Introduction
Glomerulosclerosis (GS) is the final pathway leading
to the loss of renal function caused by a phenotypic transition of
mesangial cells and an increase in extracellular matrix formation
(1). The lesions are characterized
by a loss of podocytes (2,3) and an accumulation of extracellular
matrix (4). The effect of these
etiological factors manifest as changes to glomerular mechanics
(including high glomerular filtration and pressure), metabolism
(such as diabetes) and a variety of regulatory molecules (such as
cytokines) (5–7). Transforming growth factor-β (TGF-β)
is a multifunctional cytokine that regulates cell proliferation,
differentiation and apoptosis. It has been previously demonstrated
that TGF-β can promote renal fibrosis by stimulating the synthesis
and inhibiting the degradation of the extracellular matrix
(8). Furthermore, TGF-β can induce
podocyte apoptosis by activating the mothers against
decapentaplegic homolog 9 (Smad) signaling pathway (9). Together, these results suggest that
TGF-β is an important pathogenic factor involved in the
pathogenesis of GS. Thus, the treatment of GS with TGF-β is
currently being investigated.
Quercetin (QU) is a natural flavonoid that is
present in various flowers, and leaves and fruits of plants
(10). Recent pharmacological
studies have demonstrated that QU exerts strong anti-tumor,
anti-oxidant, anti-fibrosis and anti-viral effects by regulating
the activity of multiple signaling pathways (11,12).
Another study suggested that QU inhibits the proliferation and
invasion of prostate tumors by preventing TGF-β-induced epithelial
mesenchymal transition (EMT) (13). EMT is a complex biological process
in which epithelial cells are transformed into interstitial
phenotypes with the loss of specific endothelial cell markers,
including mesenchymal phenotype initiating expression of
fibroblast-specific protein-1 (FSP-1) and α-smooth muscle actin
(α-SMA). In addition, QU reduces the expression of TGF-β to inhibit
renal interstitial fibrosis (14).
Based on these results, QU may alleviate GS by inhibiting the TGF-β
signaling pathway.
In the present study, the mechanism and feasibility
of QU treatment for GS were examined by evaluating the effects of
QU on physiological parameters of GS mice and associated proteins
in the TGF-β signaling pathway.
Materials and methods
Animal model
A total of 60 male, 6–8 week old Sprague-Dawley
rats, weighing 220±20 g, were purchased from the Experimental
Animal Center of Gansu University of Chinese Medicine (Lanzhou,
China). The protocol was approved prior to commencing this study by
the animal ethics committee of Gansu University of Chinese
Medicine. All rats were housed under standard conditions (12/12
light/dark cycle, 40% humidity and 21–25°C) and fed a standard rat
diet. The rats were randomized into three groups: Control group
(NC; n=10), sham operation group (SHO; n=10) and GS model group
(GS; n=40). The GS group was prepared by uninephrectomy of the left
kidney. Briefly, rats were anesthetized by intraperitoneal
injection of 30–60 mg/kg pentobarbital. Following routine
disinfection, an incision of 1–1.5 cm was made in the back to
expose the kidney. The kidney fat and adrenal glands were stripped
away, the left renal portal vessels were ligated, the left kidney
was resected and the incision was sutured. Subsequently, these rats
were treated with adriamycin (ADR; Shanxi Pude Pharmaceutical Co.,
Shanxi, China) at a dose of 3 mg/kg by single tail vein injection
at days 8 and 29. The SHO group was operated on without removal of
the kidney and received tail vein injection with saline solution,
while the NC group was maintained under normal conditions without
surgery or injection.
Drug treatment and biochemical
analysis
A total of 30 randomly selected GS rats were divided
equally into three groups (n=10) and treated with different doses
of QU (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) once daily by
oral gavage for 8 weeks: Low-dose group (QU I, 25 mg/kg/day),
intermediate-dose group (QU II, 50 mg/kg/day) and high-dose group
(QU III, 100 mg/kg/day). The NC group, SHO group, GS group, QU I
group, QU II group and QU III group were treated with equal volumes
of saline solutions. Following 8 weeks of drug intervention, the
mental status (15), activity, fur
glossiness, appetite, defecation and weight alterations were
observed and recorded for each group. The 24-h urine total protein
excretion (UTP/24 h) was measured by the sulfosalicylic acid
method. Briefly, 0.1 ml 4% sulfosalicylic acid reagent was added to
2–3 ml of urine supernatant in a test tube. The tube was gently
shaken for 1 min and immediately observed for turbidity. The blood
was collected from the caudal vein of the rats and the levels of
serum total protein (TP), serum albumin (Alb), triglyceride (TG),
serum total cholesterol (TC), urea nitrogen (BUN), and serum
creatinine (SCr) were measured with ELISA kits according to the
manufacturer's protocol: Rat triglyceride (TG; RA20187); Rat
Albumin (Alb; RA20636); Rat Total cholesterol (TC; RA20136); Rat
creatinine (Cr; RA20115) from Bio-Swamp (Wuhan, Hubei, China). Rat
total protein (TP; RJ16656) and Rat urea nitrogen (BUN; RJ16084)
were purchased from Shanghai Renjie Biotechnology Co., Ltd.
(Shanghai, China).
Pathological analysis
After rats in each group were anesthetized
(pentobarbital, 45 mg/kg, intraperitoneal injection) and rapidly
sacrificed by cervical dislocation. These rats were sacrificed and
the right kidneys were removed and fixed with 4% formaldehyde
(Beijing Solarbio Science & Technology Co., Ltd., Beijing,
China) at 21–25°C for 15 min. The kidney tissues were embedded in
paraffin and 4-µm sections were cut. Certain paraffin sections were
separately stained with hematoxylin and eosin (H&E; 1% H for 7
min, 1% E for 4 min at 21–25°C), periodic acid-Schiff base (PAS;
1.1% Schiff's reagent for 30 min at 20°C) and Masson's trichrome
(Masson; 1% H for 5 min, 1% Ponceau S for 7 min, 2% Aniline blue
for 5 min at 21–25°C). The remaining paraffin sections were dewaxed
and microwaved for 10 min in citrate buffer for antigen retrieval.
Next, the sections were treated with 3% hydrogen peroxide
(H2O2) in methanol at 21–25°C for 30 min to
block the activity of endogenous peroxidase followed by rinsing in
PBS three times. The sections were incubated with antibodies
(Abcam, Cambridge, UK) against podocyte marker proteins [zona
occludens protein 1 (ZO-1; 1:100; cat. no. ab214228), Nephrin
(1:2,000; cat. no. ab216341) and P-cadherin (1:100; cat. no.
ab137729)], EMT marker proteins [α-SMA (1:50; cat. no. ab5694) and
FSP-1 (1:2,000; cat. no. ab197896)] and TGF-β and its associated
signaling pathway proteins [TGF-β receptor 1 (TGFBR1; 1:50; cat.
no. ab31013), TGFBR2 (1:100; cat. no. ab186838), Smad2/Smad3
(1:100; cat. no. ab217553), Smad4 (1:100; cat. no. ab40759), Smad7
(1:100; cat. no. ab216428), glycogen synthase kinase (GSK)-3β
(1:50; cat. no. ab75745) and β-catenin (1:500; cat. no. ab32572)]
overnight at 4°C. Next, goat anti-rabbit-horseradish peroxidase
(HRP)-conjugated IgG antibody (1:1,000; cat. no. ab6721; Abcam) was
added for 1 h at 37°C. Finally, the sections were developed with 1%
diaminobenzidine for 5 min, counterstained with 1% hematoxylin for
2 min at 21–25°C and mounted with neutral gum. Images of the
sections were captured at magnification, ×400 using a BX53 inverted
light microscope (Olympus Corporation, Tokyo, Japan).
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the kidney tissue using
TRIzol (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) and reverse transcription synthesis of cDNA was performed in a
single-step method using the Eastep® RT Master Mix kit
(Promega Corporation, Madison, WI, USA), according to the
manufacturer's protocol. qPCR was performed using the
GoTaq® 2-Step RT-qPCR System (Promega Corporation)
following the manufacturer's protocol. The thermocycling conditions
for RT-qPCR were 5 min at 99°C, then 15 sec at 94°C, 30 sec at 59°C
(40 cycles), 45 sec at 72°C. The fold-change in mRNA expression was
calculated with the 2−ΔΔCq method (16). Primer sequences are presented in
Table I.
 | Table I.Primer sequences of the
glomerulosclerosis associated proteins. |
Table I.
Primer sequences of the
glomerulosclerosis associated proteins.
| Gene | Primer
sequence |
|---|
| TGF-β1 | F:
5′-ATGAACCGACCCTTCCTGCT-3′ |
|
| R:
5′-TGTGTCCAGGCTCCAAATGT-3′ |
| Smad7 | F:
5′-CTCTTGCGAACATTACGGCT-3′ |
|
| R:
5′-CGAGATCAAGGTCGACCTGC-3′ |
| GADPH | F:
5′-CTACCCACGGCAAGTTCAAT-3′ |
|
| R:
5′-GGATGCAGGGATGATGTTCT-3′ |
| Smad2/3 | F:
5′-GGAAAGGGTTGCCACATGTT-3′ |
|
| R:
5′-AGAATCTCCGTGTGCCGAGG-3′ |
| Smad4 | F:
5′-ACGGCCATCTTCAGCACCAC-3′ |
|
| R:
5′-AGAATGCACAATCGCCGGAG-3′ |
| ZO-1 | F:
5′-GGCATTATTCGCCTTCATAC-3′ |
|
| R:
5′-GGAACACAACAATCGGATAC-3′ |
| GSK3β | F:
5′-CTTCAGGACAAGCGATTTA-3′ |
|
| R:
5′-CCAGCACCAGGTTAAGGTAG-3′ |
| Nephrin | F:
5′-CCCTCCGGGACCCTACTG-3′ |
|
| R:
5′-TCTGGGAGGATGGGATTGG-3′ |
| P-Cadherin | F:
5′-AAGTGCTGCAGCCAAAGACAGA-3′ |
|
| R:
5′-AGGTAGACCCACCTCAATCATCCTC-3′ |
| SMA | F:
5′-CAGCTATTGCCGTTCCAATTGA-3′ |
|
| R:
5′-CCAGGGCTTCATCATTGCA-3′ |
| FSP | F:
5′-ATACTCAGGCAACGAGGGTG-3′ |
|
| R:
5′-CTTCCGGGGCTCCTTATC-3′ |
| TGF-βIR | F:
5′-ACCATTTGGAGCCAGAACAC-3′ |
|
| R:
5′-GGTCTGAAGAGCTGAGCCTG-3′ |
| TGF-βIIR | F:
5′-CGAGCTCGGTGGAAGGTCTCATTTTATTG-3′ |
|
| R:
5′-CCCAAGCTTGGGATGTAAAAGACAAACAATG-3′ |
| β-catenin | F:
5′-GCTGACCAAAACTGCTAAATGACGA-3′ |
|
| R:
5′-TGTAGGGTCCCAGCGGTACAA-3′ |
Western blotting
Total protein was extracted from renal tissue using
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology, Shanghai, China) and protein concentration was
determined using bicinchoninic protein assay reagent (Beyotime
Institute of Biotechnology). First, 20 µg protein sample were
separated by 10% SDS-PAGE (Beijing Solarbio Science &
Technology Co., Ltd.) and transferred to polyvinylidene fluoride
membranes (EMD Millipore, Billerica, MA, USA), and then blocked
using 5% non-fat milk-Tris-buffered saline containing 0.05% Tween
20 (TBST) at 21–25°C for 2 h. Following washing with TBST, the
membranes were incubated with primary antibodies at 4°C overnight,
the primary antibodies were as follows: Anti-TGF-β1 (1:500; cat.
no. ab92486; Abcam); anti-ZO1 (1:1,000; ab96587; Abcam);
anti-Nephrin (1:1,000; cat. no. ab216341; Abcam); anti-P-cadherin
(1:2,000; cat. no. ab137729; Abcam); anti-α-SMA (1:100; cat. no.
ab5694; Abcam), anti-FSP-1 (1:1,000; cat. no. ab197896; Abcam);
anti-TGFBR2 (1:1,000; cat. no. ab186838; Abcam), anti-TGFBR1
(1:200; cat. no. ab31013; Abcam), anti-Smad2/Smad3 (1:1,000; cat.
no. ab202445; Abcam), anti-Smad4 (1:5,000; cat. no. ab40759;
Abcam), anti-Smad7 (1:500; cat. no. ab216428; Abcam), anti-GSK-3β
(1:1,000; cat. no. ab75745; Abcam), anti-β-catenin (1:5,000; cat.
no. ab32572; Abcam) and anti-GAPDH (1:2,500; cat. no. ab9485;
Abcam), followed by incubation with an HRP-conjugated secondary
antibody (1:10,000; cat. no. ab6721; Abcam) for 1 h at room
temperature. Finally, proteins were visualized by enhanced
chemiluminescence ECL (cat. no. WBKLS0010; EMD Millipore). The
images were acquired and analyzed with a ChemiDoc Touch imaging
system (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Statistical analysis
The results of immunohistochemistry and western
blotting were analyzed using Image J software version 1.8.0
(National Institute of Health, Bethesda, MD, USA). The results are
expressed as the mean ± standard deviation and each experiment
repeated three times. Significant differences were established by
one-way analysis of variance followed by Bonferroni post-hoc test
using the GraphPad Instat program version 3.0 (GraphPad, Inc., La
Jolla, CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
General appearance
Rats in the NC and SHO groups consumed a normal diet
and exhibited increased weight, good mental status, sensitive
response, shiny fur and solid stools; there was no significant
difference between these two groups (data not shown). Serum
biochemical indexes also exhibited no significant difference
between these two groups (Table
II). Therefore, the operation did not affect the experiment.
Rats in the GS groups demonstrated decreased food intake and became
lethargic, dispirited, and unresponsive. Additionally, they
demonstrated severe alopecia, dark hair, loose stools, discolored
urine and reduced weight. The levels of serum TP and Alb
significantly decreased, but TG, TC, BUN, SCr and UTP significantly
increased compared with the NC and SHO groups. Following QU
intervention in rats with GS, compared with in the GS group, the
general appearance of rats (QU I, QU II and QU III groups) was
improved in a dose-dependent manner. The general appearance of the
rats in the QU III group was near normal levels. The levels of
serum TP and Alb were significantly increased, and TG, TC, BUN, SCr
and UTP significantly decreased in a dose-dependent manner in the
QU I, QU II and QU III groups compared with the GS group (Table II).
| Table II.Biochemical parameters in all
groups. |
Table II.
Biochemical parameters in all
groups.
| Parameter | NC | SHO | GS | QU I | QU II | QU III |
|---|
| Kidney (g) | 1.11±0.02 |
1.20±0.09b |
2.45±0.26a,c |
2.40±0.26c,f |
2.22±0.15c,e |
1.97±0.14c,e |
| Weight (g) | 442.00±25.68 |
451.80±21.08b |
370.71±19.97a,c |
388.29±14.43c,f |
383.61±18.23c,e |
416.42±10.45c,e |
| TP (mM) | 62.00±3.19 |
60.78±2.08b |
52.90±2.24a,c |
61.46±4.25c,e |
57.91±2.91c,e |
60.61±1.80d,e |
| TG (mM) | 1.24±0.17 |
1.60±0.23b |
12.30±1.73a,c |
6.99±0.36c,e |
4.64±1.87c,e |
2.31±0.32c,e |
| Alb (g/l) | 38.08±1.49 |
36.06±1.22b |
14.20±1.24a,c |
22.37±2.11c,e |
23.15±4.35c,e |
30.84±1.72c,e |
| TC (mM) | 1.17±0.04 |
1.48±0.09b |
10.60±0.98a,c |
5.42±0.43c,e |
5.03±1.01c,e |
2.76±0.23c,e |
| BUN (mM) | 5.64±0.43 |
6.38±0.42b |
17.39±2.53a,c |
12.67±0.58c,e |
11.03±1.03c,e |
8.99±0.43c,e |
| SCr (mM) | 29.40±1.71 |
33.40±1.98b |
98.57±10.36a,c |
71.57±443c,a |
67.06±8.60c,e |
45.29±3.04c,e |
| UTP (mg/24 h) | 31.34±2.59 |
34.66±4.97b |
363.19±18.21a,c |
299.84±20.13c,e |
310.53±28.63c,e |
267.70±12.61c,e |
Biochemical parameters
Following 8 weeks of the study, there was no
significant difference in biochemical parameters between the NC
group and SHO group (P>0.05; Table
II). The effect of the surgery on the experiment can be
considered insignificant. Compared with the SHO group, the weight
of mice in the GS group was significantly reduced, while kidney
weight increased (P<0.05; Table
II). UTP in the GS group increased by ~10-fold, accompanied by
increased levels of TG, CHO, BUN and SCr and significantly
decreased levels of TP and Alb (P<0.05; Table II) in comparison to NC and SHO.
Biochemical parameters, including TG, Alb, TC, BUN and SCr in rats
with GS returned to nearly normal levels following treatment with
QU III.
Pathological evaluation
Based on direct observation, compared with the
normal group, the kidneys of the GS group were fibrotic (Fig. 1A). Additionally, enlargement,
damage, atrophy and hardening of the glomerulus, and interstitial
lymphocytic infiltration in the GS group were observed by H&E
staining (Fig. 1B). In addition,
Masson staining demonstrated that the interstitial cells of the
renal interstitium proliferated, the mesangial area increased in
mass and the basement membrane of the glomerulus exhibited focal
proliferation in the GS group compared with sham and NC rats
(Fig. 1C). PAS staining
demonstrated that the basement membrane of GS group rats became
wider and the mesangial matrix increased compared with sham and NC
rats (Fig. 1D). Following
treatment with increasing doses of QU, it was observed that the
pathological alterations of renal tissue were gradually relieved
and the fibrosis status of the kidneys gradually subsided and
became normal in the QU-treated group.
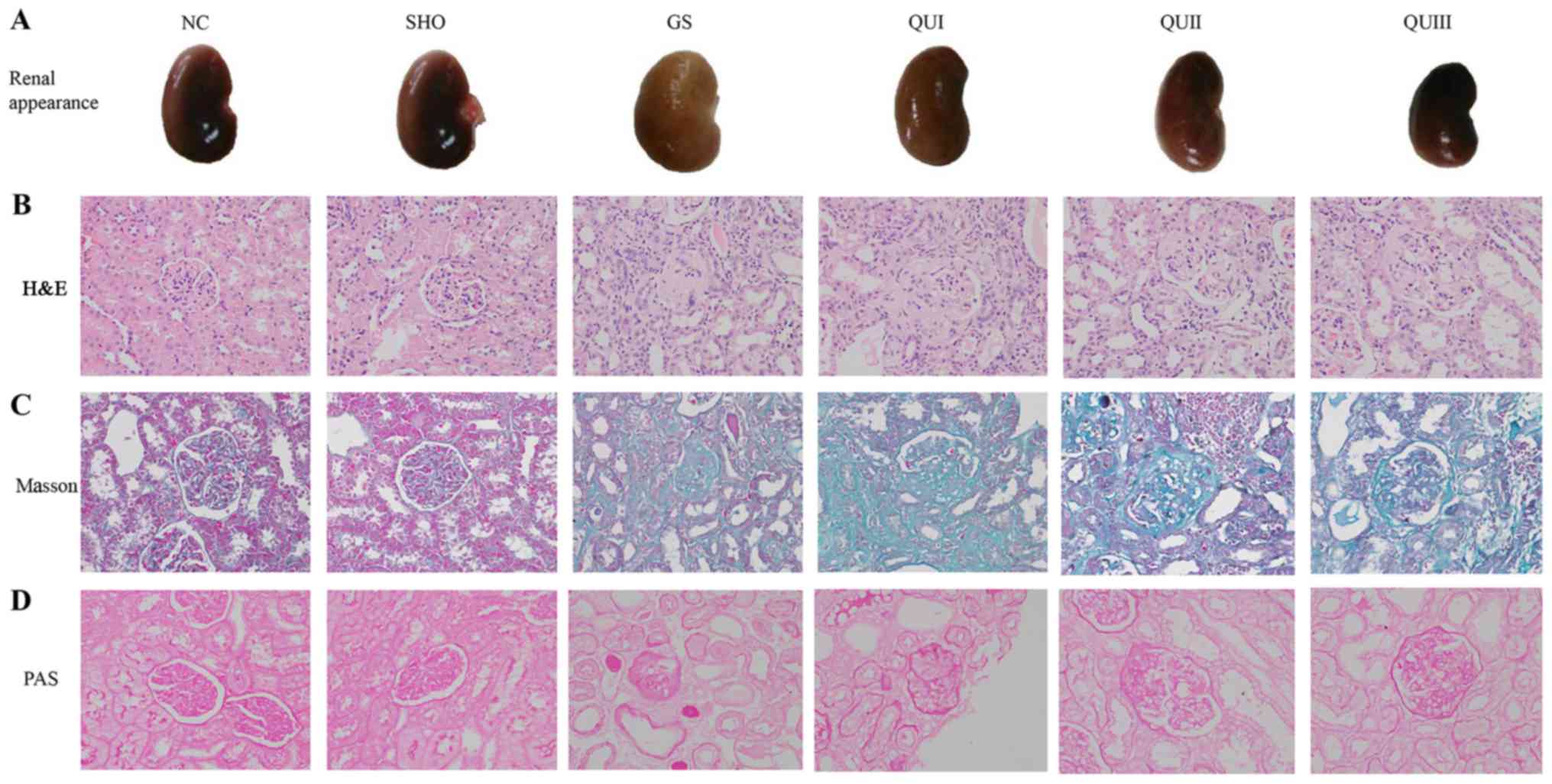 | Figure 1.Pathological analysis of renal tissue
following QU treatment in rats. (A) Observation of renal tissue in
rats. (B) H&E, (C) Masson and (D) PAS staining of rat kidney
tissue in all groups, magnification, ×400. NC, normal control; SHO,
sham operation group; GS, glomerulosclerosis; QU, quercetin; QU I,
QU 25 mg/kg/day; QU II, 50 mg/kg/day; QU III, 100 mg/kg/day;
H&E, hematoxylin and eosin; PAS, periodic acid-Schiff base. |
Effect of QU on transcription and
translation levels of GS-associated proteins
mRNA and protein in kidney tissue from each group
were extracted and detected by RT-qPCR, western blotting, and
immunohistochemistry. Podocyte marker proteins (ZO-1, Nephrin and
P-cadherin) and Smad7 mRNAs (Fig.
2A) exhibited significantly decreased expression in the GS
group compared with the NC group (P<0.001). Similarly, at the
translation level, these proteins exhibited low expression
(Fig. 2B) and distribution in the
GS group. The proteins were recovered following QU treatment and
their levels were altered in a dose-dependent manner. By contrast,
EMT marker proteins (α-SMA and FSP-1) and other TGF-β signaling
pathway-associated proteins (TGFBR2, TGFBR2, Smad2/Smad3, Smad4,
GSK-3β and β-catenin) exhibited increased expression in the GS
group compared with the NC group (Fig.
3), and distribution (Fig. 4)
of these proteins observed by immunohistochemistry exhibited
similar results. Following QU treatment, protein expression
decreased in a dose-dependent manner.
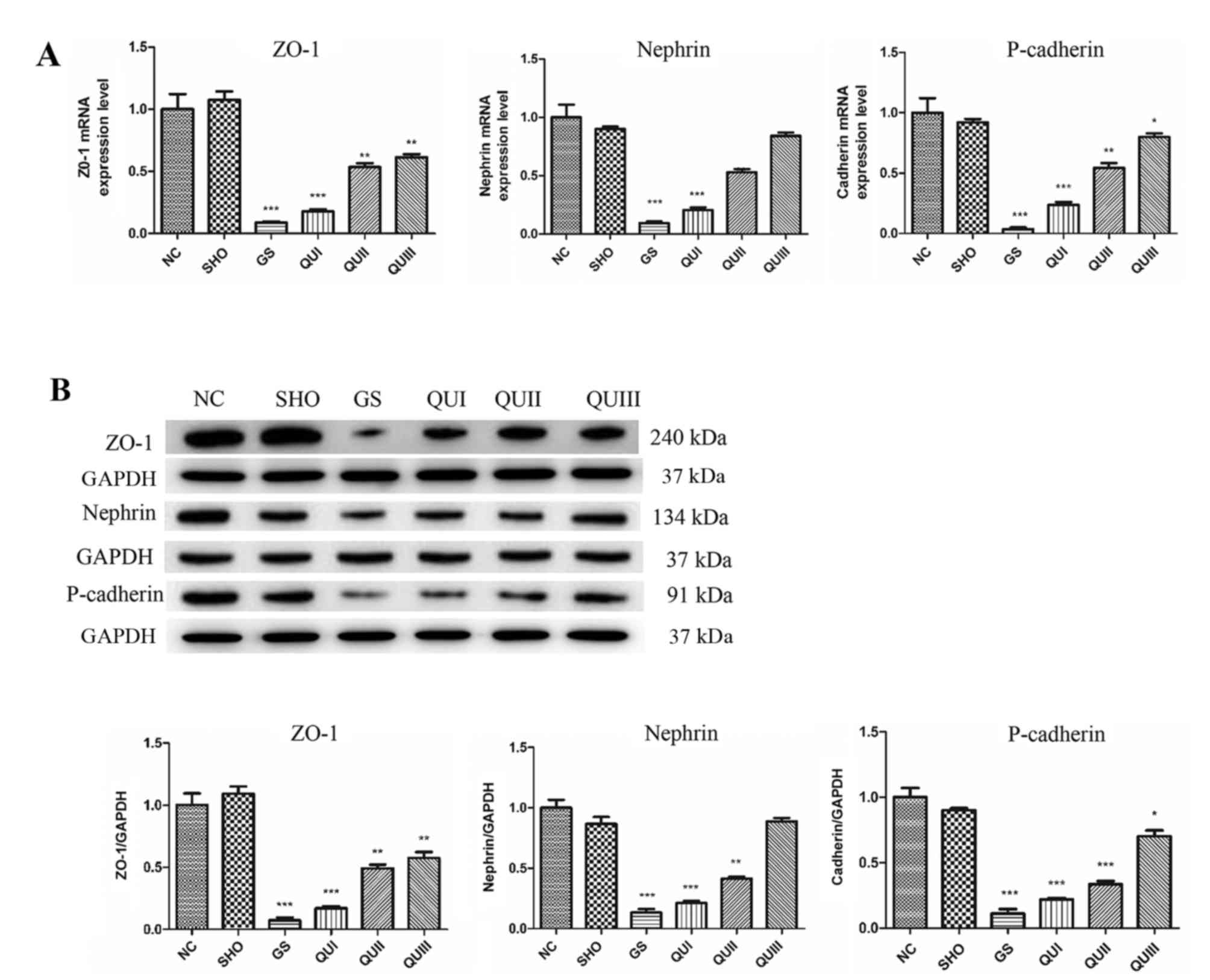 | Figure 2.Expression levels of ZO-1, Nephrin
and Cadherin following QU intervention. (A) mRNA and (B) protein
expression levels were detected by RT-qPCR and western blotting,
respectively. All data are presented as the mean ± standard
deviation of the mean. *P<0.05, **P<0.01 and ***P<0.001,
vs. the NC group. ZO-1, zona occludens protein 1; NC, normal
control; SHO, sham operation group; GS, glomerulosclerosis; QU,
quercetin; QU I, QU 25 mg/kg/day; QU II, 50 mg/kg/day; QU III, 100
mg/kg/day. |
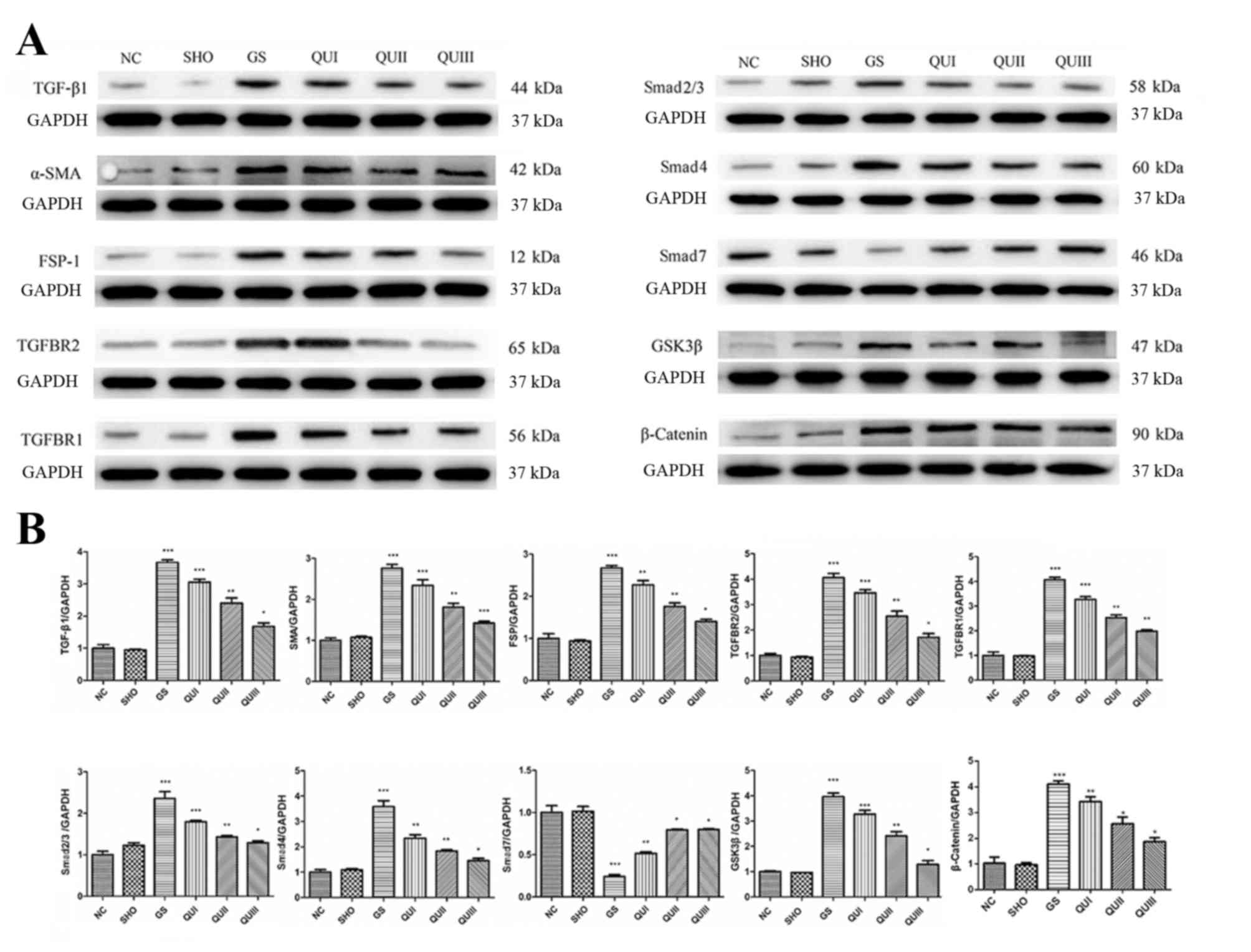 | Figure 3.Protein expression levels of
GS-associated proteins following QU intervention. (A) Western
blotting of GS-associated proteins in rats from all groups. (B)
Relative protein expression levels of GS-associated proteins in
rats from all groups. Magnification, ×400. All data are presented
as the mean ± standard deviation of the mean. *P<0.05,
**P<0.01 and ***P<0.001, vs. the NC group. NC, normal
control; SHO, sham operation group; GS, glomerulosclerosis; QU,
quercetin; QU I, QU 25 mg/kg/day; QU II, 50 mg/kg/day; QU III, 100
mg/kg/day; TGF-β, transforming growth factor-β; ZO-1, zona
occludens protein 1; SMA, smooth muscle actin; FSP-1,
fibroblast-specific protein-1; TGFBR, transforming growth factor β
receptor; Smad, mothers against decapentaplegic homolog 9; GSK,
glycogen synthase kinase. |
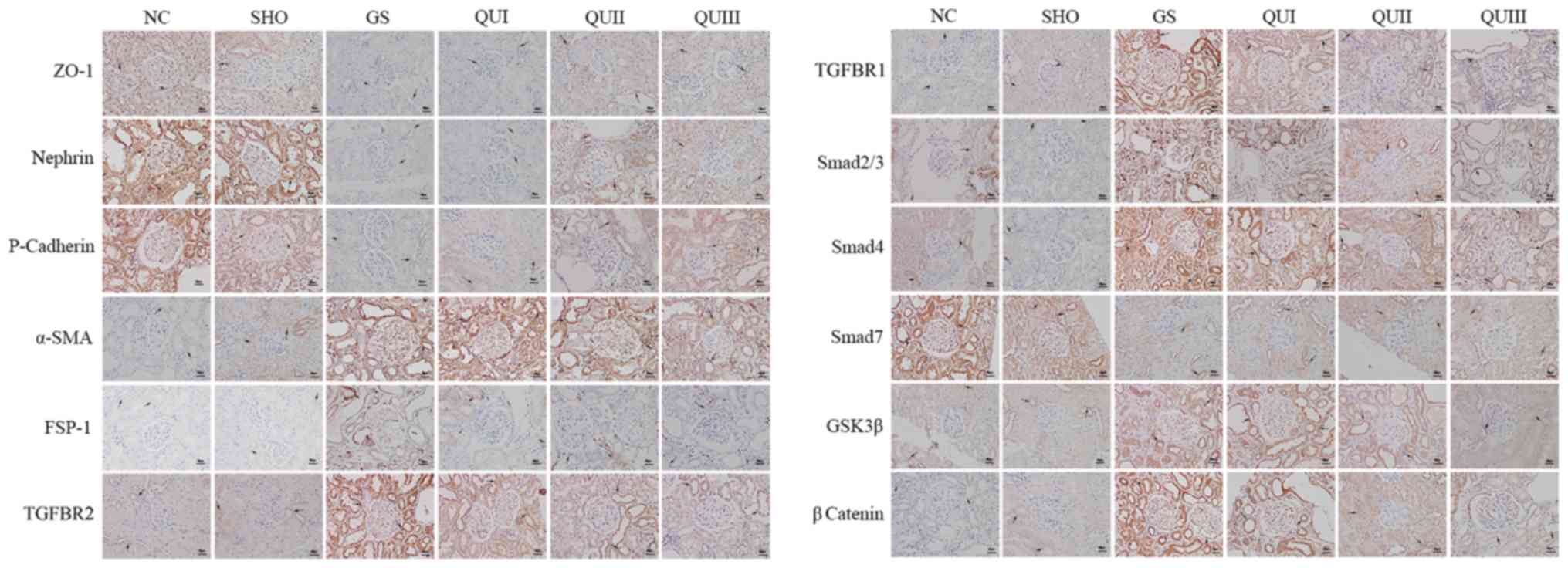 | Figure 4.Detection of the distribution of
GS-associated proteins by immunohistochemistry. Magnification,
×400. Brown represents protein expression and increasingly dark
colors represent increased protein expression. The arrows indicated
the alterations in morphology. Scale bar, 5 µm. NC, normal control;
SHO, sham operation group; GS, glomerulosclerosis; QU, quercetin;
QU I, QU 25 mg/kg/day; QU II, 50 mg/kg/day; QU III, 100 mg/kg/day;
ZO-1, zona occludens protein 1; SMA, smooth muscle actin; FSP-1,
fibroblast-specific protein-1; TGFBR, transforming growth factor β
receptor; Smad, mothers against decapentaplegic homolog 9; GSK,
glycogen synthase. |
Discussion
To investigate the efficacy and mechanism of QU in
GS, an ADR-induced GS mouse model (GS group) was constructed which
demonstrated similarities to human renal disease (17). The main experimental manifestations
of these mice were proteinuria, hyperlipidemia, hypoproteinemia,
hypoxemia and renal hypofunction (18,19).
Accordingly, certain biochemical indices of the mice were altered,
such as increased SCr, BUN, TG and UTP (20). In the present study, the associated
biochemical indices (SCr, BUN, TC, TG, UTP, TP and Alb) were
examined following establishment of the rat model, and the results
agreed with those of previous reports (20,21).
Together with the examination of general appearance and
pathological sections of the mice, these results indicated that the
renal system of rats in the GS group had swelling and fibrosis,
their functions were severely impaired, and absorption and
filtration capabilities were weakened. Following treatment with QU,
the physiological indices of GS rats began to recover. Therefore,
QU may be useful for in the treatment of GS. QU can promote the
filtration, protein absorption and lipid metabolism in the kidney,
as well as inhibit glomerular fibrosis and improve the renal
function of glomerulosclerotic rats.
GS is associated with the activation of various
cytokines signal transduction pathways. Numerous studies confirm
that TGF-β is a major cytokine causing GS (22–25).
Therefore, TGF-β and its signal transduction pathway have become
important targets for studies of renal diseases caused by various
factors (26). There are two main
TGF-β receptors: TGFBR2 and TGFBR1 (27). Activated TGF-β can bind TGF-BR1 and
then adsorb TGF-BR1 to form heterodimers on the cell membrane that
activate downstream effectors. Activated TGFBR1 induces
phosphorylation of Smad2 and Smad3, and the phosphorylation of
Smad2 or Smad3 can bind Smad4 to form oligomers, followed by
translocation into the nucleus to act as a transcription factor
(28–30). However, Smad7 can inhibit the
phosphorylation of Smad2 and Smad3 by directly interacting with the
TGFBR, and ultimately block the TGF-β signaling pathway (31). In the present study, the expression
levels of TGFBR2, TGFBR1, Smad2/Smad3 and Smad4 were elevated in
the GS group, and the expression of Smad7 was inhibited. In
addition, during EMT there was a significant increase in α-SMA and
FSP-1, which are transcriptionally regulated by the TGF-β signal
pathway to promote GS (29,32).
TGF-β signaling can lead to tumorigenesis or tumor suppression
(33,34). It induces EMT to promote the
invasion and metastasis of cancer cells (34). Disruption of the transcriptional
network in the EMT process can improve the tumor suppressive
functions of TGF-β (35).
Myofibroblasts are characterized by their high expression of α-SMA
induced by elevated expression of TGF-β1 receptors (36). The expression of FSP-1 is increased
in a TGF-β/Smad-dependent manner in systemic sclerosis skin.
Additionally, FSP-1 overexpression or stimulation induced an
activated phenotype in resting normal fibroblasts; however,
knockdown of S100A4 attenuated the pro-fibrotic effects of TGF-β
(37). The expression level of the
TGF-β upstream target proteins GSK-3β (38) and β-catenin (39) were also significantly decreased by
QU treatment of GS rats in the current study. Additionally, the
expression levels of ZO-1 (40),
Nephrin (41) and P-cadherin
(42), which are podocyte marker
proteins are negatively associated with podocyte apoptosis, were
decreased in GS rats. Following treatment with QU, the expression
of podocyte marker proteins (ZO-1, Nephrin and P-cadherin), TGF-β
signaling associated proteins (TGF-β1, TGFBR1/2, Smad2/3, Smad4,
Smad7, GSK-3β and β-catenin), EMT associated proteins (α-SMA and
FSP-1) returned to normal levels. Following QU treatment, the
development of EMT was attenuated in podocytes with GS, potentially
via downregulating TGF-β signaling proteins (TGF-β1, TGFBR1/2,
Smad2/3, Smad4, GSK-3β and β-catenin) and EMT proteins (α-SMA and
FSP-1), and upregulating Smad7. This suggests that QU can protect
the kidneys from GS by inhibiting the TGF-β signal pathway.
In the present study, it was demonstrated that QU
protects renal function and alleviates the progression of GS in
rats. The mechanism may involve inhibition of the TGF-β signaling
pathway. QU is a natural flavonoid that is widely distributed in
green plants, including fruits and vegetables. Although QU had some
negative impact on embryonic development according to an in
vitro study (43) and prenatal
exposure resulted in a small increase in the risk of cancer in mice
offspring (44), QU has been well
tolerated in human studies. Doses up to 1,000 mg/day did not
produce adverse effects on blood parameters, liver and kidney
function, hematology or serum electrolytes for several months
(45–49). As a result of the low toxicity and
side effects of QU, it may be clinically applicable. Therefore,
this study suggests that QU can be used to treat GS. The present
study used an in vivo rat model to demonstrate that QU
treatment attenuates glomerulosclerosis via the TGF-β signaling
pathway, but lacked in vitro experiments. Therefore, the
association between QU and TGF-β signal pathway in vitro and
the interference experiment of TGF-β1 inhibition in QU treatment
remains to be investigated.
Acknowledgements
Not applicable.
Funding
This work was supported by grants from the National
Natural Science Foundation of China (grant no. 81360602), and the
Science and Technology Development Project of Chengguan District,
Gansu, Lanzhou (grant no. 2016-1-8).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Author's contributions
ED designed and supervised these experiments; YL
performed most of experiments involved in this study, including the
establishment of animal models, pathological section preparation,
immunohistochemistry and so on, collated and analyzed the
experimental data and wrote the manuscript; JY was responsible for
the PCR and western blot analysis. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The protocol was approved prior to commencing this
study by the animal ethics committee of Gansu University of Chinese
Medicine.
Patient consent for publication
Not applicable.
Competing interest
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
TGF-β
|
transforming growth factor-β
|
|
GS
|
glomerulosclerosis
|
|
QU
|
quercetin
|
|
EMT
|
epithelial mesenchymal transition
|
|
FSP-1
|
fibroblast-specific protein-1
|
|
α-SMA
|
α-smooth muscle actin
|
|
ADR
|
adriamycin
|
|
SHO
|
sham operation group
|
|
UTP
|
urine total protein
|
|
TP
|
total protein
|
|
Abl
|
albumin
|
|
TG
|
triglyceride
|
|
TC
|
total cholesterol
|
|
BUN
|
urea nitrogen
|
|
SCr
|
serum creatinine
|
|
H&E
|
hematoxylin and eosin
|
|
PAS
|
periodic acid-Schiff base
|
|
TBST
|
Tris-buffered saline containing
Tween-20
|
References
|
1
|
Zou R, Xu G, Liu XC, Han M, Jiang JJ,
Huang Q, He Y and Yao Y: PPARgamma agonists inhibit TGF-beta-PKA
signaling in glomerulosclerosis. Acta Pharmacol Sin. 31:43–50.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nagata M: Podocyte injury and its
consequences. Kidney Int. 89:1221–1230. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Taneda S, Honda K, Ohno M, Uchida K, Nitta
K and Oda H: Podocyte and endothelial injury in focal segmental
glomerulosclerosis: An ultrastructural analysis. Virchows Arch.
467:449–458. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang C, Blough E, Arvapalli R, Dai X,
Triest WE, Leidy JW, Masannat Y and Wu M: Acetaminophen attenuates
glomerulosclerosis in obese Zucker rats via reactive oxygen
species/p38MAPK signaling pathways. Free Radic Biol Med. 81:47–57.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Qian Y, Feldman E, Pennathur S, Kretzler M
and Brosius FC III: From fibrosis to sclerosis: Mechanisms of
glomerulosclerosis in diabetic nephropathy. Diabetes. 57:1439–1445.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou TB and Qin YH: The signaling pathways
of LMX1B and its role in glomerulosclerosis. J Recept Signal
Transduct Res. 32:285–289. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hodgin JB, Bitzer M, Wickman L, Afshinnia
F, Wang SQ, O'Connor C, Yang Y, Meadowbrooke C, Chowdhury M,
Kikuchi M, et al: Glomerular aging and focal global
glomerulosclerosis: A podometric perspective. J Am Soc Nephrol.
26:3162–3178. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Arauz J, Rivera-Espinoza Y, Shibayama M,
Favari L, Flores-Beltrán RE and Muriel P: Nicotinic acid prevents
experimental liver fibrosis by attenuating the prooxidant process.
Int Immunopharmacol. 28:244–251. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu L, Lin W, Zhang Q, Cao W and Liu Z:
TGF-β induces miR-30d down-regulation and podocyte injury through
Smad2/3 and HDAC3-associated transcriptional repression. J Mol Med
(Berl). 94:291–300. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nabavi SF, Russo GL, Daglia M and Nabavi
SM: Role of quercetin as an alternative for obesity treatment: You
are what you eat! Food Chem. 179:305–310. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ma JQ, Li Z, Xie WR, Liu CM and Liu SS:
Quercetin protects mouse liver against CCl4-induced
inflammation by the TLR2/4 and MAPK/NF-κB pathway. Int
Immunopharmacol. 28:531–539. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pan HC, Jiang Q, Yu Y, Mei JP, Cui YK and
Zhao WJ: Quercetin promotes cell apoptosis and inhibits the
expression of MMP-9 and fibronectin via the AKT and ERK signalling
pathways in human glioma cells. Neurochem Int. 80:60–71. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Baruah MM, Khandwekar AP and Sharma N:
Quercetin modulates Wnt signaling components in prostate cancer
cell line by inhibiting cell viability, migration, and metastases.
Tumour Biol. 37:14025–14034. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Peng H, Liu Y, Wang J, Zhao X and Wang X:
Effect of quercetin on the expression of TGF-beta1 in human
embryonic lung fibroblasts activated by the silicotic alveolar
macrophages. Wei Sheng Yan Jiu. 42:99–102. 2013.(In Chinese).
PubMed/NCBI
|
|
15
|
Steru L, Chermat R, Thierry B and Simon P:
The tail suspension test: A new method for screening
antidepressants in mice. Psychopharmacology (Berl). 85:367–370.
1985. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bertani T, Rocchi G, Sacchi G, Mecca G and
Remuzzi G: Adriamycin-induced glomerulosclerosis in the rat. Am J
Kidney Dis. 7:12–19. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhou TB, Qin YH, Lei FY, Su LN, Zhao YJ
and Huang WF: apoE expression in glomerulus and correlation with
glomerulosclerosis induced by adriamycin in rats. Ren Fail.
33:348–354. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu S, Jia Z, Zhou L, Liu Y, Ling H, Zhou
SF, Zhang A, Du Y, Guan G and Yang T: Nitro-oleic acid protects
against adriamycin-induced nephropathy in mice. Am J Physiol Renal
Physiol. 305:F1533–F1541. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang YU, Zhou N, Wang H, Wang S and He J:
Effect of Shenkang granules on the progression of chronic renal
failure in 5/6 nephrectomized rats. Exp Ther Med. 9:2034–2042.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou TB, Qin YH, Lei FY, Su LN, Zhao YJ
and Huang WF: All-trans retinoic acid regulates the expression of
apolipoprotein E in rats with glomerulosclerosis induced by
Adriamycin. Exp Mol Pathol. 90:287–294. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ding Y and Choi ME: Regulation of
autophagy by TGF-β: Emerging role in kidney fibrosis. Semin
Nephrol. 34:62–71. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wan YG, Che XY, Sun W, Huang YR, Meng XJ,
Chen HL, Shi XM, Tu Y, Wu W and Liu YL: Low-dose of multi-glycoside
of Tripterygium wilfordii Hook. f., a natural regulator of
TGF-β1/Smad signaling activity improves adriamycin-induced
glomerulosclerosis in vivo. J Ethnopharmacol. 151:1079–1089. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fukuda A, Minakawa A, Sato Y, Iwakiri T,
Iwatsubo S, Komatsu H, Kikuchi M, Kitamura K, Wiggins RC and
Fujimoto S: Urinary podocyte and TGF-β1 mRNA as markers for disease
activity and progression in anti-glomerular basement membrane
nephritis. Nephrol Dial Transplant. 32:1818–1830. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yoon JJ, Lee YJ, Namgung S, Han BH, Choi
ES, Kang DG and Lee HS: Samchuleum attenuates diabetic renal injury
through the regulation of TGF-β/Smad signaling in human renal
mesangial cells. Mol Med Rep. 17:3099–3108. 2018.PubMed/NCBI
|
|
26
|
Kim YS, Xu ZG, Reddy MA, Li SL, Lanting L,
Sharma K, Adler SG and Natarajan R: Novel interactions between
TGF-{beta}1 actions and the 12/15-lipoxygenase pathway in mesangial
cells. J Am Soc Nephrol. 16:352–362. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
McKnight AJ, Savage DA, Patterson CC,
Sadlier D and Maxwell AP: Resequencing of genes for transforming
growth factor beta1 (TGFB1) type 1 and 2 receptors (TGFBR1,
TGFBR2), and association analysis of variants with diabetic
nephropathy. BMC Med Genet. 8:52007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Das F, Ghosh-Choudhury N, Bera A, Dey N,
Abboud HE, Kasinath BS and Choudhury GG: Transforming growth factor
β integrates Smad 3 to mechanistic target of rapamycin complexes to
arrest deptor abundance for glomerular mesangial cell hypertrophy.
J Biol Chem. 288:7756–7768. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang L, Liu C, Meng XM, Huang C, Xu F and
Li J: Smad2 protects against TGF-β1/Smad3-mediated collagen
synthesis in human hepatic stellate cells during hepatic fibrosis.
Mol Cell Biochem. 400:17–28. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang H, Li G, Wu JJ, Wang L, Uhler M and
Simeone DM: Protein kinase A modulates transforming growth factor-β
signaling through a direct interaction with Smad4 protein. J Biol
Chem. 288:8737–8749. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schiffer M, Bitzer M, Roberts IS, Kopp JB,
ten Dijke P, Mundel P and Böttinger EP: Apoptosis in podocytes
induced by TGF-beta and Smad7. J Clin Invest. 108:807–816. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Larsson SO, Hedner U and Nilsson IM: On
fibrinolytic split products in serum and urine in uraemia. Scand J
Urol Nephrol. 5:234–242. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Guasch G, Schober M, Pasolli HA, Conn EB,
Polak L and Fuchs E: Loss of TGFbeta signaling destabilizes
homeostasis and promotes squamous cell carcinomas in stratified
epithelia. Cancer Cell. 12:313–327. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Heldin CH, Vanlandewijck M and Moustakas
A: Regulation of EMT by TGFβ in cancer. FEBS Lett. 586:1959–1970.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
David CJ, Huang YH, Chen M, Su J, Zou Y,
Bardeesy N, Iacobuzio-Donahue CA and Massagué J: TGF-β tumor
suppression through a lethal EMT. Cell. 164:1015–1030. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sriram S, Gibson DJ, Robinson P, Pi L,
Tuli S, Lewin AS and Schultz G: Assessment of anti-scarring
therapies in ex vivo organ cultured rabbit corneas. Exp Eye Res.
125:173–182. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tomcik M, Palumbo-Zerr K, Zerr P, Avouac
J, Dees C, Sumova B, Distler A, Beyer C, Cerezo LA, Becvar R, et
al: S100A4 amplifies TGF-β-induced fibroblast activation in
systemic sclerosis. Ann Rheum Dis. 74:1748–1755. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yan Q, Luo H, Wang B, Sui W, Zou G, Chen H
and Zou H: Correlation between PKB/Akt, GSK-3β expression and
tubular epithelial-mesenchymal transition in renal allografts with
chronic active antibody-mediated rejection. Exp Ther Med.
13:2217–2224. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Toraldo G, Bhasin S, Bakhit M, Guo W,
Serra C, Safer JD, Bhawan J and Jasuja R: Topical androgen
antagonism promotes cutaneous wound healing without systemic
androgen deprivation by blocking β-catenin nuclear translocation
and cross-talk with TGF-β signaling in keratinocytes. Wound Repair
Regen. 20:61–73. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wagner MC, Rhodes G, Wang E, Pruthi V,
Arif E, Saleem MA, Wean SE, Garg P, Verma R, Holzman LB, et al:
Ischemic injury to kidney induces glomerular podocyte effacement
and dissociation of slit diaphragm proteins Neph1 and ZO-1. J Biol
Chem. 283:35579–35589. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Jia J, Ding G, Zhu J, Chen C, Liang W,
Franki N and Singhal PC: Angiotensin II infusion induces nephrin
expression changes and podocyte apoptosis. Am J Nephrol.
28:500–507. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Teixeira Vde P, Blattner SM, Li M, Anders
HJ, Cohen CD, Edenhofer I, Calvaresi N, Merkle M, Rastaldi MP and
Kretzler M: Functional consequences of integrin-linked kinase
activation in podocyte damage. Kidney Int. 67:514–523. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Pérez-Pastén R, Martinez-Galero E and
Chamorro-Cevallos G: Quercetin and naringenin reduce abnormal
development of mouse embryos produced by hydroxyurea. J Pharm
Pharmacol. 62:1003–1009. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Vanhees K, de Bock L, Godschalk RW, van
Schooten FJ and van Waalwijk van Doorn-Khosrovani SB: Prenatal
exposure to flavonoids: Implication for cancer risk. Toxicol Sci.
120:59–67. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Heinz SA, Henson DA, Austin MD, Jin F and
Nieman DC: Quercetin supplementation and upper respiratory tract
infection: A randomized community clinical trial. Pharmacol Res.
62:237–242. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Jin F, Nieman DC, Shanely RA, Knab AM,
Austin MD and Sha W: The variable plasma quercetin response to
12-week quercetin supplementation in humans. Eur J Clin Nutr.
64:692–697. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Nieman DC, Henson DA, Davis JM, Dumke CL,
Gross SJ, Jenkins DP, Murphy EA, Carmichael MD, Quindry JC,
McAnulty SR, et al: Quercetin ingestion does not alter cytokine
changes in athletes competing in the western states endurance run.
J Interferon Cytokine Res. 27:1003–1011. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Scholten SD and Sergeev IN: Long-term
quercetin supplementation reduces lipid peroxidation but does not
improve performance in endurance runners. Open Access J Sports Med.
4:53–61. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Shanely RA, Knab AM, Nieman DC, Jin F,
McAnulty SR and Landram MJ: Quercetin supplementation does not
alter antioxidant status in humans. Free Radic Res. 44:224–231.
2010. View Article : Google Scholar : PubMed/NCBI
|














