Introduction
The three principal muscle types in mammals are
cardiac, smooth and skeletal muscles, with skeletal muscle being
the most abundant tissues in the human body (1). Although previous studies identified
sex-specific differences in the physiopathological characteristics
of the cardiovascular system (2),
the sex-specific physiopathological properties of skeletal muscles
remain unclear. A previous study demonstrated that >3,000 genes
were differentially expressed between male and female skeletal
muscle (3).
In mammals, there are three principal types of
estrogens: Estrone, 17β-estradiol (E2) and estriol.
E2, which exhibits increased biological activity
compared with other estrogens, is secreted primarily by the growing
follicles in the ovaries during the ovarian cycle (4). Notably, skeletal muscles are targeted
by E2 (5). During
perimenopausal and postmenopausal periods, there is a significant
decrease in muscle strength that may be increased by hormonal
replacement therapy (HRT), suggesting that estrogens are important
modulators of muscle physiology and are able to affect gene
expression and mitochondrial function, thus maintaining the
bioenergetic status of muscle cells (6).
In addition to the regulation of reproductive
functions, estrogens may affect various physiological functions,
including cellular metabolism (7,8).
Decreased levels of E2 following menopause or
ovariectomy (OVX) are associated with hyperphagia, obesity, hepatic
steatosis and triglycerides (TG) accumulation in skeletal muscle
cells (9–11); however the molecular mechanisms
underlying E2 function remain unclear. Due to their
lipophilic properties, estrogens are able to diffuse through the
cellular membranes, interacting with intracellular receptors
(12). In total, there are two
types of estrogen receptors (ESR): ESR1 and ESR2.
Previous studies demonstrated that the effects of
E2 on energy homeostasis are primarily mediated by ESR1
(10,13). Activating ESR1 using
propylpyrazoletriol (PPT) may affect metabolism, insulin resistance
and mitochondrial function in OVX mice with metabolic syndrome
(14); however, the molecular
mechanism underlying ESR1 function remains unclear.
Skeletal muscles are able to oxidize non-esterified
fatty acids, or to store fatty acids (FAs) as TG, accumulating
intramuscular triacylglycerol (IMTAG) (15). Although physiological levels of
IMTAG do not impair the metabolic functions of skeletal muscles,
the accumulation of IMTAG may be associated with promoted insulin
resistance, due to increased levels of lipid intermediates and the
subsequent activation of inflammatory or stress-associated pathway
(16). Estrogen increases the
levels of the FA transporter CD36 molecule (CD36) in the plasma
membrane of cardiomyocytes, thus serving cardioprotective roles
(17). Therefore, in the present
study, it was hypothesized that E2 may increase
CD36-mediated FA uptake, inhibiting the synthesis of FA and TG in
skeletal muscle.
The transcription factor peroxisome proliferator
activated receptor α (PPARα) is expressed primarily in tissues
exhibiting high rates of FA oxidation, including cardiac and
skeletal muscles (18). PPARα may
regulate the protein expression levels of factors involved in FA
catabolism. PPARα activation was identified to improve the
musculoskeletal effects of exercise during estrogen deficiency
(19); however, its roles in
CD36-mediated FA uptake and the synthesis of acetyl-CoA carboxylase
α (ACACA), fatty acid synthetase (FASN) and perilipin 2 (PLIN2)
remain unclear.
Although the potential health risks and benefits of
long-term HRT remain unclear, future studies using short-term HRT
or specific ER agonists including PPT and diarylpropionitrile
(DPN), may provide novel insights into the effects of E2
on skeletal muscles, thus benefiting postmenopausal patients
(20). Therefore, it was
hypothesized that short-term HRT may inhibit TG synthesis in the
skeletal muscles of female rats following OVX, and this effect may
be associated with the role of FA in promoting the activity of
PPARα following the interaction between E2, and ESR1 or
2. To investigate this hypothesis, in vivo experiments were
performed to examine the effects of E2 on weight loss
and TG synthesis inhibition in female rats following OVX.
Additionally, in vitro experiments were performed using ER
agonists, including PPT and DPN, to activate ESR1 or ESR2, thus
promoting FA transport, inhibiting TG synthesis and improving
insulin resistance. Palmitic acid (PA) is the principal saturated
FA in the blood, and was selected to treat myotubes formed by
differentiated C2C12 cells in order to investigate the mechanisms
underlying E2-mediated TG synthesis and insulin
resistance.
Materials and methods
Animals
A total of 30 female Sprague-Dawley rats (age, 6
months; weight, 300–350 g) were purchased from The Experimental
Animal Center at Anhui Medical University (Hefei, China). All rats
had free access to food and water and were maintained in standard
conditions of controlled temperature and humidity (22±1°C, 60–70%)
under a 12-h light/dark cycle for one week. All animal experiments
were reviewed and approved by The Ethics Committee of Anhui Medical
University.
Experimental protocol
Rats were randomly divided into three groups: i)
Sham surgery (SHAM; n=10); ii) rats that underwent ovariectomy
without treatment (OVX; n=10); and iii) rats that underwent
ovariectomy and treated with E2 (OVX + E2;
n=10). All groups underwent laparotomy; however, the OVX and OVX +
E2 groups underwent bilateral ovariectomy. The OVX +
E2 rats were injected with E2 (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) at a concentration of 0.06
mg/kg/day for 3 months following surgery. Body weight and food
intake were measured every week.
Glucose tolerance test (GTT) and
insulin tolerance test (ITT)
Insulin sensitivity was assessed using GTT and ITT
as previously described (21).
Briefly, for the GTT assay, all rats were fasted overnight and
administered glucose (2 g/kg) by intraperitoneal injection. Blood
samples were obtained from the tail veins at 0, 30, 60, 90 and 120
min following injection. The blood glucose was measured with a
glucometer (OneTouch Ultra; Johnson & Johnson, New Brunswick,
NJ, USA). ITT assays were performed by the intraperitoneal
injection of neutral insulin (1 U/kg) following fasting for 4–6 h.
The blood glucose was measured at 0, 30, 45, 60, 90 and 120 min.
All data were plotted as blood glucose concentration over time.
Subsequently, the area under curve (AUC) was calculated.
Serum lipid levels and E2
assays
After 6 months, rats were fasted for 12 h and
anesthetized by intraperitoneal injection of sodium pentobarbital
(60 mg/kg). Blood (~10 ml) was collected in tubes and serum was
separated by centrifuging the samples at 1,000 × g for 10 min at
room temperature and stored at −80°C prior to biochemical analysis.
Fasting TG, total cholesterol (TC), low-density lipoprotein
cholesterol (LDL-C) and high-density lipoprotein cholesterol
(HDL-C) levels were measured using an automatic biochemical
analyzer (AU640; Olympus Corporation, Tokyo, Japan). The serum
levels of E2 were determined by radioimmunoassay
(Beijing North Institute of Biological Technology, Beijing, China)
and the serum levels of free FA (FFA) were measured using a
Non-esterified Free Fatty Acids Assay kit (Nanjing Jiancheng
Bio-Engineering Institute Co., Ltd., Nanjing, China) according to
the manufacturer's protocol. Serum PA levels were examined by gas
chromatography-mass spectrometry according to the method of Han
et al (22).
IMTAG analysis
Soleus muscles were homogenized with normal saline
buffer (467 mmol/l NaCl, 10 mmol/l KCl, 1 mmol/l
NaH2PO4, 4 mmol/l NaHCO3, 8.4
mmol/l Na2SO4, 30 mmol/l HEPES, 10 mmol/l
EDTA; pH 7.1 with KOH) to obtain 10% muscle homogenates.
Subsequently, the levels of IMTAG were analyzed using the lipase
glycerol kinase calorimetric method as previously described
(23).
Histology analysis
Uterus, liver and soleus muscle samples were fixed
in 10% formalin for 1 week at room temperature, embedded in
paraffin, cut into 5-µm thick sections, and stained with 0.4%
hematoxylin for 5 min and 0.5% eosin for 4 min at room temperature.
All sections were imaged using a Nikon Eclipse 80i fluorescence
microscope (magnification, ×100; Nikon Corporation, Tokyo,
Japan).
Cell culture and drug treatment
The mouse myogenic cell line C2C12 was purchased
from the American Type Culture Collection (Manassas, VA, USA),
cultured in high-glucose Dulbecco's Modified Eagle's medium at 37°C
with 5% CO2 in a cell incubator, and differentiated into
myotubes with 2% horse serum (HyClone; GE Healthcare Life Sciences,
Logan, UT, USA) for 4 days. PA and E2 (Sigma-Aldrich;
Merck KGaA) were dissolved in ethanol and added to the culture
medium. The cells were divided into six groups: Untreated (CON),
absolute ethyl alcohol, 0.5 mmol/l PA, 0.5 mmol/l PA +
1×10−8 mol/l E2 (PA + E2), 0.5
mmol/l PA + 1×10−8 mol/l PPT (PA + PPT; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) and 0.5 mmol/l PA +
1×10−8 mol/l DPN (PA + DPN; Tokyo Chemical Industry Co.,
Ltd., Tokyo, Japan). E2, PPT and DPN were added to the
medium 8 h prior to treatment with PA at 37°C. Cells were treated
with PA for 24 h at 37°C, and cells were harvested for reverse
transcription-semi-quantitative polymerase chain reaction
(RT-sqPCR) and western blot analysis.
RT-sqPCR assay
Soleus muscles or myotubes formed by C2C12 cells
were used to detect the expression levels of ESR1, ESR2, PPARα,
ACACA, PLIN2, CD36, FASN and GAPDH with RT-sqPCR using a PCR
thermocycler (Omni Controls, Inc. Tampa, FL, USA). Total RNA was
extracted using TRIzol® reagent (Thermo Fisher
Scientific, Inc., Waltham, MA, USA) and reverse transcribed with
OligodT primers and Moloney murine leukemia virus reverse
transcriptase (Fermentas; Thermo Fisher Scientific, Inc.,
Pittsburgh, PA, USA) according to the manufacturer's protocols. The
primers (Table I) were designed
with Primer Premier 5 software (Premier Biosoft International, Palo
Alto, CA, USA) and synthesized by Sangon Biotech Co., Ltd.,
(Shanghai, China). The reaction was performed in 25 µl using Takara
Ex Taq Polymerase (Takara Biotechnology Co., Ltd., Dalian, China),
with the following thermocycling conditions: Initial denaturation
at 95°C for 3 min, 32 cycles of 30 sec at 95°C, 30 sec at 55°C and
30 sec at 72°C, with a final extension at 72°C for 10 min. A total
of 30 cycles were conducted for GAPDH. The PCR products (6 µl in
each well) were analyzed by electrophoresis on a 3% agarose gel;
the dye used for visualization was ethidium bromide. Gel-Pro
Analyzer 3.1 software (Media Cybernetics, Inc., Rockville, MD, USA)
was used to analyze the intensity of each band, and the relative
expression levels of the genes were normalized to GAPDH. Each
experiment was repeated three times.
 | Table I.Primers for reverse
transcription-semi-quantitative polymerase chain reaction. |
Table I.
Primers for reverse
transcription-semi-quantitative polymerase chain reaction.
| A, Rat |
|---|
|
|---|
| Gene | Primer sequence
(5′-3′) | Product size,
bp |
|---|
| ESR1 | F:
CTGCCAAGGAGACTCGCTAC | 315 |
|
| R:
AAGTGCCCATTTCATTTCG |
|
| ESR2 | F:
TTGTGCCAGCCCTGTTACTA | 200 |
|
| R:
TACGCCGGTTCTTGTCTAT |
|
| PPARα | F:
TGCTGTCCTCCTTGATGAAC | 270 |
|
| R:
GCTTGAGCACGTGCACAATC |
|
| ACACA | F:
AACCAGCACTCCCGATTC | 175 |
|
| R:
AGGCCAAACCATCCTGTAA |
|
| PLIN2 | F:
CTCTCGGCAGGATCAAAGAC | 171 |
|
| R:
CGTAGCCGACGATTCTCTTC |
|
| FASN | F:
CGGCGAGTCTATGCCACTAT | 398 |
|
| R:
ACACAGGGACCGAGTAATGC |
|
| CD36 | F:
CTCTGACATTTGCAGGTCCA | 214 |
|
| R:
CACAGGCTTTCCTTCTTTGC |
|
| GAPDH | F:
GCGAGATCCCGCTAACATCA | 178 |
|
| R:
CTCGTGGTTCACACCCATCA |
|
|
| B,
Mouse |
|
| Gene | Primer sequence
(5′-3′) | Product size,
bp |
|
| ESR1 | F:
TCCTAACTTGCTCCTGGACAGG | 78 |
|
| R:
GTAGCCAGCAACATGTCA |
|
| ESR2 | F:
TTCTTTCTCATGTCAGGCACA | 123 |
|
| R:
CTCGAAGCGTGTGAGCATT |
|
| PPARα | F:
CAAGTGCCTGTCTGTCGG | 236 |
|
| R:
TCTGGTCGTTGTTGGGCG |
|
| ACACA | F:
CCTGGAGTGGCAGTGGTCTTCG | 176 |
|
| R:
TCCTCCTCCCTCTGAGGCCTTG |
|
| PLIN2 | F:
CCCGCAACCTGACCCAGCAG | 114 |
|
| R:
CGCCTGCCATCACCCCCAAG |
|
| CD36 | F:
GAGCAACTGGTGGATGGTTT | 204 |
|
| R:
GCAGAATCAAGGGAGAGCAC |
|
| FASN | F:
GGAGGTGGTGATAGCCGGTAT | 140 |
|
| R:
TGGGTAATCCATAGAGCCCAG |
|
| GAPDH | F:
CATCTTCCAGGAGCGAGACC | 635 |
|
| R:
TGAAGTCGCAGGAGACAACC |
|
Western blot assay
Soleus muscles or myotubes formed by C2C12 cells
were homogenized in ice-cold radioimmunoprecipitation assay buffer
(Beyotime Institute of Biotechnology, Haimen, China). The total
concentration of protein in sample was determined using a
bicinchoninic acid assay. In total, 30 µg protein was loaded in
each lane. Proteins were separated by 10% SDS-PAGE and transferred
to polyvinylidene difluoride membranes. Following blocking in a
solution of 1X TBS, 0.1% Tween-20 (TBST) containing 5% non-fat dry
milk for 1 h at room temperature, the membranes were incubated with
the following primary antibodies overnight at 4°C: PPARα (1:400;
cat. no. bs-23398R; Bioss Antibodies Inc., Woburn, MA, USA), ACACA
(1:500; cat. no. 21923-1-AP; ProteinTech Group, Inc., Chicago, IL,
USA), phosphorylated (p-)ACACA (1:500; cat. no. 11818; Cell
Signaling Technology, Inc., Danvers, MA, USA), PLIN2 (1:500; cat.
no. bs-1164R; Bioss Antibodies Inc.), CD36 (1:500; cat. no.
sc-7309), FASN (1:500; cat. no. sc-48357; both Santa Cruz
Biotechnology, Inc.), MAPK8 (1:500; cat. no. BS3630), p-MAPK8
(1:500; cat. no. BS4763; all Biogot Technology Co., Ltd., Nanjing,
China), AKT serine/threonine kinase (AKT; 1:1,000; cat. no. 9272),
p-AKT (1:1,000; cat. no. 4060; both Cell Signaling Technology,
Inc.) and β-actin (1:1,000; cat. no. TA-09; Beijing Zhongshan
Golden Bridge Biotech, China). Then, the membranes were incubated
with the secondary antibodies, horseradish peroxidase-conjugated
goat anti-mouse (1:5,000; cat. no. S0002) and goat anti-rabbit
(1:5,000; cat. no. S0001; both Affinity Biosciences, Cincinnati,
OH, USA) for 1 h at room temperature. Detection was performed with
enhanced chemiluminescence substrate (Thermo Fisher Scientific,
Inc.) and quantification was performed with ImageJ software 1.48
(National Institutes of Health, Bethesda, MD, USA). The protein
expression levels were normalized to β-actin. Subsequently, the
ratio of phosphorylated/total protein was calculated. Each
experiment was repeated three times.
Statistical analysis
Data are presented as the mean ± standard error of
the mean (in tables) or standard deviation (in figures). P<0.05
was considered to indicate a statistically significant difference.
Statistical analyses were performed using one-way analysis of
variance followed by Fisher's Least Significant Difference (LSD)
test. Statistical analyses were performed using SPSS software
(version 23.0; IBM Corp., Armonk, NY, USA).
Results
Effects of E2 on body
weight and serum biochemical parameters in OVX rats
Female OVX rats exhibited a significant increase in
body weight compared with age-matched SHAM controls (Table II). Food intake in OVX rats was
increased, suggesting that surgical removal of ovaries induced
hyperphagia. OVX rats exhibited significant increases in the serum
levels of TG, TC, LDL-C, FFA and PA, but significant decreases in
the serum levels of HDL-C and E2. Treatment with
E2 decreased body weights, decreased the serum levels of
TG, TC, LDL-C, FFA and PA, and increased the serum levels of HDL-C
and E2 in the OVX group.
| Table II.Effects of treatment with
E2 in ovariectomized female rats. |
Table II.
Effects of treatment with
E2 in ovariectomized female rats.
| Parameters | Sham | OVX | OVX +
E2 |
|---|
| Body weight, g | 299.50±22.52 |
406.25±30.75c |
312.75±31.34f |
| Food intake, g | 19±1.23 |
26±2.65b |
21±1.78d |
| Triglycerides,
mmol/l | 0.44±0.027 |
0.56±0.070a |
0.38±0.056f |
| Total cholesterol,
mmol/l | 1.61±0.023 |
2.51±0.072c |
2.04±0.026f |
| High-density
lipoprotein cholesterol, mmol/l | 0.70±0.0058 |
0.54±0.071b |
0.68±0.12f |
| Low-density
lipoprotein cholesterol, mmol/l | 0.93±0.23 |
1.81±0.25c |
1.27±0.23d |
| Estradiol,
pmol/l | 93.5±4.65 |
72.5±12.71b |
128.75±6.24f |
| Free fatty acids,
mmol/l | 0.41±0.05 |
1.73±0.17a |
0.84±0.11d |
| Palmitic acid,
µg/ml | 210.81±2.63 |
655.29±18.25b |
350.96±5.47e |
Effects of E2 on
pathological alterations following OVX
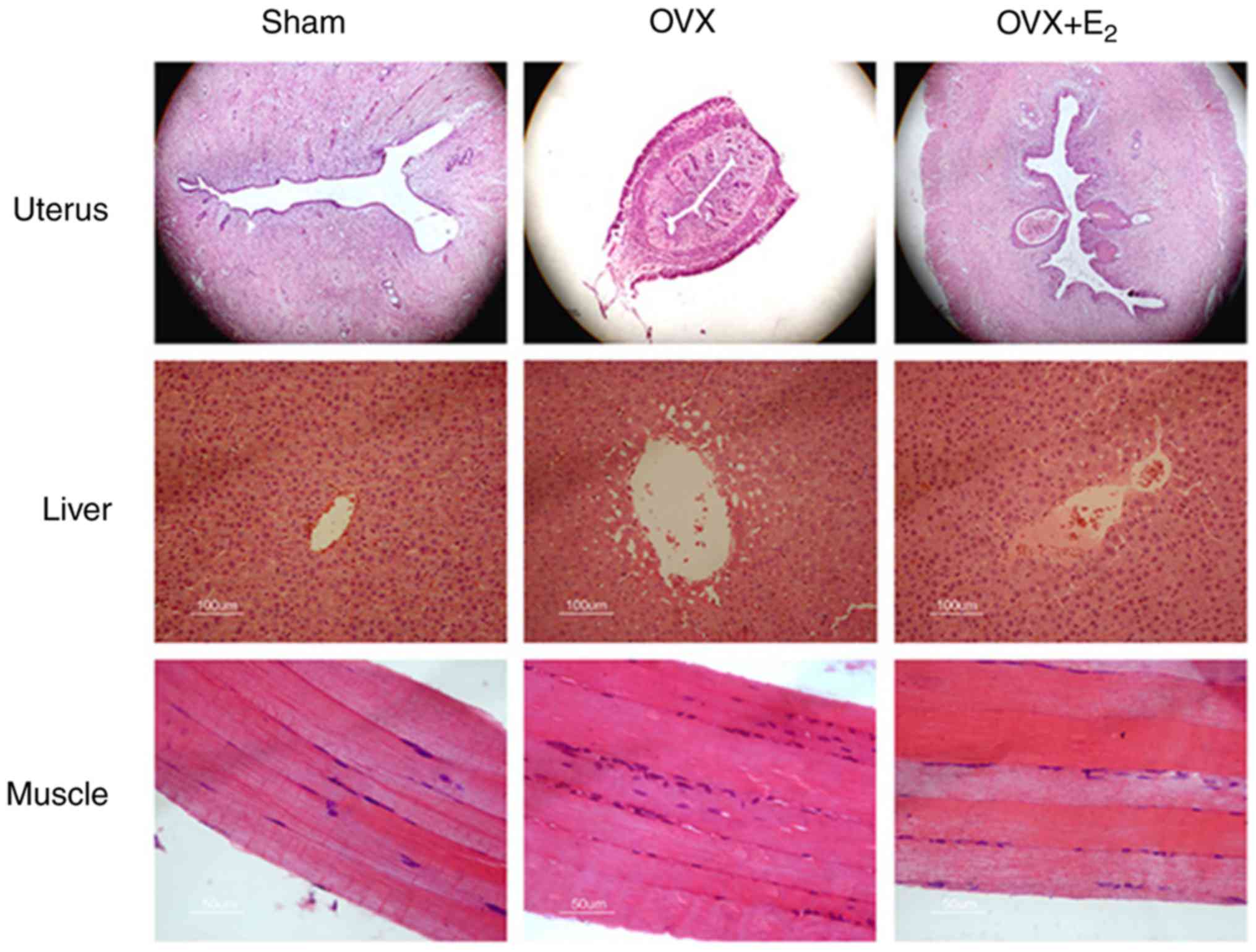
The volumes of the uterine gland and the uterus
decreased in OVX rats compared with the sham control group
(Fig. 1). The deposition of lipids
increased in liver tissue, and infiltration of inflammatory cell
into skeletal muscles were detected in the OVX group. Treatment
with E2 partially reversed the size of the uterine
gland, and decreased lipid deposition in the liver and aggregation
of inflammatory cells in the skeletal muscles.
Effects of E2 on insulin
resistance and circulating levels of IMTAG in OVX rats
GTT and ITT analyses suggested that rats in the OVX
group exhibited insulin resistance compared with the SHAM group.
However, rats in the OVX + E2 group exhibited a
significant improvement in insulin resistance (Fig. 2A-D), in addition to a decrease in
body mass (Table II). Following
insulin resistance, the protein expression levels of p-AKT and
p-MAPK serve as opposing markers for insulin signaling. Western
blotting suggested that the p-AKT/AKT ratio was significantly
decreased and the p-MAPK8/MAPK8 ratio was markedly increased in the
OVX group compared with the control. However, the p-AKT/AKT and
p-MAPK8/MAPK8 ratios were restored following treatment with
E2, in line with the GTT and ITT results (Fig. 2E and F). The levels of TG in muscle
tissues were measured, and decreased estrogen levels were
identified to be associated with a significant increase in the
intracellular levels of IMTAG in OVX rats compared with the control
(Fig. 2G). TG deposition was not
observed in the muscle tissue following H&E staining (Fig. 1). In contrast, the intracellular
levels of IMTAG were significantly decreased in the OVX +
E2 group compared with OVX alone (Fig. 2G). This suggested that treatment
with E2 may decrease the synthesis of TG.
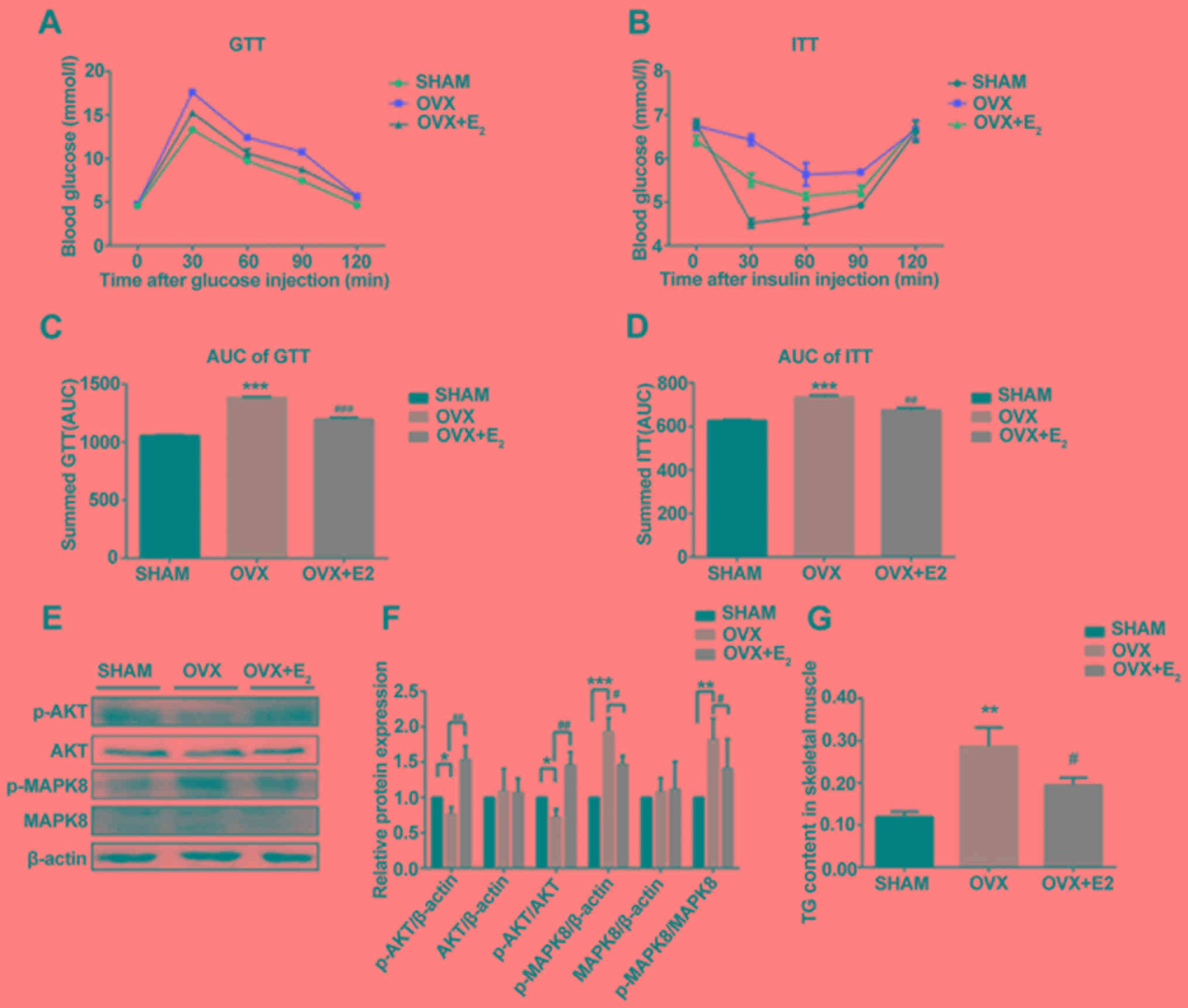 | Figure 2.Effects of E2 on insulin
resistance. (A) GTT analysis. (B) ITT analysis. (C) AUC of GTT
analysis. (D) AUC of ITT analysis. (E) Protein expression levels of
p-AKT, AKT, p-MAPK8 and MAPK8 in skeletal muscles. (F)
Densitometric analysis of (E). (G) Levels of TG in skeletal muscles
in various conditions. Data are presented as the mean ± standard
deviation. **P<0.01, ***P<0.001 vs. sham control group;
#P<0.05, ##P<0.01,
###P<0.001 vs. OVX group. GTT, glucose tolerance
test; ITT, insulin tolerance test; p-, phosphorylated; MAPK,
mitogen-activated protein kinase; AKT, AKT serine/threonine kinase;
TG, triglycerides; AUC, area under the curve; OVX, ovariectomy;
E2, 17β-estradiol. |
Effects of E2 on gene
expressions in OVX rats
The results of the present study suggested that the
expression levels of ESR1 significantly decreased in OVX rats
compared with the control group; however, ESR2 expression was
increased in OVX rats. The expression levels of ESR1 and ESR2 were
significantly restored in OVX rats following treatment with
E2 (Fig. 3A and B).
Since treatment with E2 decreased the levels of TG
(Fig. 2G), the mRNA and protein
expression levels of three proteins involved in TG synthesis were
examined. The mRNA expression levels of ACACA, PLIN2 and FASN
increased significantly in the OVX group compared with the control
group, but were decreased in the OVX + E2 group compared
with OVX alone (Fig. 3A and B).
The protein expression levels of PLIN2 and FASN exhibited the same
trend; however, that of ACACA was markedly unaltered. Therefore,
the protein expression levels of p-ACACA, the activated form of
ACACA, were examined. The results of western blotting suggested
that the p-ACACA/ACACA ratio was decreased in the OVX group and was
significantly restored following treatment with E2
(Fig. 3C and D). Additionally, the
present RT-sqPCR and western blotting results suggested that the
mRNA and protein expression levels of CD36 and PPARα were decreased
in OVX rats, and treatment with E2 partially reversed
these effects. The alterations in the expression levels of ESR1,
CD36 and PPARα were similar following OVX and treatment with
E2, suggesting that these factors may be involved in the
same pathway.
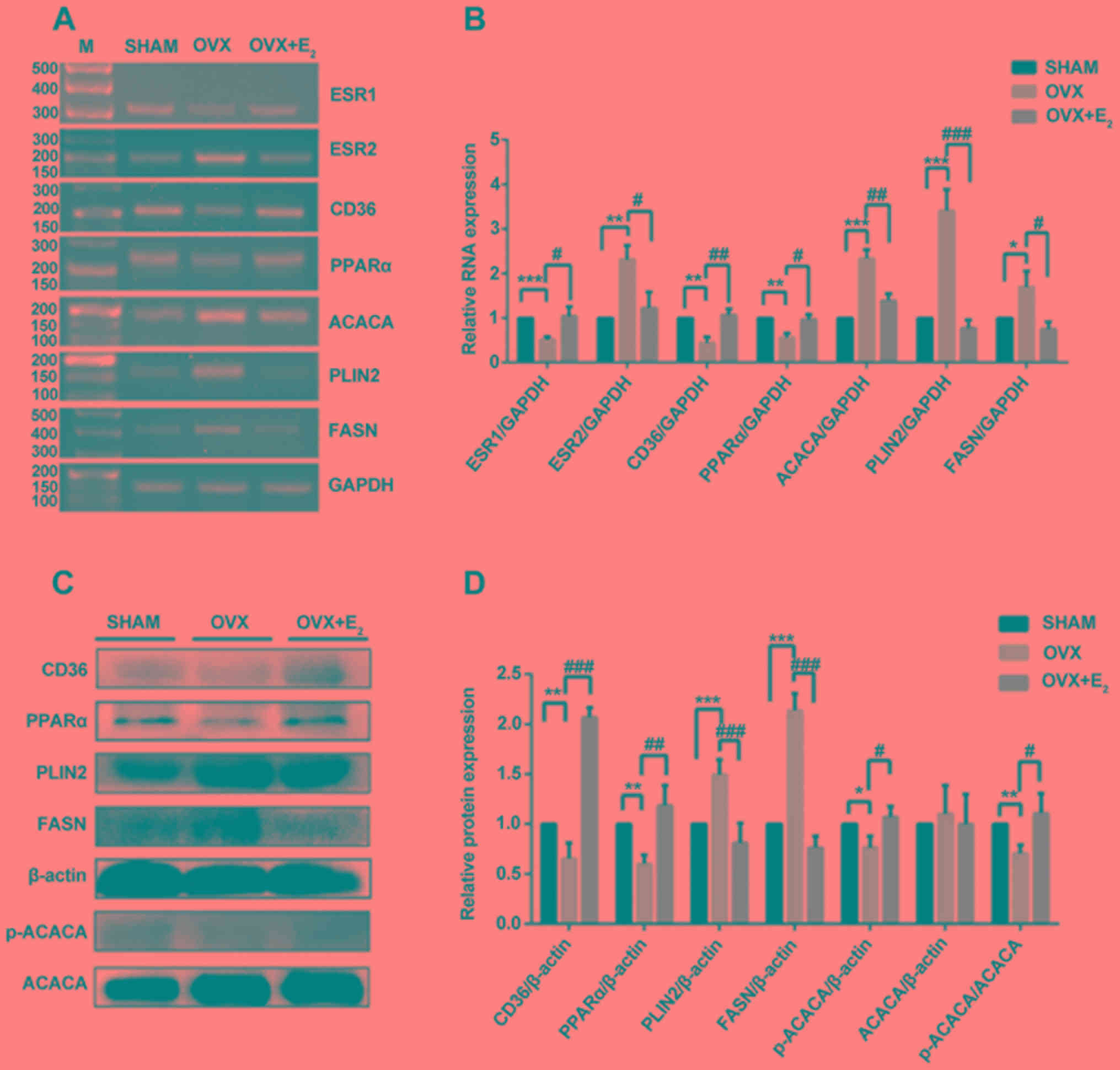 | Figure 3.Analysis of gene expression in
skeletal muscles. (A) RT-sqPCR and (B) densitometry analysis of
ESR1, ESR2, CD36, PPARα, ACACA, PLIN2 and FASN in skeletal muscles
in three conditions. (C) Western blot and (D) densitometry analyses
of CD36, PPARα, PLIN2, FASN, p-ACACA and ACACA in skeletal muscles.
Data are presented as the mean ± standard deviation. *P<0.05,
**P<0.01, ***P<0.001 vs. sham control group.
#P<0.05, ##P<0.01,
###P<0.001 vs. OVX group. RT-sqPCR, reverse
transcription-semi-quantitative polymerase chain reaction; PLIN2,
perilipin 2; CD36, CD36 molecule; ACACA, acetyl-CoA carboxylase α;
FASN, fatty acid synthase; PPARα, peroxisome proliferator activated
receptor α; ESR, estrogen receptor; OVX, ovariectomy;
E2, 17β-estradiol; p-, phosphorylated; M, marker. |
Effects of E2, PPT and DPN
on the expression levels of metabolism-associated factors in C2C12
cells treated with PA
Treatment with PA significantly decreased the
expression levels of ESR1 mRNA in differentiated C2C12 cells
compared with the control. In contrast, pretreatment with
E2 or PPT led to a significant increase in the
expression of ESR1 compared with PA treatment alone. However, the
expression level of ESR2 significantly increased following
treatment with PA compared with the control, and was partially
restored by pretreating C2C12 cells with E2 or PPT;
although the difference between pretreated cells and non-pretreated
cells was not significant (Fig. 4A and
B). Similarly, the mRNA and protein expression levels of PLIN2
and FASN significantly increased following treatment with PA, and
these effects were reversed by pretreating C2C12 cells with
E2 or PPT, which was similarly to the mRNA expression
profile of ACACA (Fig. 4A and B).
Notably, the protein expression level of ACACA was markedly
unaltered; however, the p-ACACA/t-ACACA ratio was increased in the
PA + E2 and PA + PPT groups compared with PA treatment
alone (Fig. 4C and D).
Additionally, the present RT-sqPCR and western blotting results
suggested that the expression levels of PPARα and CD36 were
significantly decreased in the PA group compared with the control,
and were increased in the PA + E2 and PA + PPT groups
compared with PA treatment alone (Fig.
4A-D). However, the expression levels of the aforementioned
factors were not significantly altered between the PA group and the
PA + DPN group. Treatment with PA significantly decreased the
p-AKT/AKT ratio and increased the p-MAPK8/MAPK ratio compared with
the control. Pretreatment with E2 or PPT reversed these
effects, and pretreatment with DPN restored the levels of
p-MAPK8/MAPK, suggesting that ESR1 activation improved insulin
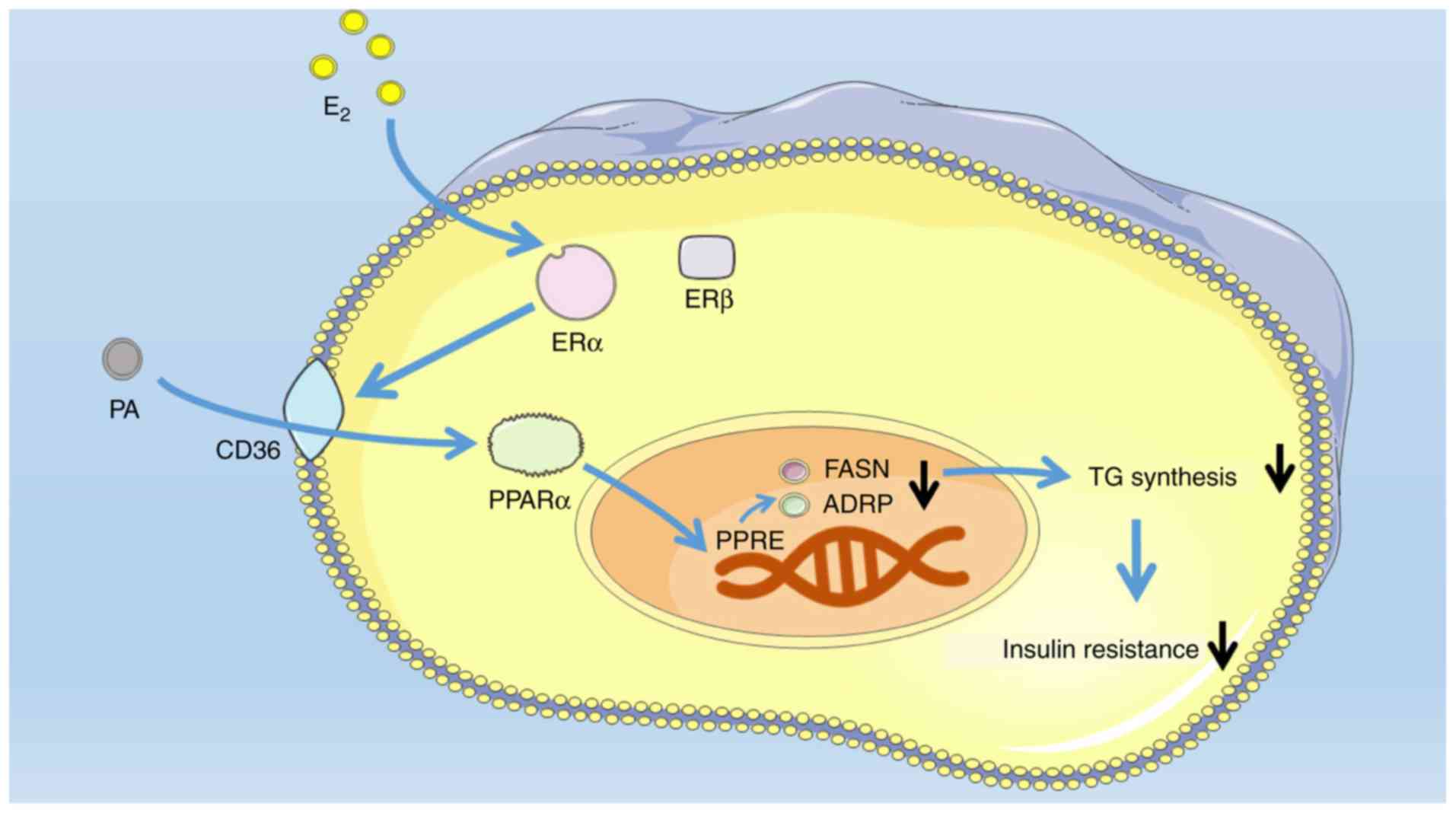
resistance in differentiated C2C12 cells (Fig. 4E and F). Collectively, the present
study investigated the role of E2 and PA in the
metabolic alterations occurring in skeletal muscle cells (Fig. 5).
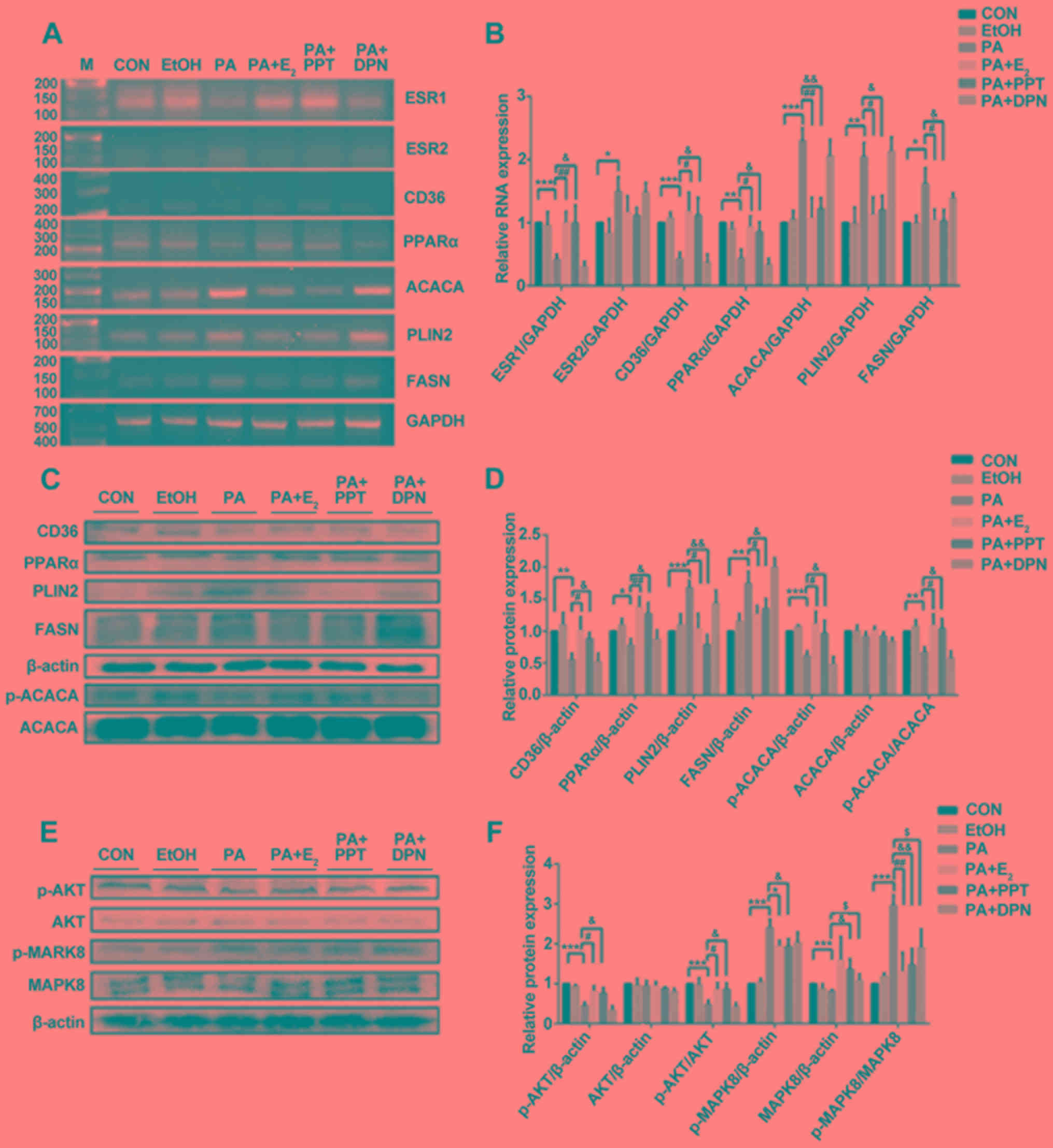 | Figure 4.Analysis of gene expression in
differentiated C2C12 cells. (A) RT-sqPCR and (B) densitometry
analysis of ESR1, ESR2, CD36, PPARα, ACACA, PLIN2 and FASN
expression in C2C12 cells. (C) Western blot and (D) densitometry
analyses of CD36, PPARα, PLIN2, FASN, p-ACACA and ACACA in C2C12.
(E) Western blot and (F) densitometry analyses of p-AKT, AKT,
p-MAPK8 and MAPK8 in C2C12. Data are presented as the mean ±
standard deviation. *P<0.05, **P<0.01, ***P<0.001 vs. CON
group; #P<0.05, ##P<0.01 vs. PA +
E2; &P<0.05,
&&P<0.01, vs. PA + PPT; $P<0.5
vs. PA + DPN. RT-sqPCR, reverse transcription-semi-quantitative
polymerase chain reaction; PLIN2, perilipin 2; CD36, CD36 molecule;
ACACA, acetyl-CoA carboxylase α; FASN, fatty acid synthase; PPARα,
peroxisome proliferator activated receptor α; ESR, estrogen
receptor; OVX, ovariectomy; E2, 17β-estradiol; p-,
phosphorylated; MAPK, mitogen-activated protein kinase; AKT, AKT
serine/threonine kinase; CON, control; PA, palmitic acid; PPT,
propylpyrazoletriol; DPN, diarylpropionitrile; EtOH, ethanol; M,
marker. |
Discussion
Due to the increasing incidence of female obesity
and the weight gain observed following menopause (24), the identification of novel
strategies to decrease the occurrence rate of postmenopausal
obesity is required. The present study suggested that treatment
with E2 decreased body weight, intracellular levels of
IMTAG, and serum levels of TG, TC and LDL-C following OVX.
Additionally, OVX led to an increase in the expression levels of
PLIN2, FASN and ACACA and a decrease in the expression levels of
ESR1, CD36 and PPARα. Notably, treatment with E2
reversed these effects, suggesting that E2 may inhibit
TG synthesis, thus improving insulin resistance through the
ESR1-CD36-PPARα pathway in skeletal muscles.
OVX and ER-knockout mice were previously identified
to develop obesity, and treatment with estrogen reversed these
effects (25). The present study
suggested that the body weight and the protein expression of
p-MAPK8 increased following OVX, whereas the protein expression
level of p-AKT increased. In addition to the phosphorylation levels
of MAPK8 and AKT, the GTT and ITT results suggested the occurrence
of insulin resistance following OVX. Treatment with E2
decreased the body weight in OVX rats, and insulin resistance was
improved, in line with previous investigations in human (26).
In normal skeletal muscle cells, FA uptake and TG
synthesis are regulated by negative feedback mechanisms, so that
lipid accumulation does not severely affect cellular function
(27). Impaired regulation of FA
uptake and TG synthesis in skeletal muscle is involved in insulin
resistance (28). Following
treatment with E2, and ESR1 and ESR2 agonists, muscle
lipogenesis and TG accumulation were identified to be significantly
decreased (29). The present
results suggested that the levels of IMTAG increased in OVX rats,
whereas the levels of IMTAG decreased following treatment with
E2, suggesting an involvement of de novo TG
synthesis in skeletal muscle following OVX. Additionally, treatment
with E2 decreased synthesis of TG, as suggested by IMTAG
detection; however, no alterations in lipid deposition were
observed in skeletal muscles.
The expression of PLIN2 is specific of certain types
of muscle fibers, and its expression level is increased in muscle
fibers containing high levels of IMTAG (30). The present results suggested that
the mRNA and protein expression levels of PLIN2 were increased in
skeletal muscles following OVX and in C2C12 cells treated with PA.
However, treatment with E2 and activation of ESR1
decreased the expression level of PLIN2 in muscle tissues and
cells, suggesting that E2 may be involved in the
expression level of PLIN2 by activating ESR1.
PPARα regulates the expression levels of various
genes involved in lipolysis and lipoprotein metabolism (31). Furthermore, OVX altered the
expression levels of factors involved in lipogenesis, including
FASN and ACACA, and in lipolysis, including PPARα. Notably,
treatment with E2 and activation of ESR1 reversed the
effects of OVX, in line with the previous study by Minnaard et
al (32). A discrepancy
between the mRNA and protein expression levels of ACACA was
identified, possibly due to post-transcriptional modifications of
ACACA. A limitation of the present study is that the mRNA
expression levels of the genes investigated were assessed only by
RT-sqPCR; however, quantitative PCR was not performed.
Previous studies using a rat model of OVX
demonstrated that certain types of ER ligands may exhibit
protective effects in skeletal muscle (33). In addition, it was observed that
ESR1, but not ESR2, may be able to maintain mitochondrial function
and metabolic homeostasis, and may exhibit a protective effect
against inflammation (34,35). The present results suggested that
the expression levels of ESR1, PPARα and CD36 was decreased
following OVX or treatment with PA, and the levels of these factors
increased following treatment with E2 or pretreatment
with PPT, suggesting that these three factors may be part of the
same pathway.
CD36 is broadly expressed, and it was identified to
be involved in FA and lipid metabolism in pathological conditions
(36). CD36 is a transporter of
FA, and it may represent a molecular target to protect myocytes
against lipotoxicity (36).
However, the effect of ESR1 activation on the expression level of
CD36 remains unclear. The present results suggested that treatment
with PPT, an ESR1 agonist, increased the mRNA and protein
expression levels of CD36 suggesting that the activity of ESR1 in
skeletal muscle may be modulated to treat diseases associated with
metabolic syndrome.
The ESR2 signaling pathway is involved in the
regulation of skeletal muscle growth and regeneration by
stimulating anabolic pathways, activating satellite cells, and
modulating immune response (33).
In the present study, the expression level of ESR2 was examined in
skeletal muscle and in cells treated with PA. Notably, the
expression level of ESR2 was increased following OVX compared with
the control. Treatment with PA increased the expression level of
ESR2; however, pretreatment with DPN did not affect the expression
levels of CD36, PPARα, ACACA, PLIN2 and FASN in C2C12 cells treated
with PA, suggesting that ESR2 may not be involved in the
ESR1-CD36-PPARα pathway.
Collectively, the present results suggested that
E2, by activating ESR1, may modulate TG synthesis in
skeletal muscle during menopause, and it may represent a novel
complementary therapy to treat postmenopausal obesity. Notably,
ESR1 and ESR2 were identified to have distinct functions, and
further loss-of-function experiments are required to examine the
function of each receptor in muscle cells.
Acknowledgements
The authors would like to thank Miss Li Gui and Mr
Dake Huang at the Comprehensive Laboratory, Anhui Medical
University for H&E staining.
Funding
The present study was funded by The Scientific
Research Foundation of Anhui Medical University (grant no.
2017XKJ027), The National Innovation and Entrepreneurship Project
(grant no. 201710366019), The Natural Science Foundation of Anhui
Province Education Department (grant no. KJ2017A186) and The Fund
of Anhui Natural Science Foundation of China (grant no.
1808085MH233).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YL conceived and designed the study. QL, RL and GC
performed the cell culture, RT-sqPCR and western blotting
experiments. JW conducted the GTT/ITT experiment, and detected
serum lipid and E2. BH performed the IMTAG and histology
analysis. RL analyzed the data. CL and XZ conducted the animal
experiments. YL drafted the manuscript.
Ethics approval and consent to
participate
All animal experiments were reviewed and approved by
The Ethics Committee of Anhui Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Endo K, Weng H, Naito Y, Sasaoka T,
Takahashi A, Fukushima Y and Iwai N: Classification of various
muscular tissues using miRNA profiling. Biomed Res. 34:289–299.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pierdominici M, Ortona E, Franconi F,
Caprio M, Straface E and Malorni W: Gender specific aspects of cell
death in the cardiovascular system. Curr Pharm Des. 17:1046–1055.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Welle S, Tawil R and Thornton CA:
Sex-related differences in gene expression in human skeletal
muscle. PLoS One. 3:e13852008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dewailly D, Robin G, Peigne M, Decanter C,
Pigny P and Catteau-Jonard S: Interactions between androgens, FSH,
anti-Müllerian hormone and estradiol during folliculogenesis in the
human normal and polycystic ovary. Hum Reprod Update. 22:709–724.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Galluzzo P, Rastelli C, Bulzomi P,
Acconcia F, Pallottini V and Marino M: 17beta-Estradiol regulates
the first steps of skeletal muscle cell differentiation via
ER-alpha-mediated signals. Am J Physiol Cell Physiol.
297:C1249–C1262. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sipilä S, Finni T and Kovanen V: Estrogen
influences on neuromuscular function in postmenopausal women.
Calcif Tissue Int. 96:222–233. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hevener AL, Clegg DJ and Mauvais-Jarvis F:
Impaired estrogen receptor action in the pathogenesis of the
metabolic syndrome. Mol Cell Endocrinol. 418:306–321. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tena-Sempere M: Neuroendocrinology in
2016: Neuroendocrine control of metabolism and reproduction. Nat
Rev Endocrinol. 13:67–68. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Martínez de Morentin PB, Lage R,
González-García I, Ruíz-Pino F, Martins L, Fernández-Mallo D,
Gallego R, Fernø J, Señarís R, Saha AK, et al: Pregnancy induces
resistance to the anorectic effect of hypothalamic malonyl-CoA and
the thermogenic effect of hypothalamic AMPK inhibition in female
rats. Endocrinology. 156:947–960. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mauvais-Jarvis F, Clegg DJ and Hevener AL:
The role of estrogens in control of energy balance and glucose
homeostasis. Endocr Rev. 34:309–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Moran AL, Warren GL and Lowe DA: Removal
of ovarian hormones from mature mice detrimentally affects muscle
contractile function and myosin structural distribution. J Appl
Physiol (1985). 100:548–559. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Klinge CM: Estrogens regulate life and
death in mitochondria. J Bioenerg Biomembr. 49:307–324. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
López M and Tena-Sempere M: Estrogens and
the control of energy homeostasis: A brain perspective. Trends
Endocrinol Metab. 26:411–421. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hamilton DJ, Minze LJ, Kumar T, Cao TN,
Lyon CJ, Geiger PC, Hsueh WA and Gupte AA: Estrogen receptor alpha
activation enhances mitochondrial function and systemic metabolism
in high-fat-fed ovariectomized mice. Physiol Rep. 4:e129132016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
van Hall G: The physiological regulation
of skeletal muscle fatty acid supply and oxidation during
moderate-intensity exercise. Sports Med. 45 (Suppl 1):S23–S32.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kewalramani G, Bilan PJ and Klip A: Muscle
insulin resistance: Assault by lipids, cytokines and local
macrophages. Curr Opin Clin Nutr Metab Care. 13:382–390. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tepavcevic S, Koricanac G, Zakula Z,
Milosavljevic T, Stojiljkovic M and Isenovic ER: Interaction
between insulin and estradiol-17β in regulation of cardiac glucose
and free fatty acid transporters. Horm Metab Res. 43:524–530. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Stavinoha MA, RaySpellicy JW, Essop MF,
Graveleau C, Abel ED, Hart-Sailors ML, Mersmann HJ, Bray MS and
Young ME: Evidence for mitochondrial thioesterase 1 as a peroxisome
proliferator-activated receptor-alpha-regulated gene in cardiac and
skeletal muscle. Am J Physiol Endocrinol Metab. 287:E888–E895.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mosti MP, Ericsson M, Erben RG, Schüler C,
Syversen U and Stunes AK: The PPARα agonist fenofibrate improves
the musculoskeletal effects of exercise in ovariectomized rats.
Endocrinology. 157:3924–3934. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gorres BK, Bomhoff GL, Morris JK and
Geiger PC: In vivo stimulation of oestrogen receptor α increases
insulin-stimulated skeletal muscle glucose uptake. J Physiol.
589:2041–2054. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
van de Heijning BJM, Oosting A, Kegler D
and van der Beek EM: An increased dietary supply of medium-chain
fatty acids during early weaning in rodents prevents excessive fat
accumulation in adulthood. Nutrients. 9:E6312017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Han LD, Xia JF, Liang QL, Wang Y, Wang YM,
Hu P, Li P and Luo GA: Plasma esterified and non-esterified fatty
acids metabolic profiling using gas-chromatography mass
spectrometry and its application in the study of diabetic mellitus
and diabetic nephropathy. Anal Chim Acta. 689:85–91. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tarchalski J, Guzik P and Wysocki H:
Correlation between the extent of coronary atherosclerosis and
lipid profile. Mol Cell Biochem. 246:25–30. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Salpeter SR, Walsh JM, Ormiston TM,
Greyber E, Buckley NS and Salpeter EE: Meta-analysis: Effect of
hormone-replacement therapy on components of the metabolic syndrome
in postmenopausal women. Diabetes Obes Metab. 8:538–554. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Manrique C, Lastra G, Habibi J, Mugerfeld
I, Garro M and Sowers JR: Loss of estrogen receptor signaling leads
to insulin resistance and obesity in young and adult female mice.
Cardiorenal Med. 2:200–210. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Marchand GB, Carreau AM, Weisnagel SJ,
Bergeron J, Labrie F, Lemieux S and Tchernof A: Increased body fat
mass explains the positive association between circulating
estradiol and insulin resistance in postmenopausal women. Am J
Physiol Endocrinol Metab. 314:E448–E456. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Manco M, Mingrone G, Greco AV, Capristo E,
Gniuli D, De Gaetano A and Gasbarrini G: Insulin resistance
directly correlates with increased saturated fatty acids in
skeletal muscle triglycerides. Metabolism. 49:220–224. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Strehlow K, Rotter S, Wassmann S, Adam O,
Grohé C, Laufs K, Böhm M and Nickenig G: Modulation of antioxidant
enzyme expression and function by estrogen. Circ Res. 93:170–177.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Huang L, Tepaamorndech S, Kirschke CP,
Newman JW, Keyes WR, Pedersen TL and Dumnil J: Aberrant fatty acid
metabolism in skeletal muscle contributes to insulin resistance in
zinc transporter 7 (znt7)-knockout mice. J Biol Chem.
293:7549–7563. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Weigt C, Hertrampf T, Kluxen FM, Flenker
U, Hülsemann F, Fritzemeier KH and Diel P: Molecular effects of ER
alpha- and beta-selective agonists on regulation of energy
homeostasis in obese female Wistar rats. Mol Cell Endocrinol.
377:147–158. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Duval C, Müller M and Kersten S: PPARalpha
and dyslipidemia. Biochim Biophys Acta. 1771:961–971. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Minnaard R, Schrauwen P, Schaart G,
Jorgensen JA, Lenaers E, Mensink M and Hesselink MK: Adipocyte
differentiation-related protein and OXPAT in rat human skeletal
muscle: Involvement in lipid accumulation and type 2 diabetes
mellitus. J Clin Endocrinol Metab. 94:4077–4085. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Velders M, Schleipen B, Fritzemeier KH,
Zierau O and Diel P: Selective estrogen receptor-β activation
stimulates skeletal muscle growth and regeneration. FASEB J.
26:1909–1920. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ribas V, Drew BG, Zhou Z, Phun J, Kalajian
NY, Soleymani T, Daraei P, Widjaja K, Wanagat J, de Aguiar Vallim
TQ, et al: Skeletal muscle action of estrogen receptor α is
critical for the maintenance of mitochondrial function and
metabolic homeostasis in females. Sci Transl Med. 8:334ra542016.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ribas V, Nguyen MT, Henstridge DC, Nguyen
AK, Beaven SW, Watt MJ and Hevener AL: Impaired oxidative
metabolism and inflammation are associated with insulin resistance
in ERalpha-deficient mice. Am J Physiol Endocrinol Metab.
298:E304–E319. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Glatz JFC and Luiken JJFP: Dynamic role of
the transmembrane glycoprotein CD36 (SR-B2) in cellular fatty acid
uptake and utilization. J Lipid Res. 59:1084–1093. 2018. View Article : Google Scholar : PubMed/NCBI
|