Introduction
Neuropeptide B is a 29 amino acid peptide with a
C-6-brominated tryptophan residue at the N terminus (1,2). Its
biological activities are mediated by activation of two GPCR
receptors, termed NPBW1 (GPR7) and NPBW2 (GPR8). Both types of
receptors are expressed in humans, whereas rodents only express
NPBW1 (3). NPB and its receptors
are predominantly expressed in the central nervous system (3). In the brain, NPB is implicated in
controlling a variety of functions, including modulation of the
neuroendocrine axis, pain, appetite or circadian rhythm (4–7).
However, NPB and its receptor are also present in peripheral
tissues such as thyroid and adrenal glands, gonads and endocrine
pancreas (8). There is growing
evidence that NPB as well as NPBW1 play prominent roles in
controlling energy homeostasis. Animal studies have shown that mice
lacking NPBW1 (NPBW1-/-) develop mild adult onset obesity, and have
lower energy expenditure and higher blood glucose levels (9). Furthermore, increased body weight has
been reported in mice lacking NPB (NPB-/-mice) (10). We found that NPB and its receptor
are present in rat adipocytes (11) where NPB stimulates lipolysis and
suppresses leptin expression, and secretion (11). Others have reported that NPB serum
level is upregulated in humans who suffer from anorexia nervosa
(12), while it is reduced in type
1 diabetic patients (13).
Overall, these results collectively indicate that the NPB/NPBW1
system is involved in controlling body weight and energy
homeostasis and its alternation may contribute to obesity.
Energy homeostasis and metabolism are modulated by
insulin which is released from pancreatic beta cells in a
glucose-dependent fashion (14).
The loss and dysfunction of pancreatic beta cell function are
hallmarks of type 1 and type 2 diabetes (15,16).
By contrast, non-diabetic obese individuals display increased beta
cell mass (15). Although the
contribution of NPB/NPBW1 to modulation of energy homeostasis and
body weight regulation is well-documented in the literature
[reviewed in (4,17)], the role of NPB in controlling
pancreatic beta cell functions-insulin expression and secretion as
well as beta cell replication-is unknown. Furthermore, NPBW1
signaling in pancreatic beta cells is poorly understood. Thus, in
the present study we assessed the effects of NPB on insulin
expression and secretion as well as cell proliferation in
insulin-producing INS-1E cells [beta cell surrogate (18)] and rat pancreatic islets.
Materials and methods
Reagents
(Des-Br)-Neuropeptide B-29 was purchased from
Phoenix Pharmaceuticals (Burlingame, CA, USA). Cell culture media
and supplements were from Biowest (Nuaillé, France).
3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide
(MTT) was from Calbiochem (Merck, Darmstadt, Germany). The BrdU
Cell Proliferation kit and Cell Death Detection ELISA PLUS kit were
from Roche Diagnostic. Phospho-ERK1/2 (cat. no. 9101S) and ERK1/2
(cat. no. 9102S) antibodies and HRP-linked anti-rabbit antibody
(cat. no. 7074S) were from Cell Signaling Technology (Danvers, MA,
USA). GAPDH antibody was from Sigma-Aldrich (St. Louis, MO, USA).
Other reagents were purchased from Sigma-Aldrich, unless otherwise
stated.
Cell culture
Rat insulin-producing INS-1E cells were kindly
provided by Professor Pierre Maechler (Médical Universitaire,
Genève, Switzerland). Cells were cultured in RPMI-1640 medium
supplemented with 10% FBS, 2 mmol/l glutamine, 10 mmol/l HEPES
buffer, 1 mmol/l sodium pyruvate, 50 µmol/l beta-mercaptoethanol
and 100 kU/l penicillin, 100 mg/l streptomycin.
Isolation of pancreatic islets
Male Wistar rats (body weight 300–350 g) were from
Department of Toxicology (Poznań University of Medical Sciences,
Poznań, Poland). Animals were sacrificed by decapitation and then
the abdominal cavity was opened and the pancreas was filled out by
10 ml of Hanks' buffer containing (in mmol/l): 137 NaCl, 5.37 KCl,
4.17 NaHCO3, 1.26 CaCl2, 0.84
MgSO4, 0.44 KH2HPO4, and 0.34
Na2HPO4, pH 7.4) supplemented in 1 Wünsch
Units/ml of Liberase DL [0.02% (w/v) collagenase; Roche Diagnostic,
Germany]. The filled pancreas was immediately cut and placed in a
Falcon tube containing 3 ml of Hanks' buffer with Liberase DL.
Samples were digested in a water bath (37°C) for 15 min. This
enzymatic process was terminated by the addition of 90 ml 10% FCS
in Hanks' buffer. Pancreatic islets were washed several times in
Hanks' buffer until the islet preparation was clear. The washing
procedure was based on mixing the sample and allowing sedimentation
of islets for 3 min. Then, Hanks' buffer with debris was aspirated
and sedimented islets were retained. Finally, islets were picked by
pipette and transferred to RPMI-1650 medium supplemented with 1%
BSA. Then, islets were placed into an incubator for regeneration.
After 3 h, the islets were ready for use in all the described
experiments.
Reverse transcription-quantitative (RT-q) PCR. Total
RNA was isolated using Extrazol (DNA Gdansk, Gdansk, Poland). One
microgram of total RNA was reverse transcribed to cDNA using
FIREScript RT cDNA Synthesis Mix (Solis BioDyne, Tartu, Estonia). A
multiplex RT-qPCR reaction was performed using a QuantStudio 12K
Flex (Thermo Fisher Scientific, Waltham, MA, USA). Primers and
TaqMan probes were from Life Technologies (Carlsbad, CA, USA).
Primers with their Applied Biosystems Assay IDs are as follows:
Npbw1, Rn01772104_s1; Npb, Rn00596187_g1;
Ins1, Rn02121433_g1; Ins2, Rn01774648_g1; Gck,
Rn00561265_m1; Glut2, Rn00563565_m1; Hnf4α,
Rn04339144_m1; Mafa, Rn00845206_s1; Pdx1,
Rn00755591_m1; Pgc1α, Rn00580241_m1; Hprt1,
Rn01527840_m1. Gene expression was evaluated by the
2−∆∆Cq method; Hprt1 was used as the endogenous
control.
Western blot analysis
The Western blot procedure was performed as
previously described (19).
Briefly, total protein was isolated using RIPA buffer containing 50
mmol/l Tris-HCl (pH 8.0), 150 mmol/l NaCl, 1.0% NP-40, 0.5% sodium
deoxycholate, 0.1% SDS and a protease inhibitor cocktail (Roche
Diagnostics). Proteins separated by SDS-PAGE gel electrophoresis
were transferred into a nitrocellulose membrane and non-specific
binding was blocked using 5% bovine serum albumin in Tris Buffered
Saline containing Tween-20 (TBST) for 1 h at room temperature (RT).
Thereafter, the membrane was incubated overnight with primary
anti-phosphorylated ERK1/2 rabbit polyclonal antibody diluted to
1:1,000 at 4°C. After washing in TBST, the membrane was incubated
with anti-rabbit secondary antibody diluted to 1:5,000 for 1 h at
RT. Signals were visualized by enhanced chemiluminescence (ECL kit,
Pierce Biotechnology, Rockford, IL, USA). Membranes were further
stripped and reprobed for total ERK1/2 and as a loading control
GAPDH.
Insulin secretion
INS-1E cells were seeded into 24-well plates
(1.5×105 cells/well) and cultured for 48 h. Following 1
h preincubation in glucose-free Krebs-Ringer-HEPES buffer (KRHB)
containing (units: mmo/l) 136 NaCl, 4.7 KCl, 1 CaCl2,
1.2 MgSO4, 1.2 KH2PO2, 2
NaHCO3, 10 HEPES (pH 7.4) and 0.1% free fatty acid BSA,
cells were washed with KRHB and exposed to 2.8 mmol/l or 16.8
mmol/l glucose in KRHB with or without 100 nmol/l NPB for 60 min.
Doses of NPB were chosen based on previous in vitro studies
(11,20). Next, the medium was collected and
centrifuged at 250 × g for 5 min. Insulin content was determined in
the supernatants using a High Range Rat Insulin ELISA kit
(EIA-3985, DRG Instruments GmbH, Germany, Marburg). Data were
normalized to protein concentrations determined by a BCA Protein
Assay kit (Thermo Scientific, Waltham, MA, USA).
In the case of pancreatic islets, groups of five rat
pancreatic islets of similar size were incubated in KRHB containing
0.1% free fatty acid BSA with or without 100 nmol/l NPB and 2.8 or
16.7 mmol/l glucose for 60 min. Insulin concentration in the
incubation medium was measured as described above for INS-1E
cells.
Viability and cell proliferation
INS-1E cells were seeded into a 96-well plate
(4×104 cells/well) and cultured for 48 h. After
overnight preincubation in serum-free medium, cells were incubated
in serum-free medium supplemented with 0.1% free fatty acid BSA and
1, 10 or 100 nmol/l NPB for 24 or 48 h. Cell viability was
determined by an MTT assay; cell proliferation was studied using a
Cell Proliferation ELISA BrdU kit (Roche Diagnostics) according to
the manufacturer's procedure.
Cell death
INS-1E cells (1.5×105 cells/well) or rat
pancreatic islets (8 islets/well) were cultured in 24-well plates
and treated with or without NPB (100 nmol/l) for 48 h. Cell death
was measured by a Cell Death Detection ELISA PLUS kit (Roche
Diagnostics), according to the manufacturer's protocol.
Statistical analysis
Data are presented as the mean ± standard error of
the mean. Statistical analysis was performed using GraphPad Prism
version 8.0 (GraphPad Software, Inc.), and either a Student's
t-test or one-way analysis of variance followed by the Bonferroni
post hoc test. Each experiment was repeated independently at least
two times. P<0.05 was considered to indicate a statistically
significant difference.
Results
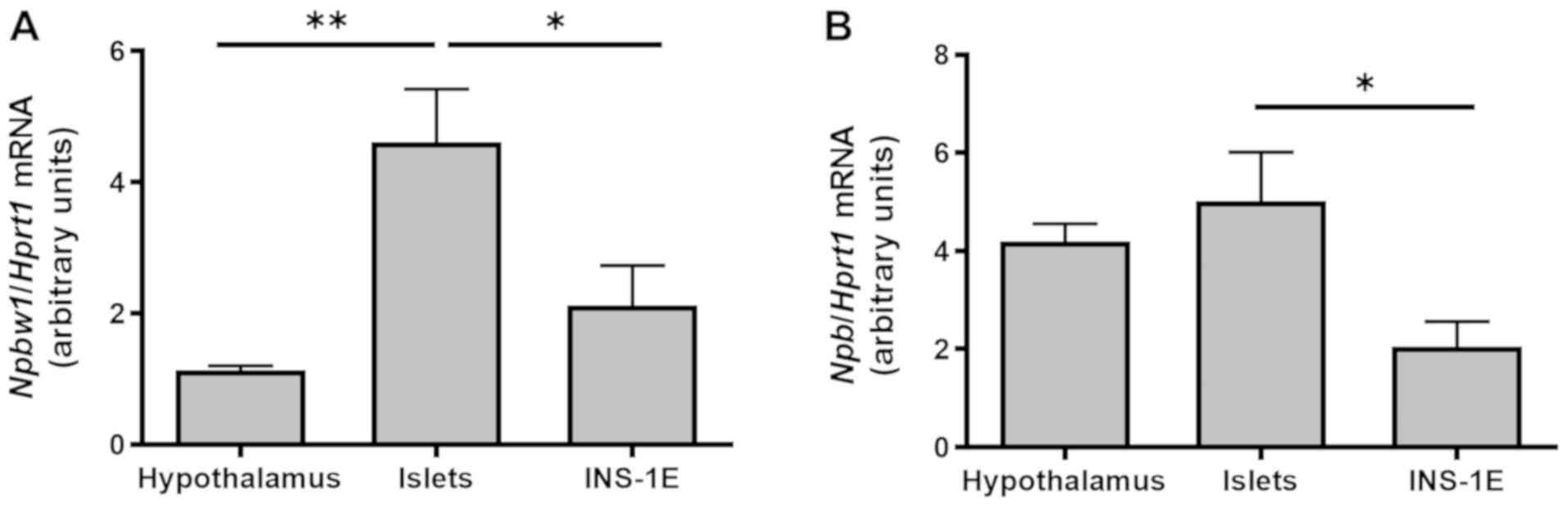
Npbw1 and Npb mRNA are expressed in
INS-1E cells and rat pancreatic islets
Npbw1 and Npb mRNA were expressed in
the hypothalamus (positive control) (4), INS-1E cells and isolated rat
pancreatic islets. The highest level of Npbw1 mRNA
expression was observed in rat pancreatic islets (Fig. 1A), while Npb mRNA expression
levels in the hypothalamus and pancreatic islets were comparable
(Fig. 1B). Expression of Npb mRNA
in INS-1E cells was lower than in pancreatic islets (Fig. 1B, P<0.05).
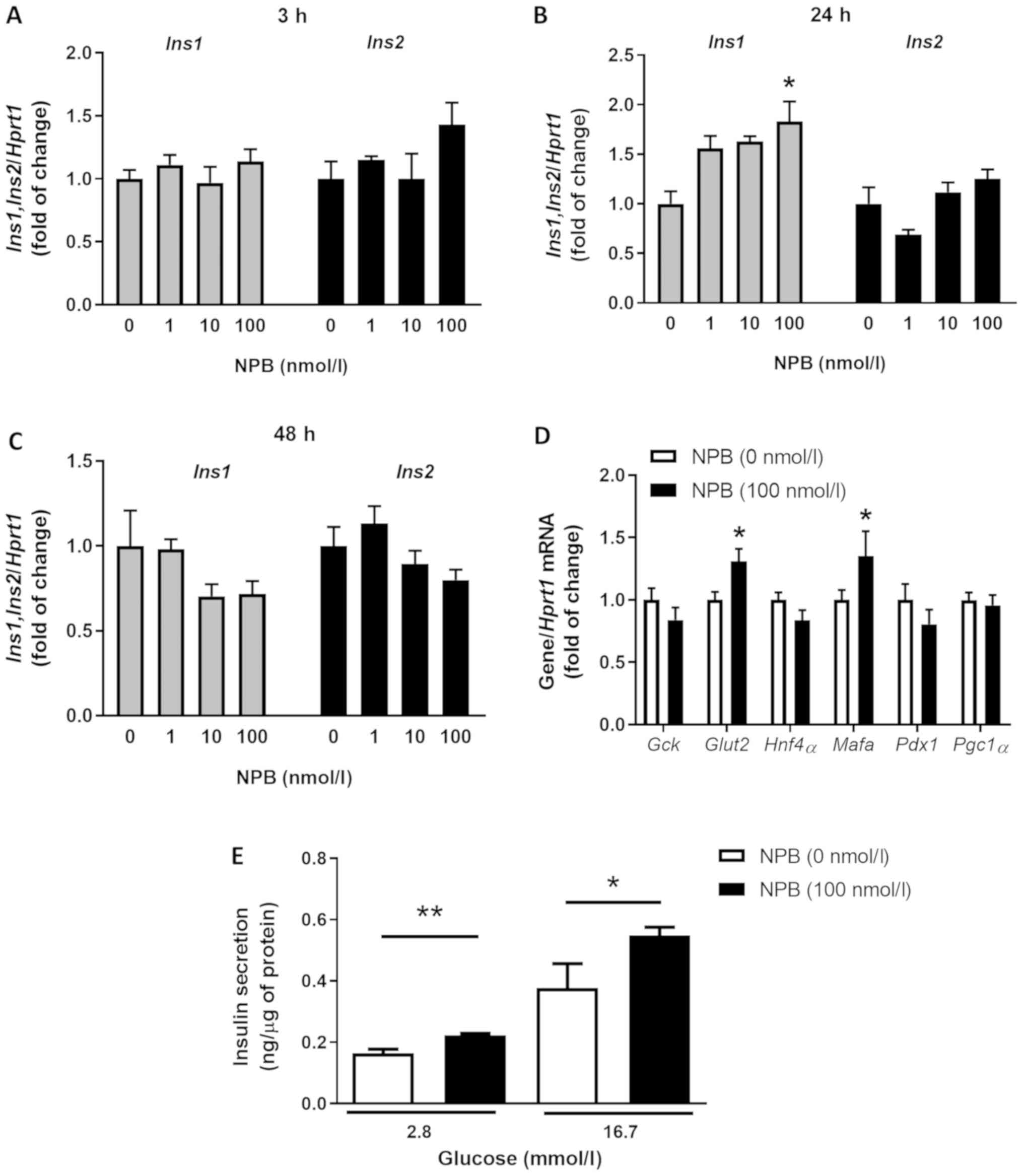
NPB stimulates insulin expression and
secretion in INS-1E cells
NPB at 100 nmol/l increased Ins1 mRNA
expression (in INS-1E cells after 24 h (Fig. 2B, P<0.05). In contrast, NPB at
all tested doses (1, 10 and 100 nmol/l) failed to induce
Ins2 mRNA expression in INS-1E cells after 24 h (Fig. 2B). Furthermore, all NPB doses
failed to affect Ins1 and Ins2 mRNA expression
assessed in cells incubated for 3 or 48 h (Fig. 2A and C).
In addition, cells treated with 100 nmol/l NPB had
increased expression of Mafa and Glut2 mRNA (Fig. 2D, P<0.05). In contrast, NPB had
no effects on mRNA levels of all other tested genes (Fig. 2D). Since 100 nmol/l NPB was the
most efficient dose at increasing insulin mRNA expression, we
evaluated the effect of this NPB dose on insulin exocytosis in
INS-1E cells. NPB increased insulin release at 2.8 and 16.7 mmol/l
glucose (Fig. 2E, P<0.01 and
P<0.05). Overall, these data showed that NPB enhanced insulin
mRNA expression and secretion in INS-1E cells.
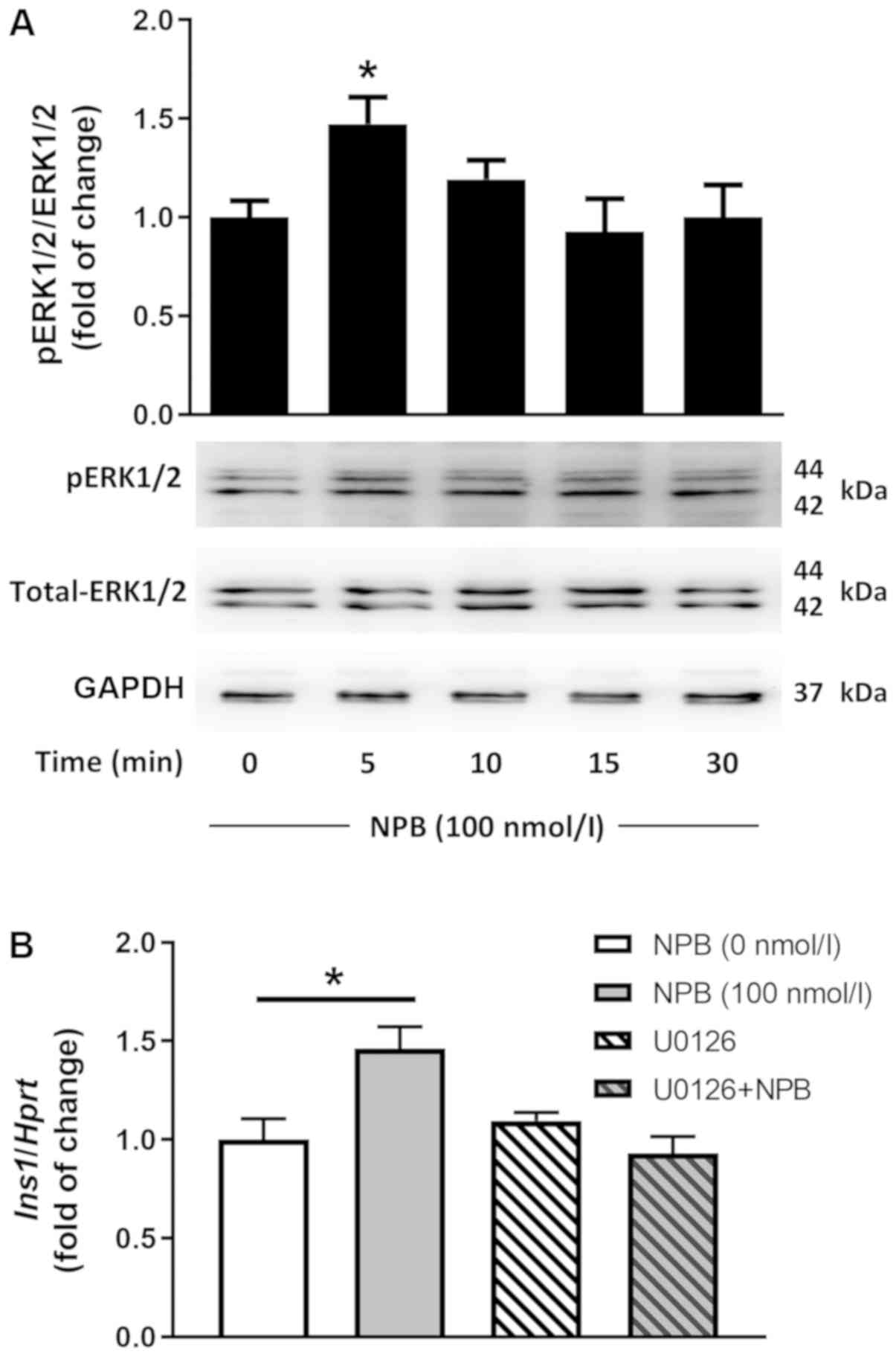
NPB stimulates Ins1 mRNA expression
via ERK1/2-dependent mechanism
ERK1/2 modulates insulin mRNA expression in
pancreatic beta cells (21).
Therefore, we studied the effects of NPB on ERK1/2 phosphorylation.
NPB (100 nmol/l) stimulated ERK1/2 phosphorylation in INS-1E cells
after 5 min (Fig. 3A,
P<0.05).
To study whether ERK1/2 mediates the effects of NPB
on Ins1 mRNA levels, we utilized MEK1/2-dependent ERK1/2
phosphorylation blocker U0126 (22). In the presence of U0126 (10
µmol/l), NPB failed to increase Ins1 mRNA expression
(Fig. 3B). These results show that
NPB stimulates Ins1 mRNA expression via ERK1/2.
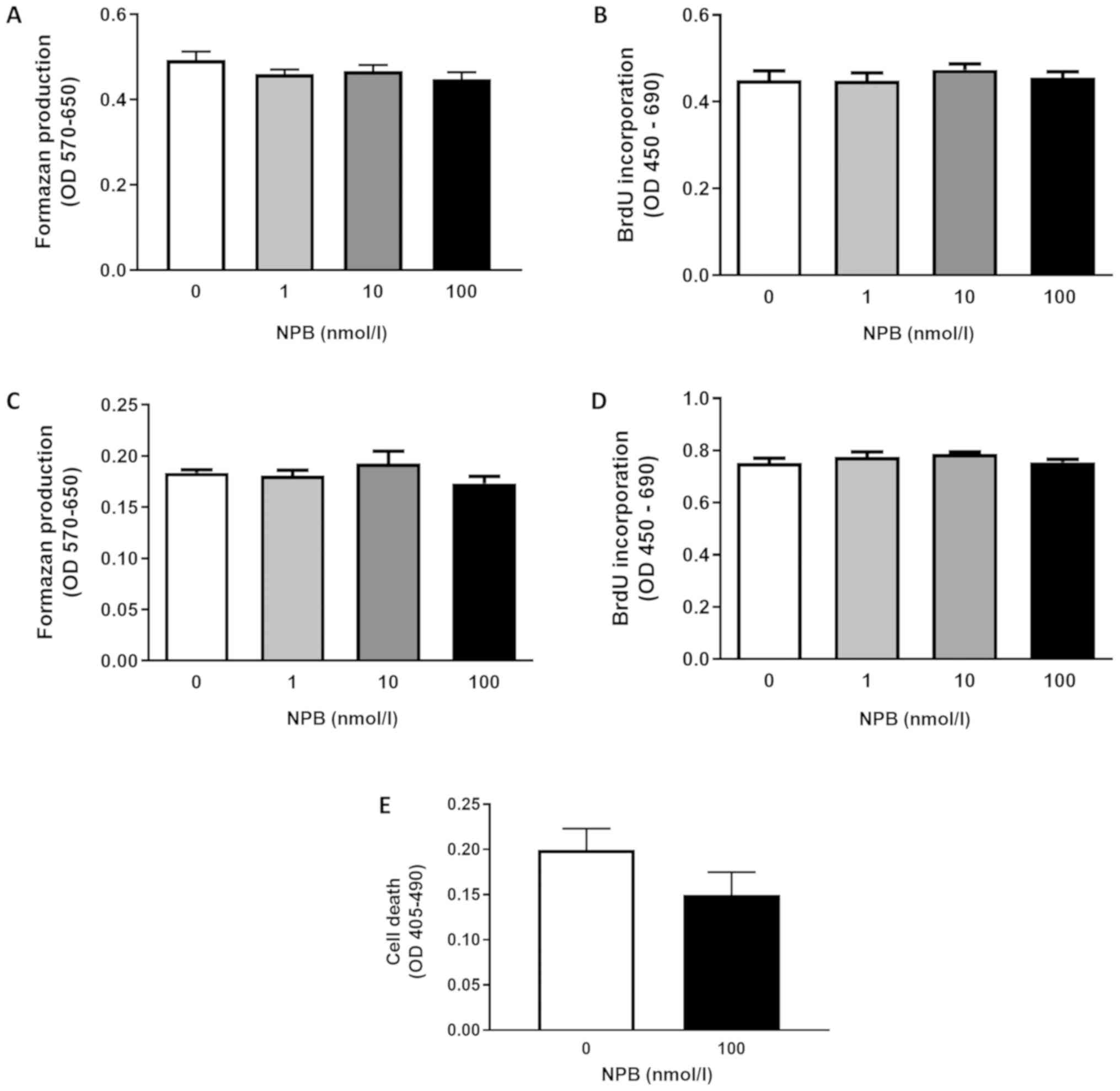
NPB fails to modulate INS-1E growth
and viability of INS-1E cells
As shown in Fig. 4,
NPB at all tested doses (1, 10 and 100 nmol/l) failed to influence
INS-1E cell viability (Fig. 4A and
C) or proliferation (Fig. 4B and
D) after 24 or 48 h. Furthermore, NPB (100 nmol/l) had no
effects on INS-1E cell death assessed after 48 h (Fig. 4E). These results show that NPB is
not involved in controlling INS-1E cell proliferation and
viability.
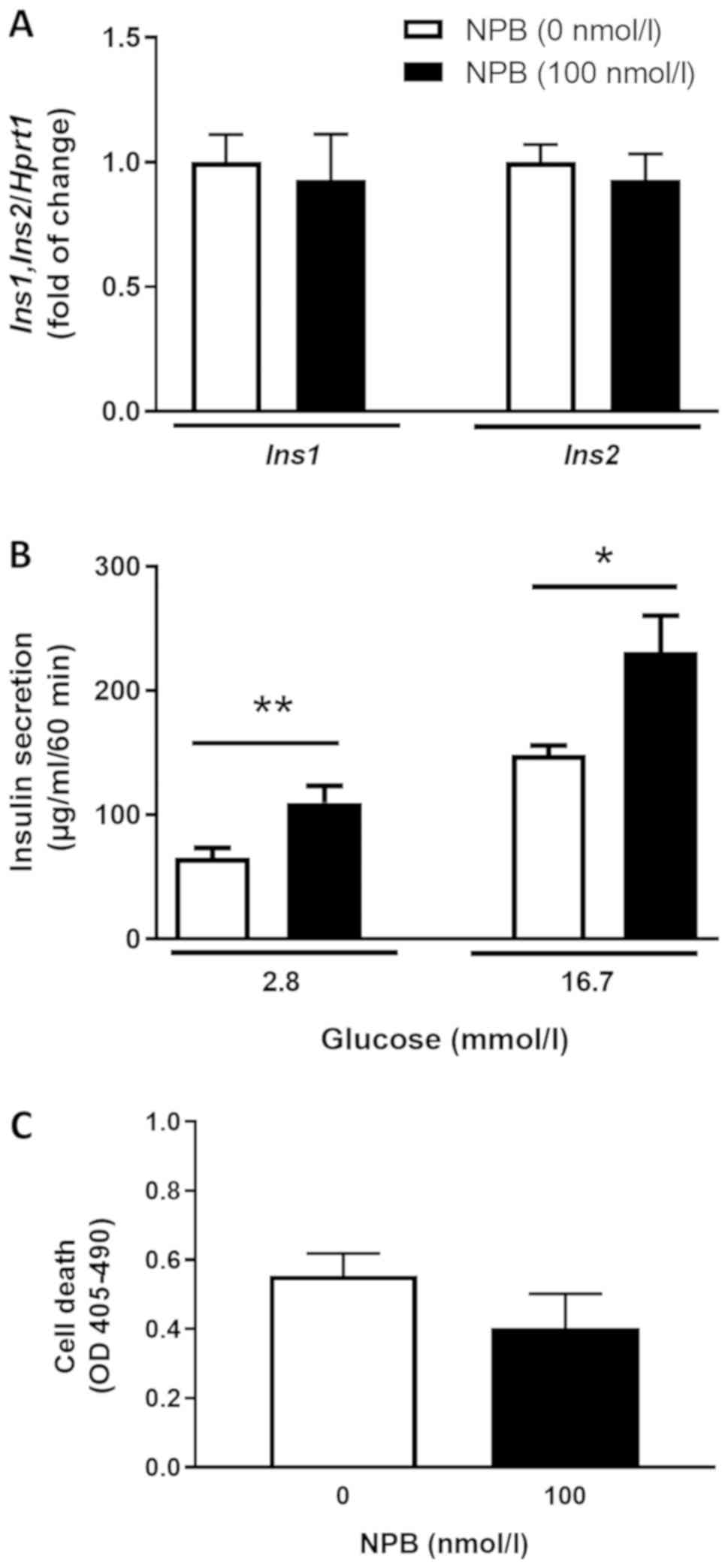
NPB stimulates insulin secretion but
not expression in isolated pancreatic islets
To confirm our findings, we studied the effects of
NPB on Ins1, Ins2 mRNA expression, insulin secretion and
cell death in isolated pancreatic islets. We found that NPB (100
nmol/l) had no effects on insulin mRNA expression in isolated
pancreatic islets after 24 h (Fig.
5A). In contrast, NPB (100 nmol/l) enhanced insulin secretion
from pancreatic islets at 2.8 and 16.7 mmol/l glucose (Fig. 5B, P<0.01 and P<0.05). On the
other hand, 100 nmol/l NPB did not affect cell death in isolated
pancreatic islets (Fig. 5C).
Discussion
In the present study, we report that NPB modulates
insulin secretion and expression in INS-1E cells. Furthermore, we
demonstrate that NPB stimulates insulin secretion in isolated rat
pancreatic islets without affecting insulin mRNA expression and
beta cell death.
First of all, we found that Npb and its
receptor Npbw1 mRNA are expressed in INS-1E cells and
isolated rat pancreatic islets. The presence of NPBW1 and its
ligand in insulin-producing cells is in line with previous data
demonstrating that NPB and Npbw1 mRNA are expressed in rat
pancreatic islets (8). The
presence of NPB in pancreatic islets suggests that this peptide may
modulate endocrine cells functions via a paracrine mechanism.
However, since our data are restricted to mRNA expression alone,
further studies are needed to confirm the presence of NPB on a
protein level. Nevertheless, when discussing the role of the
NPB/NPW system in pancreatic islets it is worth noticing that there
is evidence indicating that another ligand of NPBW1 receptor NPW
peptide has been detected with beta cells in rat pancreatic islet
(23).
Expression and secretion of insulin from pancreatic
beta cells are modulated by a variety of nutritional factors, among
which glucose plays the essential role (24). However, there is emerging evidence
that numerous appetite-controlling peptides significantly
contribute to these processes as well (25–27).
Therefore, we studied the influence of NPB on insulin mRNA
expression. We found that NPB increased Ins1 but not
Ins2 mRNA expression in INS-1E cells exposed to NPB for 24
h. Notably, these effects were not detected in cells exposed to NPB
for 3 or 48 h, which clearly indicates that the effects of NPB are
strictly time-dependent. It cannot be excluded that the lack of an
effect on Ins1 mRNA expression in long-time incubations with
NPB was due to the receptor desensitization which can be caused by
prolonged exposition of GPCR to their ligands (28).
To explore the mechanism by which NPB increases
Ins1 mRNA expression we assessed its effect on the
expression of genes involved in insulin mRNA expression and beta
cell metabolism: Gck, Glu2, Hnf4α, Mafa, Pdx1, Pgc1α
(14,24,29,30).
We found that NPB enhanced Mafa and Glut2 mRNA
expression only in INS-1E cell. Previous data showed that in
pancreatic beta cells the transcription factor Mafa
stimulates insulin mRNA expression in a glucose-dependent manner
(31). Since NPB also stimulated
the expression of the main glucose transporter Glut2 in
rodent beta cells (32), the
intracellular glucose content may increase which, in turn, may lead
to stimulation of Mafa, with concomitant upregulation of
insulin mRNA expression. Previous data have demonstrated that
binding of Mafa to the glucose-responsive A2C1 element of
the insulin gene promoter depends on ERK1/2 activation (33). Intracellular cascades downstream of
NPBW1 are poorly characterized. However, it has been found that NPB
stimulates adrenocortical carcinoma-derived NCI-H295 cells growth
via ERK1/2-dependent mechanism (34). Thus, we examined the effects of NPB
on ERK1/2 phosphorylation. Our data show that NPB stimulates ERK1/2
phosphorylation in INS-1E cells. Furthermore, we found that the
MEK1/2-dependent ERK1/2 phosphorylation blocker U0126 (22) completely blunted the effects of NPB
on Ins1 mRNA expression. Overall, these results suggest that
ERK1/2 activation is required to induce Ins1 mRNA expression
in response to NPB treatment.
An open question is why NPB stimulates Ins1
mRNA expression alone. Notably, there is evidence suggesting the
independent regulation of Ins1 and Ins2 mRNA
expression (35). For example, a
mouse study showed that Ins1 but not Ins2 mRNA
expression is altered by glucose (36). Furthermore, different
transcriptional regulation of Ins1 and Ins2 mRNA
expression was also reported. For example, mice lacking the neuroD
transcription factor had reduced Ins1 but not Ins2
mRNA expression (37). Moreover,
it was shown that Mafa knockout (KO) mice had suppressed
Ins1 but not Ins2 mRNA expression comparing with wild
type animals (38). Thus, NPB may
stimulate expression and/or activity of transcription factors
(e.g., Mafa) which are predominantly involved in controlling
Ins1 but not Ins2 mRNA expression.
In addition, our study demonstrates that NPB
stimulates insulin secretion from INS-1E cells. These effects were
observed at low (2.8 mmol/l) as well as at high (16.7 mol/l)
glucose concentrations. These results are comparable with previous
studies indicating that activation of the NPBW1 receptor leads to
increased insulin secretion in rat islets (23). Dezaki et al (23) reported that NPW stimulates insulin
release from pancreatic islets; however, this effect was detected
at 8.3 mM glucose only and not at 2.8 mM. In this respect, NPB has
a higher potency at activating NPBW1 receptor as compared to NPW
(39). Thus, this may partially
explain the ability of NPB to enhance glucose release even at low
glucose concentrations.
Our study lacks an exploration of the mechanism by
which NPB triggers insulin exocytosis in beta cells. However,
studies on NPW have shown that this peptide stimulates
intracellular calcium influx in rat beta cells via voltage-gated
L-type channels (23). Since both
NPW and NPB interact with the same type of receptor (NPBW1)
(39), this strongly suggests that
NPB may increase insulin secretion by activating the same type of
calcium channels.
Type 1 and type 2 diabetes are characterized by a
loss of beta cells (40).
Identification of new molecular targets able to protect beta cells
from death and promote their replication is of clinical importance
(41). Therefore, we assessed the
effects of NPB on INS-1E cell growth and death. However, we found
that NPB did not influence INS-1E cell growth, viability or death.
The role of NPB receptor signaling in controlling mitogenesis and
cell death is poorly characterized so far. NPB suppresses
proliferation of rat calvaria osteoblast-like cells in vitro
(42). On the other hand, NPB
potentiates growth of rat adrenocortical cells (43). Overall, these results suggest that
the effects of NPB on cell proliferation are cell-specific.
Nevertheless, it should be kept in mind that in our study we used
INS-1E insulinoma cells; therefore, the potential influence of NPB
on primary beta cell replication remains to be studied in the
future.
Finally, to study the relevance of our findings in
more physiological settings, we tested whether NPB is involved in
insulin expression, secretion and cell death in isolated rat
pancreatic islets. We found that NPB increased insulin secretion
without affecting insulin mRNA expression or cell death. Therefore,
NPB may be a physiological modulator of insulin exocytosis alone.
However, it must be noticed that our study is limited to static
incubation of pancreatic islets. Furthermore, pancreatic islets
release other endocrine factors in addition to insulin (44), which in turn may affect insulin
mRNA expression. For example, it has been shown that somatostatin
which is produced in pancreatic delta cells is able to suppress
transcription of the insulin gene (45). In addition, established beta cell
lines display different ion channel expressions, glucose
sensitivity and numerous aspects of cellular physiology that may
differ from those of native beta cells (46,47).
Therefore, our results derived from beta cell lines need to be
interpreted cautiously. Experiments utilizing purified primary beta
cells and/or in vivo experiments should answer the questions
about the role of NPB in primary beta cell physiology.
In conclusion, we found that NPB stimulates insulin
mRNA expression via an ERK1/2-dependent mechanism. Furthermore, we
demonstrated that NPB stimulates insulin secretion in INS-1E cells
and isolated rat pancreatic islets.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Science Center of Poland (grant nos. 2016/23/D/NZ4/03557
and 2016/21/B/NZ9/00943).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MB, MSa and MSk designed the study and wrote the
manuscript. MB, MSa, TW, MJ and MSk performed the experiments. MZS
and KWN interpreted the data, and revised the manuscript. All
authors read and approved the final submitted manuscript.
Ethics approval and consent to
participate
According to the Act on the protection of Animals
used for Scientific or Educational Purposes in Poland adopted on
15th January 2015 and according to earlier regulations, experiments
focused on the analysis of tissues obtained after the death of
animals that were not treated with any experimental procedure do
not require permission of the Local Ethical Commission for
investigation on Animals. Therefore, the requirement for ethical
approval in the present study was waived. All applicable
international, national and/or institutional guidelines for the
care and use of animals were followed.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
BrdU
|
5-bromo-2′-deoxyuridine
|
|
ERK1/2
|
extracellular signal-regulated protein
kinases 1 and 2
|
|
GAPDH
|
glyceraldehyde 3-phosphate
dehydrogenase
|
|
Gck
|
glucokinase
|
|
Glut2
|
glucose transporter 2
|
|
GPCR
|
G protein-coupled receptor
|
|
Hnf4α
|
hepatocyte nuclear factor 4α
|
|
Hprt
|
hypoxanthine-guanine
phosphoribosyltransferase
|
|
Mafa
|
β-cell transcriptional factor
musculoaponeurotic fibrosarcoma oncogene family A
|
|
MTT
|
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
|
|
NPB
|
neuropeptide B
|
|
NPBW1
|
neuropeptides B/W receptor 1, GPR7
|
|
NPBW2
|
neuropeptides B/W receptor 2, GPR8
|
|
Pdx1
|
pancreatic and duodenal homeobox 1
|
|
Pgc1α
|
peroxisome proliferator-activated
receptor γ coactivator 1-α
|
References
|
1
|
Tanaka H, Yoshida T, Miyamoto N, Motoike
T, Kurosu H, Shibata K, Yamanaka A, Williams SC, Richardson JA,
Tsujino N, et al: Characterization of a family of endogenous
neuropeptide ligands for the G protein-coupled receptors GPR7 and
GPR8. Proc Natl Acad Sci USA. 100:6251–6256. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fujii R, Yoshida H, Fukusumi S, Habata Y,
Hosoya M, Kawamata Y, Yano T, Hinuma S, Kitada C, Asami T, et al:
Identification of a neuropeptide modified with bromine as an
endogenous ligand for GPR7. J Biol Chem. 277:34010–34016. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lee DK, Nguyen T, Porter CA, Cheng R,
George SR and O'Dowd BF: Two related G protein-coupled receptors:
The distribution of GPR7 in rat brain and the absence of GPR8 in
rodents. Brain Res Mol Brain Res. 71:96–103. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chottova Dvorakova M: Distribution and
function of neuropeptides W/B signaling system. Front Physiol.
9:9812018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yamamoto T, Saito O, Shono K and Tanabe S:
Anti-hyperalgesic effects of intrathecally administered
neuropeptide W-23, and neuropeptide B, in tests of inflammatory
pain in rats. Brain Res. 1045:97–106. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Aikawa S, Ishii M, Yanagisawa M,
Sakakibara Y and Sakurai T: Effect of neuropeptide B on feeding
behavior is influenced by endogenous corticotropin-releasing factor
activities. Regul Pept. 151:147–152. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Uchio N, Doi M, Matsuo M, Yamazaki F,
Mizoro Y, Hondo M, Sakurai T and Okamura H: Circadian
characteristics of mice depleted with GPR7. Biomed Res. 30:357–364.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hochol A, Belloni AS, Rucinski M,
Ziolkowska A, Di Liddo R, Nussdorfer GG and Malendowicz LK:
Expression of neuropeptides B and W and their receptors in
endocrine glands of the rat. Int J Mol Med. 18:1101–1106.
2006.PubMed/NCBI
|
|
9
|
Ishii M, Fei H and Friedman JM: Targeted
disruption of GPR7, the endogenous receptor for neuropeptides B and
W, leads to metabolic defects and adult-onset obesity. Proc Natl
Acad Sci USA. 100:10540–10545. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kelly MA, Beuckmann CT, Williams SC,
Sinton CM, Motoike T, Richardson JA, Hammer RE, Garry MG and
Yanagisawa M: Neuropeptide B-deficient mice demonstrate
hyperalgesia in response to inflammatory pain. Proc Natl Acad Sci
USA. 102:9942–9947. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Skrzypski M, Pruszynska-Oszmalek E,
Rucinski M, Szczepankiewicz D, Sassek M, Wojciechowicz T, Kaczmarek
P, Kołodziejski PA, Strowski MZ, Malendowicz LK and Nowak KW:
Neuropeptide B and W regulate leptin and resistin secretion, and
stimulate lipolysis in isolated rat adipocytes. Regul Pept.
176:51–56. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Grzelak T, Tyszkiewicz-Nwafor M,
Dutkiewicz A, Mikulska AA, Dmitrzak-Weglarz M, Slopien A, Czyzewska
K and Paszynska E: Neuropeptide B and vaspin as new biomarkers in
anorexia nervosa. Biomed Res Int. 2018:97275092018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Grzelak T, Wedrychowicz A, Grupinska J,
Grupinska J, Pelczynska M, Sperling M, Mikulska AA, Naughton V and
Czyzewska K: Neuropeptide B and neuropeptide W as new serum
predictors of nutritional status and of clinical outcomes in
pediatric patients with type 1 diabetes mellitus treated with the
use of pens or insulin pumps. Arch Med Scie. 15:619–631. 2019.
View Article : Google Scholar
|
|
14
|
Fu Z, Gilbert ER and Liu D: Regulation of
insulin synthesis and secretion and pancreatic Beta-cell
dysfunction in diabetes. Curr Diabetes Rev. 9:25–53. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Klöppel G, Löhr M, Habich K, Oberholzer M
and Heitz PU: Islet pathology and the pathogenesis of type 1 and
type 2 diabetes mellitus revisited. Surv Synth Pathol Res.
4:110–125. 1985.PubMed/NCBI
|
|
16
|
Porte D Jr and Kahn SE: Beta-cell
dysfunction and failure in type 2 diabetes: Potential mechanisms.
Diabetes. 50 (Suppl 1):S160–S163. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sakurai T: NPBWR1 and NPBWR2: Implications
in energy homeostasis, pain, and emotion. Front Endocrinol
(Lausanne). 4:232013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Merglen A, Theander S, Rubi B, Chaffard G,
Wollheim CB and Maechler P: Glucose sensitivity and
metabolism-secretion coupling studied during two-year continuous
culture in INS-1E insulinoma cells. Endocrinology. 145:667–678.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Billert M, Skrzypski M, Sassek M,
Szczepankiewicz D, Wojciechowicz T, Mergler S, Strowski MZ and
Nowak KW: TRPV4 regulates insulin mRNA expression and INS-1E cell
death via ERK1/2 and NO-dependent mechanisms. Cell Signal.
35:242–249. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mazzocchi G, Rebuffat P, Ziolkowska A,
Rossi GP, Malendowicz LK and Nussdorfer GG: G protein receptors 7
and 8 are expressed in human adrenocortical cells, and their
endogenous ligands neuropeptides B and w enhance cortisol secretion
by activating adenylate cyclase- and phospholipase C-dependent
signaling cascades. J Clin Endocrinol Metab. 90:3466–3471. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Khoo S, Gibson TB, Arnette D, Lawrence M,
January B, McGlynn K, Vanderbilt CA, Griffen SC, German MS and Cobb
MH: MAP kinases and their roles in pancreatic beta-cells. Cell
Biochem Biophys 40 (3 Suppl). S191–S200. 2004. View Article : Google Scholar
|
|
22
|
Favata MF, Horiuchi KY, Manos EJ, Daulerio
AJ, Stradley DA, Feeser WS, Van Dyk DE, Pitts WJ, Earl RA, Hobbs F,
et al: Identification of a novel inhibitor of mitogen-activated
protein kinase kinase. J Biol Chem. 273:18623–18632. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dezaki K, Kageyama H, Seki M, Shioda S and
Yada T: Neuropeptide W in the rat pancreas: Potentiation of
glucose-induced insulin release and Ca2+ influx through
L-type Ca2+ channels in beta-cells and localization in
islets. Regul Pept. 145:153–158. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Andrali SS, Sampley ML, Vanderford NL and
Ozcan S: Glucose regulation of insulin gene expression in
pancreatic beta-cells. Biochem J. 415:1–10. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Seufert J, Kieffer TJ and Habener JF:
Leptin inhibits insulin gene transcription and reverses
hyperinsulinemia in leptin-deficient ob/ob mice. Proc Natl Acad Sci
USA. 96:674–679. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Colombo M, Gregersen S, Xiao J and
Hermansen K: Effects of ghrelin and other neuropeptides (CART, MCH,
orexin A and B, and GLP-1) on the release of insulin from isolated
rat islets. Pancreas. 27:161–166. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Skrzypski M, Khajavi N, Mergler S, Billert
M, Szczepankiewicz D, Wojciechowicz T, Nowak KW and Strowski MZ:
Orexin A modulates INS-1E cell proliferation and insulin secretion
via extracellular signal-regulated kinase and transient receptor
potential channels. J Physiol Pharmacol. 67:643–652.
2016.PubMed/NCBI
|
|
28
|
Böhm SK, Grady EF and Bunnett NW:
Regulatory mechanisms that modulate signalling by G-protein-coupled
receptors. Biochem J. 322:1–18. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yoon JC, Xu G, Deeney JT, Yang SN, Rhee J,
Puigserver P, Levens AR, Yang R, Zhang CY, Lowell BB, et al:
Suppression of beta cell energy metabolism and insulin release by
PGC-1alpha. Dev Cell. 5:73–83. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang H, Maechler P, Antinozzi PA,
Hagenfeldt KA and Wollheim CB: Hepatocyte nuclear factor 4alpha
regulates the expression of pancreatic beta-cell genes implicated
in glucose metabolism and nutrient-induced insulin secretion. J
Biol Chem. 275:35953–35959. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kataoka K, Han SI, Shioda S, Hirai M,
Nishizawa M and Handa H: MafA is a glucose-regulated and pancreatic
beta-cell-specific transcriptional activator for the insulin gene.
J Biol Chem. 277:49903–49910. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
De Vos A, Heimberg H, Quartier E, Huypens
P, Bouwens L, Pipeleers D and Schuit F: Human and rat beta cells
differ in glucose transporter but not in glucokinase gene
expression. J Clin Invest. 96:2489–2495. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lawrence MC, McGlynn K, Park BH and Cobb
MH: ERK1/2-dependent activation of transcription factors required
for acute and chronic effects of glucose on the insulin gene
promoter. J Biol Chem. 280:26751–26759. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Andreis PG, Rucinski M, Neri G, Conconi
MT, Petrelli L, Parnigotto PP, Malendowicz LK and Nussdorfer GG:
Neuropeptides B and W enhance the growth of human adrenocortical
carcinoma-derived NCI-H295 cells by exerting MAPK p42/p44-mediated
proliferogenic and antiapoptotic effects. Int J Mol Med.
16:1021–1028. 2005.PubMed/NCBI
|
|
35
|
Deltour L, Leduque P, Blume N, Madsen O,
Dubois P, Jami J and Bucchini D: Differential expression of the two
nonallelic proinsulin genes in the developing mouse embryo. Proc
Natl Acad Sci USA. 90:527–531. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Roderigo-Milne H, Hauge-Evans AC, Persaud
SJ and Jones PM: Differential expression of insulin genes 1 and 2
in MIN6 cells and pseudoislets. Biochem Biophys Res Commun.
296:589–595. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gu C, Stein GH, Pan N, Goebbels S,
Hörnberg H, Nave KA, Herrera P, White P, Kaestner KH, Sussel L and
Lee JE: Pancreatic beta cells require NeuroD to achieve and
maintain functional maturity. Cell Metab. 11:298–310. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Nishimura W, Takahashi S and Yasuda K:
MafA is critical for maintenance of the mature beta cell phenotype
in mice. Diabetologia. 58:566–574. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Brezillon S, Lannoy V, Franssen JD, Le
Poul E, Dupriez V, Lucchetti J, Detheux M and Parmentier M:
Identification of natural ligands for the orphan G protein-coupled
receptors GPR7 and GPR8. J Biol Chem. 278:776–783. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cnop M, Welsh N, Jonas JC, Jorns A, Lenzen
S and Eizirik DL: Mechanisms of pancreatic beta-cell death in type
1 and type 2 diabetes: Many differences, few similarities.
Diabetes. 54 (Suppl 2):S97–S107. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen C, Cohrs CM, Stertmann J, Bozsak R
and Speier S: Human beta cell mass and function in diabetes: Recent
advances in knowledge and technologies to understand disease
pathogenesis. Mol Metab. 6:943–957. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ziolkowska A, Rucinski M, Tyczewska M and
Malendowicz LK: Neuropeptide B (NPB) and neuropeptide W (NPW)
system in cultured rat calvarial osteoblast-like (ROB) cells: NPW
and NPB inhibit proliferative activity of ROB cells. Int J Mol Med.
24:781–787. 2009.PubMed/NCBI
|
|
43
|
Hochol A, Albertin G, Nussdorfer GG,
Spinazzi R, Ziolkowska A, Rucinski M and Malendowicz LK: Effects of
neuropeptides B and W on the secretion and growth of rat
adrenocortical cells. Int J Mol Med. 14:843–847. 2004.PubMed/NCBI
|
|
44
|
Steiner DJ, Kim A, Miller K and Hara M:
Pancreatic islet plasticity: Interspecies comparison of islet
architecture and composition. Islets. 2:135–145. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Redmon JB, Towle HC and Robertson RP:
Regulation of human insulin gene transcription by glucose,
epinephrine, and somatostatin. Diabetes. 43:546–551. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hiriart M and Aguilar-Bryan L: Channel
regulation of glucose sensing in the pancreatic beta-cell. Am J
Physiol Endocrinol Metab. 295:E1298–E1306. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Skrzypski M, Billert M, Mergler S, Khajavi
N, Nowak KW and Strowski MZ: Role of TRPV channels in regulating
various pancreatic β-cell functions: Lessons from in vitro studies.
Biosci Trends. 11:9–15. 2017. View Article : Google Scholar : PubMed/NCBI
|