Introduction
It is well known that bone homeostasis is a dynamic
balance maintained by bone formation and bone resorption (1,2).
Osteoclasts are derived from hematopoietic precursors and function
in bone resorption. The classic receptor activator of nuclear
factor-κB (RANK)/RANK ligand (RANKL)/osteoprotegerin signaling
pathway regulates the activity and differentiation of osteoclasts.
RANKL is considered to be an important factor that regulates
osteoclast differentiation and bone metabolism (3). Excessive bone resorption is closely
associated with the formation of osteoclasts, which can lead to
bone disease, including osteoporosis (4). It has been shown that osteoporosis
significantly increases the risk of fracture, particularly hip
fractures, in postmenopausal women (5). Therefore, regulating osteoclast
differentiation and activity is important for preserving bone mass
and reducing the incidence of fractures. At present, the treatment
of osteoporosis predominantly includes estrogen replacement therapy
and medical treatment, including the use of calcitriol and caltrate
D; however, clinical evidence has demonstrated that these
treatments are associated with serious side effects, including
cancer (6,7). Therefore, identifying novel
treatments is critical.
As a non-invasive and inexpensive method,
low-frequency pulsed electromagnetic fields (PEMF) have shown
efficacy for a wide range of diseases of the skeletomuscular system
(8,9). In previous years, it has been
reported that PEMF may inhibit osteoclast differentiation in
vitro (10,11), however, the underlying mechanisms
remain to be fully elucidated. PEMF exposure is likely to produce
satisfying therapeutic effects for certain bone diseases, including
osteoporosis, which is closely associated with osteoclast function
(4,12,13).
Therefore, in vitro studies examining the effects of PEMF on
osteoclasts and the potential underlying mechanisms are essential
to understand the efficacy of PEMF for treating
osteoclast-associated diseases.
Reactive oxygen species (ROS) are produced
intracellularly as byproducts during mitochondrial electron
transport or through the action of certain enzymes, including NADPH
oxidase and cyclooxygenase (14).
ROS consist of radical and non-radical oxygen species, including
the superoxide anion (O2-), hydroxyl radical
(OH−) and hydrogen peroxide (H2O2)
(15). Changes in ROS levels are
implicated in regulating cellular signal transduction. The
excessive production of ROS results in oxidative stress, which in
turn may cause apoptosis, ischemia and inflammation (16). However, low, non-toxic levels of
ROS may act as secondary messengers in several receptor signaling
pathways (17–19). Various studies have shown that ROS
are implicated in bone metabolism and promote osteoclast
differentiation and bone resorption (20–23).
The increased production of ROS during osteoclast formation appears
to activate the peroxisome proliferator-activated-receptor-γ
coactivator 1β, which regulates mitochondrial biogenesis, thus
facilitating osteoclast differentiation (24). A previous study illustrated the
destructive effects of antioxidants on osteoclastogenesis in mouse
models lacking the forkhead box O transcription factor, which
drives transcription of the antioxidant catalase (25). To the best of our knowledge, the
inhibitory effects of PEMF on RANKL-induced osteoclast
differentiation have not been investigated. In the present study,
the effects of PEMF on osteoclast differentiation were examined. It
was demonstrated that PEMF served as a RANKL-mediated inhibitor of
osteoclastogenesis. The mechanism underlying this inhibitory effect
may be via the suppression of ROS generation, which is required for
osteoclast differentiation. Therefore, the results of the present
study demonstrated that PEMF may inhibit osteoclast differentiation
by scavenging intracellular ROS.
Materials and methods
Cell culture and PEMF exposure
RAW264.7 cells were purchased from the Cell Bank of
The Type Culture Collection of Chinese Academy of Sciences
(Shanghai, China). For the osteoclastogenesis experiment, the cells
were cultured in 24-well culture plate at 1×106
cells/ml, followed by culture in α-minimum essential media (α-MEM;
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) containing
10% fetal bovine serum (FBS, Gibco; The Fisher Scientific, Inc.)
and 1% penicillin/1% streptomycin in the presence of 50 ng/ml RANKL
(PeproTech, Inc., Rocky Hill, NJ, USA). The medium was placed at
37°C in a humidified atmosphere containing 5% CO2 for 4
days and refreshed every 48 h.
For PEMF, the cells were placed in an incubator
containing a 75 Hz sinusoidal PEMF with a density of 1 mT. The
magnetic field used for the test was produced via a Helmholtz coil.
The cells in the control group were placed in another incubator
under the same conditions, but without a PEMF in the incubator.
Cell Counting Kit-8 (CCK-8)
proliferation assay
Cell viability was quantified according to the
manufacturer's protocol using the CCK-8 kit (Dojindo Molecular
Technologies, Inc., Kumamoto, Japan). Briefly, the RAW264.7 cells
were seeded in 96-well plates at a cell density of 1,000 cells/well
and cell proliferation was determined at different time points
(each group had a set of three sub-wells). The PEMF group was
exposed to PEMF for 4 days (3 h/day), whereas control cells were
cultured under the same conditions without PEMF. After 24, 48, 72
and 96 h, 10 µl CCK-8 solution was added to each well. Following
incubation at 37°C for 4 h, the optical density (OD) of each well
was determined using an enzyme mark instrument (Multiskan FC;
Thermo Fisher Scientific, Inc.) at a wavelength of 450 nm.
Tartrate-resistant acid phosphatase
(TRAP) staining, cell counting and TRAP activity assay
Following exposure to PEMF, the cells were washed
with PBS and fixed. A TRAP staining kit (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) was used to identify the formation of mature
osteoclasts according to the manufacturer's protocol. The cell
culture plate was transferred to an optical microscope, and cells
that were TRAP-positive with more than three nuclei were confirmed
to be osteoclasts. Five random fields of view were selected, and
the number of osteoclasts was counted. TRAP activity was measured
at a wavelength of 540 nm with a microplate reader.
Determination of intracellular
ROS
The level of intracellular ROS generated by
H2O2 was measured using a ROS assay kit
(Beyotime Institute of Biotechnology, Haimen, China).
H2O2 was used as a ROS inducer. DCFH-DA
interacts with ROS in viable cells to generate
2′,7′-dichlorofluorescein (DCF). DCF is highly fluorescent at 530
nm. The cells were washed with PBS three times and cultured for 30
min at 37°C in the dark following the addition of DCFH-DA at a
final concentration of 10 µM/ml. The relative expression of ROS was
evaluated using a fluorescent microscope, and the fluorescence
intensity of DCF was determined using a fluorometric plate reader.
To further investigate the involvement of ROS in the promotion of
osteoclast precursor differentiation, diphenylene-iodonium chloride
(DPI; Sigma-Aldrich; Merck KGaA), a widely used NADH oxidase 1
inhibitor that completely scavenges generated ROS, was added to the
cultured system (RANKL).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
TRIzol® reagent (Thermo Fisher
Scientific, Inc.) was used to extract intracellular total RNA, and
a Prime Script RT kit (Takara Bio, Inc., Otsu, Japan) was used to
synthesize single-stranded cDNA. qPCR was performed using an ABI
7500 real-time PCR system (Thermo Fisher Scientific, Inc.). qPCR
was conducted using cDNA (1 µl) with SYBR Green-1 (20 µl; Takara
Bio, Inc.) according to the manufacturer's protocol under the
following conditions: Activation at 95°C for 10 min, then 40 cycles
of amplification (95°C for 10 sec, 60°C for 24 sec and 72°C for 20
sec) and a final extension at 72°C for 1 min. All the reactions
were performed in triplicate and target gene expression was
normalized to the reference gene β-actin. The 2−Δ∆Cq
method (26) was applied to
calculate the relative expression level of the target genes. The
PCR products were subjected to melting curve analysis and a
standard curve to confirm the correct amplification. The primers
used for PCR were as follows: Cathepsin K (CTSK), forward
5′-AGAACGGAGGCATTGACTCT-3′, reverse 5′-GATGGACACAGAGATGGGTC-3′;
TRAP, forward 5′-CGATCACAATCTGCAGTACC-3′, reverse
5′-ACCCAGTGAGTCTTCAGTCC-3′; nuclear factor of activated T cells
cytoplasmic 1 (NFATc1), forward 5′-CGCAAGTACAGTCTCAATGG-3′, reverse
5′-CAGGTATCTTCGGTCACACT-3′; and β-actin, forward
5′-AGGCCAACCGTGAAAAGATG-3′ and reverse
5′-TGGCGTGAGCGAGACCATAG-3′.
Western blotting
The treated RAW264.7 cells (5×104 cells
per well) were lysed using cOmplete™ Lysis-M and Phos-STOP (Roche
Diagnostics, Indianapolis, IN, USA). The concentration was measured
using a Bicinchoninic Acid protein assay kit (Pierce; Thermo Fisher
Scientific, Inc.), Proteins (40 µg/lane) were separated via 10%
SDS-PAGE. The proteins were then transferred to polyvinylidene
difluoride (PVDF) membranes, and were blocked for 1.5 h at room
temperature with 3% non-fat dry milk in TBS-Tween-20 (150 mM NaCl,
pH 7.4, 25 mM Tris-HCl and 0.2% Tween-20). The membranes were
incubated with specific primary antibodies overnight at 4°C. The
primary rabbit polyclonal antibodies used were as follows:
Anti-β-actin (1:1,000; cat. no. ab8226, Abcam, Cambridge, MA, USA);
anti-CTSK (1:2,000; cat. no. 11239-1-AP, ProteinTech Group, Inc.,
Chicago, IL, USA); anti-NFATc1 (1:3,000; cat. no. 4389, Cell
Signaling Technology, Inc., Danvers, MA, USA) and anti-TRAP
(1:2,000; cat. no. 10325-1-AP, ProteinTech Group, Inc.). Membranes
were then incubated for 1 h at room temperature with horseradish
peroxidase-conjugated goat anti-rabbit immunoglobulin G secondary
antibodies (1:1,000; cat. no. 7074, Cell Signaling Technology,
Inc.). Immunoreactive bands on the PVDF membranes were detected by
enhanced chemiluminescence (Tanon Science and Technology Co., Ltd.,
Shanghai, China).
Statistical analysis
Data are presented as the mean ± standard deviation.
Each experiment was repeated at least three times. Statistical
analyses were performed using one-way analysis of variance with
subsequent post hoc multiple comparisons with Dunnett's test using
SPSS version 20 (IBM Corp., Armonk, NY, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
PEMF inhibits the osteoclastic
differentiation, but not the viability of RANKL-induced RAW264.7
cells
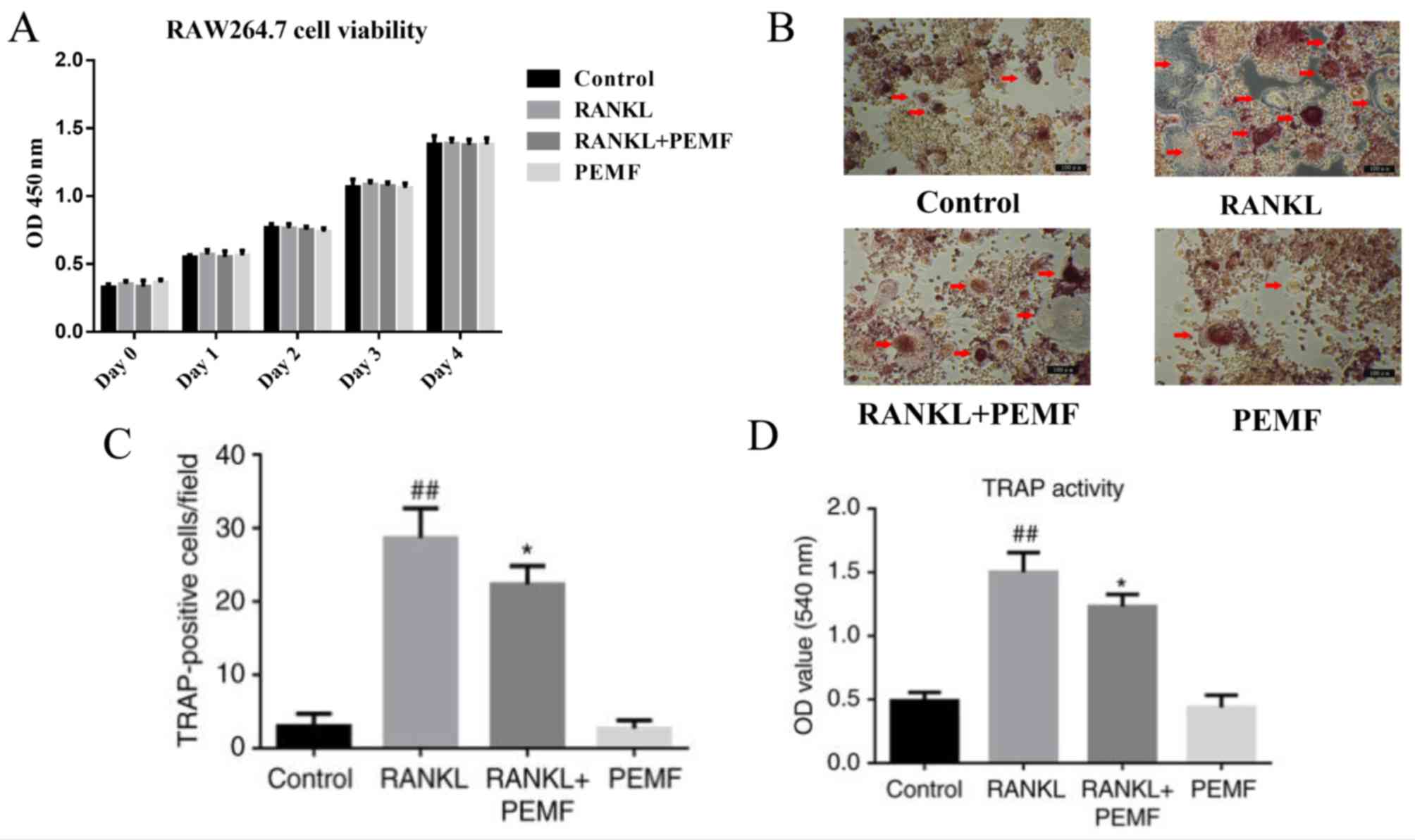
Cell viability was assessed using a CCK-8 assay
following exposure of the RAW264.7 cells to PEMF. The RAW264.7
cells were treated with or without RANKL (50 ng/l) for 4 days under
PEMF (75 Hz, 1 mt, 3 h/day). The results revealed no significant
difference in cell viability between groups (Fig. 1A).
In the present study, the inhibitory effect of PEMF
on osteoclast differentiation was investigated. RANKL (50 ng/ml)
induced the differentiation of RAW264.7 murine macrophage cells
into osteoclasts. Osteoclasts were visualized under an optical
microscope (Fig. 1B). However,
PEMF significantly reduced the number of TRAP-positive
multinucleated osteoclasts (Fig.
1C). Compared with the control group, RANKL treatment increased
the activity of TRAP by 150%, whereas PEMF reduced the activity of
TRAP stimulated by RANKL (Fig.
1D). No significant difference was observed between the control
group and PEMF group.
PEMF inhibits the expression of
osteoclastogenesis-associated genes
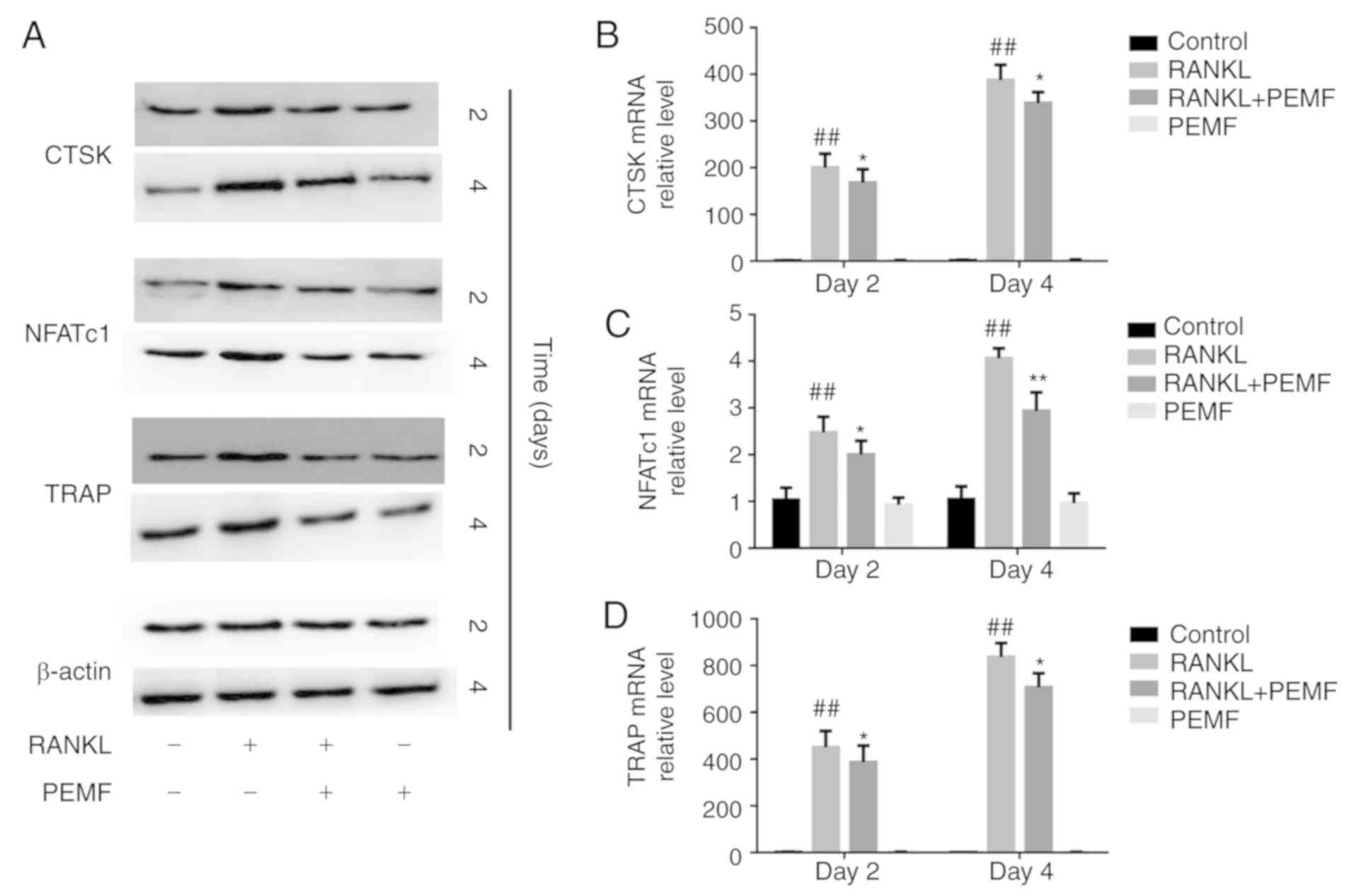
The expression of several
osteoclastogenesis-associated genes, including CTSK, NFATc1 and
TRAP, were determined by western blotting (Fig. 2A) and RT-qPCR analysis (Fig. 2B-D) on days 2 and 4. The results
showed that PEMF inhibited the RANKL-induced increase in the
expression of CTSK, NFATc1 and TRAP, which was consistent with the
above results.
Antioxidant capacity and intracellular
ROS levels are reduced by PEMF
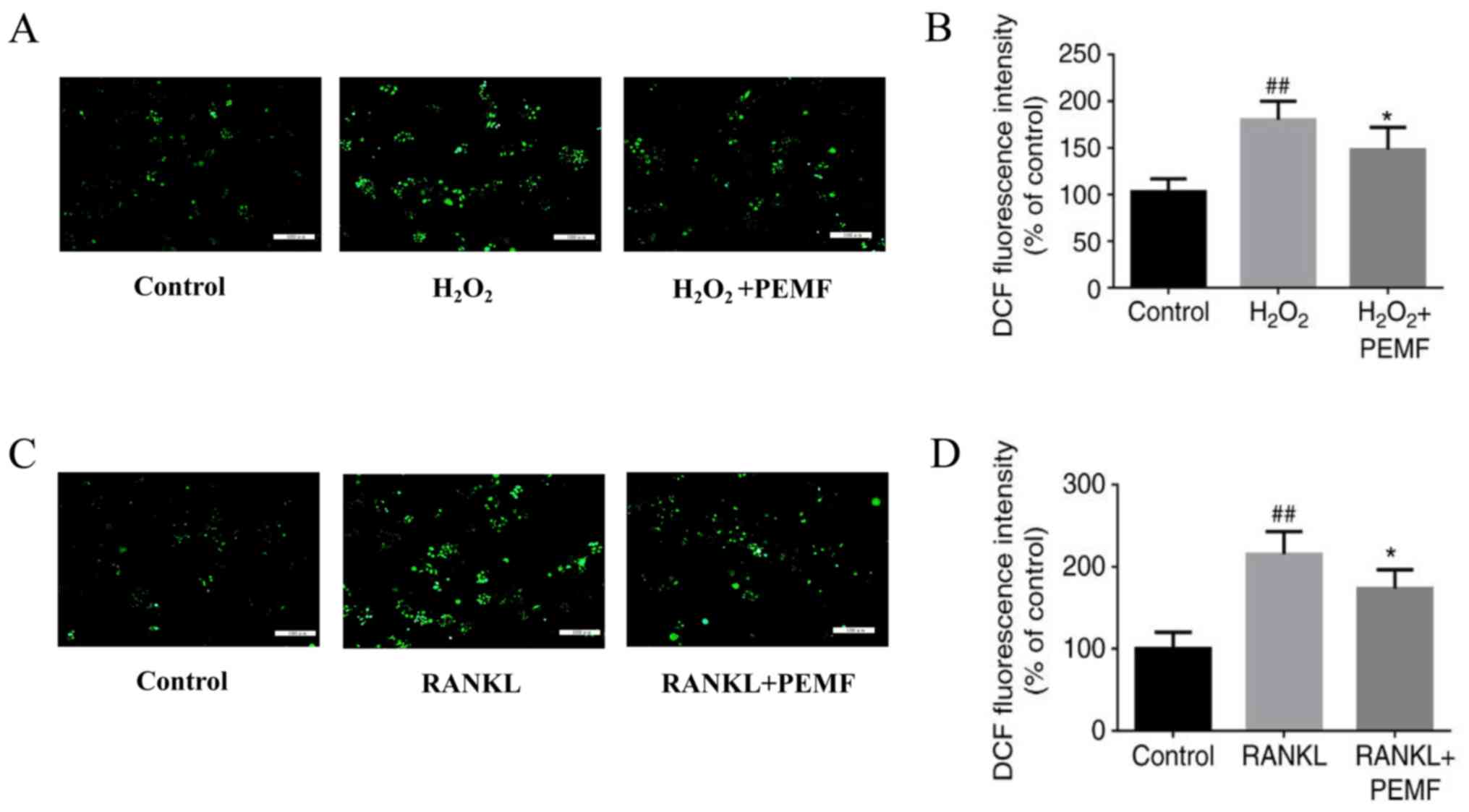
To confirm whether PEMF can scavenge intracellular
ROS, the accumulation of intracellular ROS was evaluated by
fluorescence microscopy. The RAW264.7 cells were exposed to PEMF
for 3 h following treatment with 1 mM H2O2
for 30 min. A previously reported study demonstrated that during
RANKL-induced differentiation of RAW264.7 cells into osteoclasts,
maximum intracellular ROS levels were reached at day 2 (27). Therefore, the inhibitory effect of
PEMF on ROS production was assessed following 2 days of induction
with RANKL (50 ng/ml). The ROS-positive cells were visualized as
bright green spots, and H2O2 treatment
increased the level of intracellular ROS, compared with that in the
control group. In addition, treatment with PEMF inhibited the
H2O2-induced increase in intracellular ROS
(Fig. 3A). All cells were
collected and analyzed on a fluorometric plate reader to measure
the OD values of DCF. The fluorescence intensity of DCF was reduced
in the PEMF + H2O2 group compared with that
in the H2O2-treated group (Fig. 3B). In the RAW264.7 cells treated
with RANKL and exposed to PEMF for 2 days (3 h/day), it was
determined that RANKL treatment significantly increased ROS, and
PEMF prevented this increase (Fig.
3C). Similar results were observed for the OD values of DCF
(Fig. 3D). These results suggested
that PEMF had antioxidant capacity and reduced intracellular
ROS.
Reducing intracellular ROS levels
inhibits RANKL-induced osteoclast differentiation
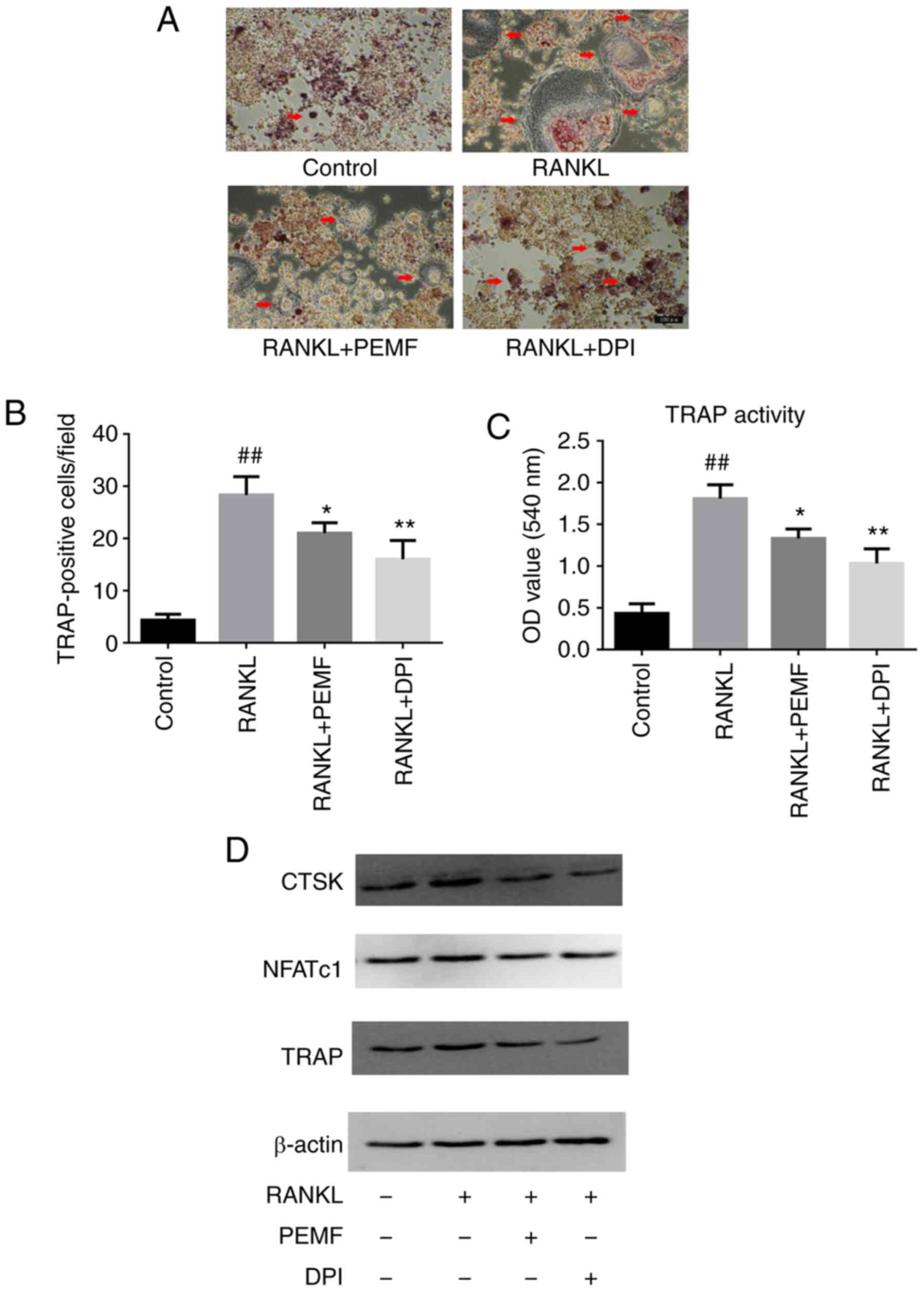
The results showed that DPI administration inhibited
the effect of RANKL on osteoclast differentiation and reduced the
number of TRAP-positive multinuclear cells (Fig. 4A and B). The decreased activity of
TRAP (Fig. 4C) was consistent with
the observed alterations in the TRAP staining. DPI treatment also
downregulated the expression of osteoclastogenesis-associated genes
(Fig. 4D). These data revealed ROS
to be the key factor in RANKL-induced osteoclast formation, and
scavenging intracellular ROS effectively inhibited osteoclast
formation.
Discussion
Osteoclasts are unique in their ability to resorb
mineralized bone and have critical functions in bone remodeling and
physical skeletal morphogenesis (28). In recent decades, it has been
confirmed that electromagnetic fields, including PEMF, have
potential in curing skeletal muscle system diseases and/or injury,
including fracture healing, particularly in tendon-to-bone healing
and joint and bone injury (29–31).
Abnormalities in osteoclast number and activity are associated with
osteoporosis. In the present study, it was evident that 75 Hz, 1 mt
PEMF exhibited a distinctly inhibitory effect on RANKL-induced
osteoclast differentiation. Therefore, it is of clinical
significance.
Previous studies have examined the action of PEMF in
osteoporosis, attributing its mechanism of action to osteoblast
precursor proliferation (32) and
the inhibition of osteoclast activity (33), in addition to the enhancement of
osteoblastic mineralization potential (9). Treatment of bone marrow cells derived
from ovariectomized rats with PEMF led to significant suppression
of osteoclast formation and osteoclast-associated cytokine
expression (34). Furthermore,
certain properties of deteriorated bone, including its stiffness,
the maximum load and the yield load, are inhibited following PEMF
exposure, indicating that more bone mass was retained and there was
greater mechanical bone strength to resist fractures (35). Wang et al (36) also reported that RAW264.7 cells
incubated with RANKL and exposed to 15 Hz PEMF (2 h/day) at 3 mt
for 7 days had decreased bone resorbing capacity, due to the
promotion of osteoclast apoptosis. These results suggested that
PEMF exerts biological effects on osteoclasts and/or osteoblasts.
To better understand how PEMF affects osteoclasts, the present
study also examined the effects of PEMF on osteoclast
differentiation. Although it was demonstrated that PEMF inhibited
RANKL-induced osteoclast differentiation by scavenging
intracellular ROS, other possible mechanisms cannot be excluded.
Another important regulating factor in osteoclasts is calcium.
During RANKL-induced osteoclastogenesis, Ca2+
upregulates the downstream signal NFATc1, which is considered the
master regulator of osteoclastogenesis (37). Previous studies have reported that
PEMF affects the influx of Ca2+ in several cell types
(38,39). Therefore, Ca2+ may be
another indication that PEMF affects osteoclast
differentiation.
In the classic RANKL signaling pathway, the
production of ROS is mediated by NADPH oxidase I in response to
RANKL (40). In the present study,
it was found that RANKL significantly increased intracellular ROS,
consistent with previous reports. ROS are byproducts of cell
metabolism and are also important signaling molecules. The ability
of ROS to serve as signaling molecules in pathways has been well
established, particularly in osteoclast differentiation, where ROS
are involved in the nuclear factor (NF)-κB and mitogen-activated
protein kinase (MAPK) signaling pathways (19). Additionally, the ROS-mediated
direct modification of inositol 1,4,5-trisphosphate receptor thiol
groups promotes Ca2+ release from the endoplasmic
reticulum (41). As mentioned
previously, Ca2+ is involved in osteoclast
differentiation. Through NF-κB, MAPK and Ca2+ signaling,
ROS are involved in the differentiation and formation of
osteoclasts. DPI scavenges RANKL-induced intracellular ROS,
effectively preventing the differentiation of osteoclast
precursors. Furthermore, RANKL may upregulate the expression of
superoxide dismutase 2 (SOD2) and sirtuin 3 (Sirt3), which exert
feedback on osteoclastogenesis by inhibiting RANKL signaling via
the downregulation of ROS (42).
This mechanism physiologically prevents osteoclast overgrowth.
Sirt3-deficient mice exhibit severe bone loss caused by elevated
osteoclastogenesis (43). By
contrast, evidence has revealed that several diseases are
associated with ROS, including osteoporosis (15,44).
Certain antioxidants, including alliin and lycopene, inhibit
osteoclast formation and bone resorption via the inhibition of ROS
generation (45,46).
Exposure to PEMF leads to the upregulation of
proteins associated with proper protein folding (47–50).
A previous study revealed that PEMF effectively prevents the
pro-oxidant effects of H2O2 in SK-N-BE(2)
human neuroblastoma cells by increasing Mn-dependent superoxide
dismutase (MnSOD)-based antioxidant protection (51). Furthermore, Falone et al
(52) found that PEMF markedly
decreased ROS in human osteoclasts by inducing the expression of
antioxidant enzymes, including glutathione peroxidase 3, SOD2,
catalase and glutathione-s-reductase. However, whether the same
mechanism exists in osteoclasts remains to be elucidated and
requires further investigation.
There are certain limitations to the present study.
First, the exact mechanism of PEMF scavenging of intracellular ROS
in osteoclasts remains to be fully elucidated. As research in this
area advances, there is no doubt that this issue can be addressed.
Second, although RAW264.7 cells are widely used to induce the
formation of osteoclasts in response to RANKL, they are not
identical to osteoclasts derived from the body. For example, when
comparing in vivo and in vitro conditions, there are
discrepancies in vitamin D compound synthesis during osteoclastic
bone resorption (53). Third, the
present study did not determine the optimal PEMF parameters,
including exposure time, strength and frequency, and did not
perform bone resorption analysis or NOX expression assays.
Therefore, further investigation is required.
In conclusion, the findings of the present study
demonstrated that PEMF inhibited RANKL-induced osteoclast
differentiation by scavenging intracellular ROS. These findings
provide theoretical support for using PEMF in the treatment of
osteoporosis and also indicates that ROS may be a potential
therapeutic target in the treatment of osteoporosis.
Acknowledgements
The authors would like to thank Dr Ou Yang
(Department of Human Anatomy, Basic Medical College, Southern
Medical University) for the technical assistance of providing
laboratory access.
Funding
This study was supported by the Science and
Technology Planning Project of Guangdong Province, China (grant no.
2013B021800312).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JT conceived and designed the study. YP, HL, QY, YY,
HX, YL and ZH performed the experiments. YP drafted the manuscript.
HL, QY, YY, HX, YL and ZH reviewed and edited the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lemaire V, Tobin FL, Greller LD, Cho CR
and Suva LJ: Modeling the interactions between osteoblast and
osteoclast activities in bone remodeling. J Theor Biol.
229:293–309. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bloemen V, Schoenmaker T, de Vries TJ and
Everts V: Direct cell-cell contact between periodontal ligament
fibroblasts and osteoclast precursors synergistically increases the
expression of genes related to osteoclastogenesis. J Cell Physiol.
222:565–573. 2010.PubMed/NCBI
|
|
3
|
Boyle WJ, Simonet WS and Lacey DL:
Osteoclast differentiation and activation. Nature. 423:337–342.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Reid IR: Short-term and long-term effects
of osteoporosis therapies. Nat Rev Endocrinol. 11:418–428. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Harvey N, Dennison E and Cooper C:
Osteoporosis: Impact on health and economics. Nat Rev Rheumatol.
6:99–105. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Adami HO, Persson I, Hoover R, Schairer C
and Bergkvist L: Risk of cancer in women receiving hormone
replacement therapy. Int J Cancer. 44:833–839. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lewiecki EM: Treatment of osteoporosis
with denosumab. Maturitas. 66:182–186. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jing D, Li F, Jiang M, Cai J, Wu Y, Xie K,
Wu X, Tang C, Liu J, Guo W, et al: Pulsed electromagnetic fields
improve bone microstructure and strength in ovariectomized rats
through a Wnt/Lrp5/β-catenin signaling-associated mechanism. PLoS
One. 8:e793772013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hannemann PF, Mommers EH, Schots JP, Brink
PR and Poeze M: The effects of low-intensity pulsed ultrasound and
pulsed electromagnetic fields bone growth stimulation in acute
fractures: A systematic review and meta-analysis of randomized
controlled trials. Arch Orthop Trauma Surg. 134:1093–1106. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chang K, Chang WH, Tsai MT and Shih C:
Pulsed electromagnetic fields accelerate apoptotic rate in
osteoclasts. Connect Tissue Res. 47:222–228. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
He Z, Selvamurugan N, Warshaw J and
Partridge NC: Pulsed electromagnetic fields inhibit human
osteoclast formation and gene expression via osteoblasts. Bone.
106:194–203. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou J, Liao Y, Zeng Y, Xie H, Fu C and Li
N: Effect of intervention initiation timing of pulsed
electromagnetic field on ovariectomy-induced osteoporosis in rats.
Bioelectromagnetics. 38:456–465. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Assiotis A, Sachinis NP and Chalidis BE:
Pulsed electromagnetic fields for the treatment of tibial delayed
unions and nonunions. A prospective clinical study and review of
the literature. J Orthop Surg Res. 7:242012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim H, Kim IY, Lee SY and Jeong D: Bimodal
actions of reactive oxygen species in the differentiation and
bone-resorbing functions of osteoclasts. FEBS Lett. 580:5661–5665.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Domazetovic V, Marcucci G, Iantomasi T,
Brandi ML and Vincenzini MT: Oxidative stress in bone remodeling:
Role of antioxidants. Clin Cases Miner Bone Metab. 14:209–216.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Davies KJ: Oxidative stress: The paradox
of aerobic life. Biochem Soc Symp. 61:1–31. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Forman HJ, Fukuto JM and Torres M: Redox
signaling: Thiol chemistry defines which reactive oxygen and
nitrogen species can act as second messengers. Am J Physiol Cell
Physiol. 287:C246–C256. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lander HM: An essential role for free
radicals and derived species in signal transduction. FASEB J.
11:118–124. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Callaway DA and Jiang JX: Reactive oxygen
species and oxidative stress in osteoclastogenesis, skeletal aging
and bone diseases. J Bone Miner Metab. 33:359–370. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lean JM, Davies JT, Fuller K, Jagger CJ,
Kirstein B, Partington GA, Urry ZL and Chambers TJ: A crucial role
for thiol antioxidants in estrogen-deficiency bone loss. J Clin
Invest. 112:915–923. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Moon HJ, Ko WK, Han SW, Kim DS, Hwang YS,
Park HK and Kwon IK: Antioxidants, like coenzyme Q10, selenite, and
curcumin, inhibited osteoclast differentiation by suppressing
reactive oxygen species generation. Biochem Biophys Res Commun.
418:247–253. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Moon HJ, Kim SE, Yun YP, Hwang YS, Bang
JB, Park JH and Kwon IK: Simvastatin inhibits osteoclast
differentiation by scavenging reactive oxygen species. Exp Mol Med.
43:605–612. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Garrett IR, Boyce BF, Oreffo RO, Bonewald
L, Poser J and Mundy GR: Oxygen-derived free radicals stimulate
osteoclastic bone resorption in rodent bone in vitro and in vivo. J
Clin Invest. 85:632–639. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ishii KA, Fumoto T, Iwai K, Takeshita S,
Ito M, Shimohata N, Aburatani H, Taketani S, Lelliott CJ,
Vidal-Puig A and Ikeda K: Coordination of PGC-1beta and iron uptake
in mitochondrial biogenesis and osteoclast activation. Nat Med.
15:259–266. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bartell SM, Kim HN, Ambrogini E, Han L,
Iyer S, Serra Ucer S, Rabinovitch P, Jilka RL, Weinstein RS, Zhao
H, et al: FoxO proteins restrain osteoclastogenesis and bone
resorption by attenuating H2O2 accumulation.
Nat Commun. 5:37732014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee SH, Ding Y, Yan XT, Kim YH and Jang
HD: Scopoletin and scopolin isolated from Artemisia iwayomogi
suppress differentiation of osteoclastic macrophage RAW 264.7 cells
by scavenging reactive oxygen species. J Nat Prod. 76:615–620.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Teitelbaum SL and Ross FP: Genetic
regulation of osteoclast development and function. Nat Rev Genet.
4:638–649. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tucker JJ, Cirone JM, Morris TR, Nuss CA,
Huegel J, Waldorff EI, Zhang N, Ryaby JT and Soslowsky LJ: Pulsed
electromagnetic field therapy improves tendon-to-bone healing in a
rat rotator cuff repair model. J Orthop Res. 35:902–909. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Osti L, Buono AD and Maffulli N: Pulsed
electromagnetic fields after rotator cuff repair: A randomized,
controlled study. Orthopedics. 38:e223–e228. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Veronesi F, Torricelli P, Giavaresi G,
Sartori M, Cavani F, Setti S, Cadossi M, Ongaro A and Fini M: In
vivo effect of two different pulsed electromagnetic field
frequencies on osteoarthritis. J Orthop Res. 32:677–685. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hartig M, Joos U and Wiesmann HP:
Capacitively coupled electric fields accelerate proliferation of
osteoblast-like primary cells and increase bone extracellular
matrix formation in vitro. Eur Biophys J. 29:499–506. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lohmann CH, Schwartz Z, Liu Y, Guerkov H,
Dean DD, Simon B and Boyan BD: Pulsed electromagnetic field
stimulation of MG63 osteoblast-like cells affects differentiation
and local factor production. J Orthop Res. 18:637–646. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
He J, Zhang Y, Chen J, Zheng S, Huang H
and Dong X: Effects of pulsed electromagnetic fields on the
expression of NFATc1 and CAII in mouse osteoclast-like cells. Aging
Clin Exp Res. 27:13–19. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jing D, Cai J, Wu Y, Shen G, Li F, Xu Q,
Xie K, Tang C, Liu J, Guo W, et al: Pulsed electromagnetic fields
partially preserve bone mass, microarchitecture, and strength by
promoting bone formation in hindlimb-suspended rats. J Bone Miner
Res. 29:2250–2261. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang P, Liu J, Yang Y, Zhai M, Shao X, Yan
Z, Zhang X, Wu Y, Cao L, Sui B, et al: Differential
intensity-dependent effects of pulsed electromagnetic fields on
RANKL-induced osteoclast formation, apoptosis, and bone resorbing
ability in RAW264.7 cells. Bioelectromagnetics. 38:602–612. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Negishi-Koga T and Takayanagi H:
Ca2+-NFATc1 signaling is an essential axis of osteoclast
differentiation. Immunol Rev. 231:241–256. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pall ML: Electromagnetic fields act via
activation of voltage-gated calcium channels to produce beneficial
or adverse effects. J Cell Mol Med. 17:958–965. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cornacchione M, Pellegrini M, Fassina L,
Mognaschi ME, Di Siena S, Gimmelli R, Ambrosino P, Soldovieri MV,
Taglialatela M, Gianfrilli D, et al: β-Adrenergic response is
counteracted by extremely-low-frequency pulsed electromagnetic
fields in beating cardiomyocytes. J Mol Cell Cardiol. 98:146–158.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lee NK, Choi YG, Baik JY, Han SY, Jeong
DW, Bae YS, Kim N and Lee SY: A crucial role for reactive oxygen
species in RANKL-induced osteoclast differentiation. Blood.
106:852–859. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Decuypere JP, Monaco G, Missiaen L, De
Smedt H, Parys JB and Bultynck G: IP(3) receptors, mitochondria,
and Ca signaling: Implications for aging. J Aging Res.
2011:9201782011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kim H, Lee YD, Kim HJ, Lee ZH and Kim HH:
SOD2 and Sirt3 control osteoclastogenesis by regulating
mitochondrial ROS. J Bone Miner Res. 32:397–406. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Huh JE, Shin JH, Jang ES, Park SJ, Park
DR, Ko R, Seo DH, Kim HS, Lee SH, Choi Y, et al: Sirtuin 3 (SIRT3)
maintains bone homeostasis by regulating AMPK-PGC-1β axis in mice.
Sci Rep. 6:225112016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sendur OF, Turan Y, Tastaban E and Serter
M: Antioxidant status in patients with osteoporosis: A controlled
study. Joint Bone Spine. 76:514–518. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chen Y, Sun J, Dou C, Li N, Kang F, Wang
Y, Cao Z, Yang X and Dong S: Alliin attenuated RANKL-induced
osteoclastogenesis by scavenging reactive oxygen species through
inhibiting Nox1. Int J Mol Sci. 17(pii): E15162016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Rao LG, Krishnadev N, Banasikowska K and
Rao AV: Lycopene I-effect on osteoclasts: Lycopene inhibits basal
and parathyroid hormone-stimulated osteoclast formation and mineral
resorption mediated by reactive oxygen species in rat bone marrow
cultures. J Med Food. 6:69–78. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Falone S, Grossi MR, Cinque B, D'Angelo B,
Tettamanti E, Cimini A, Di Ilio C and Amicarelli F: Fifty hertz
extremely low-frequency electromagnetic field causes changes in
redox and differentiative status in neuroblastoma cells. Int J
Biochem Cell Biol. 39:2093–2106. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Simkó M: Cell type specific redox status
is responsible for diverse electromagnetic field effects. Curr Med
Chem. 14:1141–1152. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Mannerling AC, Simko M, Mild KH and
Mattsson MO: Effects of 50-Hz magnetic field exposure on superoxide
radical anion formation and HSP70 induction in human K562 cells.
Radiat Environ Biophys. 49:731–741. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Osera C, Fassina L, Amadio M, Venturini L,
Buoso E, Magenes G, Govoni S, Ricevuti G and Pascale A:
Cytoprotective response induced by electromagnetic stimulation on
SH-SY5Y human neuroblastoma cell line. Tissue Eng Part A.
17:2573–2582. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ehnert S, Fentz AK, Schreiner A, Birk J,
Wilbrand B, Ziegler P, Reumann MK, Wang H, Falldorf K and Nussler
AK: Extremely low frequency pulsed electromagnetic fields cause
antioxidative defense mechanisms in human osteoblasts via induction
of •O2− and H2O2. Sci
Rep. 7:145442017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Falone S, Marchesi N, Osera C, Fassina L,
Comincini S, Amadio M and Pascale A: Pulsed electromagnetic field
(PEMF) prevents pro-oxidant effects of H2O2
in SK-N-BE(2) human neuroblastoma cells. Int J Radiat Biol.
92:281–286. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Suda T, Takahashi F and Takahashi N: Bone
effects of vitamin D-discrepancies between in vivo and in vitro
studies. Arch Biochem Biophys. 523:22–29. 2012. View Article : Google Scholar : PubMed/NCBI
|