Introduction
Periodontitis is a chronic inflammatory disease with
periodontal ligament inflammation and alveolar bone destruction
(1). It is initiated by bacterial
plaque, which induces inflammatory immune responses to perturb the
bone homeostasis between bone construction and bone resorption
(2). Although several studies have
explored the complex regulatory mechanisms underlying alveolar bone
loss, in the pathogenies of periodontitis, these are unclear
(3).
Interleukin-17A (IL-17A) is a pro-inflammatory
cytokine typically secreted by Th17 cells and contributes to the
pathophysiology of periodontitis, notably aggravating gingival
inflammation and alveolar bone loss (4). Emerging clinical studies have
revealed that human periodontitis is related to the increased
expression of IL-17 in peripheral blood, gingival crevicular fluid,
gingival tissues, and alveolar bone when compared to a healthy
periodontal group (5–7). Additionally, the increased number of
IL-17-positive cells and osteoclasts were accumulated in the
alveolar bone of periodontitis patients, indicating a causal
relevance of IL-17 and osteoclasts in periodontitis (8). However, the underlying mechanism of
IL-17A-mediated inflammatory periodontal bone destruction is yet to
be elucidated.
Furthermore, osteoclasts are specialized
multinucleated cells derived from macrophage precursors and
activated to absorb the alveolar bone, ultimately resulting in bone
loss in periodontitis (9).
Notably, IL-17A was revealed to possess valid osteoclastogenic
properties that stimulate the expression of the receptor activator
of nuclear factor-κB ligand (RANKL), interleukin 1β (IL-1β), tumor
necrosis factor-α (TNF-α), and prostaglandin E2. Of note, the
targets of IL-17A contain osteoblasts, fibroblasts, and other
stromal cells, indicating that IL-17A mediates osteoclast activity
by promoting osteoclastogenesis factors on other cells (10–12).
However, the study of osteoclast differentiation under the direct
effect of IL-17A on osteoclast precursors is poorly understood.
Autophagy is an intracellular metabolic pathway,
which orchestrates diverse aspects of cellular stress response, and
the core molecular machinery of autophagy consists of
autophagy-related proteins (13,14).
Recently, autophagy has been demonstrated to be involved in
osteoclast differentiation and bone resorption (15,16).
Autophagy-related proteins, including Atg5, Atg7, Atg4B, and
microtubule-associated protein 1 light chain 3 (LC3) are
significantly responsible for the formation of ruffled border and
facilitation of osteoclast polarization, ultimately resulting in
bone resorption (17). Moreover,
emerging evidence has indicated that several pro-inflammatory
cytokines, including IL-1β and TNF-α, trigger autophagy in murine
osteoclasts (18,19). However, whether autophagy is
involved in IL-17A-mediated osteoclastogenesis requires further
exploration.
In the present study, the effects of IL-17A on
osteoclast differentiation and bone resorption of BMMs were
investigated, and whether autophagy was involved in this process
was analyzed. Moreover, the effect of IL-17A on osteoclast activity
was detected on the alveolar bone surfaces in a rat periodontitis
model.
Materials and methods
Materials
Soluble recombinant mouse IL-17A was obtained from
PeproTech, Inc. (Rocky Hill, NJ, USA). Macrophage
colony-stimulating factor (M-CSF) and RANKL were obtained from
R&D Systems, Inc. (Minneapolis, MN, USA). All cell culture
media and supplements were procured from Gibco; Thermo Fisher
Scientific, Inc. (Waltham, MA, USA). Reagents for real-time
quantitative polymerase chain reaction (PCR) were obtained from
Takara Bio, Inc. (Otsu, Japan). RNA was extracted using TRIzol
reagent from Thermo Fisher Scientific, Inc., according to the
manufacturer's protocol. Antibodies against LC3B (cat. no. 192890;
1:2,000) and β-actin (cat. no. 8226; 1:1,000) were purchased from
Abcam (Cambridge Science Park, Cambridge, UK). An antibody against
IL-17A (cat. no. GB11110; 1:100) was purchased from Servicebio
(Wuhan, Hubei, China). Goat anti-rabbit immunoglobulin G secondary
antibody (cat. no. BA1054; 1:5,000) was obtained from Boster
Biological Technology, Ltd. (Wuhan, Hubei, China). A bicinchoninic
acid (BCA) kit and enhanced chemiluminescence (ECL) detection
reagent was purchased from Thermo Fisher Scientific, Inc. TRAP
staining was conducted using a Leukocyte Acid Phosphatase kit
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). Cell viability was
assessed using Cell Counting Kit-8 (CCK-8) (Sigma-Aldrich; Merck
KGaA).
Animals
All animal experiments were performed in accordance
with the principles and procedures of the National Institutes of
Health (NIH) for the Care and Use of Laboratory Animals. The
approval number granted by the Animal Ethics Committee of The
Second Affiliated Hospital of Zhejiang University School of
Medicine is 2017–052 (2017-2-10). All surgeries were performed
under sodium pentobarbital anesthesia, and all efforts were made to
minimize suffering. A total of 10 female 7-week-old C57BL/6 mice
(22±2 g), and 30 male 7-weeks of age Sprague-Dawley rats (200±50
g), were purchased from the China Experimental Animal Center
(Hangzhou, China). Throughout the experiment, the animals were
housed in individual cages under an appropriate temperature
(21±2°C), with 12-h light/dark cycles and humidity (50±5%) and fed
standard laboratory food and tap water ad libitum.
Cell culture
BMMs were obtained as previously described (20). Briefly, C57BL/6 female mice were
euthanized by cervical dislocation after being anesthetized with
sodium pentobarbital (150 mg/kg body weight, intraperitoneally).
The femora and tibiae were dissected from mice, and the ends were
cut off with scissors. Then, a 1-µl syringe was used to flush the
marrow cavity with α-MEM medium supplemented with 10% fetal bovine
serum, 100 U/µl penicillin, 100 U/µl streptomycin, and 30 ng/µl
M-CSF. The mixture was homogenized and incubated in a Petri dish at
37°C and 5% CO2 for 3 days. The adherent cells were
collected as BMMs.
Osteoclast formation
BMMs were seeded into 96-well plates at a density of
1×104 cells/well in the presence of M-CSF (30 ng/µl),
RANKL (50 ng/µl), and different concentrations of recombinant
IL-17A (0, 0.1, 1 or 10 ng/µl). The media were changed every
alternate day until multinucleated cells appeared under a Leica
light microscope (Leica, Solms, Germany).
TRAP staining
When the multinucleated osteoclasts were formed, the
medium was aspirated, the cells were washed with phosphate-buffered
saline (PBS), fixed with 4% paraformaldehyde at room temperature
for 20 min, and stained for TRAP using a leukocyte acid phosphatase
kit. After the staining, multinucleated osteoclasts which contained
3 or more nuclei, could be identified by counting the number of
nuclei in an osteoclast using a Leica light microscope (Leica).
Bone resorption assay in vitro
BMMs were seeded on bone slices at a density of
1×104 cells/slice in 96-well culture plates in 3
replicates in the presence of 30 ng/µl M-CSF. After 24 h, the cells
were treated with 30 ng/µl M-CSF, 50 ng/µl RANKL, and different
concentrations of IL-17A (0, 0.1, 1 and 10 ng/µl). The medium was
replaced every alternate day until the formation of mature
osteoclasts. Then, the bone slices were removed, washed with PBS,
and dried. The bone resorption pits were observed using a scanning
electron microscope (TM-100; Hitachi, Ltd., Tokyo, Japan).
Detection of F-actin ring
formation
After mature osteoclasts formed, the cells were
fixed with 4% paraformaldehyde at room temperature for 20 min,
permeabilized with 0.1% Triton X-100 at room temperature for 10
min, blocked with 1% BSA for 30 min, and incubated with
TRITC-labeled phalloidin (Invitrogen; Thermo Fisher Scientific,
Inc.,) for 2 h. DAPI was used for staining the cell nuclei.
Subsequently, the F-actin ring formation was observed using a Leica
fluorescence microscope (Leica).
Transmission electron microscopy
Cells were fixed with 2.5% glutaraldehyde for 30
min, and post-fixed in 1% buffered osmium tetroxide for 1 h at
20°C. Then, the samples were dehydrated by an increasing ethanol
gradient and embedded in epoxy resin. Ultrathin sections (60–80 nm)
were sliced using an ultramicrotome (Leica EM UC7; Leica), and
observed under a transmission electron microscope (Tecnai G2 20
TWIN; FEI; Thermo Fisher Scientific, Inc.).
Cell viability assay
BMMs were plated in 96-well plates (1×103
cells/well) for 24 h. Then, the cells treated with IL-17 were
incubated with or without 3-MA (2 nM) for 48 h. Cell viability was
assessed using a CCK-8 assay. Briefly, 10 µl of CCK-8 solution was
added to each well at the indicated time-points, and then the plate
was incubated for 2 h in a 5% CO2 incubator. Absorbance
was assessed at 450 nm with a spectrophotometer (Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
RNA extraction and real-time
quantitative polymerase chain reaction
Total RNA of the cells was extracted using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) and reverse
transcribed to synthesize single-stranded cDNA using PrimeScript
Reverse Transcription Master Mix Kit (Takara Bio, Inc.). The
expression of the target genes was quantified using a SYBR
polymerase chain reaction Master Mix Kit (Takara Bio, Inc.) on a
StepOne Plus real-time polymerase chain reaction system (Applied
Biosystems; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. The relative expression level of the
target genes was calculated using the relative quantitative method
2−ΔΔCq (21), and
normalized to the mouse glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) gene. The primer sequences are listed in Table I.
 | Table I.Primers used for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primers used for reverse
transcription-quantitative polymerase chain reaction.
| Gene symbol | Primer sequence
(5′-3′) |
|---|
| LC3 | F:
CGGAGCTTTGAACAAAGAGTG |
|
| R:
TCTCTCACTCTCGTACACTTC |
| Beclin-1 | F:
AGCTCAGTACCAGCGGGAGT |
|
| R:
TGGAAGGTGGCATTGAAGAC |
| Atg5 | F:
TGTGCTTCGAGATGTGTGGTT |
|
| R:
GTCAAATAGCTGACTCTTGGCAA |
| c-Fos | F:
CGGGTTTCAACGCCGACTA |
|
| R:
TTGGCACTAGAGACGGACAGA |
| NFATc1 | F:
GGAGAGTCCGAGAATCGAGAT |
|
| R:
TTGCAGCTAGGAAGTACGTCT |
| CatK | F:
CTCGGCGTTTAATTTGGGAGA |
|
| R:
TCGAGAGGGAGGTATTCTGAGT |
| TRAP | F:
CACTCCCACCCTGAGATTTGT |
|
| R:
CCCCAGAGACATGATGAAGTCA |
| GAPDH | F:
GACTTCAACAGCAACTCCCAC |
|
| R:
TCCACCACCCTGTTGCTGTA |
Western blot analysis
The total protein of the cells was extracted using
cell lysate buffer and quantified using a bicinchoninic acid kit
(Thermo Fisher Scientific, Inc.). An equivalent of 25 µg of the
total protein was resolved on a ١٢٪ gel using sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and transferred to a
polyvinylidene fluoride (PVDF) membrane (Bio-Rad Laboratories,
Inc.). Subsequently, the membrane was blocked with 5% skim milk at
room temperature for 2 h and probed with the primary antibodies
anti- LC3B (cat. no. 192890; 1:2,000) and anti-β-actin (cat. no.
8226; 1:1,000) purchased from Abcam. Incubation with the membranes
was performed overnight at 4°C. Then, the membrane was washed and
incubated with a horseradish peroxidase-conjugated secondary
antibody (1:5,000; cat. no. BA1054; Boster Biological Technology,
Ltd.) at room temperature for 1.5 h. The immunoreactive bands were
visualized using an ECL kit, and the data were analyzed using
ImageJ software, version 1.48 [National Institutes of Health (NIH),
Bethesda, MD, USA].
Experimental periodontitis
induction
Sprague-Dawley rats were randomly divided into 3
experimental groups: NC (normal control), NS (normal saline) and
the IL-17 group. To induce a rat periodontitis model, as previously
described (22,23), the rats were anesthetized with
sodium pentobarbital (40 mg/kg body weight) via intraperitoneal
injection; 3.0 cotton threads were placed bilaterally around the
intraorbital portion of the rat maxillary first molar to induce
experimental periodontitis in NS and IL-17 groups, while the NC
group was subjected to sham treatment. In addition, 20 µl normal
saline and 20 µl recombinant IL-17A (5 µg/µl) were injected into
the bilateral maxillary first molars of the rats every other day in
the NS and IL-17 groups under anesthesia with sodium pentobarbital
(40 mg/kg body weight) via intraperitoneal injection, respectively.
During the experiments, rats had no weight loss, no loss of
appetite, no serious infection, and no weakness or sudden
death.
Histomorphometric analysis and
immunohistochemical staining
All the rats were euthanized after being
anesthetized with sodium pentobarbital (150 mg/kg,
intraperitoneally) at 1-month after injury induction and death was
ascertained mainly based on complete cessation of a heartbeat. The
maxilla was dissected carefully, immediately fixed in 4%
paraformaldehyde for 1 day, and decalcified in 10%
ethylenediaminetetraacetic acid buffer at room temperature for 6
weeks. The samples were embedded in paraffin, and 4-µm thick
sections were obtained in the proximal mandible for hematoxylin and
eosin (H&E) staining. The distance between the cementum-enamel
junction (CEJ) and the alveolar callus (ABC) in the interproximal
region of the maxillary first molar was observed using a Leica
optical microscope (Leica). TRAP staining was performed using a
leukocyte acid phosphatase kit to quantify the osteoclasts as
described by the manufacturer. Multinucleated TRAP+
cells on the alveolar bone surface around the first molar were
enumerated as active osteoclasts. Immunohistochemical staining was
performed to detect the level of IL-17A and LC3, using the
respective antibodies (1:100 dilution) according to the
manufacturer's protocol. Finally, the sections were observed under
a Leica light microscope and positive cells were counted using
ImageJ software (NIH).
Statistical analysis
All quantitative data are presented as the mean ±
standard deviation. Each experiment was performed in triplicate.
Statistical analyses were performed using one-way analysis of
variance (ANOVA), followed by Tukey's post hoc test with GraphPrism
7 software (GraphPad Software, Inc., La Jolla, CA, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
IL-17A facilitates osteoclast
differentiation and bone resorption of mouse bone marrow
macrophages
To detect the effects of IL-17A on osteoclast
differentiation and bone resorption, BMMs were cultured in medium
with or without IL-17A. The hallmark of osteoclast differentiation
is the formation of TRAP+ multinucleated cells (24). The present results revealed that
the number of TRAP+ multinucleated cells was
significantly increased with increasing concentration of IL-17A
(Fig. 1A and D). In addition, the
formation of filamentous actin (F-actin) rings is a characteristic
of terminally differentiated osteoclasts, and bone resorption pits
can be quantified for determining the bone loss (24). Immunofluorescence staining revealed
that the circumference of the actin ring was larger as compared to
the control groups, which indicated that increasing the areas of
contact with the bone surface enhanced the function of bone
resorption after the addition of IL-17A (Fig. 1C). In addition, the areas of the
bone absorption pits were markedly promoted by IL-17A treatment
(Fig. 1B and E). These results
indicated increased osteoclastogenesis of BMMs in vitro
after addition of IL-17A.
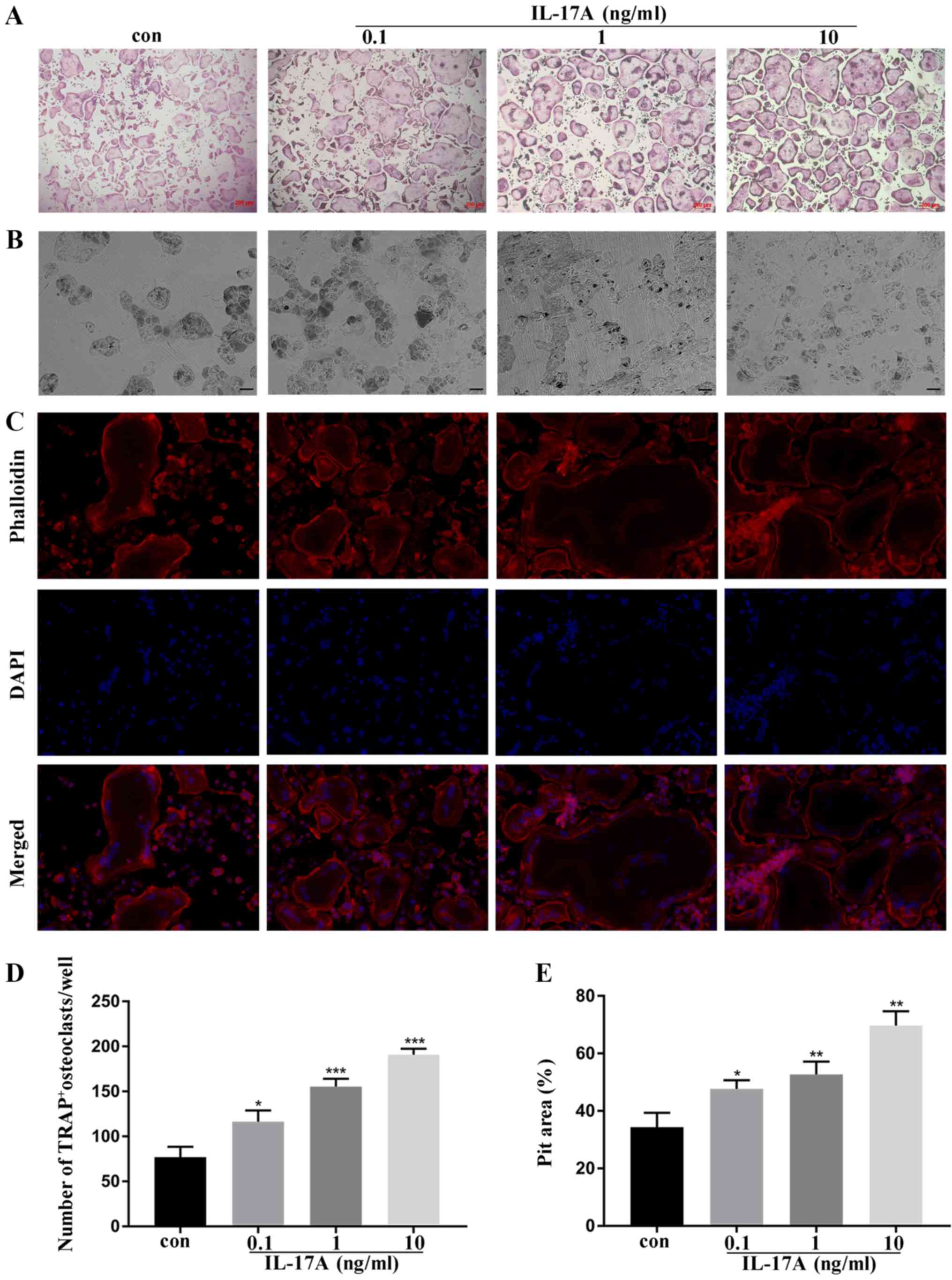 | Figure 1.IL-17A exerts positive effects on
osteoclast formation in bone marrow macrophages in vitro.
(A) The representative TRAP+ multinucleated cells/well are
presented (magnification, ×10), scale bar, 200 µm. (B) The
resorption pits of the bone slices were visualized with a scanning
electron microscope (magnification, ×200) scale bar, 50 µm. (C)
Immunofluorescence staining was performed with phalloidin (red) and
DAPI (blue). Images were obtained and analyzed using a Leica
fluorescence microscope (magnification, ×40) scale bar, 20 µm. (D)
Quantitative analysis was implemented to assess the TRAP+
multinucleated cells/well. (E) Quantitative analysis was performed
on resorption pit areas. *P<0.05, **P<0.01, ***P<0.001
compared to the control group (mean ± standard deviation). IL-17A,
interleukin IL-17A; TRAP, tartrate-resistant acid phosphatase;
F-actin, filamentous actin; con, control group. |
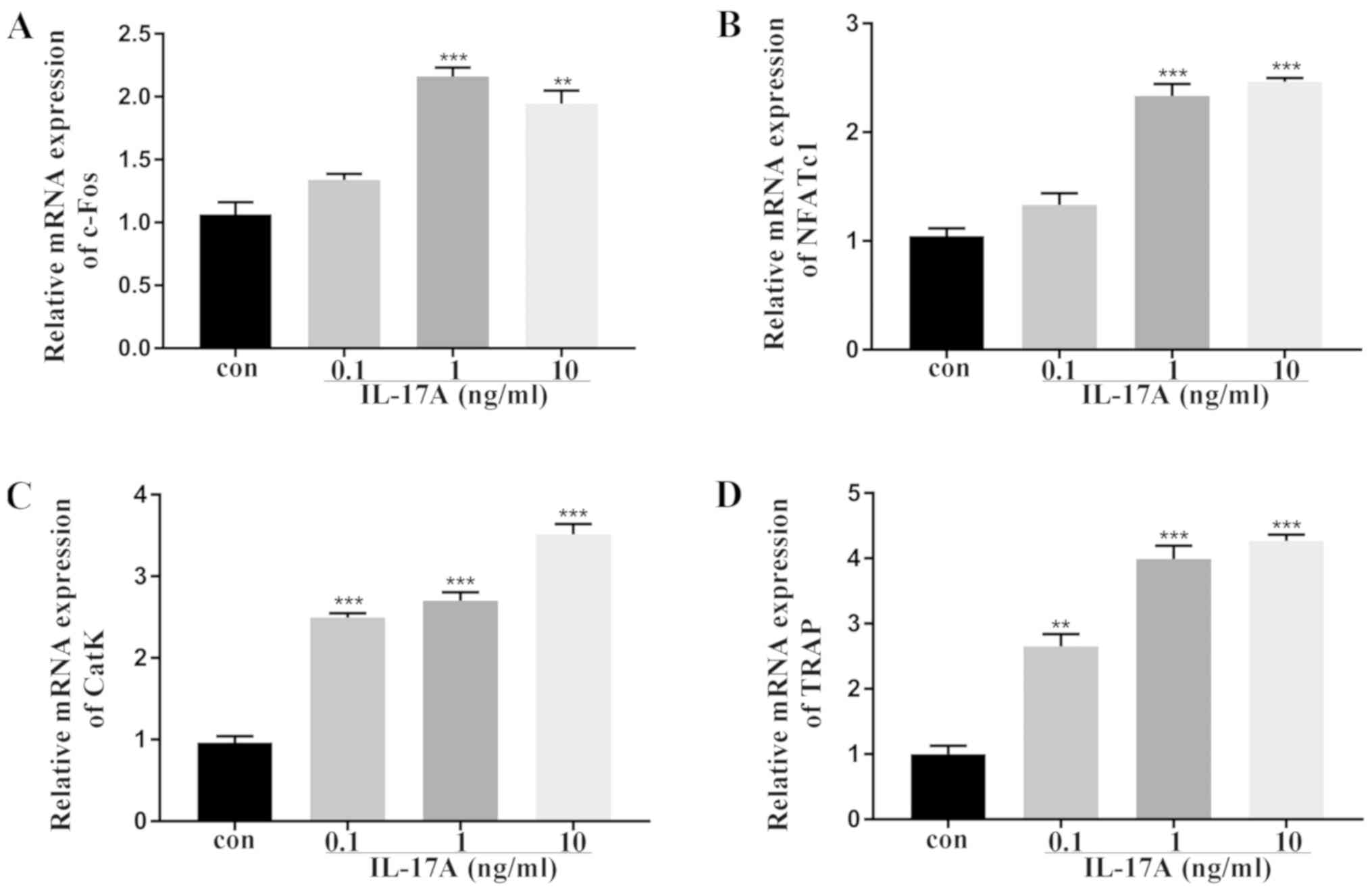
Effect of IL-17A on the expression of
osteoclast-related genes in vitro
To further investigate the effects of IL-17A on
osteoclast differentiation, the expression of osteoclast-related
genes was examined by real-time quantitative PCR. c-Fos and NFATc1
are the osteoclastogenic transcription factors required for early
osteoclast differentiation, while TRAP and CatK are designated as
terminal osteoclast differentiation markers (24–26).
Furthermore, slightly increased expression of c-Fos and NFATc1 was
observed post-IL-17A treatment (Fig.
2A and B). Moreover, IL-17A significantly increased the
expression TRAP and CatK as compared to the control group (Fig. 2C and D).
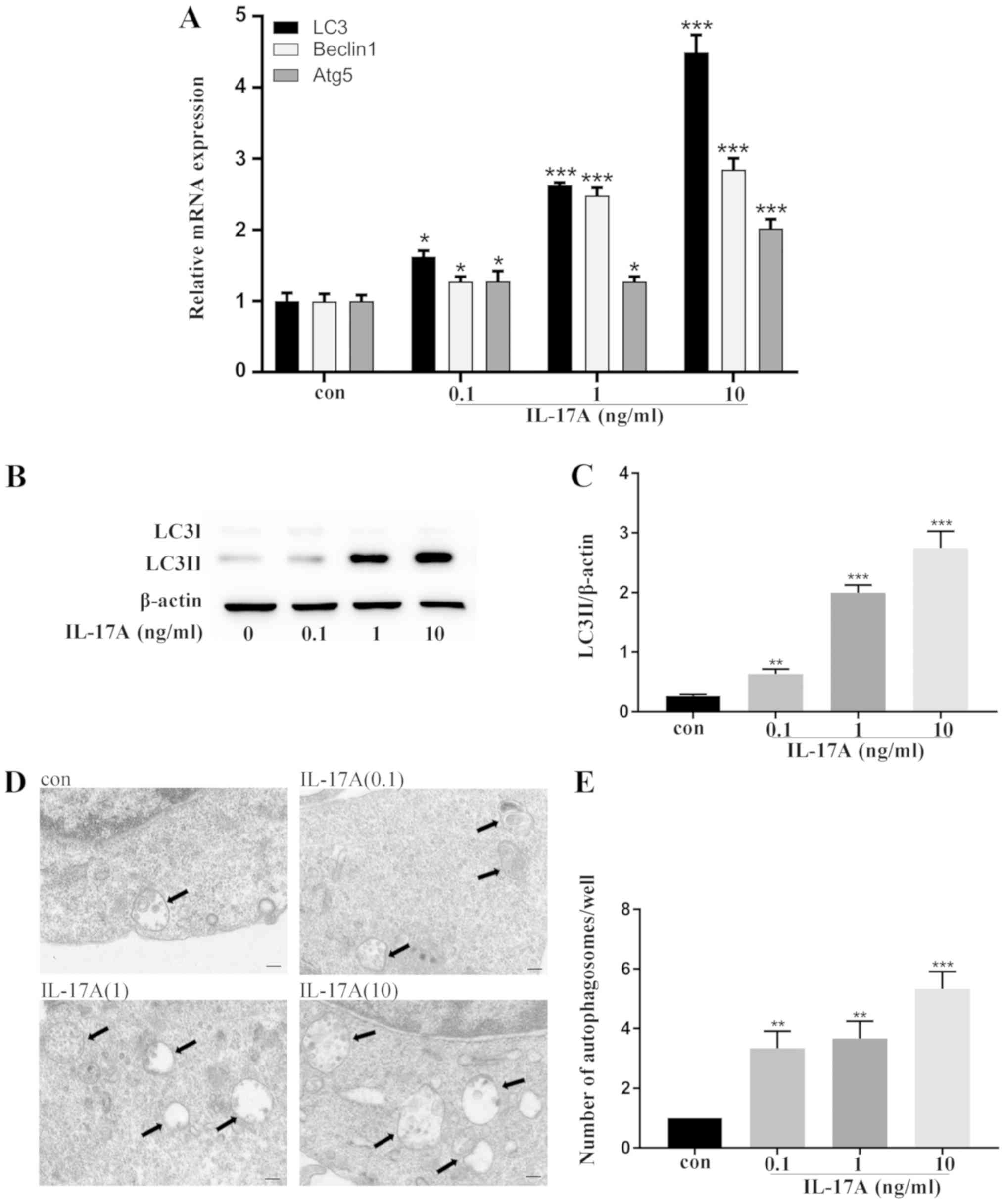
IL-17A facilitates autophagy activity
during osteoclast differentiation
The specific mechanism underlying IL-17A-facilitated
osteoclast differentiation was hypothesized to be the autophagic
pathway. Thus, real-time quantitative PCR was employed and it was
revealed that the expression of the autophagy-related genes, LC3,
Beclin1 and Atg5 was increased (Fig.
3A). Next, the expression of LC3-II protein, which is the
autophagy marker in autophagosome membranes was investigated
(14). Western blot analysis
revealed that the expression of LC3-II was upregulated after the
addition of IL-17A (Fig. 3B and
C), indicating enhanced autophagosome formation. Notably, an
increased number of autophagosomes with typical bilayer membrane
morphology was observed by transmission electron microscopy
(Fig. 3D and E). The
aforementioned findings indicated that autophagy activity was
enhanced in IL-17A-mediated osteoclast differentiation.
Inhibition of autophagy attenuates
IL-17A-mediated osteoclastogenesis
To elucidate whether autophagy in IL-17A-treated
BMMs exerted a positive effect, autophagy inhibitor 3-MA, a
prominent inhibitor reducing autophagy flux in the early stages,
was added. Consequently, no detrimental effect was noted on the
viability of osteoclasts after the treatment with 3-MA (Fig. 4A). The data revealed that the
expression of LC3-II was significantly downregulated with the
treatment of 3-MA (Fig. 4B and E).
Consecutively, the number of TRAP+ multinuclear cells
(Fig. 4C and F), the area of pits
on bone slices (Fig. 4D and G),
and the mRNA level of osteoclast-related genes, CatK and TRAP
(Fig. 4H and I), were decreased
after the addition of 3-MA in IL-17A-treated BMMs. These results
demonstrated that the autophagy inhibitor could weaken the effect
on IL-17A-mediated osteoclast activity.
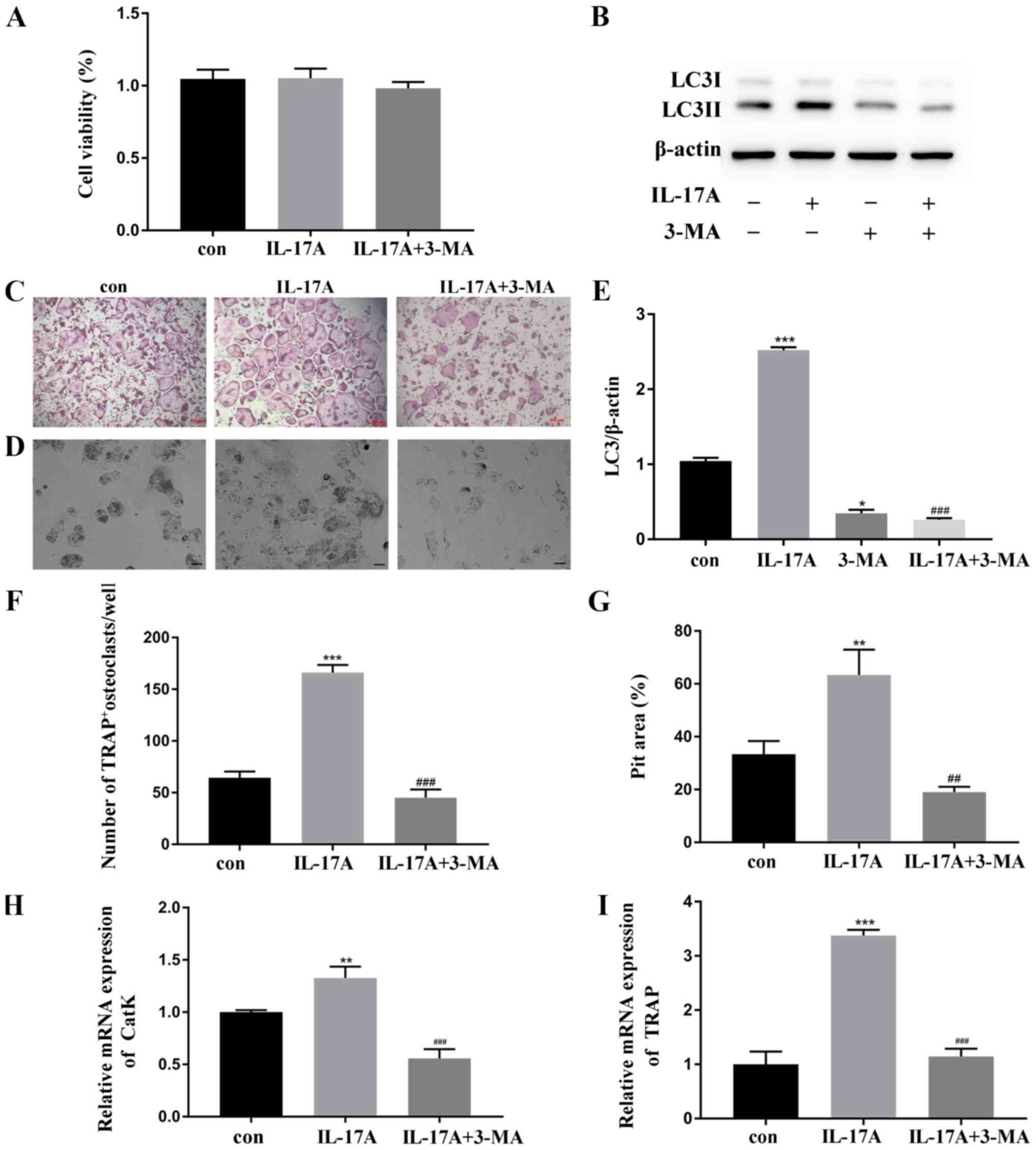 | Figure 4.Inhibition of autophagy abrogates
IL-17A-mediated osteoclastogenesis. (A) Cell viability was measured
by CCK-8 assay. BMMs were incubated with or without IL-17A (10
ng/µl) for 48 h, and 3-MA (10 nM) was added into IL-17A medium. (B)
Western blot analysis was used to analyze the expression of LC3.
(C) Fixed cells were stained for TRAP, and representative TRAP+
multinucleated cells/well are displayed. Magnification, ×10; Scale
bar, 200 µm. (D) The resorption pits of the bone slices were
visualized using a scanning electron microscope after 10 days of
culture. Magnification, ×200; scale bar, 50 µm. (E) Densitometric
analysis of LC3-II expression. The cells were pretreated with 3-MA
(2 nM) for 2 h, and then, with RANKL (50 ng/µl) plus IL-17A (10
ng/µl) for 5 days. (F) Quantitative analysis was implemented to
assess the TRAP+ multinucleated cells/well. (G) Quantitative
analysis was performed on resorption pit areas. The
osteoclast-related genes (H) CatK and (I) TRAP were analyzed by
real-time quantitative polymerase chain reaction. Data were
normalized to the GAPDH expression and presented as the fold-change
relative to the control group (mean ± standard deviation).
*P<0.05, **P<0.01, ***P<0.001 compared to the control
group. ##P<0.01, ###P<0.001, compared to the IL-17A-treated
group. IL-17A, interleukin IL-17A; 3-MA, 3-methyladenine; LC3,
microtubule-associated protein 1 light chain 3; CatK, cathepsin K;
TRAP, tartrate-resistant acid phosphatase; RANKL, receptor
activator of nuclear factor-κB ligand; GAPDH,
glyceraldehyde-3-phosphate dehydrogenase; con, control group. |
Effect of IL-17A on osteoclast
activity in vivo
A rat model of experimental periodontitis was
established to detect the effect of IL-17A on osteoclasts.
Immunohistochemistry demonstrated that a marked level of IL-17A in
the periodontal tissue was detected after IL-17A injection when
compared to normal saline injection (Fig. 5A and B). H&E staining revealed
that the attachments of the collagenous fibers were destroyed, and
a large number of inflammatory cells were revealed in both the
connective tissue and the epithelium layer in the NS and IL-17
groups when compared to the NC group. Additionally, a prominently
enhanced average distance was noted between the CEJ and ABC in the
IL-17 groups when compared to the NS group (Fig. 5C and D). In addition, TRAP staining
exposed higher numbers of TRAP+ multinuclear cells in
the IL-17 groups (Fig. 5E and F).
Moreover, an increase in LC3 staining was observed on the
inflammatory alveolar bone surfaces after IL-17 treatment (Fig. 5G and H). Collectively, these
results revealed that IL-17A could increase osteoclastogenesis to
exacerbate alveolar bone resorption in the periodontitis rat
model.
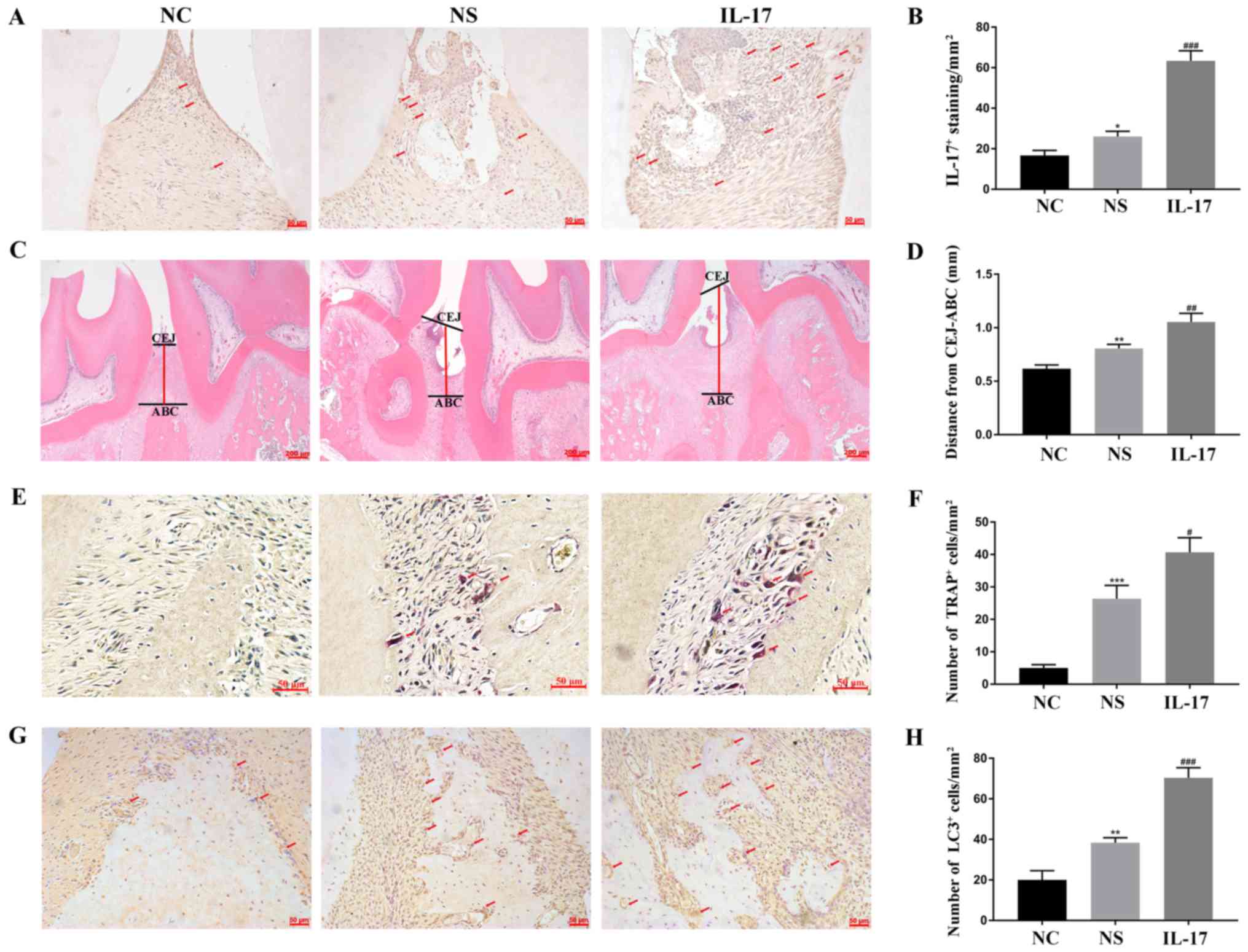 | Figure 5.Effects of IL-17A on osteoclast
activity in vivo. (A) Immunohistochemical staining was used
to determine the level of IL-17A expression (arrows). Scale bar, 50
µm. (B) IL-17A expression was analyzed quantitatively. (C) The
distance between CEJ and ABC (red line) was observed by H&E
staining. Scale bar, 200 µm. (D) Quantitative analysis was
performed on the distance. (E) TRAP staining was used to detect the
osteoclasts (red arrows). Scale bar, 50 µm. (F) Quantitative
analysis was performed on the number of TRAP+ cells. (G)
Immunohistochemical staining was used to examine the level of LC3
expression (arrows). Scale bar, 50 µm. (H) Quantitative analysis
was performed on LC3 level. *P<0.05, **P<0.01, ***P<0.001
compared to the NC group. #P<0.05, ##P<0.01, ###P<0.001
compared to the NS group. IL-17A, interleukin IL-17A; H&E
staining, hematoxylin and eosin staining; CEJ, cementum-enamel
junction; ABC, the alveolar callus; TRAP, tartrate-resistant acid
phosphatase; LC3, microtubule-associated protein 1 light chain 3;
NC, normal control group; NS, normal saline group; IL-17A, IL-17A
group. |
Discussion
In periodontitis, microbes and toxic products in
bacterial plaques induce local inflammatory reactions in the
periodontal tissues due to a large-scale production of inflammatory
mediators, which can promote osteoclast activity and aggravate
alveolar bone resorption (27).
Accumulating evidence has revealed an interdisciplinary approach
combing the study of bone and immune system, regarded as
osteoimmunology, for the treatment of the disease (28).
Osteoclasts are exclusive bone resorbing cells that
could secrete lysosomal enzymes into the extracellular space to
degrade bone matrix (24). The
effects of IL-17A, a pro-inflammatory cytokine, were examined on
osteoclast differentiation and bone resorption in vitro. It
was found that the number of TRAP+ multinucleated cells,
and the formation of actin rings accompanied by the pit area of
bone resorption were increased in response to increased
concentrations of IL-17A, thereby indicating that IL-17A could
facilitate osteoclast differentiation and bone resorption. A
similar result was deduced that IL-17 exhibited a positive effect
on osteoclast differentiation of human-derived bone marrow cells
(29). Further study revealed that
IL-17 upregulated the expression of RANK on human osteoclast
precursors to enhance the sensitivity to RANKL signaling,
culminating in bone resorption (30). Notably, the differentiation of
osteoclasts from macrophage precursors is dependent on the presence
of RANKL, and RANKL-RANK interaction is essential for the
activation of various signaling cascades of osteoclastogenesis
(26,31). Considering these findings, it was
speculated that in the present study, the strong enhancement of
osteoclast activity promoted by IL-17A of BMMs is attributed to the
synergistic effect with RANKL.
However, Kitami et al (32) observed suppression of osteoclast
differentiation of RAW264.7 as a result of the treatment of IL-17.
IL-17 markedly suppressed the osteoclastogenesis of canine bone
marrow-derived macrophages at a maximal concentration (33). Collectively, these studies
contradicted the present experimental results, which may be
attributed to various factors, including different experimental
conditions and species. Therefore, theories on the activity of
IL-17A in osteoclast differentiation are yet controversial, and
thus, exploring the specific underlying mechanisms is
imperative.
Osteoclast-specific genes contribute to various
stages of osteoclast formation and activation (26). c-Fos is one of the key
transcription factors in the AP-1 family, which is required for the
early phase of osteoclast differentiation (34). NFATc1 is the hallmark of osteoclast
fate, and is essential for the survival and differentiation of
osteoclast precursors (35). CatK
and TRAP are largely regarded as terminal osteoclast
differentiation markers and highly expressed by activated
osteoclasts to mediate the function of bone resorption (36,37).
Moreover, mature osteoclasts exhibit bone resorption capacity by
secreting protons, CatK and so forth. In the present study, the
expression of c-Fos, NFATc1, CatK, and TRAP genes was
increased with the treatment of IL-17A, indicating that IL-17A
could promote the osteoclast differentiation in BMMs.
Autophagy, a conserved lysosomal degradation
process, is essential for cell survival, differentiation, and
development and is regulated by some autophagy-related genes
(38). Emerging evidence has
revealed that autophagy was activated in response to several
pro-inflammatory cytokines, such as IL-1β and TNFα accompanied by a
robust increase in osteoclast differentiation and bone resorption
(18,39). The present findings supported the
hypothesis that IL-17A, a pro-inflammatory cytokine, enhanced the
autophagic activity including the increased expression of an
autophagy-related gene, accumulation of LC3-II, as well as
autophagosome formation along with osteoclast differentiation.
Moreover, the addition of autophagy inhibitor 3-MA could reduce the
level of osteoclast-related markers, thereby attenuating the
osteoclast formation and bone resorption after the treatment with
IL-17A. Thus, this result confirmed our speculation that autophagy
exhibits the positive effect on osteoclast activity under IL-17
stimulation.
In addition, Ke et al (40) observed that IL-17 facilitated
osteoclast precursor autophagy at a low concentration, and the
activation of the RANKL/JNK pathway may be involved in
IL-17-enhanced osteoclastogenesis. In the present study, it was
also revealed that p-JNK and p-p38 were increased after addition of
IL-17 (data not shown), which suggested that mitogen-activated
protein kinase (MAPK) pathways including JNK and p38 may mediate
autophagic activity of osteoclasts after addition of IL-17.
However, more specific mechanisms require revealing to elucidate
which pathway may exist in IL-17-enhanced autophagy and
osteoclastogenesis.
It is well accepted that autophagy, as a highly
conserved intracellular metabolic mechanism, has an anti-apoptotic
effect. Xue et al (41),
revealed that IL-17 modulated apoptosis of osteoclast precursors,
RAW264.7, to influence osteoclastogenesis. However, RAW264.7 is a
cell line which differs from the primary mouse bone marrow
macrophages that we used (42).
Therefore, whether IL-17A has an effect on the apoptosis of
osteoclasts from primary bone marrow macrophages requires further
study.
Notably, recent studies have demonstrated that the
knockdown of Beclin1 (43) or p62
(44) using small interfering RNA
(siRNA) resulted in the inhibition of osteoclastogenesis, and
further deletion of Atg7 prevented the TNFα-induced bone loss in
mice (45). Similar results
revealed that the pharmacological or genetic inactivation of
autophagy could ameliorate bone loss by inhibiting osteoclast
differentiation. This phenomenon could implicate the direct
translation targeting autophagy, which in turn, would provide
promising therapeutic approaches in the prevention and treatment of
inflammatory bone resorption.
Nonetheless, the number of osteoclasts was increased
along with the exacerbation of alveolar bone resorption in response
to IL-17A treatment in a rat periodontitis model. A previous study
revealed that IL-17A gene transfer increased the level of
osteoclasts and biomarkers of bone resorption in collagen-induced
arthritis, indicating that IL-17A is crucial for pathological bone
loss via direct activation of osteoclasts (46). Further studies demonstrated that
blocking of IL-17 with IL-17 antibody or IL-17-deficient mice
suppressed joint damage in experimental arthritis, rendering IL-17
as an intervention target for inflammatory bone resorption in
rheumatoid arthritis (12,47–49).
Notably, rheumatoid arthritis exhibits specific similarities with
periodontitis with respect to inflammatory diseases, wherein
inflammatory cytokines accelerate bone and joint destruction
(50). However, although IL-17 is
ascribed to joint disruption, the specific mechanism in
periodontitis has not been reported yet, necessitating further
investigation.
In conclusion, the present study revealed that
IL-17A directly promoted osteoclast differentiation and exacerbated
bone resorption in vitro and in vivo, and autophagic
activity was involved in the IL-17A-mediated osteoclast
differentiation of BMMs. These findings contribute to the molecular
mechanism underlying IL-17A in alveolar bone destruction, and
provide insight into the clinical therapeutic targets for
periodontitis. However, these rudimentary studies cannot completely
elucidate the complex correlation between the immune system and
alveolar bone destruction. Future studies should be primarily
focused on the specific mechanism of IL-17A in periodontitis in
order to investigate the systemic treatment by blocking IL-17 for
the prevention of periodontitis.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81771072 and
81800972), and the Natural Science Foundation of Zhejiang Province
(grant nos. LY18H140002 and LY15H140003).
Availability of data and materials
The datasets used and analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
LS and JT performed the experiments, analyzed the
data and wrote the manuscript. ZW cultured the cells and analyzed
the data. MX performed western blotting and analyzed data. QT and
YW analyzed data and revised the manuscript. LC conceived and
designed the study. PD designed the study and revised the
manuscript. All authors read and approved the manuscript and agree
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All mice and rat experiments were performed in
accordance with the principles and procedures of the National
Institutes of Health (NIH) for the Care and Use of Laboratory
Animals. The approval number granted by the Animal Ethics Committee
of The Second Affiliated Hospital of Zhejiang University School of
Medicine is 2017–052. All surgeries were performed under sodium
pentobarbital anesthesia, and all efforts were made to minimize
suffering.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
IL-17A
|
interleukin 17A
|
|
BMMs
|
bone marrow macrophages
|
|
3-MA
|
3-methyladenine
|
|
TRAP
|
tartrate-resistant acid
phosphatase
|
|
NFATc1
|
nuclear factor of activated T cells
1
|
|
CatK
|
cathepsin K
|
|
RANKL
|
receptor activator of nuclear
factor-κB ligand
|
|
LC3
|
microtubule-associated protein 1 light
chain 3
|
|
M-CSF
|
macrophage colony-stimulating
factor
|
References
|
1
|
Page RC and Kornman KS: The pathogenesis
of human periodontitis: An introduction. Periodontol 2000. 14:9–11.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hajishengallis G: Immunomicrobial
pathogenesis of periodontitis: Keystones, pathobionts, and host
response. Trends Immunol. 35:3–11. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cochran DL: Inflammation and bone loss in
periodontal disease. J Periodontol 79 (8 Suppl). S1569–S1576. 2008.
View Article : Google Scholar
|
|
4
|
Cheng WC, Hughes FJ and Taams LS: The
presence, function and regulation of IL-17 and Th17 cells in
periodontitis. J Clin Periodontol. 41:541–549. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen XT, Chen LL, Tan JY, Shi DH, Ke T and
Lei LH: Th17 and Th1 lymphocytes are correlated with chronic
periodontitis. Immunol Invest. 45:243–254. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen XT, Tan JY, Lei LH and Chen LL:
Cytokine levels in plasma and gingival crevicular fluid in chronic
periodontitis. Am J Dent. 28:9–12. 2015.PubMed/NCBI
|
|
7
|
Cardoso CR, Garlet GP, Crippa GE, Rosa AL,
Junior WM, Rossi MA and Silva JS: Evidence of the presence of T
helper type 17 cells in chronic lesions of human periodontal
disease. Oral Microbiol Immunol. 24:1–6. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
da Costa TA, Silva MJ, Alves PM, Chica JE,
Barcelos EZ, Giani MA, Garlet GP, Silva JS, Junior VR, Rodrigues DB
and Cardoso CR: Inflammation biomarkers of advanced disease in
nongingival tissues of chronic periodontitis patients. Mediators
Inflamm. 2015:9837822015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hienz SA, Paliwal S and Ivanovski S:
Mechanisms of bone resorption in periodontitis. J Immunol Res.
2015:6154862015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fossiez F, Djossou O, Chomarat P,
Flores-Romo L, Ait-Yahia S, Maat C, Pin JJ, Garrone P, Garcia E,
Saeland S, et al: T cell interleukin-17 induces stromal cells to
produce proinflammatory and hematopoietic cytokines. J Exp Med.
183:2593–2603. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jovanovic DV, Di Battista JA,
Martel-Pelletier J, Jolicoeur FC, He Y, Zhang M, Mineau F and
Pelletier JP: IL-17 stimulates the production and expression of
proinflammatory cytokines, IL-beta and TNF-alpha, by human
macrophages. J Immunol. 160:3513–3521. 1998.PubMed/NCBI
|
|
12
|
Lubberts E, van den Bersselaar L,
Oppers-Walgreen B, Schwarzenberger P, Coenen-de Roo CJ, Kolls JK,
Joosten LA and van den Berg WB: IL-17 promotes bone erosion in
murine collagen-induced arthritis through loss of the receptor
activator of NF-kappa B ligand/osteoprotegerin balance. J Immunol.
170:2655–2662. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Feng Y, He D, Yao Z and Klionsky DJ: The
machinery of macroautophagy. Cell Res. 24:24–41. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hale AN, Ledbetter DJ, Gawriluk TR and
Rucker EB III: Autophagy: Regulation and role in development.
Autophagy. 9:951–972. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pierrefite-Carle V, Santucci-Darmanin S,
Breuil V, Camuzard O and Carle GF: Autophagy in bone: Self-eating
to stay in balance. Ageing Res Rev. 24:206–217. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gelman A and Elazar Z: Autophagic factors
cut to the bone. Dev Cell. 21:808–810. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
DeSelm CJ, Miller BC, Zou W, Beatty WL,
van Meel E, Takahata Y, Klumperman J, Tooze SA, Teitelbaum SL and
Virgin HW: Autophagy proteins regulate the secretory component of
osteoclastic bone resorption. Dev Cell. 21:966–974. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chung YH, Choi B, Song DH, Song Y, Kang
SW, Yoon SY, Kim SW, Lee HK and Chang EJ: Interleukin-1β promotes
the LC3-mediated secretory function of osteoclast precursors by
stimulating the Ca2+-dependent activation of ERK. Int J Biochem
Cell Biol. 54:198–207. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lin NY, Stefanica A and Distler JH:
Autophagy: A key pathway of TNF-induced inflammatory bone loss.
Autophagy. 9:1253–1255. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xing L and Boyce BF: RANKL-based
osteoclastogenic assays from murine bone marrow cells. Methods Mol
Biol. 1130:307–313. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang Y, Xiong Y, Chen X, Chen C, Zhu Z
and Li L: Therapeutic effect of bone marrow mesenchymal stem cells
pretreated with acetylsalicylic acid on experimental periodontitis
in rats. Int Immunopharmacol. 54:320–328. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Castro ΜL, Franco GC, Branco-de-Almeida
LS, Anbinder AL, Cogo-Muller K, Cortelli SC, Duarte S, Saxena D and
Rosalen PL: Downregulation of proteinase-activated receptor-2,
interleukin-17, and other proinflammatory genes by subantimicrobial
doxycycline dose in a rat periodontitis model. J Periodontol.
87:203–210. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Boyle WJ, Simonet WS and Lacey DL:
Osteoclast differentiation and activation. Nature. 423:337–342.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Teitelbaum SL and Ross FP: Genetic
regulation of osteoclast development and function. Nat Rev Genet.
4:638–649. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Asagiri M and Takayanagi H: The molecular
understanding of osteoclast differentiation. Bone. 40:251–264.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cekici A, Kantarci A, Hasturk H and Van
Dyke TE: Inflammatory and immune pathways in the pathogenesis of
periodontal disease. Periodontol 2000. 64:57–80. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tompkins KA: The osteoimmunology of
alveolar bone loss. Connect Tissue Res. 57:69–90. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sprangers S, Schoenmaker T, Cao Y, Everts
V and de Vries TJ: Different blood-borne human osteoclast
precursors respond in distinct ways to IL-17A. J Cell Physiol.
231:1249–1260. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Adamopoulos IE, Chao CC, Geissler R,
Laface D, Blumenschein W, lwakura Y, McClanahan T and Bowman EP:
Interleukin-17A upregulates receptor activator of NF-kappaB on
osteoclast precursors. Arthritis Res Ther. 12:R292010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Boyce BF: Advances in the regulation of
osteoclasts and osteoclast functions. J Dent Res. 92:860–867. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kitami S, Tanaka H, Kawato T, Tanabe N,
Katono-Tani T, Zhang F, Suzuki N, Yonehara Y and Maeno M: IL-17A
suppresses the expression of bone resorption-related proteinases
and osteoclast differentiation via IL-17RA or IL-17RC receptors in
RAW264.7 cells. Biochimie. 92:398–404. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wijekoon S, Bwalya EC, Fang J, Kim S,
Hosoya K and Okumura M: Chronological differential effects of
pro-inflammatory cytokines on RANKL-induced osteoclast
differentiation of canine bone marrow-derived macrophages. J Vet
Med Sci. 79:2030–2035. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Grigoriadis AE, Wang ZQ, Cecchini MG,
Hofestetter W, Felix R, Fleisch HA and Wanger EF: c-Fos: A key
regulator of osteoclast-macrophage lineage determination and bone
remodeling. Science. 266:443–448. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Asagiri M, Sato K, Usami T, Ochi S,
Nishina H, Yoshida H, Morita I, Wagner EF, Mark TW, Serfling E and
Takayanagi H: Autoamplification of NFATc1 expression determines its
essential role in bone homeostasis. J Exp Med. 202:1261–1269. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Costa AG, Cusano NE, Silva BC, Cremers S
and Bilezikian JP: Cathepsin K: Its skeletal actions and role as a
therapeutic target in osteoporosis. Nat Rev Rheumatol. 7:447–456.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hayman AR: Tartrate-resistant acid
phosphatase (TRAP) and the osteoclast/immune cell dichotomy.
Autoimmunity. 41:218–223. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang Z and Klionsky DJ: Eaten alive: A
history of macroautophagy. Nat Cell Biol. 12:814–822. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lin NY, Beyer C, Giessl A, Kireva T,
Scholtysek C, Uderhardt S, Munoz LE, Dees C, Distler A, Wirtz S, et
al: Autophagy regulates TNFα-mediated joint destruction in
experimental arthritis. Ann Rheum Dis. 72:761–768. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ke D, Fu X, Xue Y, Wu H, Zhang Y, Chen X
and Hou J: IL-17A regulates the autophagic activity of osteoclast
precursors through RANKL-JNK1 signaling during osteoclastogenesis
in vitro. Biochem Biophys Res Commun. 497:890–896. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xue Y, Liang Z, Fu X, Wang T, Xie Q and Ke
D: IL-17A modulates osteoclast precursors' apoptosis through
autophagy-TRAF3 signaling during osteoclastogenesis. Biochem
Biophys Res Commun. 508:1088–1092. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ng AY, Tu C, Shen S, Xu D, Oursler MJ, Qu
J and Yang S: Comparative characterization of osteoclasts derived
from murine bone marrow macrophages and RAW 264.7 cells using
quantitative proteomics. JBMR Plus. 2:328–340. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chung YH, Jang Y, Choi B, Song DH, Lee EJ,
Kim SM, Song Y, Kang SW, Yoon SY and Chang EJ: Beclin-1 is required
for RANKL-induced osteoclast differentiation. J Cell Physiol.
229:1963–1971. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li RF, Chen G, Ren JG, Zhang W, Wu ZX, Liu
B, Zhao Y and Zhao YF: The adaptor protein p62 is involved in
RANKL-induced autophagy and osteoclastogenesis. J Histochem
Cytochem. 62:879–888. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lin NY, Chen CW, Kagwiria R, Liang R,
Beyer C, Distler A, Luther J, Engelke K, Schett G and Distler JH:
Inactivation of autophagy ameliorates glucocorticoid-induced and
ovariectomy-induced bone loss. Ann Rheum Dis. 75:1203–1210. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Adamopoulos IE, Suzuki E, Chao CC, Gorman
D, Adda S, Maverakis E, Zarbalis K, Geissler R, Asio A,
Blumenschein WM, et al: IL-17A gene transfer induces bone loss and
epidermal hyperplasia associated with psoriatic arthritis. Ann
Rheum Dis. 74:1284–1292. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Park MJ, Park HS, Oh HJ, Lim JY, Yoon BY,
Kim HY, Cho ΜL and Cho SG: IL-17-deficient allogeneic bone marrow
transplantation prevents the induction of collagen-induced
arthritis in DBA/1J mice. Exp Mol Med. 44:694–705. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Koenders MI, Lubberts E, Oppers-Walgreen
B, van den Bersselaar L, Helsen MM, Di Padova FE, Boots AM, Gram H,
Joosten LA and van den Berg WB: Blocking of interleukin-17 during
reactivation of experimental arthritis prevents joint inflammation
and bone erosion by decreasing RANKL and interleukin-1. Am J
Pathol. 167:141–149. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Nakae S, Nambu A, Sudo K and Iwakura Y:
Suppression of immune induction of collagen-induced arthritis in
IL-17-deficient mice. J Immunol. 171:6173–6177. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Araujo VM, Melo IM and Lima V:
Relationship between periodontitis and rheumatoid arthritis: Review
of the Literature. Mediators Inflamm. 2015:2590742015. View Article : Google Scholar : PubMed/NCBI
|