Introduction
Melanoma, also known as malignant melanoma, arises
from the pigment-producing cells of the deeper layers of the skin,
attributed to skin lesions (1).
Melanocytes are abundant in the skin (2). Melanogenesis is complex regulated by
a variety of factors that interact with each other in a hormonal,
autogenous, para-endocrine or endocrine manner, through
receptor-dependent and independent mechanisms of activation
(3). Nearly 20,000 cases of
cutaneous malignant melanoma are diagnosed in China every year
(4). Increasing numbers of studies
have confirmed that ultraviolet (UV) radiation is the main risk
factor for the development of melanoma. Cumulative UV exposure
leads to UV-induced DNA damage, oxidative stress and skin
inflammation (5–8); in particular, UV leads to DNA damage
and immunosuppression, both of which cause melanoma to develop and
are involved in the pathogenesis of melanoma (9). At the same time, the interaction
between melanoma cells and the microenvironment of melanocytes in
the skin epidermis is also considered to affect melanin damage
(10,11). In addition, an immunosuppressive
field is produced in the intermediate stage of melanogenesis, which
weakens the effect of any immunotherapy for melanoma. Therefore,
uncontrolled melanogenesis, together with melanin, leads to
decreased efficacy of radiotherapy, chemotherapy, phototherapy and
immunotherapy (12). In view of
the above, traditional treatment methods include surgical
treatment, radiotherapy and chemotherapy. These treatment
strategies are often self-limiting, achieve far from satisfactory
results, and may even lead to an increase in the burden of global
public health (13,14). Therefore, the urgent task is to
find an effective drug, improve the level of comprehensive
treatment, reduce the side effects of melanoma and reduce the
mortality rate of melanoma.
Chan Su, which is obtained from the skin and parotid
venom glands of the toad (15),
has been traditionally used for the treatment of a variety of
clinical diseases in China. Bufadienolides, including
gamabufotalin, arenobufagin, telocinobufagin, bufalin and other
ingredients, are the major pharmacological constituents of Chan Su,
which is frequently used in the clinic for the treatment of cancer,
including hepatoma, gallbladder carcinoma and lung cancer (16). In previously published studies,
bufadienolide compounds have been demonstrated to induce arrest of
the tumor cell cycle and apoptosis (17,18).
In addition, bufadienolides were revealed to markedly inhibit
proliferation and induce autophagy in liver cancer cells (19), and alterations in the expression
levels of microtubule-associated proteins 1A/1B light chain 3-II
(LC3-II), p62, Bax, Bcl-2, cyclin D1 and caspase-3 are involved in
the underlying mechanism (20).
It has been previously reported in the literature
that the AKT pathway has an important role in promoting cell
apoptosis and autophagy (21).
Based on these previous findings, the present study aimed to
explore whether bufadienolides are able to impair the viability of
melanoma cells and to increase the levels of apoptosis and
autophagy via inhibition of the AKT pathway.
Materials and methods
Reagents and antibody
The bufadienolides were a gift from Dr Jiang (Macau
University of Science and Technology). Preparation of bufotoxin and
bufogenin fractions from toad venom and toad skin (200 g; Xiangshui
Bolin Pharmaceutical & Chemical Co., Ltd.) was ground into
powder and ultrasonically extracted with methanol five times, as
previously described (22). The
methanol solution was filtered, combined and evaporated at reduced
pressure to obtain the total extract (138 g bufadienolides; purity
>97%), which was used for this study. The chemical profile and
structure of bufadienolides has been presented in a previous study
(22). Bufadienolides were
dissolved in anhydrous DMSO at a concentration of 5,000 g/l, for
use as a stock solution. The Annexin V-FITC & PI Apoptosis
Detection kit (cat. no. BD556547) was purchased from BD
Biosciences. Anti-Bax (cat. no. 2772), anti-Bcl-2 (cat. no. 15071),
anti-caspase-3 (cat. no. 9662), anti-p53 (cat. no. sc-126),
anti-LC3-II (cat. no. 2775), anti-p62 (cat. no. 5114), anti-AKT
(cat. no. 9272), anti-mTOR (cat. no. 2972), anti-phosphorylated
(p)-AKT (Ser473; cat. no. 4060), anti-p70S6K (cat. no. 9202),
anti-p-p70S6K (cat. no. 9204), anti-glycogen synthase kinase
(GSK)-3β (cat. no. 9325), anti-p-GSK-3β (cat. no. (Ser9)9323) and
anti-cyclinD1 (cat. no. 2922) were purchased from Cell Signaling
Technology, Inc. GAPDH (cat. no. AP0066) antibody was purchased
from Bioworld Technology, Inc. Horseradish peroxidase (HRP)-labeled
anti-mouse IgG (cat. no. TA130001) or anti-rabbit IgG (cat. no.
TA130015) were obtained from OriGene Technologies, Inc.
Cell culture and treatment
Melanoma A-375 cells, obtained from the American
Type Culture Collection (cat. no. CRL-1619), were incubated in 10%
FBS (Ausbian) and Gibco® DMEM (Thermo Fisher Scientific,
Inc.), and subsequently maintained at 37°C in an atmosphere of 5%
CO2. The bufadienolides were dissolved in anhydrous
DMSO, and diluted with fresh medium to achieve the desired
concentrations (see below).
Cell morphology
Cells were seeded at 2×104/100 µl per
well in 500 µl DMEM in a 24-well plate and incubated for 12 h.
Subsequently, DMEM medium alone (negative control) and media
containing bufadienolides at various concentrations (0, 0.001,
0.01, 0.1, 1 mg/l) were incubated for 0, 24, 48, 72 or 96 h. The
cell morphology change was visualized and photographed under an
inverted phase contrast microscope at ×200 magnification.
Cell Counting Kit-8 (CCK-8) assay
A-375 cells were seeded in 96-well plates at a
density of 2×104/100 µl containing 0, 0.001, 0.01, 0.1
and 1 mg/l bufadienolides, and subsequently cultured for 0, 24, 48,
72 or 96 h. As a negative control, cells were incubated in the
absence of bufadienolides. In order to measure cell viability, 10
µl CCK-8 reagent (Dojindo Molecular Technologies, Inc.) was added
to each well and incubated for 2 h at 37°C in a tissue culture
hood. The absorbance [optical density (OD)] at 490 nm was measured
using a microplate reader.
Annexin V/propidium iodide (PI)
staining assay
An Annexin V-FITC/PI kit was used to assess cell
apoptosis. After treating with bufadienolides (0, 0.001, 0.01, 0.1
and 1 mg/l) for 48 h, A-375 cells were collected, and the harvested
cells were diluted with PBS to a concentration of
5×105−1×106 cells/ml, centrifuged at 4°C and
7,000 × g for 10 min, and resuspended in 200 µl 1X binding buffer.
Subsequently, Annexin V-FITC (10 µl) and PI (5 µl) were added to
each sample. The samples were incubated at room temperature for 15
min, and examined immediately on a flow cytometer (BD Biosciences),
according to the manufacturer's protocol. A BD FACSCanto running BD
CellQuest™ software version 3.3 (BD Biosciences) was used to
perform flow cytometric analysis. Early apoptosis was defined by
Annexin V+/PI-staining (Q4) and late apoptosis was defined by
Annexin V+/PI+ staining (Q2).
Gel electrophoresis and western blot
analysis
The expression levels of Bax, Bcl-2, caspase-3, p53,
LC3-II, p62, AKT, p-AKT, mTOR, p70-S6K1, p-p70-S6K1, GSK-3β,
p-GSK-3β and cyclin D1 in cells were determined using western blot
analysis. A-375 cells were placed in 6-well cell culture plates at
a concentration of 2×105 cells/well, and incubated at
37°C for 24 h. The cells were subsequently treated with
bufadienolides at the aforementioned concentrations for 48 h.
Subsequently, A-375 cells were lysed using RIPA lysis buffer
(Beijing Solarbio Science & Technology Co., Ltd.) on ice for 30
min. The protein concentration was determined using a bicinchoninic
acid protein assay kit (Beyotime Institute of Biotechnology). An
equal amount of total protein (20 µg) was loaded in each lane.
Proteins were separated by 10% SDS-PAGE, the proteins were
transferred onto PVDF membranes (EMD Millipore). The membranes were
blocked with 5% skimmed milk powder solution at room temperature
for 1 h in order to reduce the non-specific background. The blotted
membranes were incubated with anti-Bax, anti-GAPDH, anti-Bcl-2,
anti-caspase-3, anti-p53, anti-LC3-II, anti-p62, anti-AKT,
anti-mTOR, anti-p-AKT, anti-p70S6K, anti-p-p70S6K, anti-GSK-3β,
anti-p-GSK-3β and anti-cyclin D1 (all 1:1,000) at 4°C overnight in
the fridge. Subsequently, the membrane was washed three times with
1X TBS with Tween-20 (TBST), and HRP-labeled anti-mouse IgG or
anti-rabbit IgG were used as secondary antibodies (1:10,000) for 2
h at room temperature. Membranes were subsequently washed again
with 1X TBST buffer, and the protein signals were detected with an
ECL kit (EMD Millipore). Protein expression levels were quantified
by using a Bio-Rad Image station and its built-in software (version
6.0; Bio-Rad Laboratories, Inc.), and are shown as the
densitometric ratio of the target protein to the input control,
GAPDH.
Statistical analysis
Each experiment was repeated three times. All the
presented data are shown as the mean ± standard deviation.
Statistical analyses were performed using one-way ANOVA followed by
the Bonferroni post-hoc test in GraphPad Prism software (version
6.0; GraphPad Software, Inc.). P<0.05 was considered to indicate
a statistically significant difference.
Results
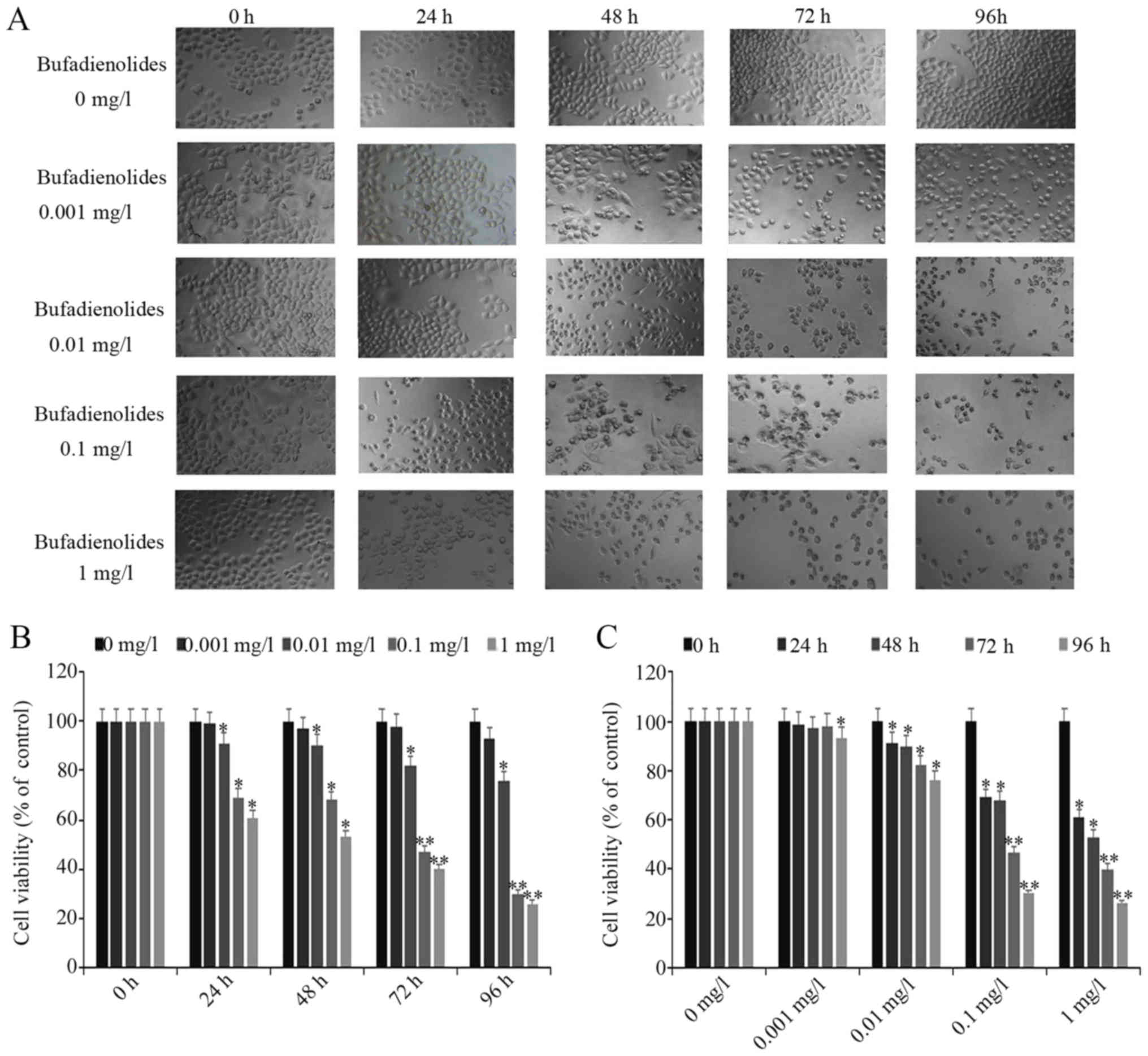
Bufadienolides inhibit the viability
of the A-375 melanoma cells
The A-375 cells were treated with the various
concentrations of bufadienolides for 24, 48, 72 and 96 h. The
IC50 values at 24, 48, 72 and 96 h were 2.56, 1.27, 0.24
and 0.04 mg/l (Table I). The cell
morphology following bufadienolide treatment was observed. From the
inverted phase contrast microscopy (Fig. 1A), it was found that following
bufadienolide treatment, the A-375 cells appeared abnormal with a
rounded shape; some cells had shrunk and had ruptured membranes,
compared with those in the control group. Additionally, the
proportion of normal and living cells decreased with an increasing
bufadienolide concentration. The results of the CCK-8 assay
revealed that at 24, 48, 72 and 96 h, bufadienolide treatment led
to a decrease in the viability of the A-375 cells in a
time-dependent manner compared with the negative control (Fig. 1B). Furthermore, when the A-375
cells were treated with bufadienolides at different doses, their
viability was inhibited in a dose-dependent manner (Fig. 1C). Therefore, these findings
confirmed that bufadienolides inhibited the viability of A-375
cells in a dose- and time-dependent manner.
 | Table I.IC50 values of
bufadienolides in A375 cells at different time points. |
Table I.
IC50 values of
bufadienolides in A375 cells at different time points.
| Time | IC50
value | Mean ± SD |
|---|
| 24 h | 2.56 mg/l | 2.56±0.70 |
| 48 h | 1.27 mg/l | 1.27±0.13 |
| 72 h | 0.24 mg/l | 0.24±0.54 |
| 96 h | 0.04 mg/l | 0.04±0.01 |
Bufadienolides regulate
apoptosis-associated protein levels and promote apoptosis in A-375
cells
Subsequently, the present study assessed whether the
bufadienolides were able to induce apoptosis in A-375 cells. It was
identified that the increases in the percentages of cells in the
early and late apoptotic phases were observed with an increasing
concentration of bufadienolides (Fig.
2A). The cell apoptosis rates in the bufadienolide-treated
cells (0, 0.001, 0.01, 0.1 and 1 mg/l) were 1.40±0.26, 7.30±1.67,
17.20±0.38, 23.40±1.80 and 28.16±0.90% (Fig. 2B). In addition, based on the
results of the western blots shown, it was observed that the
expression levels of Bcl-2 and caspase-3 were reduced in a
dose-dependent manner. However, conversely, the expression levels
of Bax and p53 were increased in A-375 cells following treatment
with bufadienolides (Fig. 2C). It
was also observed that higher concentrations of the bufadienolides
exerted more marked effects; in particular, the protein expression
levels of Bax and p53 were appreciably enhanced (Fig. 2D). These observations suggested
that bufadienolides are able to induce and promote apoptosis in
A-375 cells.
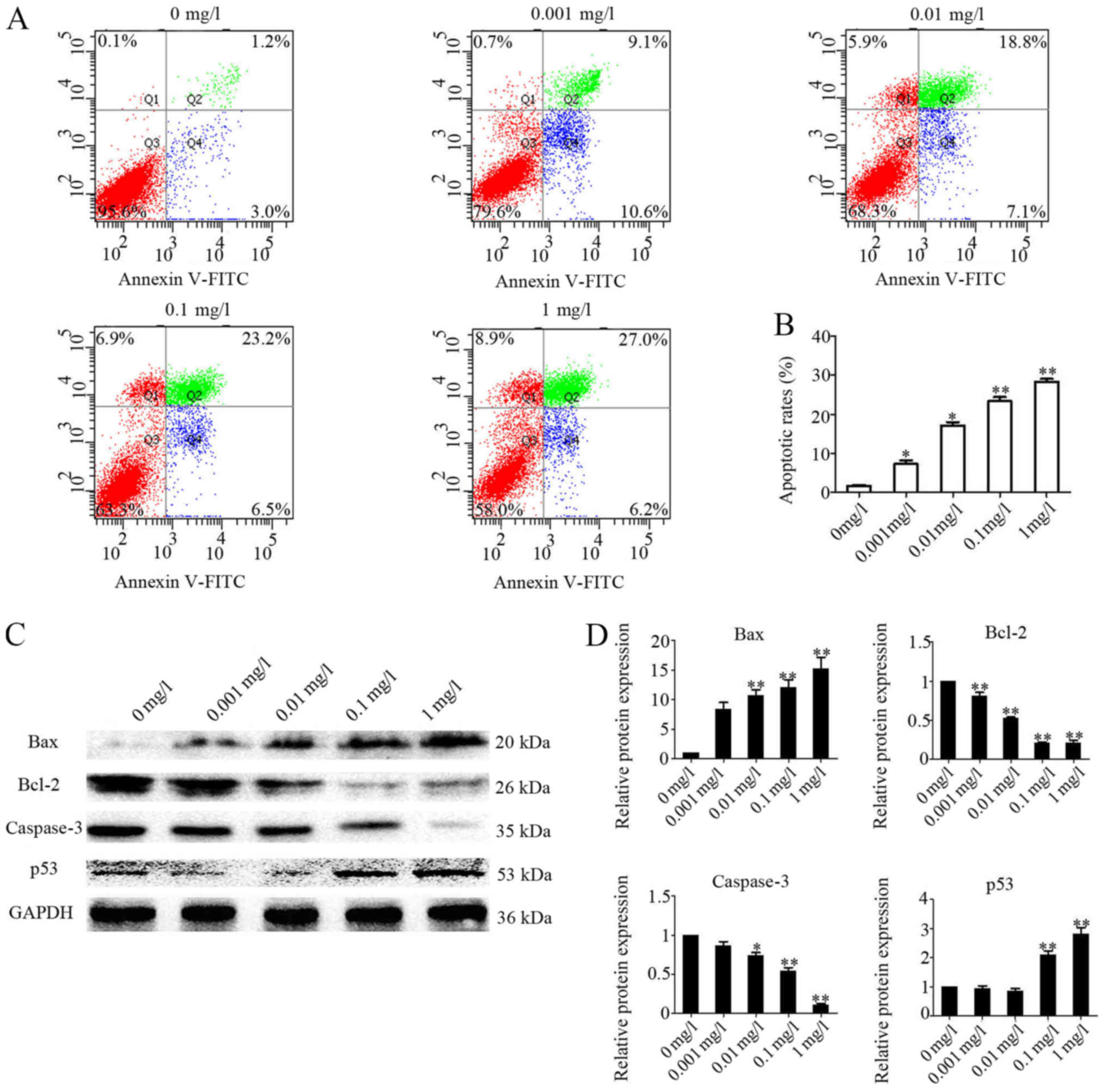 | Figure 2.Expression of apoptosis-associated
proteins in A-375 cells treated with or without bufadienolides.
A-375 cells were treated with bufadienolides (0, 0.001, 0.01, 0.1
and 1 mg/1) for 48 h in 6-well plates (2×105 cell/ml).
(A) The effects of bufadienolides on the induction of A-375 cell
apoptosis were analyzed by flow cytometric analysis. (B) The
apoptosis rate was statistically analyzed. (C) Western blotting
images for Bax, Bcl-2, caspase-3, p53 and GAPDH in A-375 cells
treated with bufadienolides. (D) Quantification of the levels of
Bax, Bcl-2, caspase-3 and p53 was performed, and the results were
normalized relative to the expression level of GAPDH. Each protein
was statistically analyzed, and the results are shown in the right
panel. *P<0.05 and **P<0.01 vs. respective negative control
(0 mg/l bufadienolides). |
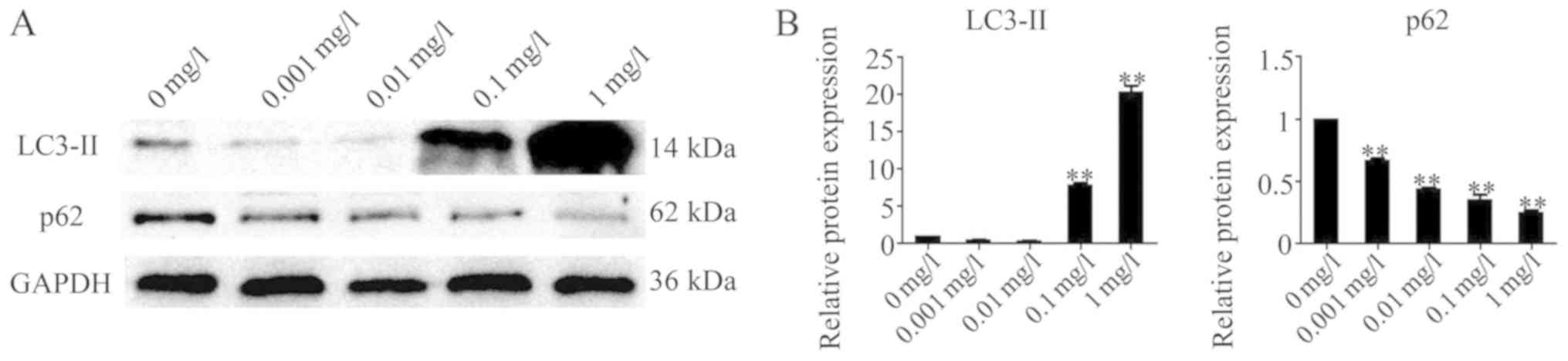
Bufadienolides regulate
autophagy-associated proteins in A-375 cells
To investigate whether the inhibition of viability
of the A-375 cells by bufadienolides was mediated via the
potentiation of autophagy, A-375 cells were treated with
bufadienolides at the specified concentrations (0.001, 0.01, 0.1
and 1 mg/l) for 24 h, and the level of autophagy was determined by
western blot analysis. These experiments revealed that treatment
with bufadienolides affected the protein expression levels of
LC3-II and p62 in A-375 cells. It was noteworthy that the level of
LC3-II was significantly increased, whereas that of p62 was reduced
(Fig. 3). Furthermore, treatment
with the higher concentrations of bufadienolides led to more
pronounced changes in the expression levels of the
autophagy-associated proteins LC3-II and p62 in A-375 cells.
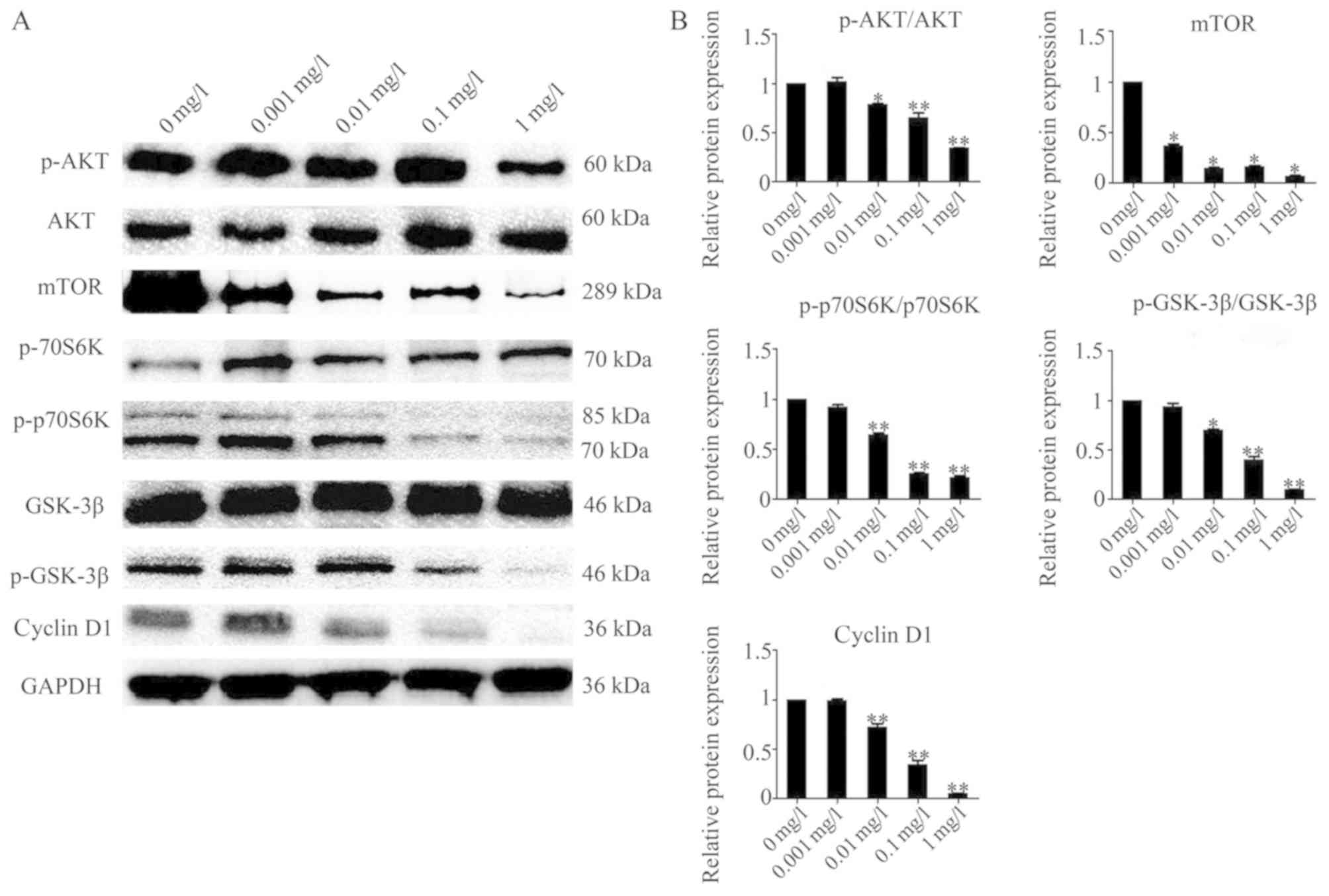
Effects of bufadienolides on the AKT
signaling pathway
To address whether the AKT signaling pathway is
involved in mediating the effects of bufadienolide treatment on
A-375 cells, the AKT pathway and its downstream proteins were
investigated by western blot analysis. No evident changes in AKT
expression were identified, whereas the level of p-AKT was
decreased upon treatment with different concentrations of
bufadienolides in the A-375 cells. There were also reductions
observed in the expression levels of p-p70S6K and p-GSK-3β, while
levels of total p70S6K and GSK-3β were not significantly changed
after exposure to different concentrations of bufadienolides. In
addition, following treatment with higher concentrations of
bufadienolides, the total mTOR and cyclin D1 levels were markedly
decreased (Fig. 4). Each bar in
the histograms represent the average of three replicates. These
findings confirmed that the effect of bufadienolides on A-375 cells
may be mediated via the regulation of different proteins downstream
of AKT.
Discussion
Melanoma, a malignant tumor of melanocytes, is the
underlying cause of 80% of mortalities associated with skin cancer
(23). In this study, it was found
that bufadienolides induced apoptosis and autophagy in A-375 cells;
furthermore, these alterations in apoptosis and autophagy were
associated with inhibition of the AKT pathway.
Two classical apoptotic pathways were selected to
examine the effect of bufadienolides on A-375 cells. First, in the
mitochondrial pathway, the Bcl-2 family of proteins exerts a major
role in tumorigenesis and tumor maintenance. A previous study
revealed that bufadienolides may mediate apoptosis through
downregulation of Bcl-2 and/or upregulation of Bax in cancer cells
(24,25). In the present study, bufadienolides
were identified to induce the apoptosis of A-375 cells, upregulate
the expression of Bax and p53, and downregulate Bcl-2 expression.
Secondly, the cell death caspases, including initiator caspases and
executioner caspases, are known to occupy a central role in the
process of apoptosis (26). In the
present study, the results also demonstrated that bufadienolides
were able to induce apoptosis of A-375 cells via the activation of
caspase-3.
In addition, autophagy is a cellular catabolic
process that helps to maintain cellular homeostasis. The regulation
of autophagy exerts an important role in tumor suppression and
promotion in numerous types of cancer (27). In cancer therapy, autophagy is
activated as an adaptive response to promote cell survival
(28). LC3-II is a marker protein
for autophagosomes, and increases in the level of this autophagy
marker have been demonstrated to inhibit tumor cell growth
(29). Furthermore, as an adaptor
protein, p62 is able to localize to the site of autophagosome
formation and interact with the autophagosome localization protein
LC3 to promote tumor cell autophagy (30,31).
In the present study, it was observed that bufadienolides
upregulated the level of LC3-II, and that p62 was itself degraded
by autophagy in a dose-dependent manner.
Accumulating evidence from recent systematic studies
has indicated that the AKT pathway exerts an immense influence on
the regulation of growth, survival and differentiation of tumor
cells (32), and is also
associated with their apoptosis and/or autophagy (33,34).
In addition, previous studies have shown that inhibition of mTOR is
related to the enhancement of apoptosis activation and autophagy
induction (35,36). The results of this study showed
that the AKT level did not change significantly after treatment of
A-375 cells with bufadienolides, but the p-AKT levels decreased,
while the mTOR expression level decreased. Inactivated mTOR leads
to a decrease in the activity of downstream proteins, p-p70S6K
(37) and p-GSK-3β (38), and inactivated GSK-3β leads to the
inactivation of cyclin D1 (39).
The results indicated that bufadienolides may
regulate AKT, p-AKT and proteins located further downstream in the
AKT pathway, including mTOR, p-p70S6K, p-GSK-3β and cyclin D1.
Based on these results, the present study is the first, to the best
of our knowledge, to demonstrate that bufadienolide induces
apoptosis and autophagy in A-375 cells, and that this may be due to
inhibition of the AKT signaling pathway. Therefore, bufadienolides
should be considered as potential drug candidates for the treatment
of melanoma by inhibiting the AKT signaling pathway. The limitation
of the present study is that the effects of bufadienolide treatment
were analyzed in only one melanoma cell line, which was not
compared with normal epidermal keratinocytes. Further studies in
vivo and in vitro in different tumor cell lines and
their corresponding normal cell lines are required to demonstrate
the anti-tumor mechanism of bufadienolides, which is part of
ongoing research. This work will ultimately increase knowledge of
the anticancer potential of these naturally occurring
compounds.
Acknowledgements
Not applicable.
Funding
This study was supported by grants from the Science
& Techno-logy Department of Sichuan Province (grant no.
2019YJ0332) and Chengdu University of TCM (grant no.
JSZX2018004).
Availability of date and materials
All the data generated or analyzed during the
present study are included in this published article.
Authors' contributions
HL and XC performed the experiments, analyzed the
data and wrote the draft version of the manuscript. XL, XC, XY, JX
and JC verified the experimental results and revised the manuscript
for important intellectual content. LY made substantial
contributions to the design of the present study, and critically
revised the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Dountsis A, Zsis C, Karagianni E and
Dahabreh J: Primary malignant melanoma of the lung: A case report.
World J Surg Oncol. 1:26–29. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang Q, Zhang R and Liu D: Long non-coding
RNA ZEB1-AS1 indicates poor prognosis and promotes melanoma
progression through targeting miR-1224-5p. Exp Ther Med.
17:857–862. 2019.PubMed/NCBI
|
|
3
|
Slominski A, Tobin DJ, Shibahara S and
Wortsman J: Melanin pigmentation in mammalian skin and its hormonal
regulation. Physiol Rev. 84:1155–1228. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chang L, Pei J, Li C, Zhang P, Zhou D, Du
W, Liu X and Jiang C: Incidence and metastasis of cutaneous
malignant melanoma with respect to ABO blood groups: A
case-controlled study in northeast of China. PLoS One.
9:e880962014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sample A and He YY: Mechanisms and
prevention of UV-induced melanoma. Photodermatol Photoimmunol
Photomed. 34:13–24. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Khan AQ, Travers JB and Kemp MG: Roles of
UVA radiation and DNA damage responses in melanoma pathogenesis.
Environ Mol Mutagen. 59:438–460. 2018. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Miura K and Green AC: Dietary Antioxidants
and Melanoma: Evidence from Cohort and Intervention Studies. Nutr
Cancer. 67:867–876. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee SJ, Lee KB, Son YH, Shin J, Lee JH,
Kim HJ, Hong AY, Bae HW, Kwon MA, Lee WJ, et al: Transglutaminase 2
mediates UV-induced skin inflammation by enhancing inflammatory
cytokine production. Cell Death Dis. 8:e31482017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Slominski AT, Brozyna AA, Zmijewski MA,
Jóźwicki W, Jetten AM, Mason RS, Tuckey RC and Elmets CA: Vitamin D
signaling and melanoma: Role of vitamin D and its receptors in
melanoma progression and management. Lab Invest. 97:706–724. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xiao D, Barry S, Kmetz D, Egger M, Pan J,
Rai SN, Qu J, McMasters KM and Hao H: Melanoma cell-derived
exosomes promote epithelial-mesenchymal transition in primary
melanocytes through paracrine/autocrine signaling in the tumor
microenvironment. Cancer Lett. 376:318–327. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Giavina-Bianchi MH, Giavina-Bianchi PF
Junior and Festa C Neto: Melanoma: Tumor microenvironment and new
treatments. An Bras Dermatol. 92:156–166. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Slominski AT and Carlson JA: Melanoma
resistance: A bright future for academicians and a challenge for
patient advocates. Mayo Clin Proc. 89:429–433. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Karimkhani C, Green AC, Nijsten T,
Weinstock MA, Dellavalle RP, Naghavi M and Fitzmaurice C: The
global burden of melanoma: Results from the Global burden of
disease study 2015. Br J Dermatol. 177:134–140. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Whiteman DC, Green AC and Olsen CM: The
growing burden of invasive melanoma: Projections of incidence rates
and numbers of new cases in six susceptible populations through
2031. J Invest Dermatol. 136:1161–1171. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim JS, Jeong TY, Cho CK, Lee YW and Yoo
HS: Antitumor effect of skin of venenum bufonis in a NCIH460 tumor
regression model. J Acupunct Meridian Stud. 3:181–187. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Deng LJ, Wang LH, Peng CK, Li YB, Huang
MH, Chen MF, Lei XP, Qi M, Cen Y, Ye WC, et al: Fibroblast
activation protein α activated tripeptide bufadienolide antitumor
prodrug with reduced cardiotoxicity. J Med Chem. 60:5320–5333.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chang H, Li J, Cao Y, Liu T, Shi S and
Chen W: Bufadienolides from venenum bufonis inhibit mTOR-mediated
cyclin D1 and retinoblastoma protein leading to arrest of cell
cycle in cancer cells. Evid Based Complement Alternat Med.
2018:32474022018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Amaral LS, Martins Ferreira J, Predes D,
Abreu JG, Noël F and Quintas LEM: Telocinobufagin and marinobufagin
produce different effects in LLC-PK1 cells: A case of functional
selectivity of bufadienolides. Int J Mol Sci. 19(pii): E27692018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sheng X, Zhu P, Qin J and Li Q: The
biological role of autophagy in regulating and controlling the
proliferation of liver cancer cells induced by bufalin. Oncol Rep.
39:2931–2941. 2018.PubMed/NCBI
|
|
20
|
Liu M, Feng LX, Sun P, Liu W, Wu WY, Jiang
BH, Yang M, Hu LH, Guo DA and Liu X: A novel bufalin derivative
exhibited stronger apoptosis-inducing effect than bufalin in A549
lung cancer cells and lower acute toxicity in mice. PLoS One.
11:e01597892016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang H, Zhang X and Zhang J: MiR-129-5p
inhibits autophagy and apoptosis of H9c2 cells induced by hydrogen
peroxide via the PI3K/AKT/mTOR signaling pathway by targeting
ATG14. Biochem Biophys Res Commun. 506:272–277. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Meng Q, Yau LF, Lu JG, Wu ZZ, Zhang BX,
Wang JR and Jiang ZH: Chemical profiling and cytotoxicity assay of
bufadienolides in toad venom and toad skin. J Ethnopharmacol.
187:74–82. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Berger MF and Garraway LA: Applications of
genomics in melanoma oncogene discovery. Hematol Oncol Clin North
Am. 23:397–414. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cui Y, Zhang Z, Zhang B, Zhao L, Hou C,
Zeng Q, Nie J, Yu J, Zhao Y, Gao T, et al: Excessive apoptosis and
disordered autophagy flux contribute to the neurotoxicity induced
by high iodine in Sprague-Dawley rat. Toxicol Lett. 297:24–33.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chou HY, Chueh FS, Ma YS, Wu RS, Liao CL,
Chu YL, Fan MJ, Huang WW and Chung JG: Bufalin induced apoptosis in
SCC-4 human tongue cancer cells by decreasing Bcl-2 and increasing
Bax expression via the mitochondria-dependent pathway. Mol Med Rep.
16:7959–7966. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zaman S, Wang R and Gandhi V: Targeting
the apoptosis pathway in hematologic malignancies. Leuk Lymphoma.
55:1980–1992. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yun CW and Lee SH: The roles of autophagy
in cancer. Int J Mol Sci. 19(pii): E34662018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Radogna F, Dicato M and Diederich M:
Cancer-type-specific crosstalk between autophagy, necroptosis and
apoptosis as a pharmacological target. Biochem Pharmacol. 94:1–11.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jena KK, Kolapalli SP, Mehto S, Nath P,
Das B, Sahoo PK, Ahad A, Syed GH, Raghav SK, Senapati S, et al:
TRIM16 controls assembly and degradation of protein aggregates by
modulating the p62-NRF2 axis and autophagy. EMBO J. 37(pii):
e983582018.PubMed/NCBI
|
|
30
|
Niklaus M, Adams O, Berezowska S, Zlobec
I, Graber F, Slotta-Huspenina J, Nitsche U, Rosenberg R, Tschan MP
and Langer R: Expression analysis of LC3B and p62 indicates intact
activated autophagy is associated with an unfavorable prognosis in
colon cancer. Oncotarget. 8:54604–54615. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sophia J, Kowshik J, Dwivedi A, Bhutia SK,
Manavathi B, Mishra R and Nagini S: Nimbolide, a neem limonoid
inhibits cytoprotective autophagy to activate apoptosis via
modulation of the PI3K/Akt/GSK-3β signalling pathway in oral
cancer. Cell Death Dis. 9:10872018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sun YZ, Cai N and Liu NN: Celecoxib
down-regulates the hypoxia-induced expression of HIF-1α and VEGF
through the PI3K/AKT pathway in retinal pigment epithelial cells.
Cell Physiol Biochem. 44:1640–1650. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Feng H, Cheng X, Kuang J, Chen L, Yuen S,
Shi M, Liang J, Shen B, Jin Z, Yan J and Qiu W: Apatinib-induced
protective autophagy and apoptosis through the AKT-mTOR pathway in
anaplastic thyroid cancer. Cell Death Dis. 9:10302018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fan YP, Liu P, Xue WK, Zhao WJ and Pan HC:
Trimebutine promotes Glioma cell apoptosis as a potential
anti-tumor agent. Front Pharmacol Front Pharmacol. 9:6642018.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shen S, Zhang Y, Wang Z, Liu R and Gong X:
Bufalin induces the interplay between apoptosis and autophagy in
glioma cells through endoplasmic reticulum stress. Int J Biol Sci.
10:212–224. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang DM, Liu JS, Deng LJ, Chen MF, Yiu A,
Cao HH, Tian HY, Fung KP, Kurihara H, Pan JX and Ye WC:
Arenobufagin, a natural bufadienolide from toad venom, induces
apoptosis and autophagy in human hepatocellular carcinoma cells
through inhibition of PI3K/Akt/mTOR pathway. Carcinogenesis.
34:1331–1342. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kim SJ, Kim JH, Jung HS, Lee TJ, Lee KM
and Chang IH: Phosphorylated p70S6K in noninvasive low-grade
urothelial carcinoma of the bladder: Correlation with tumor
recurrence. Asian J Androl. 16:611–617. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kaidanovich-Beilin O and Woodgett JR:
GSK-3: Functional Insights from Cell Biology and Animal Models.
Front Mol Neurosci. 4:402011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang Q, Zheng M, Yin Y and Zhang W:
Ghrelin stimulates hepatocyte proliferation via regulating cell
cycle through GSK3β/B-catenin signaling pathway. Cell Physiol
Biochem. 50:1698–1710. 2018. View Article : Google Scholar : PubMed/NCBI
|