Introduction
In total, 415 million people suffer from type 2
diabetes mellitus (T2DM) worldwide, and ~2/3 of cases of
diabetes-associated mortality are caused by cardiovascular
complications (1,2). T2DM was the sixth major cause of
disability in 2015, and has caused a significant economic burden on
society and individuals, with an estimated total cost of ~825
billion dollars (2). High
glucose-induced endothelial dysfunction is associated with the
development and progression of diabetic vascular complications,
including atherosclerosis and heart failure (3–5).
High glucose levels activate the nuclear factor-κB and the p38
mitogen-activated protein kinase signaling pathways, leading to
endothelial damage (6–9). Therefore, the development of novel
strategies to control the effects of high glucose levels is
necessary to prevent the complications of diabetes (10).
The Janus kinase (JAK)/signal transducer and
activator of transcription (STAT) signaling pathway is a
multifunctional mediator involved in the signal transduction from
extracellular molecules to the nucleus, regulating gene expression
(11,12). The binding of extracellular
molecules, including cytokines and growth factors, to their
corresponding receptors induces dimerization of the receptors,
which allows JAKs associated with the receptors to interact,
inducing the phosphorylation of conserved tyrosines and activation
of the downstream signal. The phosphorylated tyrosine residues and
the surrounding amino acids form a docking site. Subsequently, the
SH2 domain of STAT protein interacts with this docking site and JAK
catalyzes the phosphorylation of the STAT protein bound to the
receptor. Finally, the activated STAT protein enters the nucleus as
a dimer and binds to the promoters of its target genes, regulating
gene expression (11). Manea et
al (13) identified that the
JAK/STAT signaling pathway serves a role in the impairment of
endothelial function under high glucose conditions. In addition,
Marrero et al (14)
suggested that inhibition of the JAK/STAT signaling pathway
significantly ameliorates diabetic nephropathy. Therefore, it is
important to develop novel drugs with limited side effects able to
regulate the JAK/STAT signaling pathway, in order to prevent
diabetic complications.
Formononetin is the main active component found in
the root of Astragalus membranaceus (Fisch.) Bge, which has
been used as a herbal medicine to treat diabetes in traditional
Chinese medicine (15,16). Formononetin has been shown to be
effective in protecting diabetic endothelium (17–19);
however, whether formononetin is able to improve endothelial
function by regulating the JAK/STAT signaling pathway remains
unclear. Therefore, the present study aimed to investigate this
hypothesis, which may provide a scientific basis for the
development of novel therapeutic strategies based on this natural
medicine for the prevention and treatment of diabetic vascular
complications.
Materials and methods
Materials
HUVECs were purchased from the American Type Culture
Collection. Formononetin (purity, 98%) was purchased from Gracia
Chemicals Pvt. Ltd. Tyrphostin AG 490 (AG490; a JAK2 inhibitor) was
purchased from MedChemExpress LLC. Recombinant human interleukin
(IL)-6 was purchased from Sigma-Aldrich (Merck KGaA).
Anti-phosphorylated (p)-JAK2 (cat. no. ab32101), anti-JAK2 (cat.
no. ab39636), anti-p-STAT3 (cat. no. ab76315), anti-STAT3 (cat. no.
ab68153), anti-cleaved caspased-3 (cat. no. ab49822), anti-GAPDH
(cat. no. ab70699) and a secondary horseradish
peroxidase-conjugated goat anti-rabbit antibody (cat. no. ab7090)
were purchased from Abcam. IL-1β (cat. no. JLC6382) and
intercellular adhesion molecule 1 (ICAM-1) ELISA kit (cat. no.
JLC5547-96T) were purchased from Shanghai Jingkang Biological
Engineering Co., Ltd. PCR materials and kits, including Transzol Up
kit, TransScript II First-Strand cDNA Synthesis SuperMix kit and
TransStart Top Green qPCR Super Mix kit, were purchased from
Beijing Transgen Biotech Co., Ltd. MTT, 3-amino,
4-aminomethyl-2′,7′-difluorescein diacetate (DAF-FM DA) and a cell
lysis buffer kit were obtained from Beyotime Institute of
Biotechnology.
Cell culture
HUVECs were cultured in Dulbecco's modified Eagle's
medium (DMEM; Gibco; Thermo Fisher Scientific, Ltd.) supplemented
with 5.5 mM glucose and 10% fetal bovine serum (Zhejiang Tianhang
Biotechnology Co., Ltd.), 100 U/ml penicillin and 100 U/ml
streptomycin. Cells were cultured at 37°C in an atmosphere
containing 5% CO2 and 95% humidity. Medium was replaced
with fresh DMEM every day. Cells were passaged at 80%
confluence.
Western blotting
HUVECs were seeded in 6-well-plates
(6×104 cells/well) and treated with glucose (5 and 25
mM), IL-6 (10 ng/ml), formononetin (2, 20 and 200 µM) and AG490 (80
µM) in combination for 24 h at 36°C. Total cellular protein was
extracted using a protein extraction kit (Beyotime Institute of
Biotechnology) according to the manufacturer's protocol. Total
protein was quantified using a bicinchoninic protein assay kit.
Subsequently, 25 µg protein was loaded into each lane and proteins
were separated by 10% SDS-PAGE and transferred onto polyvinylidene
fluoride (PVDF) membranes (EMD Millipore). PVDF membranes were
blocked using TBS containing 5% skimmed milk and 0.05% Tween 20 at
37°C for 3 h, followed by incubation with the primary antibody
(1:300 dilution) at 4°C for 10 h and with secondary antibodies
(1:1,000 dilution) at 37°C for 2.5 h. Protein bands were detected
using BeyoECL Moon (Beyotime Institute of Biotechnology) and a
ChemiDoc XRS system (Bio-Rad Laboratories, Inc.) and analyzed using
ImageJ software (1.8.0; National Institutes of Health).
RNA analysis by reverse
transcription-quantitative (q)PCR
HUVECs were seeded in 6-well-plates
(6×104 cells/well) and treated with glucose (5 and 25
mM), IL-6 (10 ng/ml), formononetin (2, 20 and 200 µM) or AG490 (80
µM) in combination for 24 h at 36°C. Subsequently, total RNA was
extracted with the Transzol Up kit (Beijing Transgen Biotech Co.,
Ltd.), according to the manufacturer's protocol. A total of 5 µg
RNA from each experimental group was used to synthesize the cDNA,
according to manufacturer's protocol. Gene expression was
determined using the TransStart Top SYBR Green qPCR Super Mix kit
and analyzed using CFX manager 2.1 (Bio-Rad Laboratories, Inc.).
The thermocycling conditions were as follows: Initial denaturation
at 93°C for 4 min, followed by 33 cycles at 93°C for 16 sec and
60°C for 35 sec. The primer sequences were as follows: JAK2,
forward, (F) 5′-TCTGGGGAGTATGTTGCAGAA-3′, reverse (R)
5′-AGACATGGTTGGGTGGATACC-3′; STAT3, F 5′-CAGCAGCTTGACACACGGTA-3′, R
5′-AAACACCAAAGTGGCATGTGA-3′; IL-1β, F 5′-AGCTACGAATCTCCGACCAC-3′, R
5′-CGTTATCCCATGTGTCGAAGAA-3′; ICAM-1, F 5′-ATGCCCAGACATCTGTGTCC-3′,
R 5′-GGGGTCTCTATGCCCAACAA-3′; and β-actin, F
5′-CCTTATTCCGATCTACACAGAGC-3′ and R 5′-TATTCGGCGTAGGTCTGAGGG-3′.
β-actin was used as the internal control (20).
Cell viability assay
To detect the effect of formononetin on cellular
viability, an MTT assay was performed. HUVECs were seeded in a
96-well plate (1×104 cells/well) and were treated with
glucose (5 and 25 mM), formononetin (2, 20 and 200 µM), AG490 (80
µM) and mannitol (25 mM) in combination for 24 h at 36°C. After 24
h treatment, 20 µl MTT solution (5 mg/ml) was added into each well
for 4 h at 36°C. DMSO (150 µl/well) was used to dissolve the
formazan crystals. Cell viability was measured using a scanning
multiwell spectrophotometer at a wavelength of 570 nm.
IL-1β and ICAM-1 measurement by
ELISA
HUVECs were seeded in 6-well-plates
(6×104 cells/well) and treated with glucose (5 and 25
mM), formononetin (2, 20 and 200 µM) and AG490 (80 µM) in
combination for 24 h at 36°C. Subsequently, IL-1β and ICAM-1
protein concentrations in the cell supernatants were analyzed using
ELISA kits, according to the manufacturer's protocol.
Intracellular nitric oxide (NO)
measurement by fluorescence microscopy
HUVECs were seeded into 6-well-plates
(6×104 cells/well) and treated with glucose (5 and 25
mM), formononetin (2, 20 and 200 µM) and AG490 (80 µM) in
combination for 24 h at 36°C. Subsequently, cells were washed twice
with PBS and incubated with 5 µM DAF-FM DA (NO synthase assay kit;
Beyotime Institute for Biotechnology) at 37°C for 25 min. After
washing with buffer solution to remove the probe, NO was detected
using an Olympus fluorescence microscope (IX51S8F-3; Olympus
Corporation) at an excitation wavelength of 495 nm and an emission
wavelength of 515 nm. ImageJ software (1.8.0; National Institutes
of Health) was used to analyze fluorescence intensity.
Vascular contraction and relaxation
assay
60 Male Sprague-Dawley rats (weight, 180–200 g) were
purchased from The Animal Experiment Center of Wuhan University and
were housed according to the institutional guidelines (temperature,
25°C; relative humidity, 50%-60; light/dark cycle, 12 h) with free
access to food and water. All experimental procedures and protocols
were approved by The Institutional Animal Care and Use Committee of
the Institute for Endocrinology, Xiangyang First People's Hospital.
After acclimation for 10 days, rats were divided into six groups
(n=10/group): i) Control group; ii) high glucose group; iii) 4
mg/kg formononetin group; iv) 40 mg/kg formononetin group; v) 400
mg/kg formononetin group; and vi) 8 mg/kg AG490 group. Drugs were
delivered through intragastric administration every day. Untreated
rats in the control group were fed a standard diet (NIH-41). The
other five groups were fed a high glucose diet [10% lard oil, 20%
sucrose, 2.5% egg yolk powder, 0.5% cholesterol, 0.5% sodium
cholate and 66.5% standard diet (w/w)]. The rats were sacrificed
after 12 weeks of treatment. Thoracic aortas form the rats were
dissected to examine thoracic aortic function. For the thoracic
aortic function analysis, isolated thoracic aortas in each group
were treated with chilled Krebs-Henseleit (K-H) solution (pH 7.4).
The composition of the solution was as follows: 117.9 mM NaCl, 4.7
mM KCl, 2.6 mM MgCl2, 3.3 mM CaCl2, 25.0 mM
NaHCO3, 1.2 mM KH2PO4 and 11.0 mM
glucose. Aortas were isolated, and after removing tissue debris,
aortic rings (length, 4–7 mm) were separated from the aortas. The
aortic rings were equilibrated for 1 h at 37°C with a resting
tension of 1.5 g. Subsequently, the aortic rings were incubated
with high K+ (60 mM) solution for 20 min at 37°C to
induce maximal contraction. Then, vascular contraction induced by
phenylephrine (10−9, 10−8, 10−7
and 10−6 mol/l; Sigma-Aldrich; Merck KGaA) for 20 min at
37°C was recorded using a force-displacement transducer connected
to a polygraph. Phenylephrine was added cumulatively until the
maximum response was detected. A cumulative dose-response curve was
calculated, and, after addition of phenylephrine, the maximum
response was determined when the dose-response curve reached a
plateau. Contraction responses were calculated using the following
formula: Contraction response = [(contraction tension-resting
tension)/resting tension] ×100%. Before determining the degree of
relaxation, the aortic rings were washed three times with K-H
solution every 20 min and recontraction was induced with
10−6 mol/l phenylephrine. The cumulative dose-response
curves for acetylcholine (10−9, 10−8,
10−7, 10−6 and 10−5 mol/l;
Sigma-Aldrich; Merck KGaA) for 20 min at 37°C were calculated to
evaluate endothelial relaxation. Relaxant response was calculated
as the percentage decrease of the contraction amplitude induced by
phenylephrine before the application of acetylcholine.
Statistical analysis
SPSS version 24.0 (IBM Corp.) was used for
statistical analysis. The Shapiro–Wilk normality test was
performed, and all variables were found to be normally distributed.
Data are presented as the mean ± SEM from three experiments.
One-way analysis of variance followed by Tukey's test was used to
compare multiple groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
Effects of formononetin on the
JAK/STAT signaling pathway
High glucose levels can activate the JAK/STAT
signaling pathway in HUVECs (13).
Therefore, the effects of formononetin on the JAK/STAT signaling
pathway were investigated in HUVECs. Treatment with 25 mM glucose
for 24 h significantly increased the protein expression levels of
p-JAK2 and p-STAT3 in HUVECs (P<0.01; Fig. 1A and C). Similarly to the JAK2
inhibitor AG490, treatment with 20 and 200 µM formononetin
significantly decreased the protein expression levels of p-JAK2 and
JAK2 mRNA expression (P<0.05; Fig.
1A and B). Furthermore, 20 and 200 µM formononetin
significantly inhibited the protein expression levels of p-STAT3, a
protein downstream to JAK2, and STAT3 mRNA expression (P<0.05;
Fig. 1C and D). Additionally, the
JAK/STAT signaling agonist IL-6 was used to examine the
pharmacological effects of formononetin. Even when HUVECs were
cotreated with high glucose and IL-6, formononetin was able to
significantly inhibit p-JAK2 and p-STAT3 protein levels (P<0.01;
Fig. 2A and C), in addition to the
mRNA expression levels of JAK2 and STAT3 (P<0.01; Fig. 2B and D). Collectively, the present
results suggested that formononetin, similarly to AG490, was able
to inhibit the JAK/STAT signaling pathway in HUVECs treated with
high glucose.
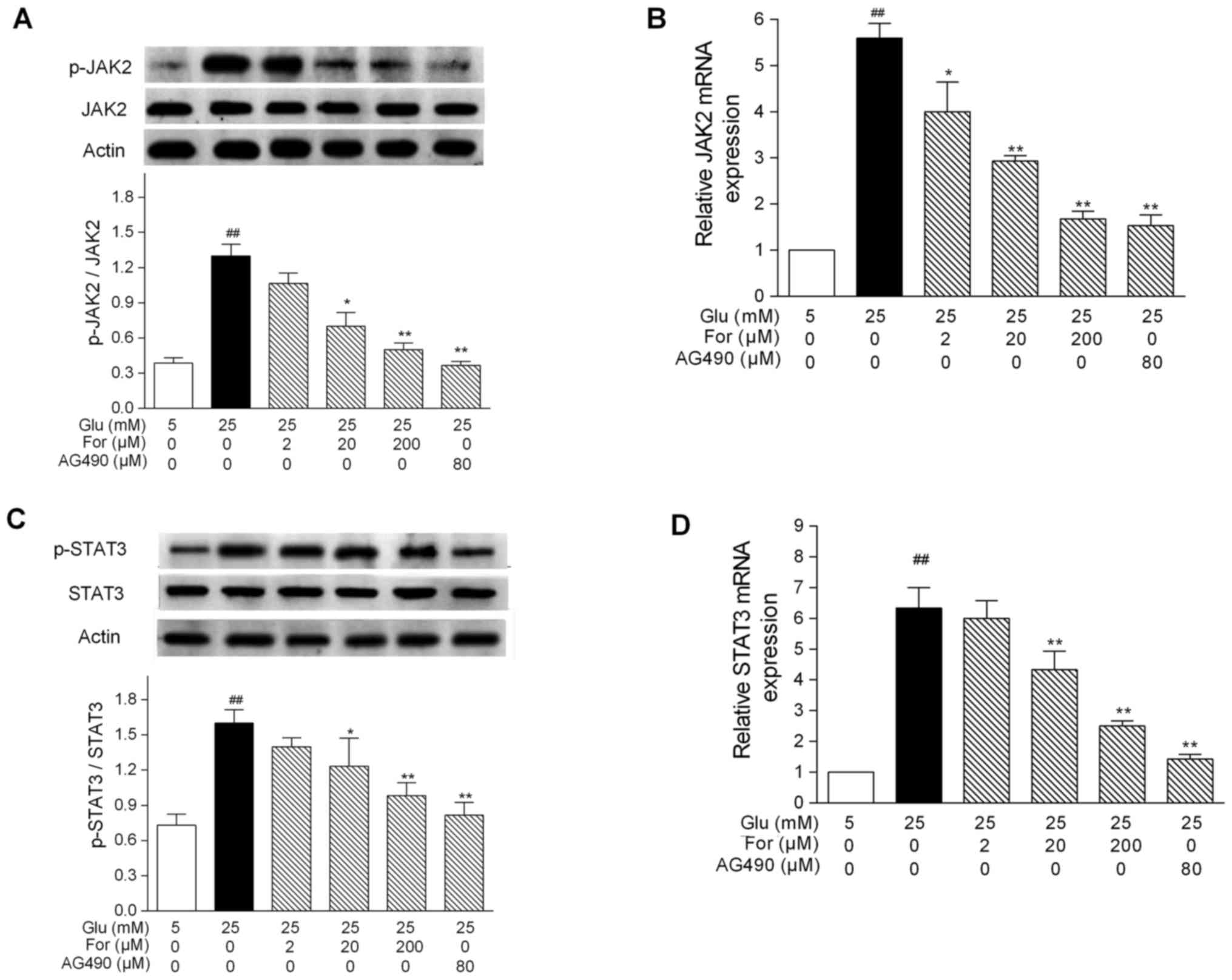 | Figure 1.Effects of formononetin on JAK2 and
STAT3 expression under high glucose conditions. HUVECs were treated
with formononetin or AG490 under high glucose conditions. Protein
expression levels of (A) JAK2 and p-JAK2, and (C) STAT3 and
p-STAT3, were determined by western blot analysis. Protein bands
were analyzed by densitometry. mRNA expression levels of (B) JAK2
and (D) STAT3, as determined by quantitative PCR. Results are
presented as the mean ± SEM from three independent experiments. The
ratio of p-protein to total/unmodified protein was normalized to
actin. ##P<0.01 vs. 5 mM glucose group; *P<0.05,
**P<0.01 vs. 25 mM glucose group. AG490, tyrphostin AG
490; For, formononetin; Glu, glucose; JAK2, Janus kinase 2; p-,
phosphorylated; STAT3, signal transducer and activator of
transcription 3. |
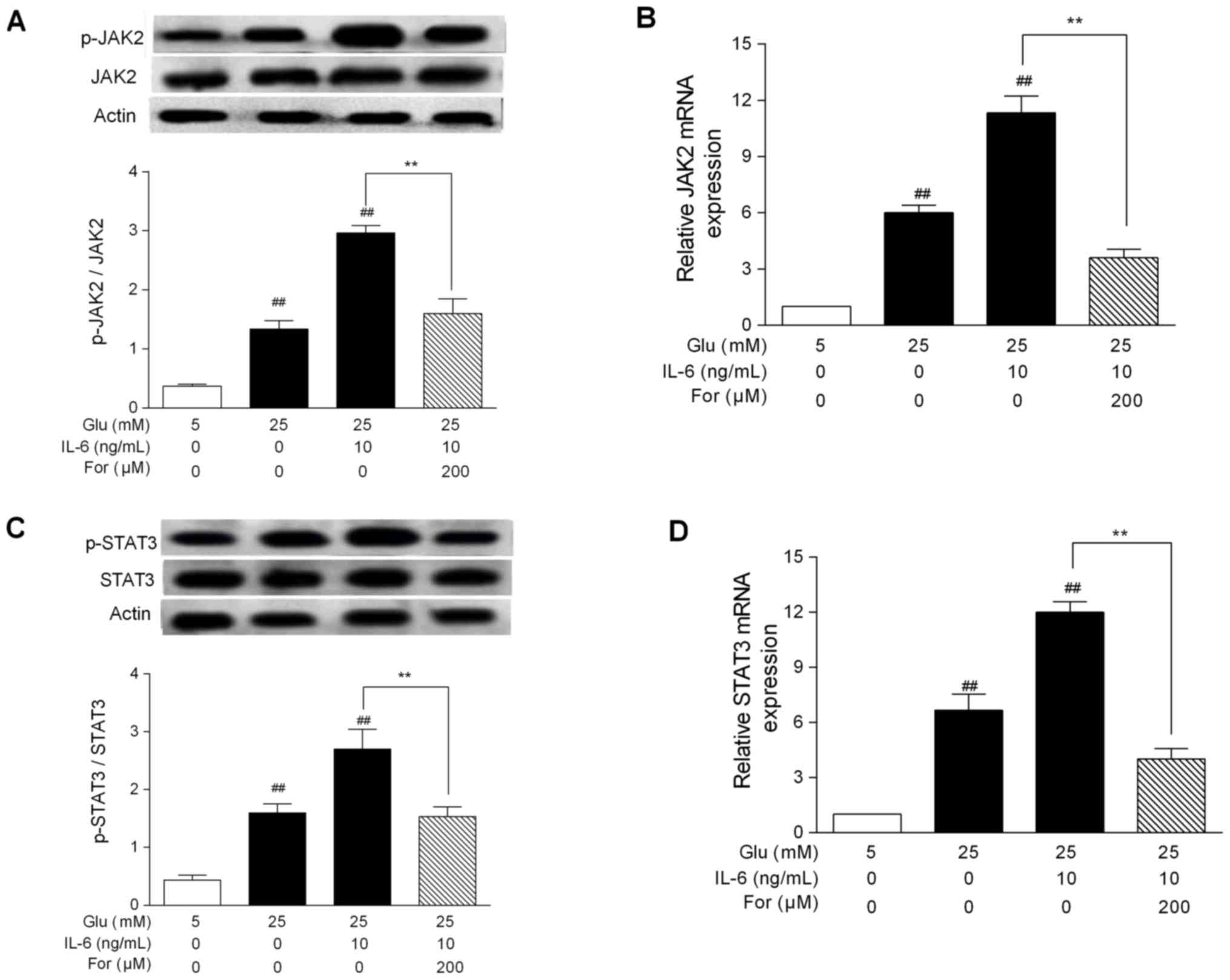 | Figure 2.Effects of formononetin on JAK2 and
STAT3 expression under high glucose conditions and following IL-6
treatment. HUVECs were cotreated with glucose + IL-6 and were
treated with formononetin. Protein expression levels of (A) JAK2
and p-JAK2, and (C) STAT3 and p-STAT3, were determined by western
blotting. Protein bands were analyzed by densitometry. mRNA
expression level of (B) JAK2 and (D) STAT3, as determined by
quantitative PCR. Results are presented as the mean ± SEM from
three independent experiments. ##P<0.01 vs. 5 mM
glucose group; **P<0.01, as indicated. AG490, tyrphostin
AG 490; For, formononetin; Glu, glucose; IL-6, interleukin-6; JAK2,
Janus kinase 2; p-, phosphorylated; STAT3, signal transducer and
activator of transcription 3. |
Effects of formononetin on high
glucose-induced cellular damage
After investigating the role of formononetin in the
JAK/STAT signaling pathway in HUVECs, the effects of formononetin
on HUVEC viability were examined under high glucose conditions.
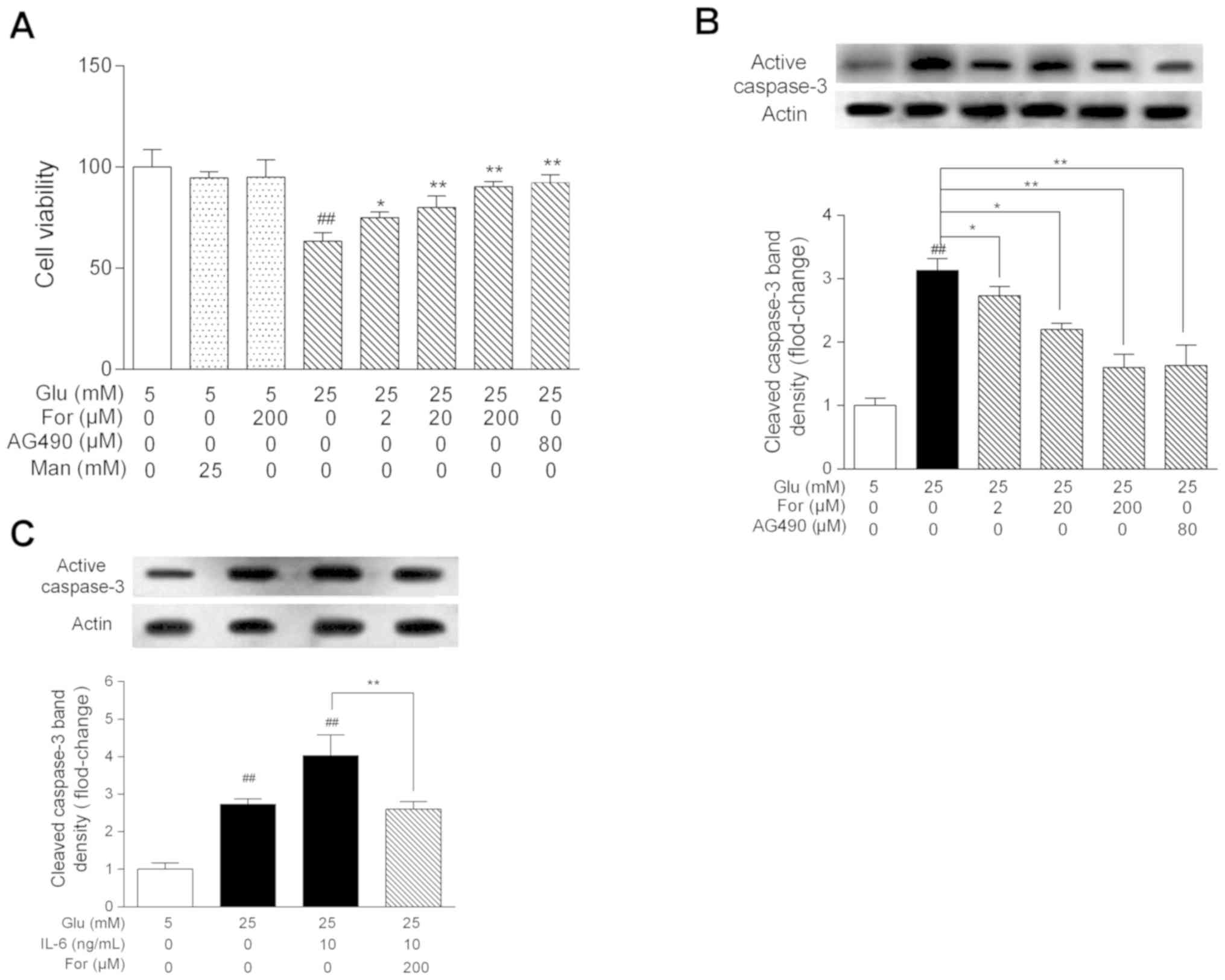
Following high glucose stress, the viability of HUVECs was
decreased to 67% (Fig. 3A). There
was no significant difference in cellular viability between the 25
mM mannitol group and the 5 mM glucose-control group, suggesting
that the decreased viability in high glucose was not related to
osmotic pressure. However, formononetin increased cellular
viability under high glucose stress in a dose-dependent manner
(P<0.05). Furthermore, formononetin could significantly decrease
the protein expression levels of active caspase-3 in HUVECs treated
with high glucose or cotreated with high glucose and IL-6
(P<0.05; Fig. 3B and C). In
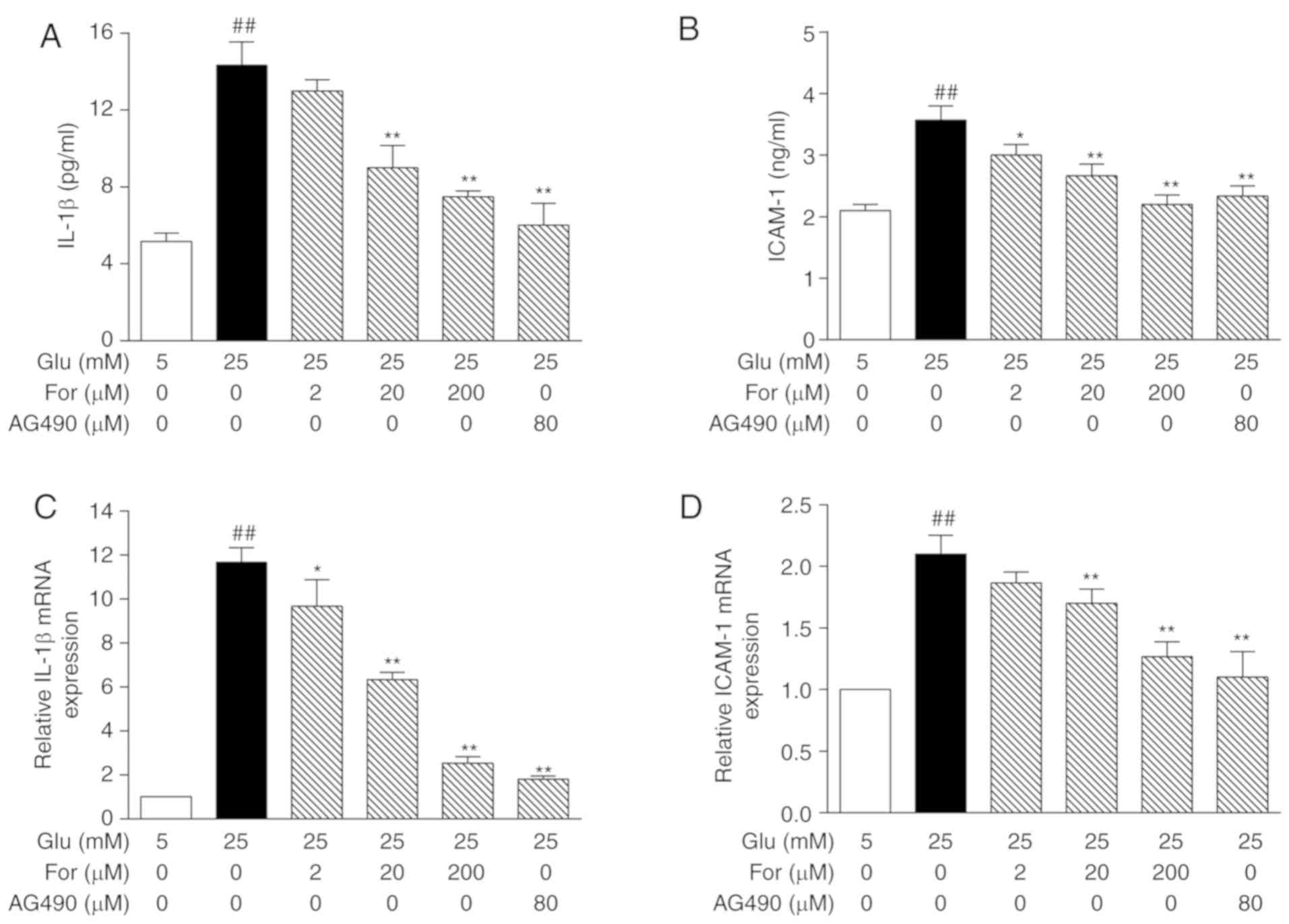
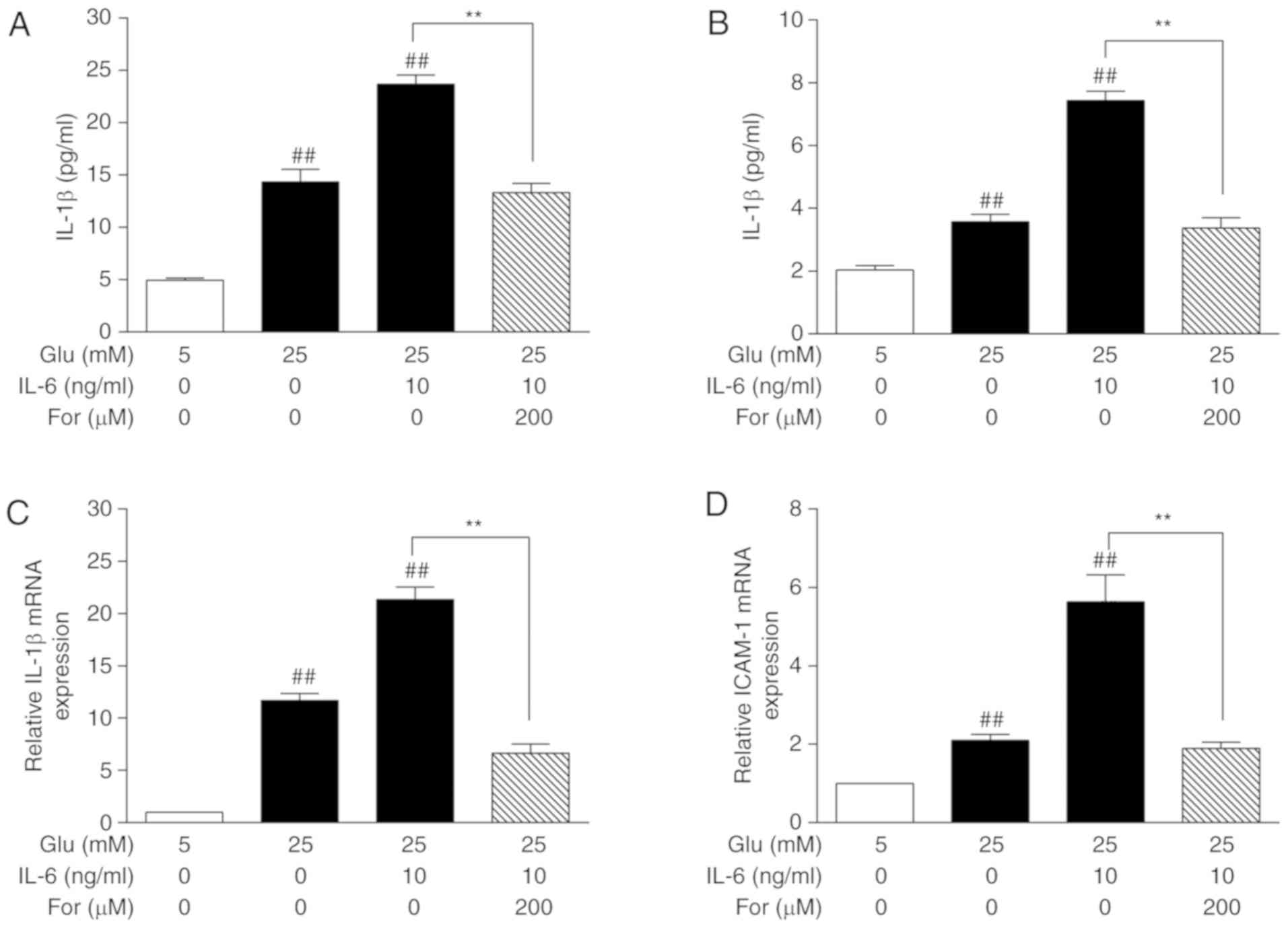
order to investigate the inhibitory effects of formononetin on
inflammation, the expression levels of IL-1β and ICAM-1 were
investigated by ELISA and qPCR. The glucose-induced protein and
mRNA expression levels of IL-1β and ICAM-1 were decreased after
formononetin treatment, also in the presence of IL-6 (P<0.05;
Figs. 4 and 5). Collectively, formononetin may protect
against endothelial damage, at least in part, through the JAK/STAT
signaling pathway.
Effects of formononetin on endothelial
function
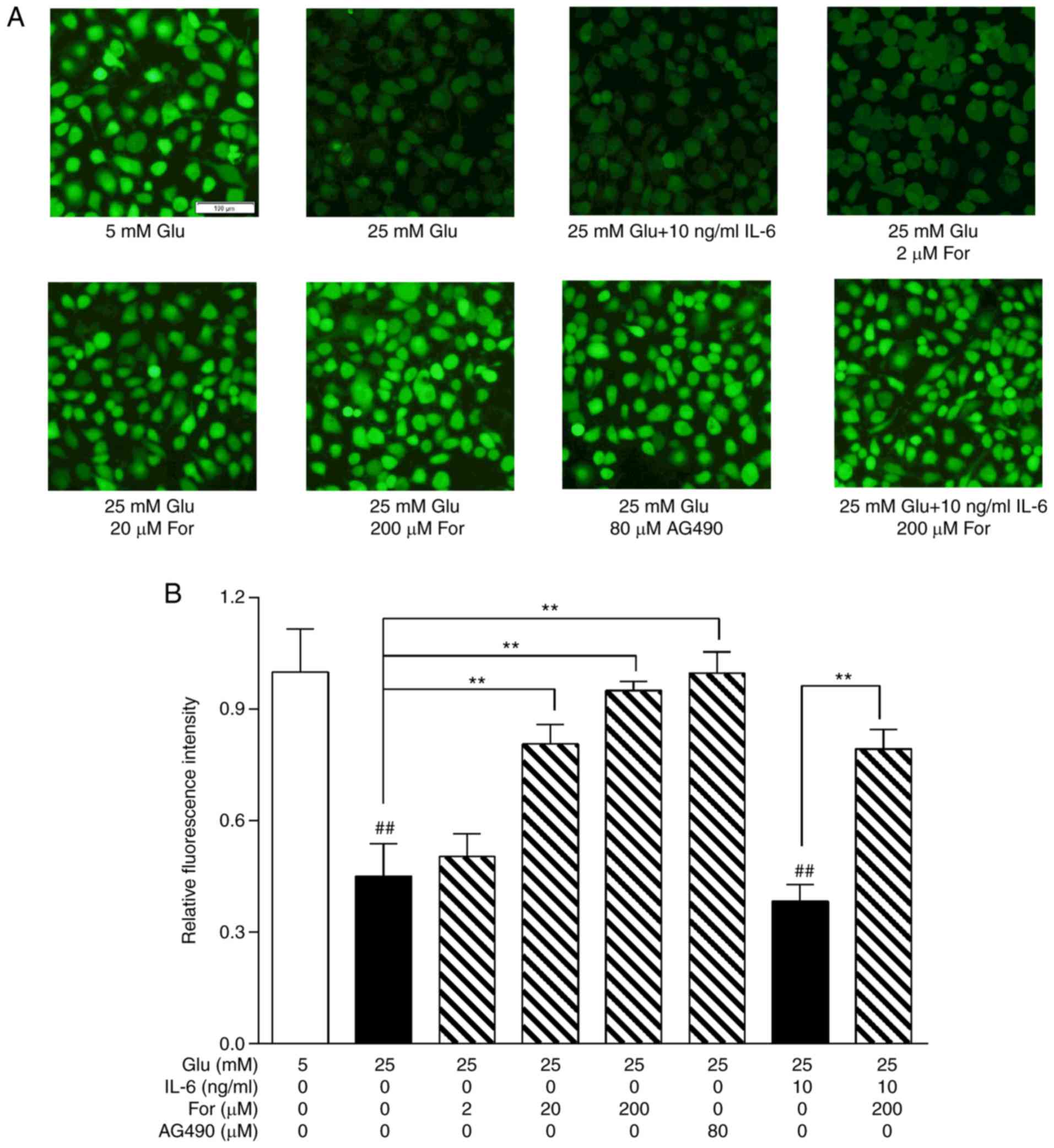
Endothelial function was examined to determine
whether formononetin could improve endothelial function. High
glucose stress decreased NO production, which is associated with
endothelial dysfunction (21).
In vitro, NO levels were significantly restored following
treatment with 20 and 200 µM formononetin, and 80 µM AG490 under 25
mM glucose conditions (P<0.01; Fig.
6). Additionally, in HUVECs cotreated with 25 mM glucose and 10
ng/ml IL-6, formononetin (200 µM) still exhibited a protective role
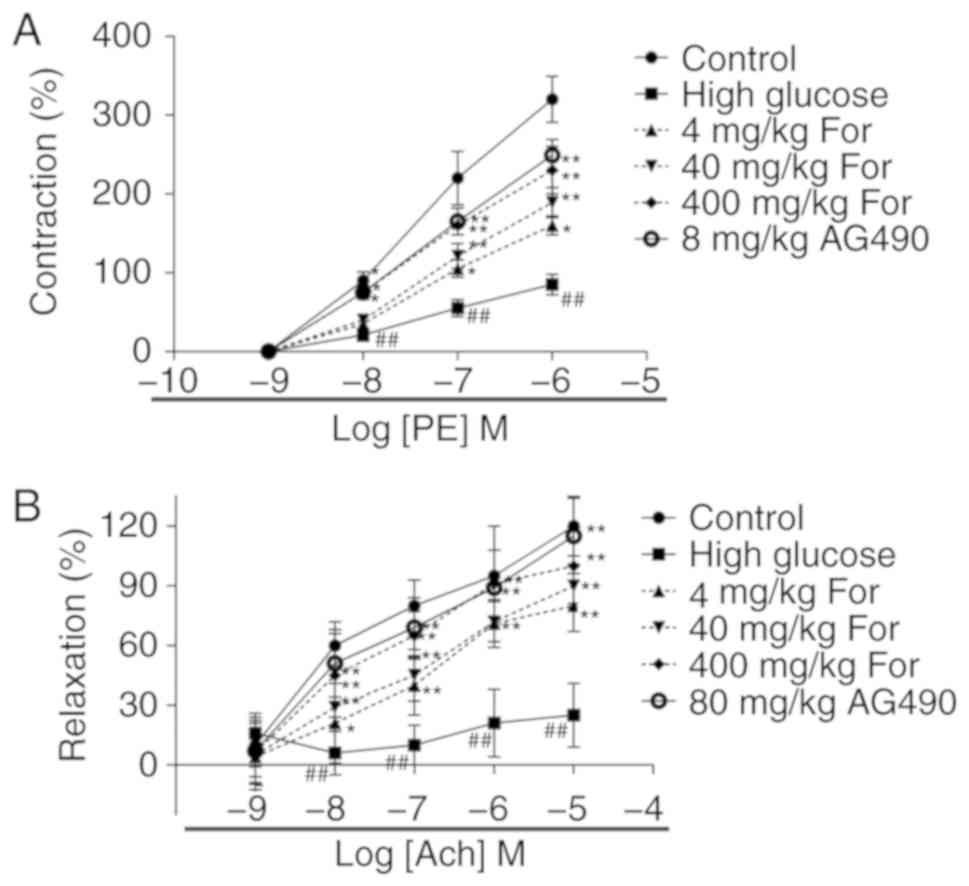
in NO production. Additionally, the contraction response to
phenylephrine was investigated in thoracic aortic rings isolated
from rats fed a high-glucose diet (Fig. 7). Compared with the control group
(rats fed a standard diet), aortic rings isolated from rats in the
high glucose group exhibited a significantly reduced maximal
contractile response induced by phenylephrine and a significantly
decreased maximal relaxation response induced by acetylcholine.
Following formononetin administration at 4, 40 and 400 mg/kg,
contraction and relaxation of aortic rings were significantly
improved (P<0.05). The JAK2 inhibitor AG490 at 80 mg/kg
exhibited similar effects to formononetin at 400 mg/kg.
Collectively, in vivo and in vitro results suggested
that formononetin improved endothelial function under high glucose
stress through the JAK/STAT signaling pathway.
Discussion
Overnutrition, a major topic of research in
endocrinology, is responsible for inducing metabolic diseases,
including T2DM (22). Insulin
resistance in patients with T2DM leads to multiple cardiovascular
diseases, including atherosclerosis and coronary disease, that are
caused by impaired glucose and lipid metabolism (23,24).
Therefore, restoring and maintaining a normal extracellular
environment is important to protect the cardiovascular system from
complications associated with diabetes. Although drugs, such as
insulin and thiazolidinediones, have been used to treat diabetes,
their long-term side effects can be significant (25). Natural products have attracted the
attention of various research groups due to their therapeutic
potential and limited side effects (26). Formononetin has been shown to be
effective in alleviating the symptoms of diabetes and related
complications, such as cardiovascular and kidney diseases (27). Additionally, formononetin has been
reported to be able to improve the function of pancreatic β-cells
and whole-body glucose homeostasis (28,29).
In the present study, formononetin was revealed to protect
endothelial function through the JAK/STAT signaling pathway, and
the present results provided a scientific basis for the development
of novel clinical strategies using formononetin to treat T2DM.
The JAK/STAT signaling cascade is one of multiple
pleiotropic signaling cascades involved in signal transduction
during development and homeostasis. In 2008, Vargha et al
(30) reported that IL-6 promotes
epithelial-to-mesenchymal transition of human peritoneal
mesothelial cells possibly through the JAK2/STAT3 signaling
pathway. Notably, the JAK/STAT signaling cascade is an important
factor activated by hyperglycemia that may cause
diabetes-associated endothelial dysfunction (13). JAK2 belongs to the family of
non-receptor protein tyrosine kinases, which consists of JAK1,
JAK2, JAK3 and tyrosine kinase 2. To transduce its downstream
signaling, activated JAK recruits the cytoplasmic transcription
factor STAT to the receptor complex. To the best of our knowledge,
seven STAT proteins have been identified (31). The JAK2/STAT3 pathway may be
involved in the occurrence of endothelial dysfunction under glucose
stress (32). In the present
study, high glucose levels activated the JAK2/STAT3 signaling
pathway. Furthermore, formononetin was able to significantly
inhibit the JAK2/STAT3 signaling pathway following high glucose
treatment or cotreatment with high glucose and IL-6, an antagonist
of the JAK2/STAT3 signaling pathway.
After identification of the inhibitory effects of
formononetin on the JAK/STAT signaling pathway, its effects on
endothelial cells under high glucose stress were examined in the
present study. Notably, formononetin could function similarly to
AG490 in improving the viability of HUVECs under high glucose
conditions. Caspase-3 is a key apoptosis-associated factor that
interacts with other members of its family, such as caspase-8 and
caspase-9 (33). The present
results suggested that formononetin could inhibit caspase-3
activation following treatment with 25 mM glucose or cotreatment
with 25 mM glucose and 10 ng/ml IL-6. The present results suggested
that formononetin could prevent HUVECs damage and apoptosis by
inhibiting the JAK/STAT signaling pathway.
Inflammation is a complex biological process
activated in response to harmful stimuli, such as pathogens and
metabolic dysfunction (34,35).
IL-1β, activated by the NLR family pyrin domain containing 3
inflammasome, is a strong inflammatory mediator involved in
endothelial cell damage (36,37).
IL-1β promotes inflammation and can also cause apoptosis.
Formononetin significantly inhibited IL-1β expression at the
protein and mRNA levels. In addition, formononetin could inhibit
ICAM-1 expression, suggesting that formononetin may inhibit the
adhesion of inflammatory cells under high glucose conditions
(38). Inflammatory mediators are
involved in the dysregulation of NO synthesis in HUVECs (39). Therefore, the effects of
formononetin on NO synthesis were investigated in the present
study. NO acts as a vasodilator and vascular homeostatic mediator
in maintaining normal endothelial function (40). However, high glucose levels induce
the JAK/STAT pathway and promote the activity of inflammatory
mediators that can impair the synthesis and release of NO (41). The present results suggested that
formononetin significantly restored NO synthesis under high glucose
conditions or in cells cotreated with high glucose and IL-6,
suggesting that formononetin could improve NO synthesis and release
in HUVECs under high glucose conditions. In addition, the effects
of formononetin on contraction and relaxation of isolated thoracic
aortic rings were investigated in rats fed a high-glucose diet. The
present results suggested that formononetin treatment significantly
improved endothelial function in rats fed a high-glucose diet.
In conclusion, the present study suggested that
formononetin could regulate the JAK/STAT signaling pathway,
improving endothelial viability and function in vivo and
in vitro. The present findings suggested that formononetin
may potentially represent a novel candidate natural product that
could be used to prevent and treat diabetic vascular
complications.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YX designed the experiment, ZZ and XZ performed the
experiments and wrote the paper. YD and ML analyzed the results.
YX, ZZ, YD and ML revised the paper.
Ethics approval and consent to
participate
All experimental procedures and protocols were
approved by The Institutional Animal Care and Use Committee of the
Institute for Endocrinology, Xiangyang First People's Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Solomon CG: Reducing cardiovascular risk
in type 2 diabetes. N Engl J Med. 348:457–459. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chatterjee S, Khunti K and Davies MJ: Type
2 diabetes. Lancet. 389:2239–2251. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Paneni F, Beckman JA, Creager MA and
Cosentino F: Diabetes and vascular disease: Pathophysiology,
clinical consequences, and medical therapy: Part I. Eur Heart J.
34:2436–2443. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Garber AJ: Diabetes and vascular disease.
N Engl J Med. 2:1–5. 1990.
|
|
5
|
Murea M, Ma L and Freedman BI: Genetic and
environmental factors associated with type 2 diabetes and diabetic
vascular complications. Rev Diabet Stud. 9:6–22. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Martinon F, Burns K and Tschopp J: The
inflammasome: A molecular platform triggering activation of
inflammatory caspases and processing of proIL-beta. Mol Cell.
10:417–426. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hink U, Tsilimingas N, Wendt M and Münzel
DT: Mechanisms underlying endothelial dysfunction in diabetes
mellitus: Therapeutic implications. Treat Endocrinol. 2:293–304.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Peng H, Hong S, Li P, Li J, Zhou X and
Zhang L: High glucose concentration increases the MAPK and
TGF-beta-2 expression in cultured P38 human umbilical vein
endothelial cells. Basic & Clinical Medicine. 27:pp. 169–173.
2007, http://en.cnki.com.cn/Article_en/CJFDTOTAL-JCYL200702010.htm
|
|
9
|
Takaishi H, Taniguchi T, Takahashi A,
Ishikawa Y and Yokoyama M: High glucose accelerates MCP-1
production via p38 MAPK in vascular endothelial cells. Biochem
Biophys Res Commun. 305:122–128. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Barr EL, Zimmet PZ, Welborn TA, Jolley D,
Magliano DJ, Dunstan DW, Cameron AJ, Dwyer T, Taylor HR, Tonkin AM,
et al: Risk of cardiovascular and all-cause mortality in
individuals with diabetes mellitus, impaired fasting glucose, and
impaired glucose tolerance: The Australian Diabetes, Obesity, and
Lifestyle Study (AusDiab). Circulation. 116:151–157. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kisseleva T, Bhattacharya S, Braunstein J
and Schindler CW: Signaling through the JAK/STAT pathway, recent
advances and future challenges. Gene. 285:1–24. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rawlings JS, Rosler KM and Harrison DA:
The JAK/STAT signaling pathway. J Cell Sci. 117:1281–1283. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Manea SA, Manea A and Heltianu C:
Inhibition of JAK/STAT signaling pathway prevents
high-glucose-induced increase in endothelin-1 synthesis in human
endothelial cells. Cell Tissue Res. 340:71–79. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Marrero MB, Banes-Berceli AK, Stern DM and
Eaton DC: Role of the JAK/STAT signaling pathway in diabetic
nephropathy. Am J Physiol Renal Physiol. 290:F762–F768. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Godefroit P, Hai S, Yu T and Lauters P:
New hadrosaurid dinosaurs from the uppermost Cretaceous of
northeastern China. Acta Palaeontol Polonica. 53:47–74. 2008.
View Article : Google Scholar
|
|
16
|
Tong Y and Hou H: Effects of Huangqi
Guizhi Wuwu Tang on diabetic peripheral neuropathy. J Altern
Complement Med. 12:506–509. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun T, Cao L, Ping NN, Wu Y, Liu DZ and
Cao YX: Formononetin upregulates nitric oxide synthase in arterial
endothelium through estrogen receptors and MAPK pathways. J Pharm
Pharmacol. 68:342–351. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu J, Ke X, Ma N, Wang W, Fu W, Zhang H,
Zhao M, Gao X, Hao X and Zhang Z: Formononetin, an active compound
of Astragalus membranaceus (Fisch) Bunge, inhibits
hypoxia-induced retinal neovascularization via the HIF-1α/VEGF
signaling pathway. Drug Des Dev Ther. 10:3071–3081. 2016.
View Article : Google Scholar
|
|
19
|
Wu JH, Li Q, Wu MY, Guo DJ, Chen HL, Chen
SL, Seto SW, Au AL, Poon CC, Leung GP, et al: Formononetin, an
isoflavone, relaxes rat isolated aorta through
endothelium-dependent and endothelium-independent pathways. J Nutr
Biochem. 21:613–620. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cosentino F, Hishikawa K, Katusic ZS and
Lüscher TF: High glucose increases nitric oxide synthase expression
and superoxide anion generation in human aortic endothelial cells.
Circulation. 96:25–28. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dabelea D and Harrod CS: Role of
developmental overnutrition in pediatric obesity and type 2
diabetes. Nutr Rev. 71 (Suppl 1):S62–S67. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wiernsperger NF and Bouskela E:
Microcirculation in insulin resistance and diabetes: More than just
a complication. Diabet Metab. 29:6S77–6S87. 2003. View Article : Google Scholar
|
|
24
|
Ndisang JF, Rastogi S and Vannacci A:
Insulin resistance, type 1 and type 2 diabetes, and related
complications 2015. J Diabetes Metab. 2015:2341352015.
|
|
25
|
Palmer SC, Mavridis D, Nicolucci A,
Johnson DW, Tonelli M, Craig JC, Maggo J, Gray V, De Berardis G,
Ruospo M, et al: Comparison of clinical outcomes and adverse events
associated with glucose-lowering drugs in patients with type 2
diabetes: A meta-analysis. JAMA. 316:313–324. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jermendy G: Can type 2 diabetes mellitus
be considered preventable? Diabetes Res Clin Pract. 68 (Suppl
1):S73–S81. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee H, Lee D, Kang KS, Song JH and Choi
YK: Inhibition of intracellular ROS accumulation by formononetin
attenuates cisplatin-mediated apoptosis in LLC-PK1 cells. Int J Mol
Sci. 19:2018.
|
|
28
|
Wang Y, Zhu Y, Gao L, Yin H, Xie Z, Wang
D, Zhu Z and Han X: Formononetin attenuates IL-1β-induced apoptosis
and NF-κB activation in INS-1 cells. Molecules. 17:10052–10064.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Qiu G, Tian W, Huan M, Chen J and Fu H:
Formononetin exhibits anti-hyperglycemic activity in
alloxan-induced type 1 diabetic mice. Exp Biol Med (Maywood).
242:223–230. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Vargha R, Bender T, Riesenhuber A,
Endemann M, Kratochwill K and Aufricht C: Effects of
epithelial-to-mesenchymal transition on acute stress response in
human peritoneal mesothelial cells. Nephrol Dial Transplant.
23:3494–3500. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schindler C, Levy DE and Decker T:
JAK-STAT signaling: From interferons to cytokines. J Biol Chem.
282:20059–20063. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liao JQ, Lin JQ, Zhang WJ, Xu L, Zhi XM,
Lin K and Wu W: Role of JAK/STAT signaling pathway in high
glucose-induced damage in human umbilical vein endothelial cells.
Chin J Pathophysiol. 32:392–397. 2016.(In Chinese).
|
|
33
|
Budihardjo I: Biochemical pathways of
caspase activation during apoptosis. Annu Rev Cell Dev Biol. 15(1):
269–290. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Griendling KK, Sorescu D and Ushio-Fukai
M: NAD(P)H oxidase: Role in cardiovascular biology and disease.
Circ Res. 86:494–501. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mehta JL and Li D: Identification,
regulation and function of a novel lectin-like oxidized low-density
lipoprotein receptor. J Am Coll Cardiol. 39:1429–1435. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dinarello CA: Biologic basis for
interleukin-1 in disease. Blood. 87:2095–2147. 1996.PubMed/NCBI
|
|
37
|
Merhi-Soussi F, Kwak BR, Magne D,
Chadjichristos C, Berti M, Pelli G, James RW, Mach F and Gabay C:
Interleukin-1 plays a major role in vascular inflammation and
atherosclerosis in male apolipoprotein E-knockout mice. Cardiovasc
Res. 66:583–593. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang L, Froio RM, Sciuto TE, Dvorak AM,
Alon R and Luscinskas FW: ICAM-1 regulates neutrophil adhesion and
transcellular migration of TNF-alpha-activated vascular endothelium
under flow. Blood. 106:584–592. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Clapp BR, Hingorani AD, Kharbanda RK,
Mohamed-Ali V, Stephens JW, Vallance P and MacAllister RJ:
Inflammation-induced endothelial dysfunction involves reduced
nitric oxide bioavailability and increased oxidant stress.
Cardiovasc Res. 64:172–178. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Förstermann U and Sessa WC: Nitric oxide
synthases: Regulation and function. Eur Heart J. 33:829–837,
837a-837d. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang X, Shaw S, Amiri F, Eaton DC and
Marrero MB: Inhibition of the JAK/STAT signaling pathway prevents
the high glucose-induced increase in TGF-beta and fibronectin
synthesis in mesangial cells. Diabetes. 51:3505–3509. 2002.
View Article : Google Scholar : PubMed/NCBI
|