Introduction
Postoperative cognitive dysfunction (POCD) is the
impairment of certain neurophysiological regions in brain after
surgical therapy, especially in older patients (1). Sevoflurane is commonly used as an
inhaled anesthetic of alkanes in clinical practice (2). Sevoflurane inhalation anesthesia has
been revealed to induce the increased apoptosis of hippocampal
neurons in aged rats, resulting in cognitive dysfunction (3). Therefore, it is a practical and
necessary task to explore the pathogenesis and mechanism of POCD,
to seek effective drugs for the inhibition of neurotoxicity and
cognitive dysfunction, to improve the life quality of patients
after surgery, and to reduce the cost of medical treatment.
Rapamycin, a macrolide antibiotic, has been
clinically used as an immunosuppressant (4). Studies have revealed that rapamycin
has a protective effect on neurodegenerative diseases with strong
anti-aging properties. Rapamycin can delay or inhibit the
occurrence and aggravation of multiple neurodegenerative diseases
(5). Therefore, rapamycin is
considered to be a potential drug for the prevention and treatment
of neurodegenerative diseases. Rapamycin has been revealed to
improve sevoflurane-induced cognitive dysfunction in rats by
activating autophagy and eliminating intracellular abnormalities
and misfolded proteins (6).
Researchers also identified that rapamycin-enhanced autophagy can
reduce pathological changes such as amyloid-β peptide (Aβ)
accumulation and Tau hyperphosphorylation in the hippocampus of
Alzheimer's disease (AD) rats, inhibit neuronal apoptosis and
improve their learning and memory functions (7). However, the mechanism of rapamycin on
improving sevoflurane-induced cognitive dysfunction in rats has not
yet been elucidated.
Autophagy plays an important role in the prevention
of neurodegenerative diseases, tumors and the infection of
pathogenic microorganisms (8–10).
Some studies have demonstrated that the occurrence and development
of neurological degenerative diseases, such as AD and Parkinson's
disease, are associated with autophagy dysfunction (11,12).
Previous studies have confirmed that the TLR4/NF-κB signaling
pathway can promote the production of reactive oxygen species in
microglia and induce apoptosis (13). The present study aimed to observe
the protective effect of rapamycin on sevoflurane-induced cognitive
dysfunction in aged rats and its effect on autophagy-related
proteins, and to explore the regulatory mechanism of the
TLR4-MyD88/TRIF-NF-κB signaling pathway.
Materials and methods
Experimental animals and group
assignment
In total, 50 male specific-pathogen-free
Sprague-Dawley rats aged 15 months and weighing 260–280 g were
purchased from the Animal Laboratory Center of China Medical
University (License No. SCXK(Liao)-2013-0001; License No.
SYXK(Liao)-2013-0007). The project was approved by the China
Medical University Institutional Animal Care and Use Committee
(IACUC No. 2017018R). All animals were maintained in individual
cages under controlled ambient temperature (24±2°C) and 40–70%
humidity in a 12-h light/dark cycle. Standard pelleted chow and
drinking water were available ad libitum. The rats were
allowed to acclimate to these conditions for at least 1 week. The
health and behavior of rats were observed daily by animal husbandry
staff at our facility. To reduce the suffering and distress, some
animal tools and nesting material were provided to the rats.
The rats were randomly assigned into five groups: i)
10 rats were placed in the blank control group (control group); ii)
10 rats were placed in the sevoflurane group (SEV group; the rats
in SEV group were anesthetized for 5 h in 2% sevoflurane); iii) 10
rats were placed in the rapamycin pretreatment group [RAP group;
the rats were intraperitoneally injected with rapamycin (20 mg/kg)
2 days before anesthesia for 5 h in 2% sevoflurane (14)]; iv) 10 rats were placed in the TLR4
inhibitor group [TLR group; the rats were intraperitoneally
injected with rapamycin (20 mg/kg) 2 days before anesthesia for 5 h
in 2% sevoflurane, and TLR4 inhibitor TAK242 (0.5 mg/kg) was
intracerebrally injected into rats at 1 h before rapamycin
treatment (15)]; v) 10 rats were
placed in the sevoflurane+ rapamycin+3-Methyladenine (3MA)
autophagy inhibitor group [3MA group; the rats in the 3MA group
were intraperitoneally injected with rapamycin (20 mg/kg) 2 days
before anesthesia for 5 h in 2% sevoflurane, and 21 mg/kg 3MA
autophagy inhibitor was intracerebrally injected at 1.5 h before
rapamycin treatment. After 24 h, physiological data were recorded,
including heart rate, mean arterial pressure, arterial carbon
dioxide tension and arterial oxygen tension. After the Morris water
maze (MWM) experiment, blood samples of rats were collected by
saphenous vein puncture, centrifuged at 12,000 × g at 4°C to obtain
serum and stored at −20°C. Then the rats were sacrificed by
overdose anesthesia, 800 mg/kg pentobarbital, intraperitoneally
(6,16). The hippocampus of rats was
collected, one half was fixed in 4% paraformaldehyde and another
half was stored in liquid nitrogen.
Morris water maze experiment
The MWM experiment, consisting of an acquisition
test and a special probe test, is a well-established method for
assessing learning and memory abilities in rodents. The acquisition
test lasted 4 days, twice daily, with 15-minute interval between
tests. On day 5, the spatial probe test was conducted. The
acquisition test was performed to assess learning and memory
abilities in rats. Rats were randomly placed into the water in a
random order at a fixed entry point of each quadrant. The duration
time to search and climb the platform in the water was recorded as
the escape latency, and the rats stayed on the platform for at
least 3 sec. If a rat failed to climb onto the platform within 120
sec, it was manually guided onto the platform with a stick and made
to stay there for 30 sec. It was then returned to the cage for the
next test. The spatial probe test was used to assess the spatial
memory of rats. After the platform was removed from the pool, the
rats were placed into the water from the opposite side of the
original target quadrant, and allowed to search for the platform.
The number of platform crossings within 120 sec was recorded and
the proportion of time spent in the original target quadrant to the
total time was calculated.
H&E staining
The hippocampus fixed in paraformaldehyde was
immerged in different concentrations of alcohol (70, 80, 90, 95,
and 100%), permeabilized in xylene, embedded in wax, and sliced
into 4-µm thick sections using a microtome. The sections were
dewaxed, stained with hematoxylin for 5 min, washed with PBS,
differentiated with 1% alcoholic hydrochloric acid, stained with
eosin for 30 sec, followed by dehydration through a graded alcohol
series and washed off. Subsequently, these sections were mounted
with neutral resin. The pathological changes in the hippocampus
were observed using a light microscope.
TUNEL assay
Cell apoptosis was assessed in accordance with the
kit instruction (Roche Diagnostics). Briefly, 5-µm paraffin
sections were dewaxed, permeabilized and sealed. After treatment
with 50 µl TUNEL reaction solution, the sections were incubated in
a wet dark box at 37°C for 60 min. The sections were then incubated
with a NEUN antibody (1:300; cat. no. ab177487; Abcam) at 37°C for
1 h. Following this, 50 µl streptavidin-HRP solution and goat
anti-rabbit IgG H&L (Cy3®; 1:1,000; cat. no. ab6939;
Abcam) was added followed by incubation in the dark box for 30 min.
Nuclei were stained with DAPI and observed under a fluorescence
microscope. After obtaining images, the number of total nuclei and
TUNEL-positive nuclei were counted. Apoptosis ratio= (the number of
TUNEL-positive nuclei)/the total number of nuclei × 100%.
ELISA
ELISA was performed to assess the expression of
brain injury markers S-100β, NSE and inflammatory factors TNF-α,
IL-1β, IL-6, and IL-10. Standard preparation (100 µl) and diluted
samples (100 µl) were added in the corresponding reaction plate
wells, lightly shaken for 30 sec, and incubated at 20–25°C for 20
min. The reaction plate was washed with a washer machine. Serum
sample (100 µl) was added in each well and incubated at 37°C for 2
h. After washing, 100 µl HRP-labeled secondary antibody was added
in each well and incubated at 37°C for 30 min. After subsequent
washing, 50 µl of substrate A and 50 µl of substrate B were added
and visualized in a dark room for 15 min. The reaction was
terminated by adding 50 µl of stop buffer. OD values were measured
at 450 nm using a microplate reader (EXL808). The standard curve
was drawn with the OD value as the ordinate and the standard
concentration as the abscissa. The curve equation and R value were
calculated. The corresponding concentration of the sample was
obtained according to the curve equation.
Western blot assay
Tissue sample was added in RIPA lysate (cat. no.
89900; Thermo Fisher Scientific, Inc.) containing a protease
inhibitor on ice for 30 min. The supernatant was collected, and the
protein concentration was measured with a BCA Protein
Quantification kit (cat. no. 23225; Thermo Fisher Scientific,
Inc.). After 10% SDS-PAGE (20 µg per lane), the samples were
transferred onto PVDF membranes. Bax (1:1,000; cat. no. ab32503;
Abcam), Bcl-2 (1:1,000; cat. no. ab59348; Abcam), caspase-3 (1:500;
cat. no. ab13847; Abcam), Beclin1 (1:2,000; cat. no. ab207612;
Abcam), LC3B (1:3,000; cat. no. ab51520; Abcam), TLR4 (1:500; cat.
no. ab13556; Abcam), MyD88 (1:1,000; cat. no. ab133739; Abcam),
NF-κB p65 (1:1,000; cat. no. ab246347; Abcam) and GAPDH (1:10,000;
cat. no. ab181602; Abcam) antibodies were added and incubated at
4°C overnight. Next, the PVDF membrane was washed with PBS, a goat
anti-rabbit IgG horseradish peroxidase-conjugated secondary
antibody (1:10,000; cat. no. ab6721; Abcam) was added and incubated
at room temperature for 2 h. The proteins were visualized with
Novex™ ECL Chemiluminescent Substrate Reagent kit (Invitrogen;
Thermo Fisher Scientific, Inc.) and gel imaging system (Gel Doc™
XR; Bio-Rad Laboratories, Inc). Absorbance values were analyzed
using ImageJ (v1.8.0; National Institutes of Health).
RT-qPCR
Brain tissue was added in TRIzol reagent (cat. no.
15596026; Invitrogen; Thermo Fisher Scientific, Inc.). According to
TRIzol reagent instructions, total RNA was extracted from tissues
and cells, and reversely-transcribed into first-strand cDNA (cat.
no. 4387406; Invitrogen; Thermo Fisher Scientific, Inc.). In
accordance with a Real-time qPCR kit (cat. no. RR820A, Takara
Biotechnology Co., Ltd.), PCR was performed with the following
conditions: pre-denaturation at 95°C for 30 sec; 40 cycles at 95°C
for 5 sec and 60°C for 20 sec; analysis of melting curve: 95°C for
1 sec, 65°C for 15 sec and 95°C for 5 sec. Results were calculated
using 2−ΔΔCq method (17). Primer sequences are listed in
Table I.
 | Table I.Reverse transcription-quantitative PCR
using gene primers. |
Table I.
Reverse transcription-quantitative PCR
using gene primers.
| Gene | Primer (5→3) |
|---|
| Beclin-1 | Forward:
GACACTGGACTTCCTCCGG |
|
| Reverse:
GATTGCTGATGTGGATAC |
| LC3 | Forward:
CGAGAGCGAGAGAGATGAAGACGG |
|
| Reverse:
GGTAACGTCCCTTTTTGCCTTG |
| GTA |
|
| p62 | Forward:
CGGAGGTCATCTCAGGAAGG |
|
| Reverse:
CGATCAGCAGAGTGGCAATAG |
| TLR4 | Forward:
AAGGGCTTCTACTCAGAG |
|
| Reverse:
AGGACCCACATGGGCACT |
| MyD88 | Forward:
GTAGCCAGCCTCTGAAAC |
|
| Reverse:
AGCCAGGATGATGTCTAC |
| NF-κB p65 | Forward:
TTTCAAAAGTGGCATTGCTT |
|
| Reverse:
TTAAGCTGTAAAATCACA |
| GAPDH | Forward:
GTCATCAACGGGAAACC |
|
| Reverse:
CATGGAGAAGGCTGGGG |
Statistical analysis
Data were analyzed using SPSS 19.0 software (IBM
Corp.), and expressed as the mean ± standard deviation. Paired
comparisons were conducted using a Student's t-test. Intergroup
comparisons were analyzed using one way ANOVA followed by Tukey's
post hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Rapamycin reduces sevoflurane-induced
brain injury in aged rats
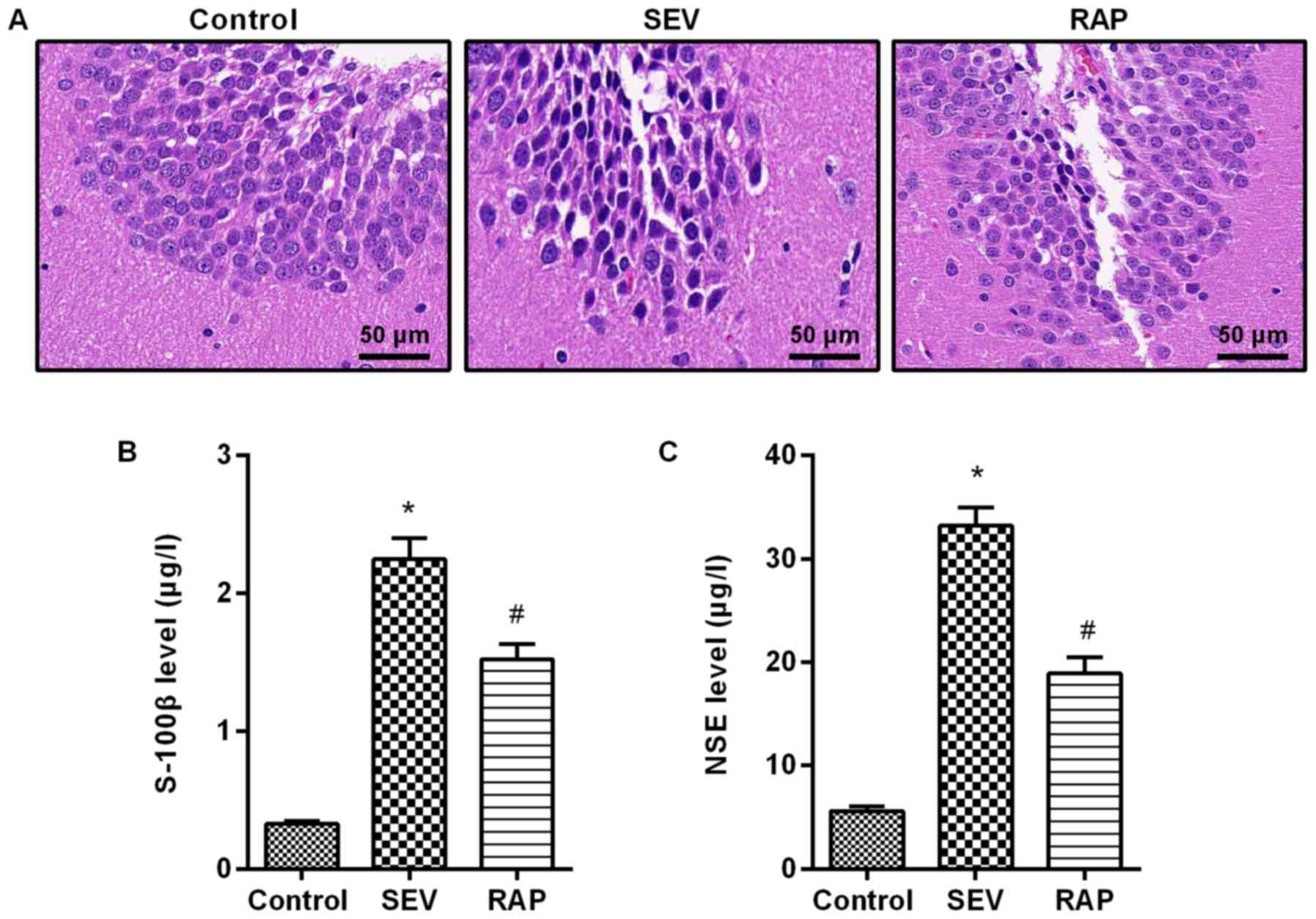
The results from H&E staining revealed that
neurons were arranged neatly and tightly, and the cytoplasm and
nucleus were plump and distinct in the control group. Neurons were
arranged disorderly and loosely, cells were smaller, and nuclear
condensation and cytoplasm were decreased in the SEV group. A small
number of cells exhibited morphological changes, and normal neurons
were still observed, which were arranged neatly and tightly in the
RAP group (Fig. 1A). By ELISA
detection it was revealed that compared with the control group,
serum S-100β and NSE expression were significantly higher in the
SEV group (P<0.05). Compared with the SEV group, serum S-100β
and NSE expression were significantly lower in the RAP group
(P<0.05; Fig. 1B and C). These
results indicated that rapamycin reduced sevoflurane-induced brain
injury in aged rats.
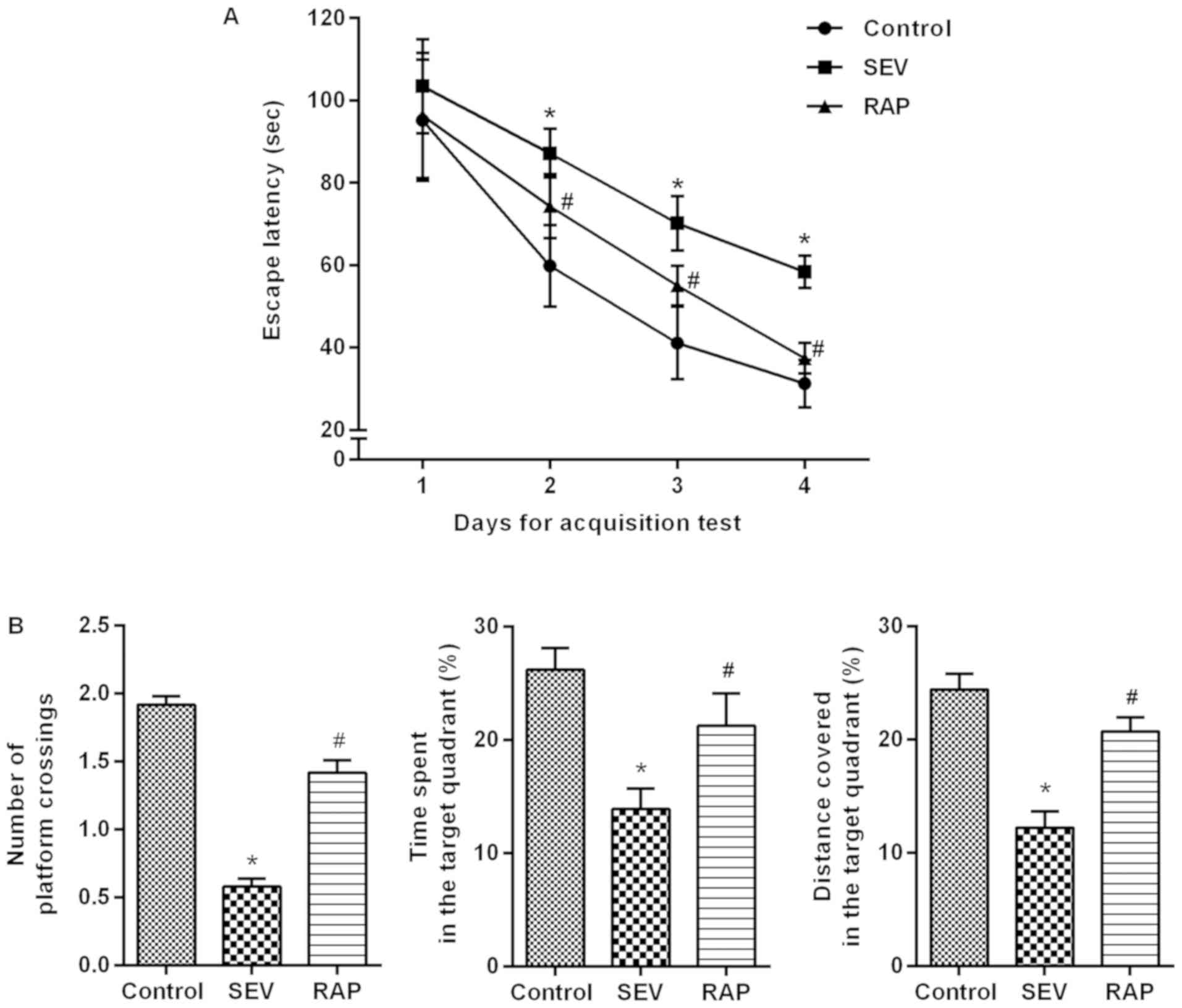
Rapamycin alleviates
sevoflurane-induced cognitive dysfunction in aged rats
Models of cognitive dysfunction induced by
sevoflurane inhalation in aged rats were intervened with rapamycin
for 24 h. An MWM experiment was conducted to determine the effects
of rapamycin on learning and memory abilities of
sevoflurane-treated rats. In the acquisition test, the escape
latency of rats in each group was gradually shortened. From 2 to 4
days, compared with control group, the escape latency was
significantly increased in the SEV group (P<0.05). Compared with
the SEV group, the escape latency of the RAP group was
significantly decreased (P<0.05; Fig. 2A). In the spatial probe test,
compared with control group, the percentage of time spent, the
distance covered in the target quadrant and the number of platform
crossings were significantly decreased in the SEV group
(P<0.05). Compared with the SEV group, the percentage of time
spent, the distance covered in the target quadrant and the number
of platform crossings were significantly increased in the RAP group
(P<0.05; Fig. 2B). It is
therefore indicated that rapamycin can alleviate
sevoflurane-induced learning and memory impairments in aged
rats.
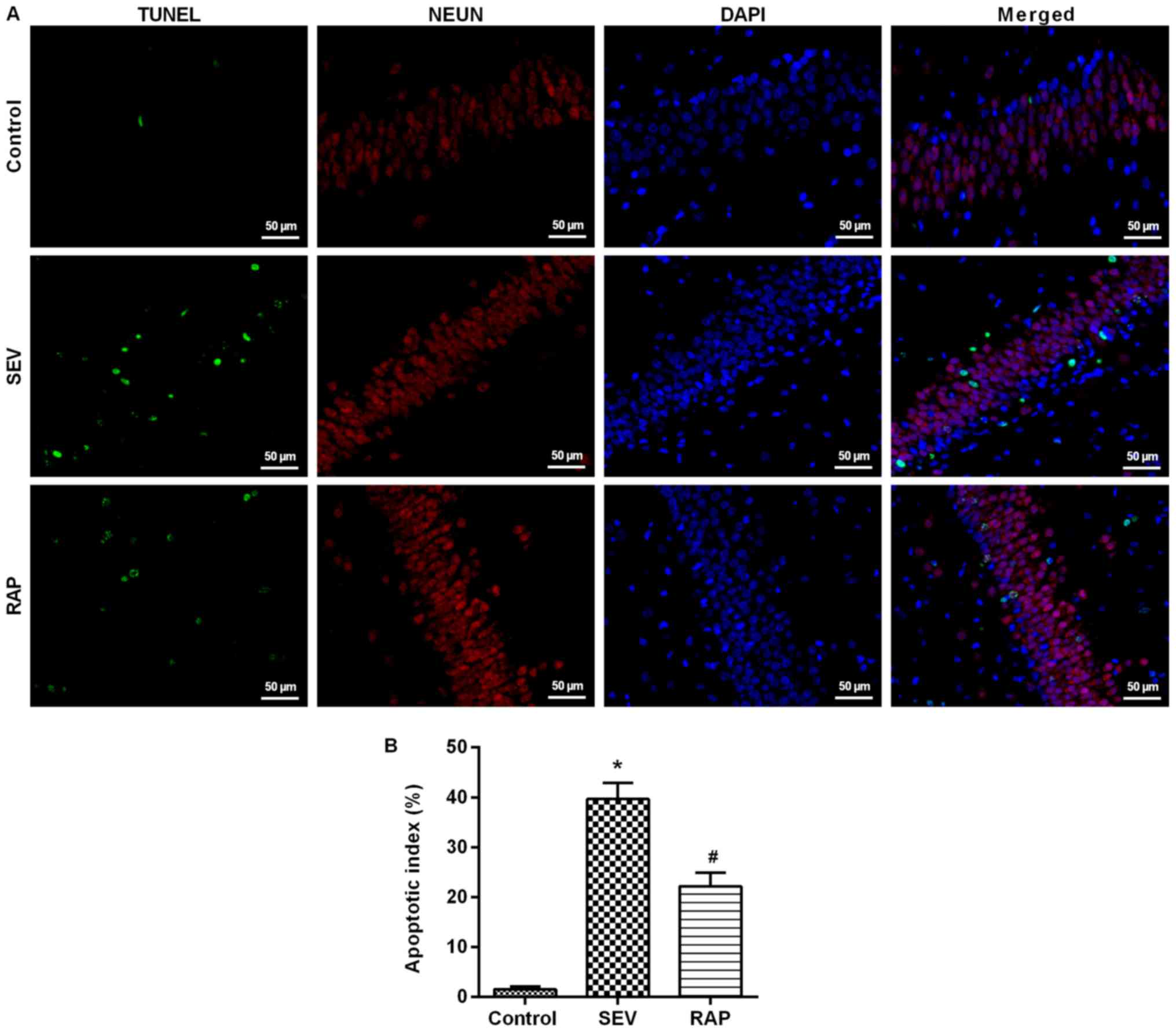
Rapamycin mitigates
sevoflurane-induced neuronal apoptosis in aged rats
TUNEL assay results revealed that compared with the
control group, the number of positive cells was significantly
higher in the SEV group (P<0.05). Compared with the SEV group,
the number of positive cells in the rat hippocampus was
significantly lower in the RAP group (P<0.05; Fig. 3). Western blot assay results
revealed that compared with the control group, Bcl-2 expression was
significantly lower, but Bax and cleaved caspase-3 expression was
significantly higher in the SEV group (P<0.05). Compared with
the SEV group, Bcl-2 expression was significantly higher, but Bax
and cleaved caspase-3 expression was significantly lower after
rapamycin pretreatment in the RAP group (P<0.05; Fig. 4B). These findings indicated that
rapamycin can inhibit sevoflurane-induced neuronal apoptosis and
prevent neuronal degeneration.
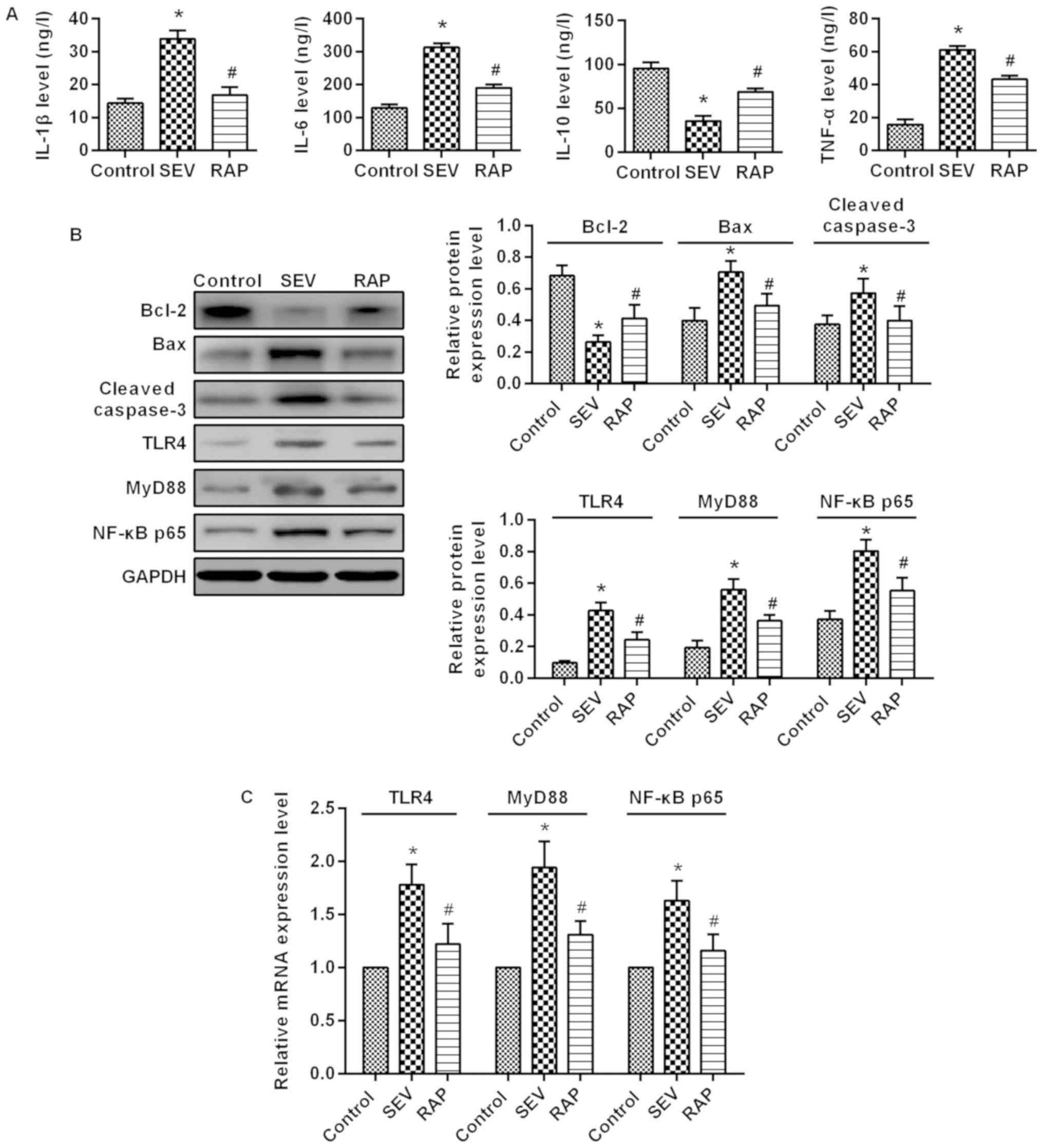
Rapamycin reduces sevoflurane-induced
inflammatory response in aged rats
ELISA results demonstrated that compared with the
control group, IL-1β, IL-6 and TNF-α expression was significantly
higher, but IL-10 expression was significantly lower in the SEV
group (P<0.05). Compared with the SEV group, IL-1β, IL-6 and
TNF-α expression was significantly lower, but IL-10 expression was
significantly higher after pretreatment in the RAP group
(P<0.05; Fig. 4A). These
results indicated that rapamycin relieves sevoflurane-induced
inflammatory responses in aged rats.
Rapamycin inhibits the expression of
the TLR4/MyD88/NF-κB signaling pathway induced by sevoflurane in
aged rats
Western blot assay (Fig. 4B) and RT-qPCR (Fig. 4C) were used to determine the
changes in the TLR4/MyD88/NF-κB signaling pathway in the rat brain.
Compared with the control group, TLR4, MyD88 and NF-κB p65
expression was significantly higher in the SEV group (P<0.05).
Compared with the SEV group, TLR4, MyD88 and NF-κB p65 expression
was significantly lower in the RAP group (P<0.05). These
findings indicated that rapamycin improves sevoflurane-induced
brain injury in aged rats possibly by inhibiting the
TLR4/MyD88/NF-κB signaling pathway.
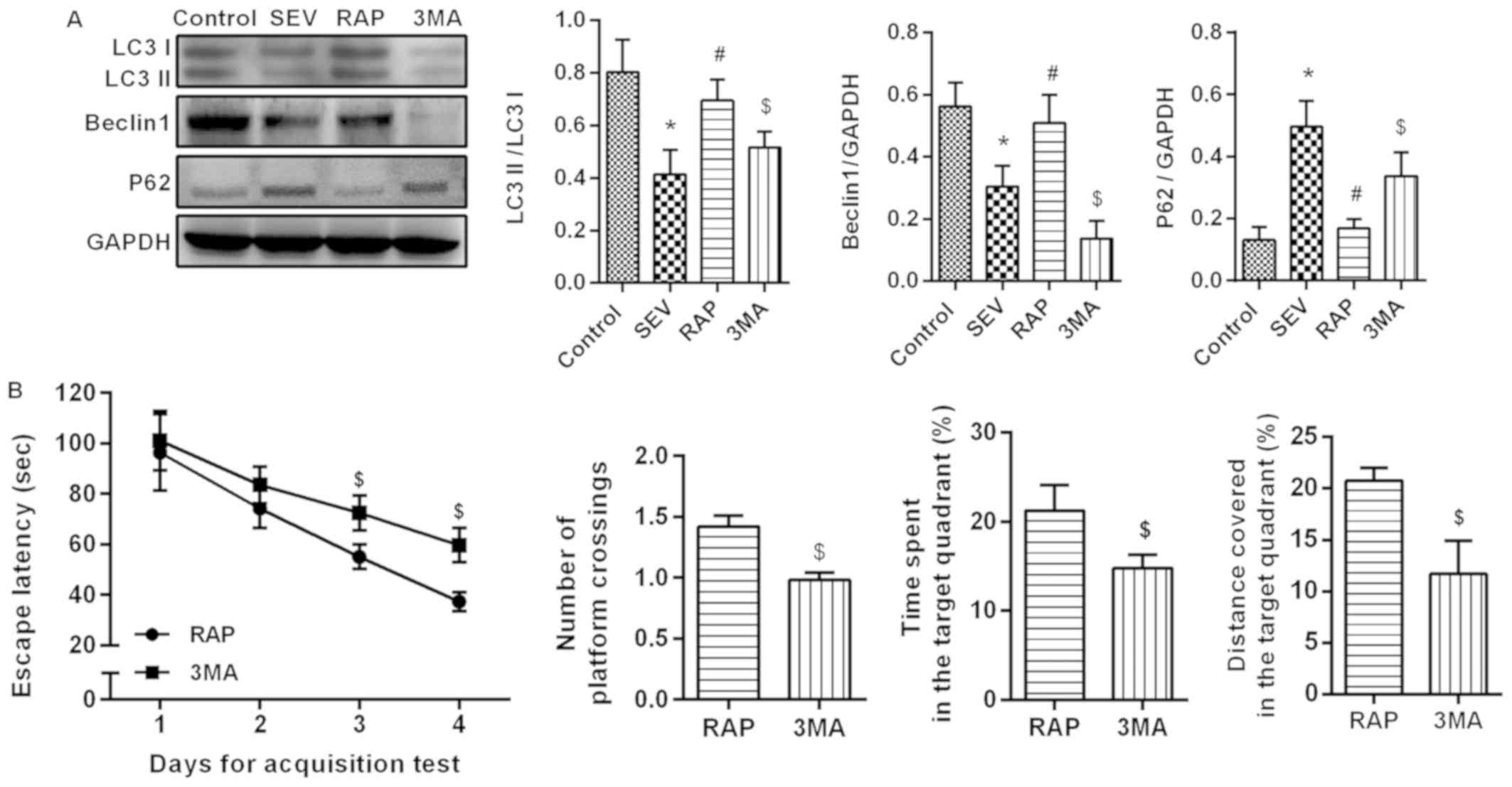
Rapamycin activates
sevoflurane-induced autophagy in aged rats
Western blot assay (Fig. 5A) results revealed that compared
with the control group, LC3II/I and Beclin1 expression was
significantly lower, but p62 expression was significantly higher in
the SEV group (P<0.05). Compared with the SEV group, LC3II/I and
Beclin1 expression was significantly higher, but p62 expression was
significantly lower in the RAP group (P<0.05). The effect of
rapamycin weakened after the administration of autophagy
inhibitors. Compared with the RAP group, LC3II/I and Beclin1
expression was significantly lower, but p62 expression was
significantly higher in the 3MA group (P<0.05). An MWM
experiment was conducted to determine the effects of autophagy
inhibitor treatment on learning and memory abilities. In the
acquisition test, the escape latency of rats in each group was
gradually shortened, and in 3–4 days, compared with the RAP group,
the escape latency was significantly increased in the 3MA group
(P<0.05). In the spatial probe test, compared with the RAP
group, the percentage of time spent, the distance covered in the
target quadrant and the number of platform crossings were
significantly decreased in the 3MA group (P<0.05´Fig. 5B). These results confirmed that
rapamycin activates sevoflurane-induced autophagy in aged rats.
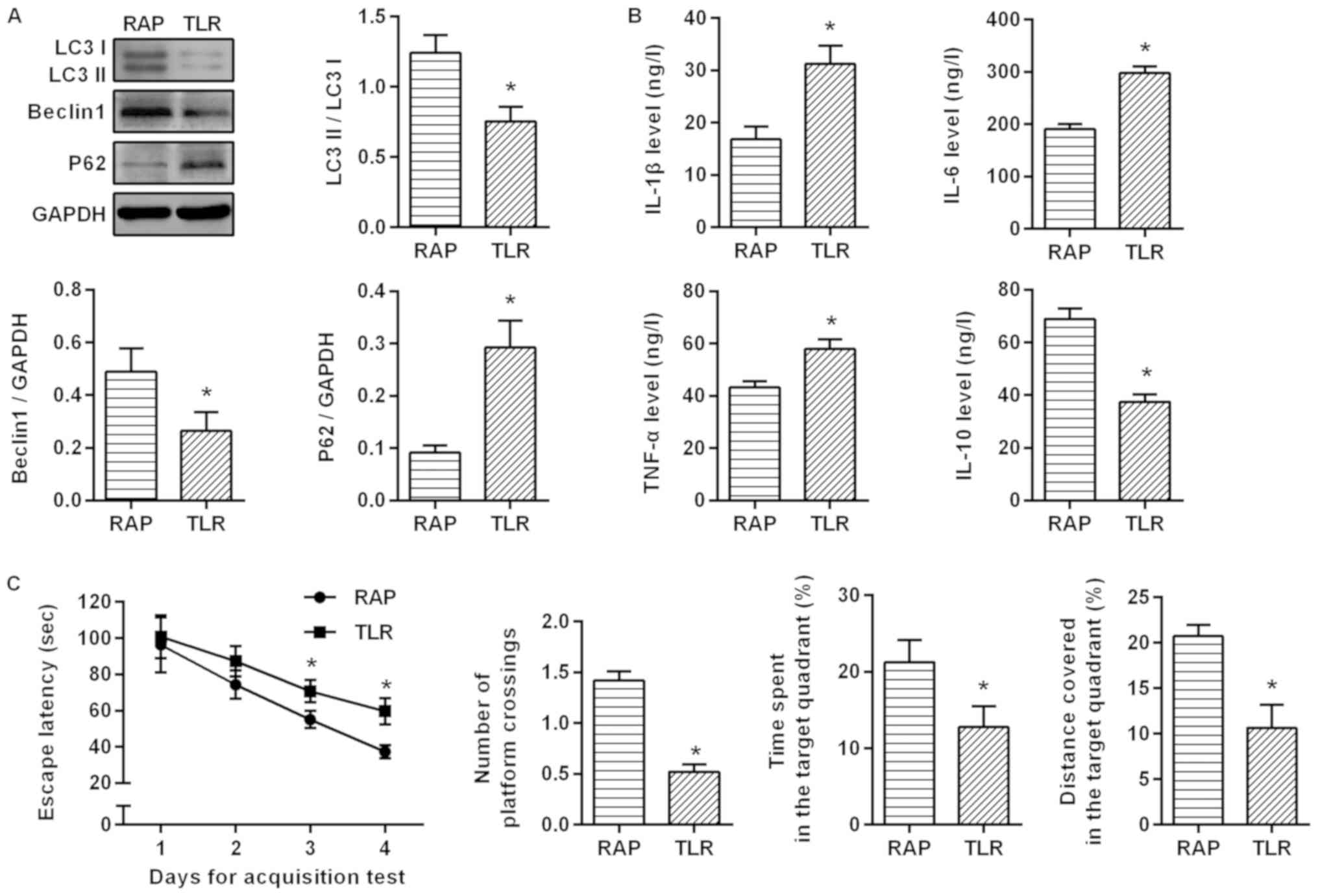
Rapamycin improves sevoflurane-induced
cognitive dysfunction in aged rats by mediating autophagy through
TLR4/MyD88/NF-κB signaling pathway
To investigate the relationship between
rapamycin-activated autophagy and the TLR4/MyD88/NF-κB signaling
pathway, the aged rats with sevoflurane-induced cognitive
dysfunction were treated with TLR4 inhibitor. It was revealed that
compared with the RAP group, LC3II/I and Beclin1 expression was
significantly lower, but p62 expression was significantly higher
after suppressing the TLR4 signaling pathway (P<0.05; Fig. 6A). ELISA was further applied to
detect inflammatory factors, and as a result, the inhibitory effect
of rapamycin on inflammatory factors was weakened. Compared with
the RAP group, IL-1β, IL-6 and TNF-α expression was significantly
higher, but IL-10 expression was significantly lower in the TLR
group (P<0.05; Fig. 6B). An MWM
experiment was conducted to determine the effects of the
TLR4/MyD88/NF-κB signaling pathway on learning and memory
abilities. In the acquisition test, the escape latency of rats in
each group was gradually shortened, and in 3–4 days, compared with
the RAP group, the escape latency was significantly increased in
the TLR group (P<0.05). In the spatial probe test, compared with
the RAP group, the percentage of time, the distance covered in the
target quadrant and the number of platform crossings were
significantly decreased in the TLR group (P<0.05; Fig. 6C). These findings indicated that by
inhibiting the TLR4 signaling pathway, the effect of rapamycin on
the activation of autophagy was weakened, and the protective effect
of rapamycin on sevoflurane-induced brain injury was markedly
weakened in the aged rats. These results confirmed that rapamycin
improves sevoflurane-induced cognitive dysfunction in aged rats by
mediating autophagy via the TLR4/MyD88/NF-κB signaling pathway.
Discussion
POCD is a reversible and fluctuating acute
neurological disorder that occurs shortly after surgery (18). POCD is mainly manifested as
postoperative mental disorder, anxiety, personality changes and
impaired memory (19). Some
studies have revealed that age was an independent risk factor for
POCD (20,21). The incidence of POCD was as high as
25% within 1 week after non-cardiac surgery in elderly patients
older than 60 years of age, and the symptoms of 10% of the patients
lasted until three months after surgery (22). The occurrence of POCD can delay the
postoperative recovery, prolong the length of hospital stay and
increase medical expenses in elderly patients, and even have a
certain impact on their long-term quality of life, which increases
the economic burden on families and society (23). In the present study, the models of
sevoflurane-induced cognitive dysfunction in the elderly in
vitro and in vivo were established, and the protective
effect of rapamycin was observed. The present results demonstrated
that rapamycin can improve learning and memory impairments in rats,
reduce the expression of inflammatory factors in rat serum,
decrease apoptosis of brain tissue, and activate autophagy in
tissues. This mechanism of action was exerted through the
TLR4/MyD88/NF-κB signaling pathway.
Rapamycin is used in the clinical treatment of organ
transplant rejection and certain cardiovascular diseases (4). Rapamycin has a therapeutic effect on
neurodegenerative diseases, such as the neuroprotective effect on
both Alzheimer's disease and Parkinson's disease (24,25).
Neurodegenerative diseases in old age are strongly associated with
age (26). Rapamycin has strong
anti-aging effects and can delay the onset of such
neurodegenerative diseases by altering age-related protein
molecules (27). Moreover, studies
have revealed that rapamycin can induce lysosome-mediated
autophagy, enhance the ability of lysosomes to clear
autophagosomes, and reduce autophagic accumulation, thereby playing
a neuroprotective role (28). By
establishing an aging cognitive dysfunction model, it was revealed
that after rapamycin pretreatment in rats, learning and memory
impairments were reduced, and the integrity of the brain tissue was
markedly less than that of the sevoflurane group; the arrangement
of neurons in the brain was neat and tight than that of the
sevoflurane group; the expression of S-100β and NSE, which is a
sign of brain tissue damage in serum, was markedly decreased, and
the expression of IL-6, IL-1β and TNF-α proinflammatory cytokines
was significantly decreased, while the expression of
anti-inflammatory cytokine IL-10 was markedly increased; the
expression of BAX and caspase-3 in the tissue was markedly reduced,
however the expression of anti-apoptotic factor Bcl-2 was
significantly increased. These findings indicated that rapamycin
exerts a protective effect on sevoflurane-induced cognitive
dysfunction both in vitro and in vivo.
Autophagy is an important mechanism for eukaryotic
cells to maintain homeostasis and survival (29). In terms of neurological diseases,
autophagy is another neuronal death mode in addition to apoptosis
and necrosis (30). Autophagy
plays roles in different diseases and plays a dual role in
neuroprotection or neuronal death (31,32).
Studies have revealed that TLR4 is an environmental receptor for
autophagy and macrophages can induce autophagy and
autophagy-mediated cell death by lipopolysaccharide (33). It was revealed that
lipopolysaccharide induces autophagy through the TRIF-dependent,
MyD88-independent TLR4 signaling pathway. Although there is in fact
a link between TLR-mediated innate immunity and autophagy, the
mechanism is still unclear. Studies have revealed that
lipopolysaccharide can induce autophagy of tubular epithelial cells
via the TLR4 pathway both in vivo and in vitro in
acute kidney injury test, counteracting endotoxin-induced renal
injury, and regulating the TLR4 downstream signaling pathway
(34). The level of autophagy was
suppressed in the isoprenaline-induced myocardial fibrosis model in
TLR4 knockout mice (35). In the
present study, it was revealed that when the TLR4/MyD88/NF-κB
signaling pathway was inhibited, the expression of LC3-II/I and
Beclin1 was significantly decreased, and the protective effect of
rapamycin was markedly weakened in older models of cognitive
dysfunction. It was further revealed that the expression of TLR4,
MyD88, and NF-κB p65 was increased compared with the rapamycin
group. These data indicated that this protective effect of
rapamycin is achieved by mediating autophagy through the
TLR4/MyD88/NF-κB p65 signaling pathway.
In conclusion, rapamycin can reduce learning and
memory impairments and inflammatory response caused by cognitive
dysfunction in the elderly in vitro and in vivo,
inhibit tissue and cell apoptosis, activate tissue and cell
autophagy, and alter the expression of TLR4, MyD88, and NF-κB p65.
These results demonstrated that rapamycin has an ameliorating
effect on sevoflurane-induced cognitive dysfunction in older age,
and this effect is achieved by mediating autophagy through the
TLR4/MyD88/NF-κB p65 signaling pathway.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Shenyang Key
Technology Research and Development plan (grant no.
17-230-9-45).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YL and JZ conceived and designed the study and
drafted the manuscript. YL, LL and YT performed experiments and
interpreted the results. YT and JZ analyzed the data. YT and JZ
contributed to acquisition of funding support. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The project was approved by the China Medical
University Institutional Animal Care and Use Committee (IACUC no.
2017018R).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Benhamou D and Brouquet A: Postoperative
cerebral dysfunction in the elderly: Diagnosis and prophylaxis. J
Visc Surg. 153:S27–S32. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Brioni JD, Varughese S, Ahmed R and Bein
B: A clinical review of inhalation anesthesia with sevoflurane:
From early research to emerging topics. J Anesth. 31:764–778. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang L, Huang K and Ning H: Autophagy
induction by hispidulin provides protection against
sevoflurane-induced neuronal apoptosis in aged rats. Biomed
Pharmacother. 98:460–468. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yoo YJ, Kim H, Park SR and Yoon YJ: An
overview of rapamycin: from discovery to future perspectives. J Ind
Microbiol Biotechnol. 44:537–553. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Maiese K: The mechanistic target of
rapamycin (mTOR) and the silent mating-type information regulation
2 homolog 1 (SIRT1): Oversight for neurodegenerative disorders.
Biochem Soc Trans. 46:351–360. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang X, Zhou Y, Xu M and Chen G:
Autophagy Is Involved in the Sevoflurane Anesthesia-Induced
Cognitive Dysfunction of Aged Rats. PLoS One. 11:e01535052016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cai Z, Zhao B, Li K, Zhang L, Li C, Quazi
SH and Tan Y: Mammalian target of rapamycin: A valid therapeutic
target through the autophagy pathway for Alzheimer's disease? J
Neurosci Res. 90:1105–1118. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nah J, Yuan J and Jung YK: Autophagy in
neurodegenerative diseases: From mechanism to therapeutic approach.
Mol Cells. 38:381–389. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gewirtz DA: The four faces of autophagy:
implications for cancer therapy. Cancer Res. 74:647–651. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Espert L, Beaumelle B and Vergne I:
Autophagy in Mycobacterium tuberculosis and HIV infections. Front
Cell Infect Microbiol. 5:492015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ntsapi C, Lumkwana D, Swart C, du Toit A
and Loos B: New insights into autophagy dysfunction related to
amyloid beta toxicity and neuropathology in alzheimer's disease.
Int Rev Cell Mol Biol. 336:321–361. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Segura-Aguilar J and Huenchuguala S:
Aminochrome Induces Irreversible Mitochondrial Dysfunction by
Inducing Autophagy Dysfunction in Parkinson's Disease. Front
Neurosci. 12:1062018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang Z, Liu D, Wang F, Liu S, Zhao S, Ling
EA and Hao A: Saturated fatty acids activate microglia via
Toll-like receptor 4/NF-κB signalling. Br J Nutr. 107:229–241.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chi OZ, Mellender SJ, Barsoum S, Liu X,
Damito S and Weiss HR: Effects of rapamycin pretreatment on
blood-brain barrier disruption in cerebral ischemia-reperfusion.
Neurosci Lett. 620:132–136. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Feng Y, Gao J, Cui Y, Li M, Li R, Cui C
and Cui J: Neuroprotective effects of resatorvid against traumatic
brain injury in rat: Involvement of neuronal autophagy and TLR4
signaling pathway. Cell Mol Neurobiol. 37:155–168. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zatroch KK, Knight CG, Reimer JN and Pang
DS: Refinement of intraperitoneal injection of sodium pentobarbital
for euthanasia in laboratory rats (Rattus norvegicus). BMC Vet Res.
13:602017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rundshagen I: Postoperative cognitive
dysfunction. Dtsch Arztebl Int. 111:119–125. 2014.PubMed/NCBI
|
|
19
|
Berger M, Nadler JW, Browndyke J, Terrando
N, Ponnusamy V, Cohen HJ, Whitson HE and Mathew JP: Postoperative
cognitive dysfunction: Minding the gaps in our knowledge of a
common postoperative complication in the elderly. Anesthesiol Clin.
33:517–550. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tang Y, Wang X, Zhang S, Duan S, Qing W,
Chen G, Ye F, Le Y and Ouyang W: Pre-existing weakness is critical
for the occurrence of postoperative cognitive dysfunction in mice
of the same age. PLoS One. 12:e01824712017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Z, Liu F, Ma H, White PF, Yumul R,
Jiang Y, Wang N and Cao X: Age exacerbates surgery-induced
cognitive impairment and neuroinflammation in sprague-dawley rats:
The role of IL-4. Brain Res. 1665:65–73. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Coburn M, Fahlenkamp A, Zoremba N and
Schaelte G: Postoperative cognitive dysfunction: Incidence and
prophylaxis. Anaesthesist. 59:177–184; quiz 185. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Silbert B, Evered L, Scott DA, McMahon S,
Choong P, Ames D, Maruff P and Jamrozik K: Preexisting cognitive
impairment is associated with postoperative cognitive dysfunction
after hip joint replacement surgery. Anesthesiology. 122:1224–1234.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li L: The molecular mechanism of
glucagon-like peptide-1 Therapy in Alzheimer's disease, based on a
mechanistic target of rapamycin pathway. CNS Drugs. 31:535–549.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jiang J, Jiang J, Zuo Y and Gu Z:
Rapamycin protects the mitochondria against oxidative stress and
apoptosis in a rat model of Parkinson's disease. Int J Mol Med.
31:825–832. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jove M, Portero-Otin M, Naudi A, Ferrer I
and Pamplona R: Metabolomics of human brain aging and age-related
neurodegenerative diseases. J Neuropathol Exp Neurol. 73:640–657.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Krzesniak M, Zajkowicz A, Matuszczyk I and
Rusin M: Rapamycin prevents strong phosphorylation of p53 on serine
46 and attenuates activation of the p53 pathway in A549 lung cancer
cells exposed to actinomycin D. Mech Ageing Dev. 139:11–21. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang Y, Ma Q, Ma X, Zhang Z, Liu N and
Wang M: Role of mammalian target of rapamycin signaling in
autophagy and the neurodegenerative process using a senescence
accelerated mouse-prone 8 model. Exp Ther Med. 14:1051–1057. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Parzych KR and Klionsky DJ: An overview of
autophagy: morphology, mechanism, and regulation. Antioxid Redox
Signal. 20:460–473. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Twayana KS and Ravanan P: Eukaryotic cell
survival mechanisms: Disease relevance and therapeutic
intervention. Life Sci. 205:73–90. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Saha S, Panigrahi DP, Patil S and Bhutia
SK: Autophagy in health and disease: A comprehensive review. Biomed
Pharmacother. 104:485–495. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Luo T, Liu G, Ma H, Lu B, Xu H, Wang Y, Wu
J, Ge P and Liang J: Inhibition of autophagy via activation of
PI3K/Akt pathway contributes to the protection of ginsenoside Rb1
against neuronal death caused by ischemic insults. Int J Mol Sci.
15:15426–15442. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen S, Yuan J, Yao S, Jin Y, Chen G, Tian
W, Xi J, Xu Z, Weng D and Chen J: Lipopolysaccharides may aggravate
apoptosis through accumulation of autophagosomes in alveolar
macrophages of human silicosis. Autophagy. 11:2346–2357. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nair AR, Masson GS, Ebenezer PJ, Del Piero
F and Francis J: Role of TLR4 in lipopolysaccharide-induced acute
kidney injury: protection by blueberry. Free Radic Biol Med.
71:16–25. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Dong RQ, Wang ZF, Zhao C, Gu HR, Hu ZW,
Xie J and Wu YQ: Toll-like receptor 4 knockout protects against
isoproterenol-induced cardiac fibrosis: The role of autophagy. J
Cardiovasc Pharmacol Ther. 20:84–92. 2015. View Article : Google Scholar : PubMed/NCBI
|