Introduction
Liver cancer is the sixth most common cancer and the
third most common cause of cancer-associated mortality worldwide
(1,2). Liver resection remains the best
therapeutic strategy to treat liver cancer, with a 5-year survival
rate of ~30%. Chronic infection with hepatitis B or hepatitis C
virus is the most common cause of liver cancer (3); however, the mechanism underlying the
development of the disease remain unclear. Advancements in
molecular technology have led to a focus on the molecular mechanism
of liver cancer progression, and the identification of potential
clinically relevant factors which may benefit the diagnosis and
treatment of liver cancer.
Long non-coding RNAs (lncRNAs) are long-chain RNA
molecules of >200 nucleotides in length that do not encode for
proteins; thus, they have been proposed to be ineffective (4). Numerous studies have reported that
lncRNAs are closely related to the multi-level regulation of gene
expression, particularly the post-transcriptional regulation of the
interaction with microRNAs (miRNAs), mRNAs, or proteins (5–8).
Recent evidence suggests that lncRNAs are involved in several
pathological processes driving cancer, including proliferation,
apoptosis, metastasis and metabolism (9–13).
In particular, certain lncRNAs have been reported to be critical in
the progression of liver cancer. LncRNA p53 upregulated regulator
of P53 levels (lncRNA-PURPL), an intergenic lncRNA, has been
identified by RNA sequencing in a variety of colorectal cancer cell
lines. It has been observed to promote the occurrence of colorectal
cancer by preventing the binding of p53 to the small
p53-stabilizing protein Myb-binding protein 1A (MYBBP1A), lowering
p53 levels (14). However, to the
best of our knowledge, no investigations into the role and
expression of PURPL in liver cancer have been conducted.
The present study reported that PURPL is
significantly upregulated in liver cancer tissues and cell lines.
We also demonstrated that PURPL could promote liver cancer cell
proliferation and cell cycle progression, while inhibiting
apoptosis via regulation of the p53 gene. These findings may
provide novel insight into the role of PURPL in the progression of
liver cancer.
Materials and methods
Tissue collection and cell
culture
Primary tumor tissue and corresponding adjacent
tissue were excised from 71 patients (60 males and 11 female; age
range: 30–80-years-old) at The First Affiliated Hospital of China
Medical University between December 2016 and May 2018. The isolated
tissue specimens were placed in a cryotube within 30 min and
immediately frozen in a −80°C freezer. All cancerous tissues were
pathologically diagnosed as liver cancer. Patients did not receive
preoperative radiotherapy or chemotherapy. All individuals provided
informed consent and the present study was approved by the
Institutional Ethics Committee of China Medical University.
Patients were divided into high- and low-PURPL expression groups
based on the median value of expression.
A total of five liver cancer cell lines, HepG2,
Huh7, HCCLM3, SK-hep1, and PLC/PRF/5, as well as the normal liver
cell line, L02, were used in this study. HepG2 cells were obtained
from the Chinese Academy of Medical Sciences (Beijing, China).
Huh7, SK-hep1, PLC/PRF/5, HCCLM3 and L02 human cell lines were
obtained from the Chinese Academy of Sciences (Shanghai, China).
SK-hep1 and L02 cells were cultured in RPMI 1640 medium (HyClone;
GE Healthcare, Logan, UT, USA). HepG2, Huh7, PLC/PRF/5 and HCCLM3
cells were cultured in Dulbecco's Modified Eagle's medium (Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA). Media were
supplemented with 10% fetal bovine serum and 100 U/ml penicillin
and 100 U/ml streptomycin (HyClone; GE Healthcare). All cells were
grown at 37°C in a humidified incubator containing 5%
CO2.
Small interfering RNA (siRNA) and
transfection
A total of three siRNAs for the silencing of PURPL
and a negative control siRNA (si-NC) were designed and synthesized
by Shanghai GenePharma Co., Ltd. (Suzhou, China). Cell lines
cultured in six-well plates were transfected with 20 µM siRNAs
dissolved in ddH2O using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) and mixed with
siRNA-PURPL or si-NC according to the manufacturer's protocols.
After 48 h, cells were collected for reverse
transcription-quantitative polymerase chain reaction (RT-qPCR),
western blotting and other experiments. The sequences of the siRNAs
were as follows: 1#PURPLsiRNA,
5′-CCACAUACAGGGUUCUUAATTUUAAGAACCCUGUAUGUGGTT-3′; 2#PURPLsiRNA,
5′-GGAAUCGAUCUGUGAGCAUTTAUGCUCACAGAUCGAUUCCTT-3′; 3#PURPLsiRNA,
5′-GGCCUACGUGAAUAAUAAUTTAUUAUUAUUCACGUAGGCCTT-3′; and si-NC sense,
5′-UUCUCCGAACGUGUCACG-3′ and si-NC antisense,
5′-ACGUGACACGUUCGGAGAATT-3′. Among them, 3#PURPLsiRNA with the
strongest silencing efficiency, was used in subsequent
experiments.
RNA extraction and RT-qPCR
Total RNA was extracted from clinical patient tissue
samples and cultured cells using Total RNA Extractor (Sangon
Biotech Co., Ltd., Shanghai, China) according to the manufacturer's
instructions. Total RNA was reverse transcribed into cDNA using the
GoScript™ Reverse Transcription Mix and Random Primers (Promega
Corporation, Madison, WI, USA) according to the the manufacturer's
instructions. The cDNA was then amplified by qPCR using the GoTaqR
qPCR Master Mix (Promega Corporation) according to the
manufacturer's instructions on an ABI PRISMR 7500 (Applied
Biosystems; Thermo Fisher Scientific, Inc.) instrument using a
two-step standard PCR amplification procedure. The first step
(pre-denaturation) included a cycle of 2 min at 95°C; the second
step (PCR reaction) included 40 cycles of 15 sec at 95°C and 1 min
at 60°C. Finally, the relative expression of PURPL and p53 mRNA in
tissue and cell samples was determined by the 2−ΔΔCq
method (15). GAPDH was used as a
control. The nucleotide sequences of the primers for qPCR were as
follows: PURPL forward, 5′-TTCTACCGCAATTCGATGGAGTCTTG-3′, reverse,
5′-GAGGCAGGAGAATGGCGTGAAC-3′; p53 forward,
5′-ACCGGCGCACAGAGGAAGAG-3′, reverse,
5′-GCCTCATTCAGCTCTCGGAACATC-3′; and GAPDH forward,
5′-CGGAGTTGTTCGTATTCGG-3′ and reverse,
5′-TACTAGCCGATGATGGCATT-3′.
Western blotting
Transfected cells and frozen liver tissues,
including liver cancer and corresponding adjacent tissues, were
subjected to western blotting to evaluate p53 protein content.
Cells and tissue samples were lysed for 30 min in
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology, Shanghai, China) on ice, the resulting lysates were
centrifuged at 10,000 × g for 15 min at 4°C, and the protein
concentration was measured using a Bicinchoninic Acid kit (FDbio
Science, Hangzhou, China). Protein samples (30 µg) were then
separated by conventional electrophoresis at 70 V, transferred
under constant pressure, blocked in 5% skim milk, and finally
incubated with anti-p53 (cat. no. 60283-l-lg, 1:1,000) or
anti-GAPDH (cat. no. 60004-l-lg, 1 mg/ml), ProteinTech Group,
Chicago, IL, USA) primary antibodies at 4°C overnight. The
following day, membranes were washed with PBS, incubated with a
secondary antibody (Goat anti-mouse IgG, horseradish peroxidase,
cat. no. ZB-2305, 1:10,000, ZSGB-BIO) for 1 h at room temperature,
and proteins were visualized by chemiluminescence using an ECL kit
and an ECL western blotting substrate (Pierce; Thermo Fisher
Scientific, Inc.). Densitometry analysis was performed using ImageJ
software version 1.8.0 (National Institutes of Health, Bethesda,
MD, USA).
Cell proliferation assays
Transfected HepG2 and Huh7 cells were seeded in
96-well microtiter plates at a density of 5×103
cells/well. At 24, 48, 72 and 96 h post-transfection, 10 µl Cell
Counting Kit-8 (CCK-8) solution (Beyotime Institute of
Biotechnology) was added to each well and the plates were incubated
in a 37°C incubator for 1.5 h. Then, the absorbance per well was
measured at 450 nm using an automatic microplate reader (BGI, Hong
Kong, China).
For the colony-forming assay, each group of cells
was seeded on a six-well plate at 1×103 cells/well and
incubated for 10 days at 37°C in 5% CO2. Then, the cells
were washed with PBS, fixed with paraformaldehyde for 15 min then
stained with 0.5% crystal violet at room temperature and colonies
were then counted.
Cell cycle analysis
To investigate the effects of PURPL silencing on
cell cycle progression, we analyzed the proportion of cells in G1,
S, and G2 phase using a BD FACSCanto™ flow cytometer (BD
Biosciences, Franklin Lakes, NJ, USA). We routinely detached cells
with trypsin (0.25%) for 5 min at 37°C, centrifuged at 850 × g at
4°C for 5 min, removed the supernatant, resuspended the cells in
pre-cooled PBS, and centrifuged them again. The supernatant was
aspirated, and the cells were fixed at 4°C by adding 1 ml of
pre-cooled 70% ethanol and subsequent incubation for 12 h. The next
day, following centrifugation (850 × g, 4°C, 5 min) and
washing(PBS), 0.5 ml of propidium iodide (PI) staining solution
(Beyotime Institute of Biotechnology) was added to each group of
cell samples, which were incubated at 37°C for 30 min in the dark.
Samples were then stored on ice until they were detected on the
flow cytometer (BD FACSCanto™).
Cell apoptosis analysis
In accordance with the manufacturer's protocols of
an apoptosis kit (Dojindo Molecular Technologies, Inc., Kumamoto,
Japan), cells were trypsinized, collected (~5×105
cells/group), washed twice with PBS, centrifuged at 850 × g at 4°C
for 5 min, and then 100 µl of Annexin V buffer was added. Then, 5
µl of PI and 5 µl of Annexin V-FITC solution were added to each
tube. The tubes were mixed by pipetting and incubated at room
temperature for 15 min, after which 400 µl of Annexin V buffer
(included in kit) was added to each tube and mixed by pipetting.
Readings were obtained on a flow cytometer (BD FACSCanto™) as
aforementioned.
Statistical analysis
The results were analyzed using GraphPad Prism 5.0
software (GraphPad Software, Inc., La Jolla, CA, USA). Data are
expressed as the mean ± standard deviation. Differences between two
groups were assessed using a Student's t-test, whereas differences
among multiple groups were analyzed using one-way analysis of
variance followed by the Least Significant Difference test. The
relationship between PURPL and p53 expression was analyzed by
Pearson correlation analysis. The association between PURPL
expression and the clinicopathological parameters of patients was
assessed via a c2 test. All experiments were performed
in triplicate. P<0.05 was considered to indicate a statistically
significant difference.
Results
LncRNA PURPL is upregulated in liver
cancer tissues and cell lines
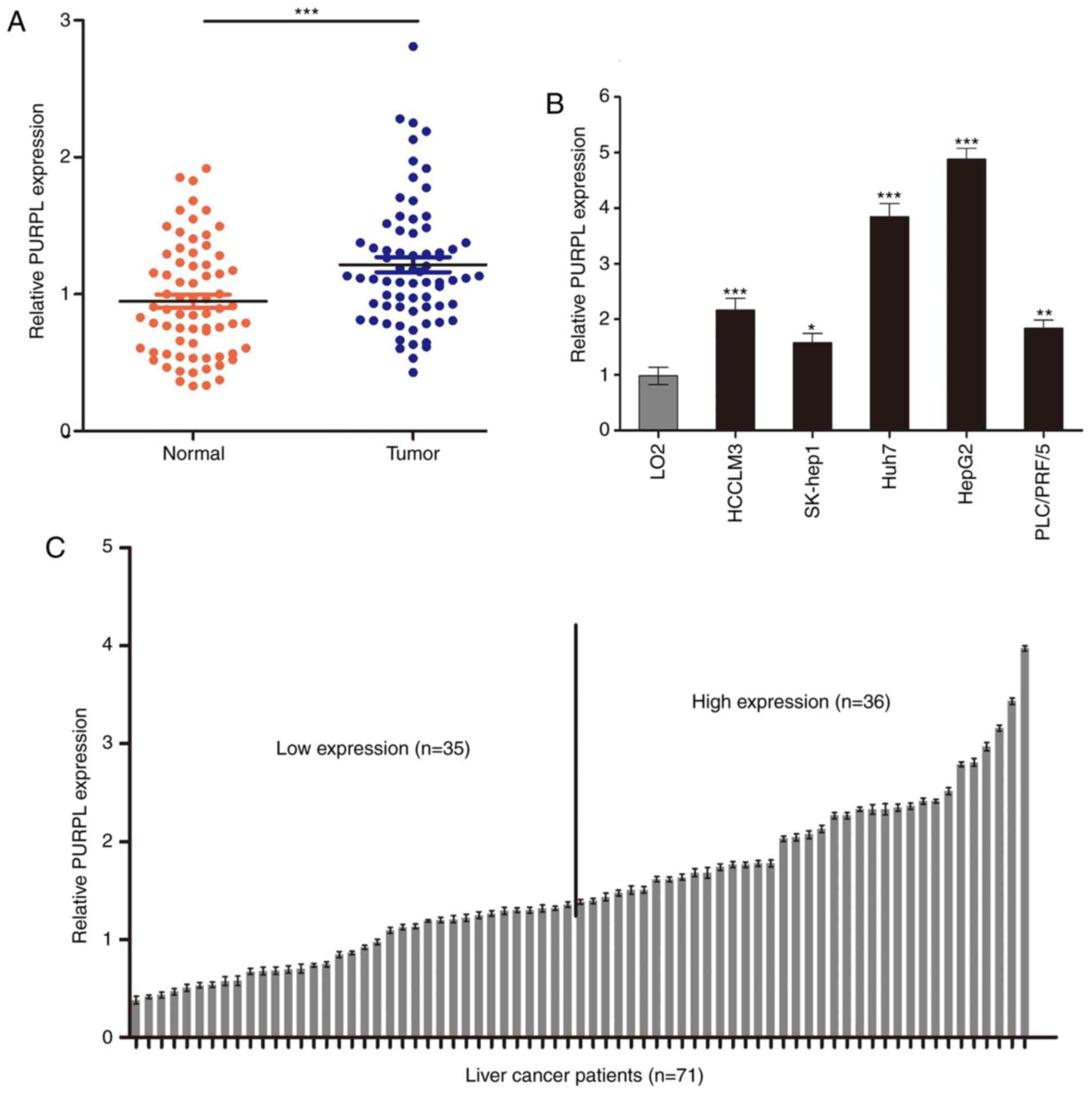
Using RT-qPCR, we examined the expression of PURPL
in 71 patients with liver cancer and several liver cancer cell
lines. The expression of PURPL was significantly increased in
cancer tissues than in adjacent tissues (P<0.001; Fig. 1A). In addition, PURPL was highly
expressed in HepG2 and Huh7 cells compared with the three other
liver cancer cell lines and L02 cells (P<0.05; Fig. 1B). To investigate the association
of PURPL expression with clinicopathological factors of patients,
the patients were divided into high- (n=36) and low-expression
(n=35) groups based on PURPL median expression levels (Fig. 1C). As presented in Table I, PURPL expression levels in
patients with liver cancer were closely associated with tumor size
(P<0.05) and tumor differentiation (P<0.01). These data
indicated that PURPL is significantly elevated in liver cancer
patients and may be closely related to the progression of liver
cancer.
 | Table I.Association between lncRNA-PURPL
expression according to reverse transcription-quantitative
polymerase chain reaction and conventional clinicopathological
parameters in 71 patients with liver cancer. |
Table I.
Association between lncRNA-PURPL
expression according to reverse transcription-quantitative
polymerase chain reaction and conventional clinicopathological
parameters in 71 patients with liver cancer.
| Characteristics | Number of
patients | lncRNA-PURPL Low
expression (%) | lncRNA-PURPL High
expression (%) | P-value |
|---|
| Total cases | 71 | 35 | 36 |
|
| Age (years) |
|
|
| 0.111 |
| ≥55 | 29 | 11 (37.93) | 18 (62.07) |
|
|
<55 | 42 | 24 (57.14) | 18 (42.86) |
|
| Sex |
|
|
| 0.705 |
| Male | 60 | 29 (48.33) | 31 (51.67) |
|
|
Female | 11 | 6 (54.56) | 5 (45.44) |
|
| Tumor size |
|
|
| 0.013a |
| ≥5cm | 39 | 14 (35.90) | 25 (64.10) |
|
|
<5cm | 32 | 21 (65.63) | 11 (34.37) |
|
| HBsAg status |
|
|
| 0.591 |
|
Positive | 57 | 29 (50.88) | 28 (49.12) |
|
|
Negative | 14 | 6 (42.86) | 8 (57.14) |
|
| Tumor
differentiation |
|
|
| 0.002b |
|
High | 21 | 17 (80.95) | 4 (19.05) |
|
|
Moderate | 30 | 12 (40.00) | 18 (60.00) |
|
|
Poor | 20 | 6 (30.00) | 14 (70.00) |
|
| PVTT |
|
|
| 0.097 |
|
Yes | 10 | 2 (20.00) | 8 (80.00) |
|
| No | 61 | 33 (54.10) | 28 (45.90) |
|
| Serum AFP |
|
|
| 0.288 |
| <20
ng/ml | 30 | 17 (56.67) | 13 (43.33) |
|
| ≥20
ng/ml | 41 | 18 (43.90) | 23 (56.10) |
|
| Liver
cirrhosis |
|
|
| 0.350 |
|
Yes | 56 | 26 (46.43) | 30 (53.57) |
|
| No | 15 | 9 (60.00) | 6 (40.00) |
|
| Metastasis |
|
|
| 0.999 |
|
Yes | 7 | 3 (42.86) | 4 (57.14) |
|
| No | 64 | 32 (50.00) | 32 (50.00) |
|
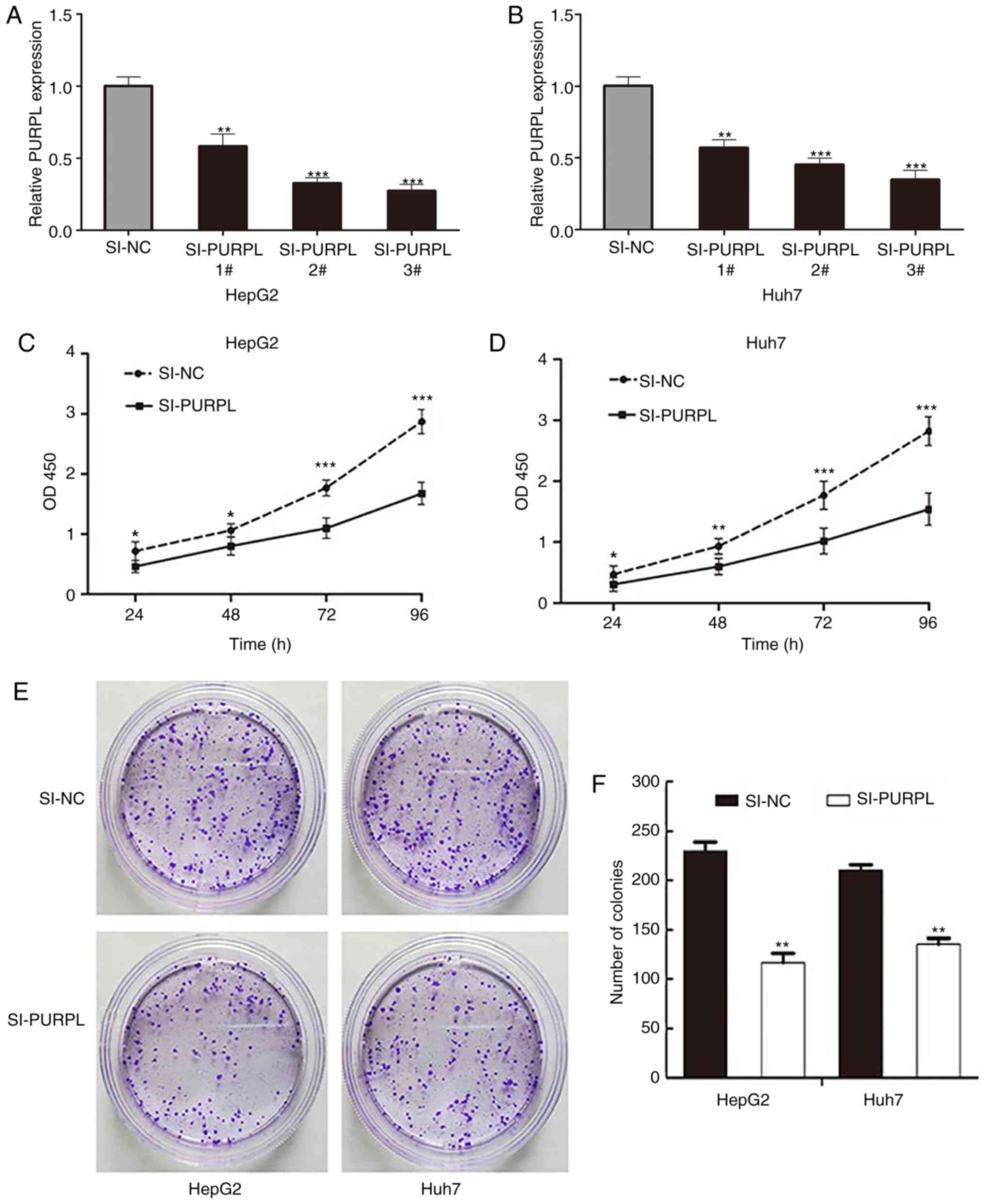
Depletion of lncRNA PURPL suppresses
liver cancer cell proliferation
To determine whether PURPL regulates the
proliferative capacity of liver cancer cells, we used three
different siRNAs to silence the PURPL gene in liver cancer cells.
#3PURPLsiRNA exhibited the strongest silencing efficiency
(P<0.05; Fig. 2A and B); thus,
#3PURPLsiRNA was selected for subsequent experiments. To determine
the effect of PURPL on proliferation of liver cancer cells, we
performed a CCK-8 assay. Silencing of PURPL significantly inhibited
the proliferation of HepG2 and Huh7 cells compared with the control
(P<0.05; Fig. 2C and D). In
addition, the number of clones formed by PURPL-silenced cells was
significantly decreased compared with the corresponding controls
(P<0.01; Fig. 2E and F).
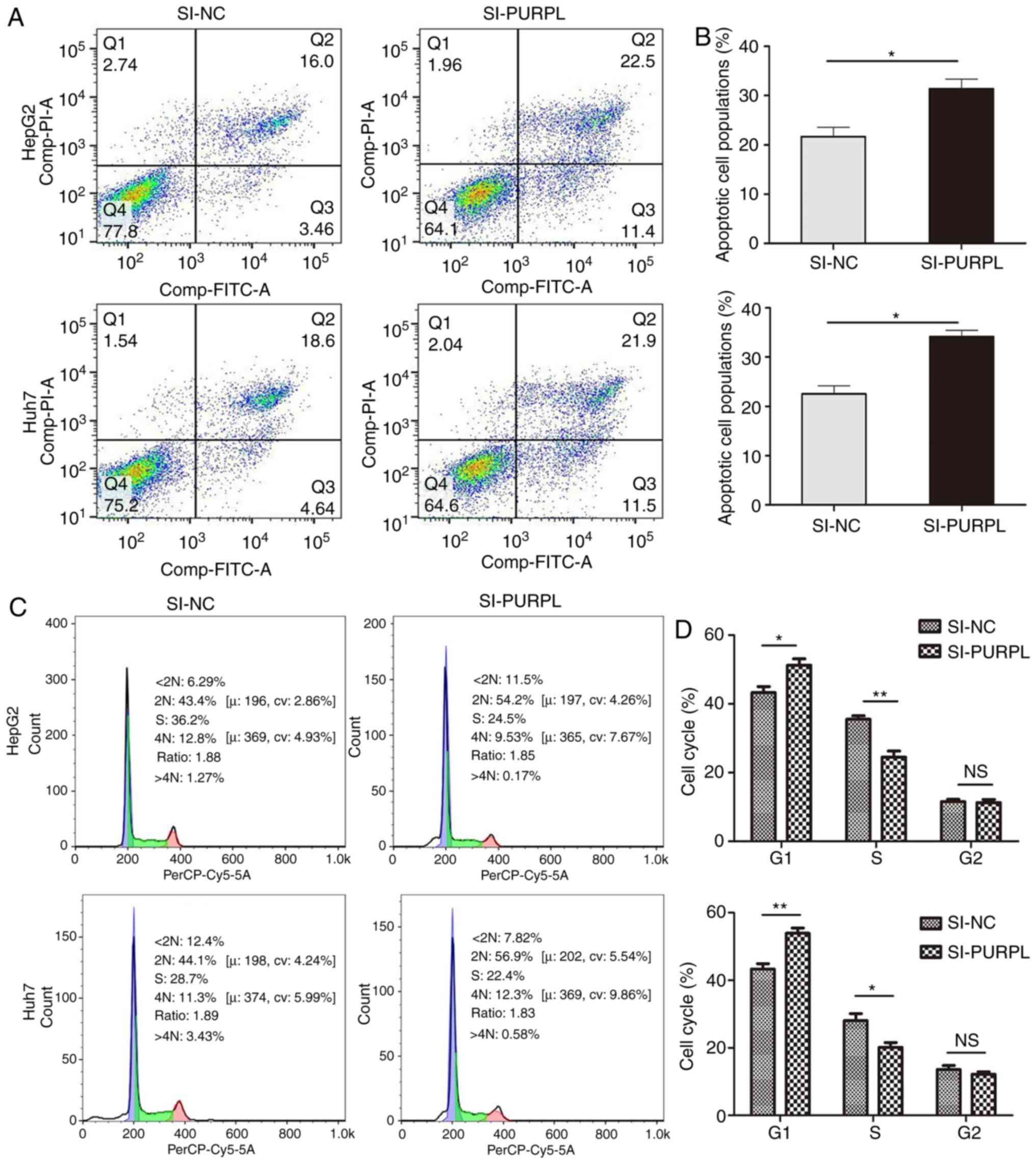
Depletion of lncRNA PURPL promotes
liver cancer cell apoptosis and leads to liver cancer cell cycle
arrest
Cell apoptosis was monitored by flow cytometry. The
number of apoptotic cells was significantly increased following
transfection with siRNA-PURPL than those transfected with si-NC
(P<0.01; Fig. 3A and B).
Regarding cell cycle progression, PURPL knockdown induced G1 phase
arrest of liver cancer cells; siRNA-PURPL transfection resulted in
a significant decrease in the number of cells in S phase compared
with the control (P<0.05; Fig. 3C
and D). Collectively, these results suggested that PURPL
knockdown may induce G1 phase arrest and promote liver cancer cell
apoptosis.
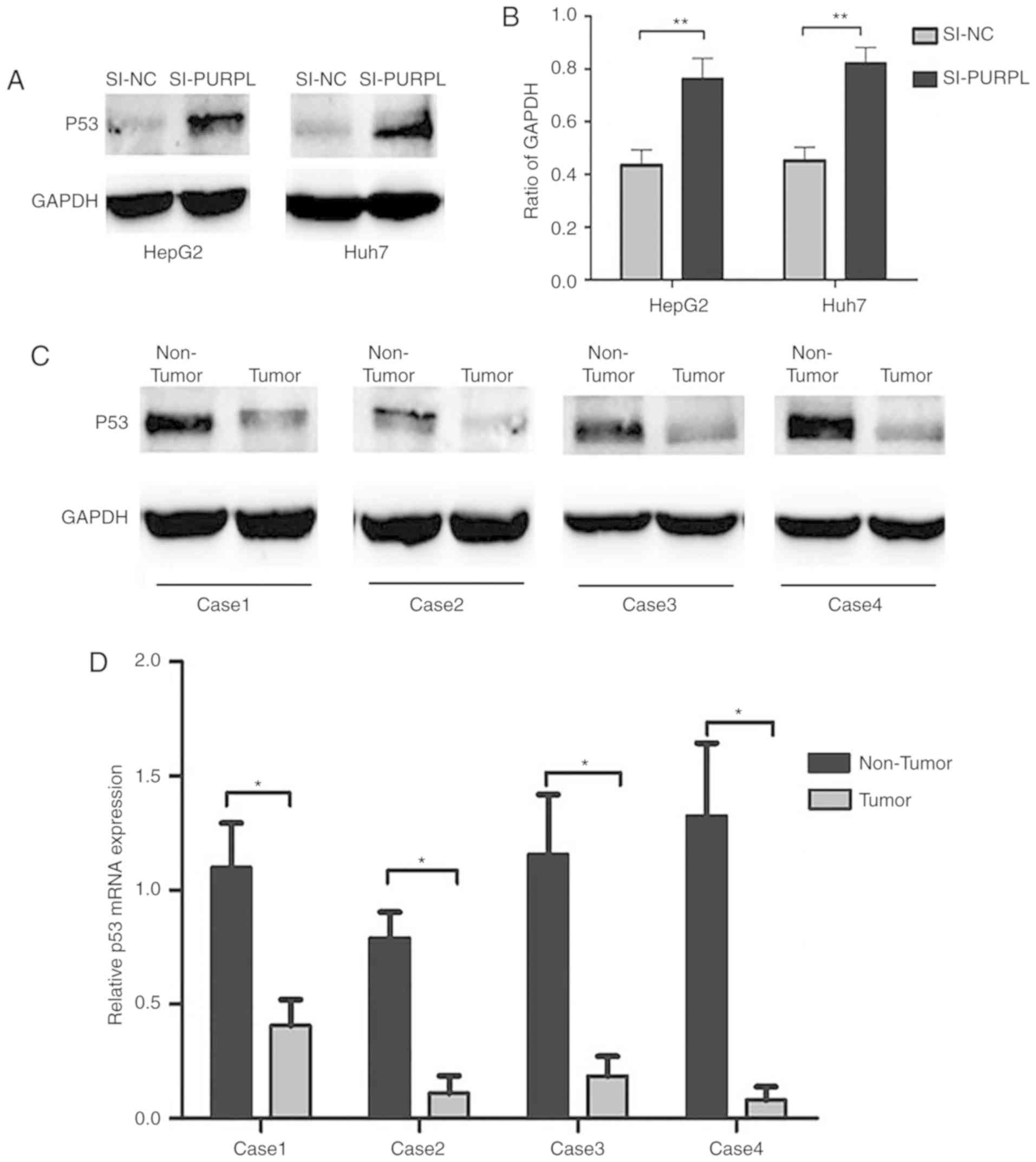
LncRNA PURPL inhibits p53 expression
in liver cancer
p53 can be modulated by lncRNAs to regulate cancer
cell progression and PURPL has been reported to inactivate p53
protein in colorectal cancer (14). To improve understanding of the role
of PURPL in liver cancer, we used western blotting to measure the
expression levels of p53 in liver cancer cells. The results
revealed that p53 was significantly upregulated in HepG2 and Huh7
cells transfected with siRNA-PURPL compared with si-NC (P<0.01;
Fig. 4A and B). Western blotting
and RT-qPCR of liver cancer tissues demonstrated downregulation of
p53 in tumor tissue compared with paracancerous tissues in four
randomly selected late-stage liver cancer patients, at the protein
and mRNA levels (P<0.05; Fig. 4C
and D). Additionally, relative p53 mRNA expression was
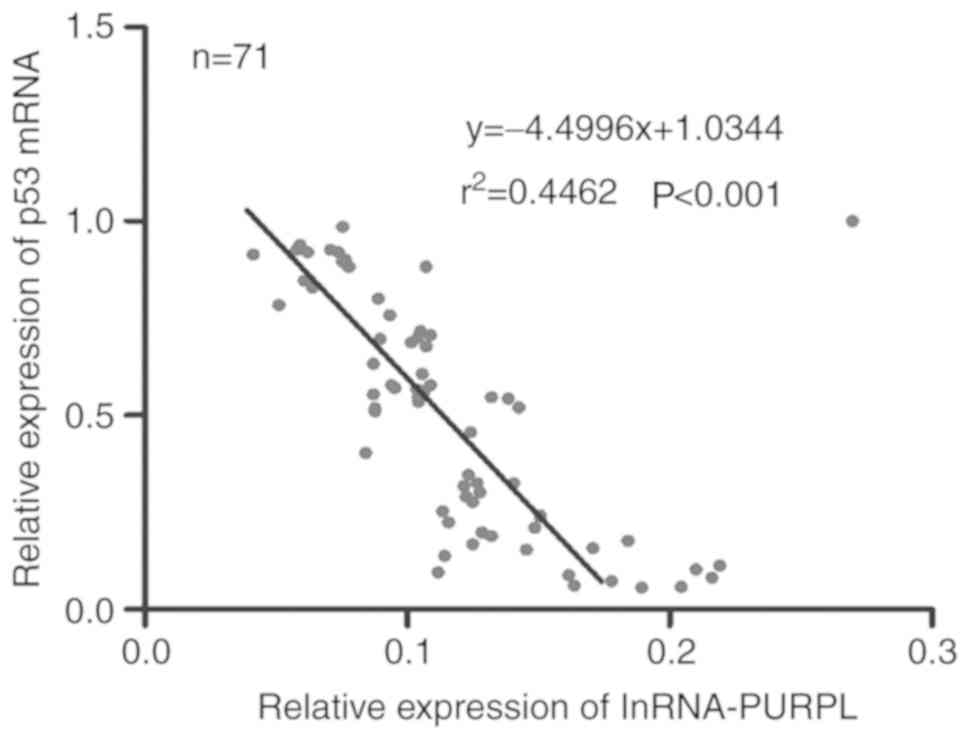
negatively correlated with PURPL expression in all 71 patients
(P<0.001; Fig. 5). These
results indicate the role of PURPL as an oncogene that may be
partially responsible for inducing p53 in liver cancer.
Discussion
Aggressive tumor invasion, metastatic dissemination,
recurrence, and drug resistance are characteristic of patients with
liver cancer (16–19). Thus, research on liver cancer has
focused on identifying genes that induce hepatocarcinogenesis and
promote the development of liver cancer, for which targeted drugs
and molecular markers capable of early diagnosis could be
developed. Increasing evidence suggests that aberrant expression of
lncRNA serves a key role in the development and progression of
liver cancer. For example, lncRNA-CCAL promotes liver cancer
progression by modulating AP-2α and Wnt/β-catenin pathways,
lncRNA-HULC promotes liver cancer by increasing the expression of
the high mobility group AT-hook 2 oncogene and sequestration of
miRNA-186; lncRNA-TSLNC8 serves as a tumor suppressor that
inactivates the interleukin-6/signal transducer and activator or
transcription 3 signaling pathway in liver cancer (20–22).
LncRNA-PURPL is highly expressed in colorectal
cancer and serves an important role in carcinogenesis. The present
study analyzed the expression of PURPL by RT-qPCR and revealed that
PURPL was highly expressed in liver cancer tissues and numerous
cell lines. Furthermore, the upregulated expression of PURPL was
positively associated with tumor size and tumor differentiation in
patients with liver cancer. In addition, silencing of PURPL
expression could inhibit liver cancer cell proliferation, arrest
cell cycle progression and promote cell apoptosis.
The p53 gene was first described in 1979 (23). It serves an important role in the
occurrence and development of various tumors via the control of DNA
damage repair, apoptosis and cell cycle regulation (23,24).
Several proteins, miRNAs and lncRNAs have been reported to affect
the occurrence and development of liver cancer by regulating the
p53 gene (25–29).
In recent years, numerous studies have investigated
on the role of lncRNAs in liver cancer and p53 gene regulation. For
example, Zhou et al (30)
reported that p53 regulation associated lncRNA inhibited liver
cancer growth and induced apoptosis in vivo and in
vitro via p53. Ren et al (31) reported that lncRNA
prostate-cancer-associated ncRNA transcript-1 expression could be
regulated by miR-215, a p53-inducible miRNA in liver cancer, and
this post-transcriptional regulation significantly affected a
variety of malignant phenomena of liver cancer cells. Additionally,
Zhang et al (32) revealed
that lncRNA-small nucleolar RNA host gene was a predictor of poor
prognosis and promoted liver cancer tumorigenesis by regulating
p53. In colorectal cancer, PURPL inhibits p53 gene expression by
blocking the interaction between p53 and MYBBP1A (14). Whether PURPL inhibits the p53 gene
via MYBBP1A or through other mechanisms of action requires further
investigation.
In conclusion, our data demonstrated that PURPL is
upregulated in liver cancer tissues, and is associated with tumor
size and tumor differentiation. Silencing of PURPL could inhibit
the proliferation of liver cancer cells, block cell cycle
progression and promote apoptosis. Furthermore, PURPL may affect
these malignant features of liver cancer cells by regulating p53.
In this regard, we elevated the expression of PURPL in liver cancer
cell lines, however this failed to alter the malignant
characteristics of liver cancer cells (data not shown). Our future
studies aim to investigate the specific mechanisms whereby PURPL
affects the development of liver cancer. Collectively, lncRNA-PURPL
was proposed to exert an important carcinogenic effect on liver
cancer, and may be a potential predictor and a novel therapeutic
target for the treatment of this disease.
Acknowledgements
Not applicable.
Funding
This study was funded by the Shenyang Municipal
Science and Technology Bureau Population and Health Research
Project (grant no. F15-139-9-25) and by the Natural Science
Foundation of Liaoning Province (grant no. 201520529).
Availability of data and materials
The datasets used or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GW made substantial contributions to the design of
the present study. XF conducted all the experiments and wrote the
manuscript. YW, WZ, SX and WW made substantial contributions to the
analysis of the data and discussed the results. All authors
approved the final version of the manuscript.
Ethics approval and consent to
participate
All patients provided written informed consent for
the use of tissues. The present study was approved by the
institutional ethics committee of China Medical University
(Shenyang, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Shi X, Zhu HR, Liu TT, Shen XZ and Zhu JM:
The Hippo pathway in hepatocellular carcinoma: Non-coding RNAs in
action. Cancer Lett. 400:175–182. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fujiwara N, Friedman SL, Goossens N and
Hoshida Y: Risk factors and prevention of hepatocellular carcinoma
in the era of precision medicine. J Hepatol. 68:526–549. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ma M, Cai B, Jiang L, Abdalla BA, Li Z,
Nie Q and Zhang X: lncRNA-Six1 Is a target of miR-1611 that
functions as a ceRNA to regulate six1 protein expression and fiber
type switching in chicken myogenesis. Cells. 7(pii): E2432018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Grelet S, Link LA, Howley B, Obellianne C,
Palanisamy V, Gangaraju VK, Diehl JA and Howe PH: A regulated PNUTS
mRNA to lncRNA splice switch mediates EMT and tumour progression.
Nat Cell Biol. 19:1105–1115. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ferrè F, Colantoni A and Helmer-Citterich
M: Revealing protein-lncRNA interaction. Brief Bioinform.
17:106–116. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhu Y, Bian Y, Zhang Q, Hu J, Li L, Yang
M, Qian H, Yu L, Liu B and Qian X: Construction and analysis of
dysregulated lncRNA-associated ceRNA network in colorectal cancer.
J Cell Biochem. Dec 7–2018.(Epub ahead of print).
|
|
9
|
Jiang N, Wang X, Xie X, Liao Y, Liu N, Liu
J, Miao N, Shen J and Peng T: lncRNA DANCR promotes tumor
progression and cancer stemness features in osteosarcoma by
upregulating AXL via miR-33a-5p inhibition. Cancer Lett. 405:46–55.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li W, Jia G, Qu Y, Du Q and Liu B and Liu
B: Long non-coding RNA (LncRNA) HOXA11-AS promotes breast cancer
invasion and metastasis by regulating epithelial-mesenchymal
transition. Med Sci Monit. 23:3393–3403. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hu B, Cai H, Zheng R, Yang S, Zhou Z and
Tu J: Long non-coding RNA 657 suppresses hepatocellular carcinoma
cell growth by acting as a molecular sponge of miR-106a-5p to
regulate PTEN expression. Int J Biochem Cell Biol. 92:34–42. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mineo M, Ricklefs F, Rooj AK, Lyons SM,
Ivanov P, Ansari KI, Nakano I, Chiocca EA, Godlewski J and Bronisz
A: The long non-coding RNA HIF1A-AS2 facilitates the maintenance of
mesenchymal glioblastoma stem-like cells in hypoxic niches. Cell
Rep. 15:2500–2509. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang HY, Zheng FS, Yang W and Lu JB: The
long non-coding RNA MIAT regulates zinc finger E-box binding
homeobox 1 expression by sponging miR-150 and promoteing cell
invasion in non-small-cell lung cancer. Gene. 633:61–65. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li XL, Subramanian M, Jones MF, Chaudhary
R, Singh DK, Zong X, Gryder B, Sindri S, Mo M, Schetter A, et al:
Long noncoding RNA PURPL suppresses basal p53 levels and promotes
tumorigenicity in colorectal cancer. Cell Rep. 20:2408–2423. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang T, Liu W, Meng W, Zhao H, Yang Q, Gu
SJ, Xiao CC, Jia CC and Fu BS: Downregulation of miR-542-3p
promotes cancer metastasis through activating TGF-β/Smad signaling
in hepatocellular carcinoma. Onco Targets Ther. 11:1929–1939. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ma Y, Yang Y, Wang F, Moyer MP, Wei Q,
Zhang P, Yang Z, Liu W, Zhang H, Chen N, et al: Long non-coding RNA
CCAL regulates colorectal cancer progression by activating
Wnt/β-catenin signalling pathway via suppression of activator
protein 2α. Gut. 65:1494–1504. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu G, Zheng K, Xia S, Wang Y, Meng X, Qin
X and Cheng Y: MicroRNA-655-3p functions as a tumor suppressor by
regulating ADAM10 and β-catenin pathway in Hepatocellular
Carcinoma. J Exp Clin Cancer Res. 35:892016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lohitesh K, Chowdhury R and Mukherjee S:
Resistance a major hindrance to chemotherapy in hepatocellular
carcinoma: An insight. Cancer Cell Int. 18:442018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Y, Chen F, Zhao M, Yang Z, Li J,
Zhang S, Zhang W, Ye L and Zhang X: The long noncoding RNA HULC
promotes liver cancer by increasing the expression of the HMGA2
oncogene via sequestration of the microRNA-186. J Biol Chem.
292:15395–15407. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang J: Long noncoding RNA TSLNC8 is a
tumor suppressor that inactivates the interleukin-6/STAT3 signaling
pathway. Hepatology. 67:171–187. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu Y, Yang Y, Wang T, Wang L, Wang X, Li
T, Shi Y and Wang Y: Long non-coding RNA CCAL promotes
hepatocellular carcinoma progression by regulating AP-2α and
Wnt/β-catenin pathway. Int J Biol Macromol. 109:424–434. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vogelstein B, Lane D and Levine AJ:
Surfing the p53 network. Nature. 408:307–310. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Vousden KH and Lu X: Live or let die: The
cell's response to p53. Nat Rev Cancer. 2:594–604. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhu H, Wang J, Yin J, Lu B, Yang Q, Wan Y
and Jia C: Downregulation of PRAME suppresses proliferation and
promotes apoptosis in hepatocellular carcinoma through the
activation of P53 mediated pathway. Cell Physiol Biochem.
45:1121–1135. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Meng X, Franklin DA, Dong J and Zhang Y:
MDM2-p53 pathway in hepatocellular carcinoma. Cancer Res.
74:7161–7167. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Su P, Wang F, Qi B, Wang T and Zhang S:
P53 Regulation-association long non-coding RNA (LncRNA PRAL)
inhibits cell proliferation by regulation of P53 in human lung
cancer. Med Sci Monit. 23:1751–1758. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pollutri D, Gramantieri L, Bolondi L and
Fornari F: TP53/MicroRNA interplay in hepatocellular carcinoma. Int
J Mol Sci. 17(pii): E20292016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu J, Rao J, Lou X, Zhai J, Ni Z and Wang
X: Upregulated TRIM11 exerts its oncogenic effects in
hepatocellular carcinoma through inhibition of P53. Cell Physiol
Biochem. 44:255–266. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhou CC, Yang F, Yuan SX, Ma JZ, Liu F,
Yuan JH, Bi FR, Lin KY, Yin JH, Cao GW, et al: Systemic genome
screening identifies the outcome associated focal loss of long
noncoding RNA PRAL in hepatocellular carcinoma. Hepatology.
63:850–863. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ren Y, Shang J, Li J, Liu W, Zhang Z, Yuan
J and Yang M: The long noncoding RNA PCAT-1 links the microRNA
miR-215 to oncogene CRKL-mediated signaling in hepatocellular
carcinoma. J Biol Chem. 292:17939–17949. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang M, Wang W, Li T, Yu X, Zhu Y, Ding
F, Li D and Yang T: Long noncoding RNA SNHG1 predicts a poor
prognosis and promotes hepatocellular carcinoma tumorigenesis.
Biomed Pharmacother. 80:73–79. 2016. View Article : Google Scholar : PubMed/NCBI
|