663–1676. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Law BY, Wang M, Ma DL, Al-Mousa F,
Michelangeli F, Cheng SH, Ng MH, To KF, Mok AY, Ko RY, et al:
Alisol B, a novel inhibitor of the sarcoplasmic/endoplasmic
reticulum Ca(2+) ATPase pump, induces autophagy, endoplasmic
reticulum stress, and apoptosis. Mol Cancer Ther. 9:718–730. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Abedini A and Schmidt AM: Mechanisms of
islet amyloidosis toxicity in type 2 diabetes. FEBS Lett.
587:1119–1127. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ahrén B, Winzell MS, Wierup N, Sundler F,
Burkey B and Hughes TE: DPP-4 inhibition improves glucose tolerance
and increases insulin and GLP-1 responses to gastric glucose in
association with normalized islet topography in mice with
beta-cell-specific overexpression of human islet amyloid
polypeptide. Regul Pept. 143:97–103. 2007. View Article : Google Scholar : PubMed/NCBI
|