Introduction
Ankylosing spondylitis (AS) is a chronic
inflammatory rheumatic disease that is characterized by pain and
stiffness between the spinal bones. AS mainly affects the axial
skeleton, leading to back pain, progressive structural and
functional impairment and reduced quality of life (1). It has been recognized that HLA-B27 is
the most important risk factor for AS (2). The prevalence of AS is ~0.20–0.40% in
China (3–5). The patients are predominantly young
adults, with more male patients than female patients, and the
disability rate 5 years after the onset of symptoms is 40–60%
(3,4,6,7).
Genome-wide association studies (GWAS) have investigated many genes
related with AS, however, to date, the pathogenesis of AS has not
been well elucidated. Recently, it was reported that aberrant
apoptosis of T cells was observed in peripheral blood and
inflammatory joints of AS patients.
MicroRNAs (miRNAs) are a large family of highly
conserved endogenous small non-coding RNAs that are ~21–23
nucleotides in length in eukaryotes (8). miRNAs do not encode proteins, and
they can inhibit the expression of multiple target genes by binding
to the 3 prime-untranslated region (3′-UTR) of target mRNAs
(9–11). In addition, miRNAs
post-transcriptionally regulate the translation and degradation of
their target genes. miRNAs are associated with inflammatory
response and the development of autoimmune diseases, for example,
rheumatoid arthritis (12), and
multiple sclerosis (13). In
recent years, research on miRNAs in AS has received more attention.
Lai et al reported the first research on miRNA expression
profiling in AS from 5 AS patients and 5 healthy controls (14). Their study revealed that the
expression of 8 miRNAs such as miR-150, miR-342-5p, miR-16,
miR-221, miR-99b, let-7b, let-7i, and miR-513-5p were upregulated
and 5 miRNAs such as miR-218, miR-30e, miR-199a-5p, miR-409-3p and
miR-215 were downregulated in the T cells of AS compared with their
normal counterparts. A recent study demonstrated that deacetylase
(HDAC) 3-mediated suppression of miR-130a in peripheral blood
mononuclear cells (PBMCs) from AS patients was involved in
increased expression of its target tumor necrosis factor (TNF)-α
(15). Moreover, that study
revealed that miR-130a was significantly downregulated in immune
cells in AS patients. These results indicated that miR-130a-3p may
play an important role in the development of AS. However, the
function and downstream molecular pathways of miR-130a-3p during
the pathogenesis of AS are unclear.
The HOX gene encodes a family of highly conserved
transcription factors. The gene contains a helix-turn-helix DNA
binding domain that has 60 amino acids, and it plays a crucial role
in growth and development, regulating many biological processes
including apoptosis, receptor signaling, differentiation, movement
and angiogenesis (16). It has
been reported that HOXB1, as a tumor suppressor gene, could be
regulated by miR-3175 in glioma (17). However, the role of HOXB1 in AS
remains unclear. In the present study, through bioinformatics
software analysis the binding site between miR-130a-3p and HOXB1
was revealed. Therefore, we surmised that miR-130a-3p may play a
role in regulating T-cell survival by regulating HOXB1.
In the present study, we aimed to confirm the
expression of miR-130a-3p in T cells of AS patients and to further
explore its effects on T cells, to elucidate whether miR-130a-3p
participates in the development of AS by regulating T-cell
survival. These findings may provide insight for the development of
new approaches to manage AS.
Materials and methods
Study population
In total, 30 paired blood samples were collected
from 30 HLA-B27-positive AS patients (female to male ratio, 1:4;
age range, 22–61 years) and 30 HLA-B27-negative healthy subjects
(female to male ratio, 1:4; age range, 19–58 years) who had no
history of autoimmune diseases in our hospital between December
2015 and February 2018. Blood samples were stored at −80°C until
use. The present study was approved by the Ethics Committee of
Puyang Oilfield General Hospital (Puyang, China), and written
informed consent was obtained from every patient.
T-cell isolation and culture
Venous blood from AS patients and healthy volunteers
was collected into tubes containing EDTA, from which PBMCs were
isolated by Ficoll-Hypaque density gradient centrifugation at 300 ×
g and 4°C for 10 min. T cells were further purified using magnetic
beads coated with anti-human CD33 (Miltenyi Biotec GmbH, Gladbach,
Germany). The T cells were cultured in RPMI-1640 medium (Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% FBS (Gibco; Thermo Fisher Scientific, Inc.), 2 mM L-glutamine
(Beyotime Institute of Biotechnology, Haimen, China), 100 U/ml
penicillin (Beyotime Institute of Biotechnology), and 100 U/ml
streptomycin (Beyotime Institute of Biotechnology) at 37°C, 5%
CO2 incubator.
Cell culture and cell
transfection
The human Jurkat cells were purchased from Shanghai
Institute of Life Sciences, Chinese Academy of Sciences (Shanghai,
China) and cultured in Dulbecco's modified Eagle's medium (DMEM)
supplemented with 10% FBS, and incubated at 37°C, in 5%
CO2 incubator. Jurkat T cells were transfected with 50
nM inhibitor control (5′-CAGUACUUUUGUGUAGUACAA-3′), 50 nM
miR-130a-3p inhibitor (5′-AUGCCCUUUUAACAUUGCACUG-3), 50 nM mimic
control (5′-UUCUCCGAACGUGUCACGUTT-3′), 50 nM miR-130a-3p mimic
(5′-CAGUGCAAUGUUAAAAGGGCAU-3′; all from Guangzhou RiboBio Co.,
Ltd., Guangzhou, China), 10 µM control-plasmid, 10 µM
HOXB1-plasmid, or miR-130a-3p mimic+HOXB1-plasmid using
Lipofectamine 2000 reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) following the manufacturer's instructions. Cell transfection
efficiency was detected 48 h after transfection.
RT-qPCR
Total RNA from blood/cells was extracted using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. The RNA concentration
was detected by NanoDrop 2000. RT-qPCR was carried out using 2X
SYBR-Green PCR Master Mix (Applied Biosystems; Thermo Fisher
Scientific, Inc.). Reaction conditions were as follows: 10 min at
95°C followed by 35 cycles of 15 sec at 95°C and 40 sec at 55°C.
The primer sequences used for RT-qPCR are listed in Table I. The relative gene expression
levels were calculated using the 2−∆∆Cq method (18) after normalization with reference
the expression of GAPDH or U6. All experiments were performed in
triplicate.
 | Table I.Primer sequences for PCR. |
Table I.
Primer sequences for PCR.
| Gene | Sequence
(5′-3′) |
|---|
| Bcl-2 | F:
5′-TTGGATCAGGGAGTTGGAAG-3′ |
|
| R:
5′-TGTCCCTACCAACCAGAAGG-3′ |
| Bax | F:
5′-CGTCCACCAAGAAGCTGAGCG-3′ |
|
| R:
5′-CGTCCACCAAGAAGCTGAGCG-3′ |
| HOXB1 | F:
5′-CGTCCACCAAGAAGCTGAGCG-3′ |
|
| R:
5′-CGTCCACCAAGAAGCTGAGCG-3′ |
| miR-130a-3p | F:
5′-GGCAGTGCAATGTTAAAAG-3′ |
|
| R:
5′-CAGTGCGTGTCGTGGAGT-3′ |
| U6 | F:
5′-CTCGCTTCGGCAGCACA-3′ |
|
| R:
5′-AACGCTTCACGAATTTGCGT-3′ |
| GAPDH | F:
5′-CTTTGGTATCGTGGAAGGACTC-3′ |
|
| R:
5′-GTAGAGGCAGGGATGATGTTCT-3′ |
Western blot analysis
Total proteins were extracted from cells using
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology), according to the manufacturer's protocol. A
bicinchoninic acid protein assay kit (Thermo Fisher Scientific,
Inc.) was used to analyze the protein concentration. The protein
samples (35 µg/lane) were separated on 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gel and
transferred to polyvinylidene fluoride (PVDF) membranes. The
membranes were blocked with 5% non-fat milk for 1.5 h, and were
incubated overnight at 4°C with the following primary antibodies:
β-actin (1:1,000; cat. no. 4970; Cell Signaling Technology, Inc.),
HOXB1 (1:1,000; cat no. ab168279; Abcam), Bcl-2 (1:1,000; cat. no.
4223; Cell Signaling Technology, Inc.) and Bax (1:1,000; cat. no.
5023; Cell Signaling Technology, Inc.). Subsequently, the membranes
were incubated with the anti-rabbit immunoglobulin G horseradish
peroxidase-conjugated secondary antibody (1:5,000; cat no. 7074;
Cell Signaling Technology, Inc.) for 2 h at room temperature.
Proteins were detected using an ECL kit (Thermo Fisher Scientific,
Inc.) and imaged. β-actin was used as an internal control, and the
AlphaView 3.4.0 (ProteinSimple, San Jose, CA, USA) software was
used for quantification analysis.
CCK-8 assay
Cell Counting Kit-8 (CCK-8) assay was performed to
assess the cell proliferation ability (16,19).
Logarithmic phase cells were seeded in a 96-well plate with
1×104 cells/well and incubated at 37°C with 5%
CO2 for 12 h, after which 10 µl CCK-8 solution was added
to each well, and the cells were incubated for a further 2 h at
37°C with 5 % CO2. The absorbance was measured at a
wavelength of 450 nm using a microplate reader.
Flow cytometric assay
Jurkat T cells were transfected with inhibitor
control, miR-130a-3p inhibitor, mimic control, miR-130a-3p mimic,
or miR-130a-3p mimic+HOXB1-plasmid for 48 h. Then, Jurkat T cells
were collected at logarithmic growth phase, and Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) apoptosis
detection kit (cat. no. 70-AP101-100; MultiSciences Biotech Co.,
Ltd., Hangzhou, China) was used to analyze cell apoptosis. Briefly,
Jurkat T cells were stained with 5 µl Annexin V- FITC and 5 µl PI
for 30 min at room temperature in the dark. A flow cytometer (BD
Biosciences, Franklin Lakes, NJ, USA) was used to perform analysis
of cell apoptosis.
Dual-luciferase reporter assay
Bioinformatics software TargetScan 7.2 (http://www.targetscan.org/vert_72/) was used to
predict target genes of miR-130a-3p (20), and the results revealed the binding
sites between miR-130a-3p and HOXB1. To investigate the
relationship between miR-130a-3p and HOXB1, a luciferase reporter
vector that contained a 3′-UTR sequence of HOXB1 was constructed.
Cells seeded in 24-well plates were co-transfected with miR-130a-3p
mimic or mimic control and the mutant (MUT) or wild-type (WT)
3′-UTR of HOXB1 using Lipofectamine 2000 for 48 h, along with
Renilla luciferase pRL-TK vector as a control. After
transfection for 48 h, the cells were lysed with RIPA buffer. The
relative luciferase activity was detected using a Dual-Luciferase
Reporter Assay System (Promega Corporation, Madison, WI, USA)
according to the manufacturer's instructions.
Statistical analyses
Each experiment was performed at least three times.
Data analyses were performed using GraphPad Prism 6.0 (GraphPad
Software, Inc., La Jolla, CA, USA). All data were expressed as the
mean ± standard deviation (SD). The significance of differences
between groups was carried out by one-way analysis of variance with
Tukey's post hoc test or Student's t-test. P<0.05 was considered
to indicate a statistically significant difference.
Results
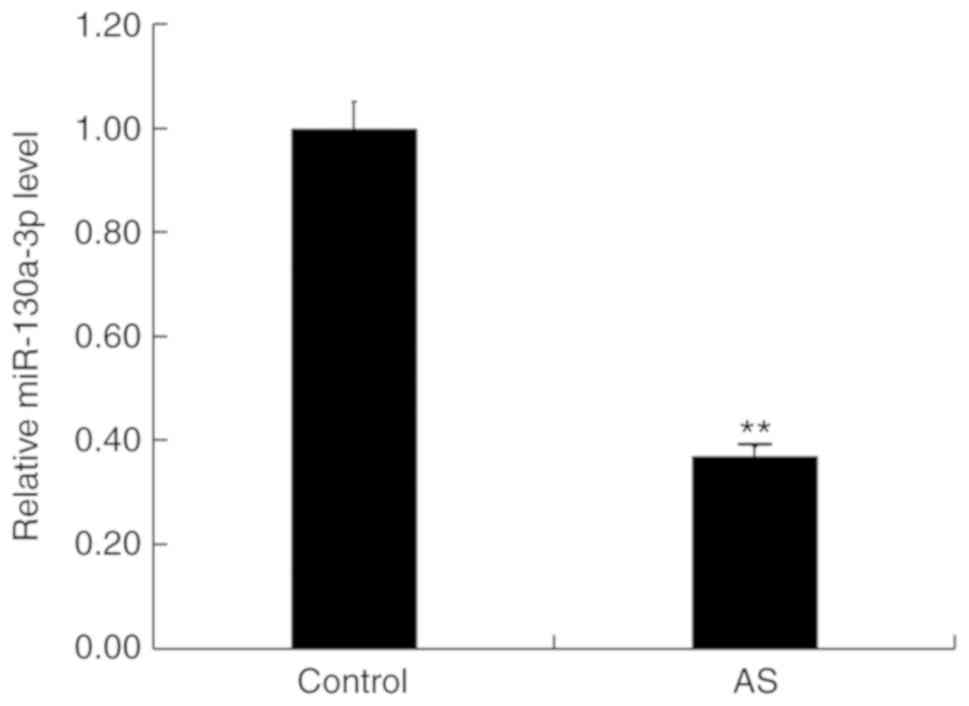
Expression level of miR-130a-3p in T
cells from AS patients
RT-qPCR was firstly performed to detect the level of
miR-130a-3p in T cells in HLA-B27-positive AS patients and
HLA-B27-negative healthy subjects. The results revealed that the
expression level of miR-130a-3p was significantly downregulated in
T cells in HLA-B27-positive AS patients compared with HLA-B27
negative healthy controls (Fig.
1).
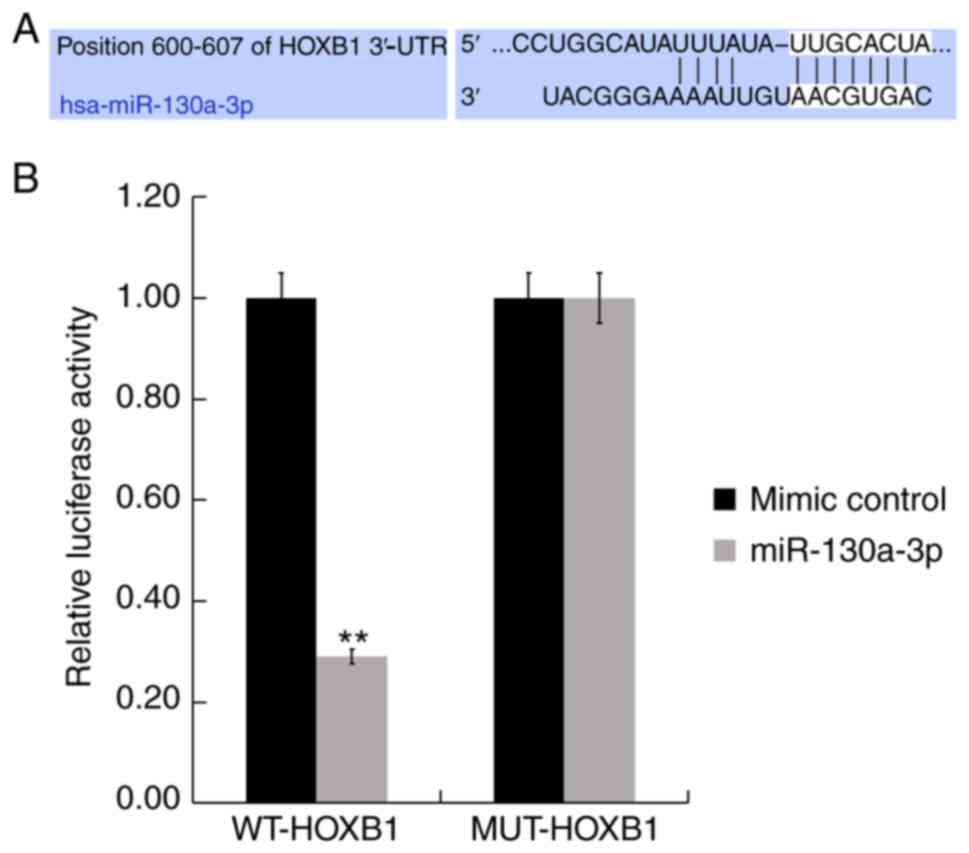
HOXB1 is a direct target of
miR-130a-3p
To identify the functional targets of miR-130a-3p,
TargetScan was used. TargetScan revealed that miR-130a-3p has
hundreds of target genes, including HOXB1 (Fig. 2A). HOXB1 regulates many biological
processes including receptor signaling, differentiation, movement,
angiogenesis, cell proliferation and apoptosis. However, the role
of HOXB1 in AS remains largely unclear, therefore, HOXB1 was
selected for further study. Furthermore, to further examine whether
miR-130a-3p directly targets HOXB1, Luc HOXB1-3′-UTR-WT and its
3′-UTR MUT plasmids were constructed. A luciferase reporter assay
revealed that miR-130a-3p mimic significantly suppressed the
luciferase activity of the wild-type HOXB1 3′-UTR (Fig. 2B). These results provided evidence
that HOXB1 was a direct target of miR-130a-3p.
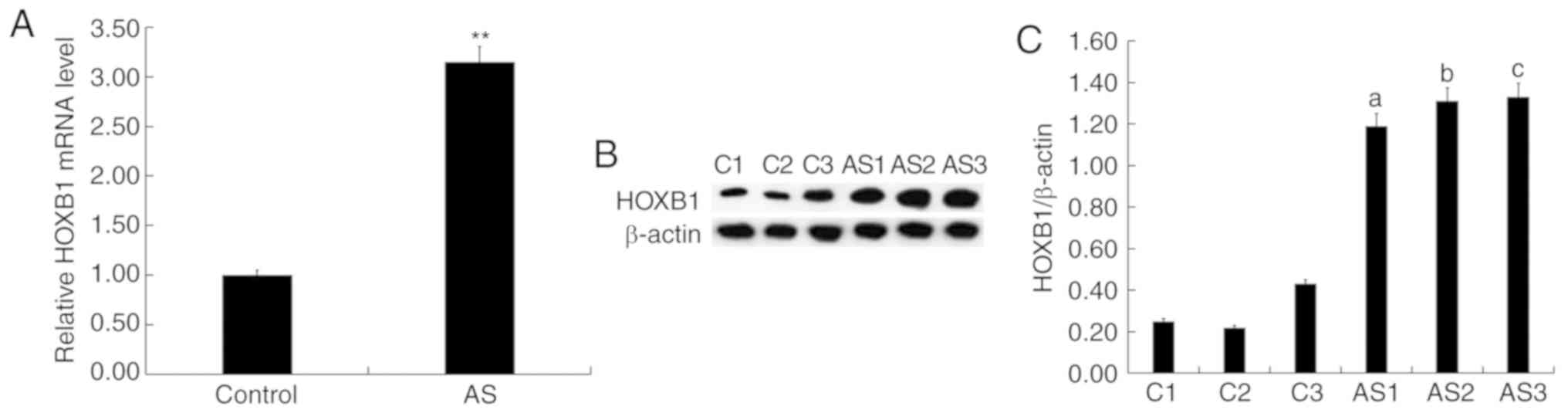
Expression level of HOXB1 in T cells
from AS patients
Next, the expression level of HOXB1 in T cells in
HLA-B27-positive AS patients and HLA-B27 negative healthy subjects
was detected by RT-qPCR and western blot assays. The results
revealed that both the mRNA and protein levels of HOXB1 in T cells
in HLA-B27-positive AS patients were higher than that in the
HLA-B27 negative healthy subjects (Fig. 3).
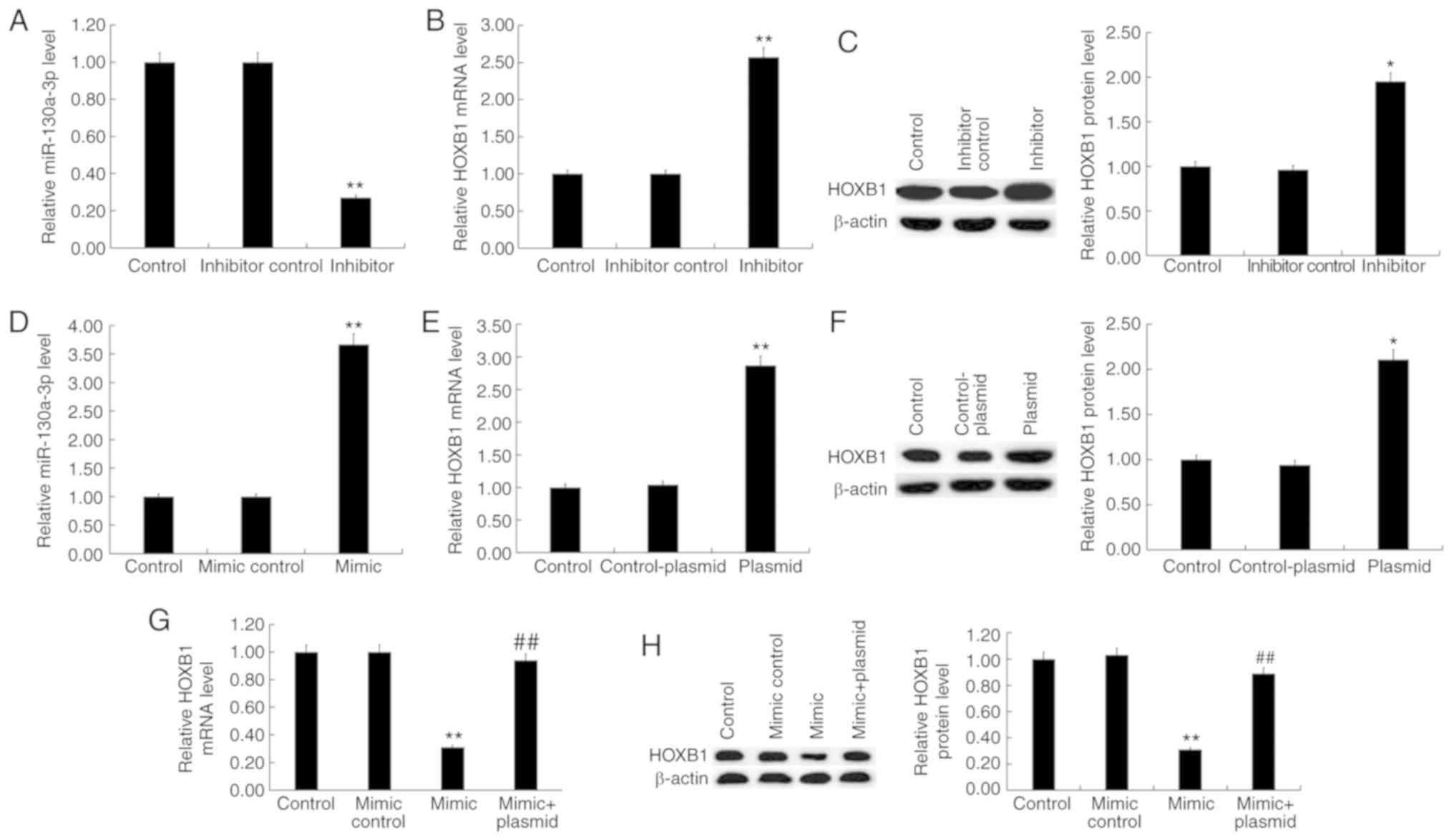
Expression level of miR-130a-3p and
HOXB1 in Jurkat T cells after transfection
To determine whether miR-130a-3p influenced T cell
proliferation and apoptosis, Jurkat T cells were transfected with
inhibitor control, miR-130a-3p inhibitor, mimic control,
miR-130a-3p mimic, control-plasmid, HOXB1-plasmid, or miR-130a-3p
mimic+HOXB1-plasmid for 48 h. RT-qPCR or and western blot assays
were performed to detect transfection efficiency of the mimic or
plasmid. RT-qPCR results revealed that miR-130a-3p inhibitor
significantly decreased the expression of miR-130a-3p in Jurkat T
cells, and it significantly upregulated the mRNA and protein levels
of HOXB1 (Fig. 4A-C). In addition,
miR-130a-3p mimic significantly increased the expression of
miR-130a-3p in Jurkat T cells, and it decreased the mRNA and
protein levels of HOXB1 (Fig. 4D, G
and H). RT-qPCR and western blot analyses also indicated that
HOXB1-plasmid significantly improved the expression level of HOXB1
in Jurkat T cells (Fig. 4E and
F).
Effect of miR-130a-3p on proliferation
and apoptosis of Jurkat T cells
In order to shed light on the function of
miR-130a-3p in T cells, the effect of miR-130a-3p on the
proliferation of Jurkat T cells was first investigated. CCK-8 assay
results revealed that compared with the control group, miR-130a-3p
inhibitor significantly inhibited the cell proliferation activity
of Jurkat T cells, while miR-130a-3p mimic significantly improved
cell proliferation activity, and this increase was reversed by
HOXB1-plasmid (Fig. 5A). To
further determine whether miR-130a-3p regulated apoptosis, flow
cytometry was performed to detect cell apoptosis. Flow cytometric
analysis revealed that miR-130a-3p inhibitor significantly induced
the cell apoptosis of Jurkat T cells, while miR-130a-3p mimic
significantly decreased apoptosis, and this decrease was reversed
by HOXB1-plasmid (Fig. 5B).
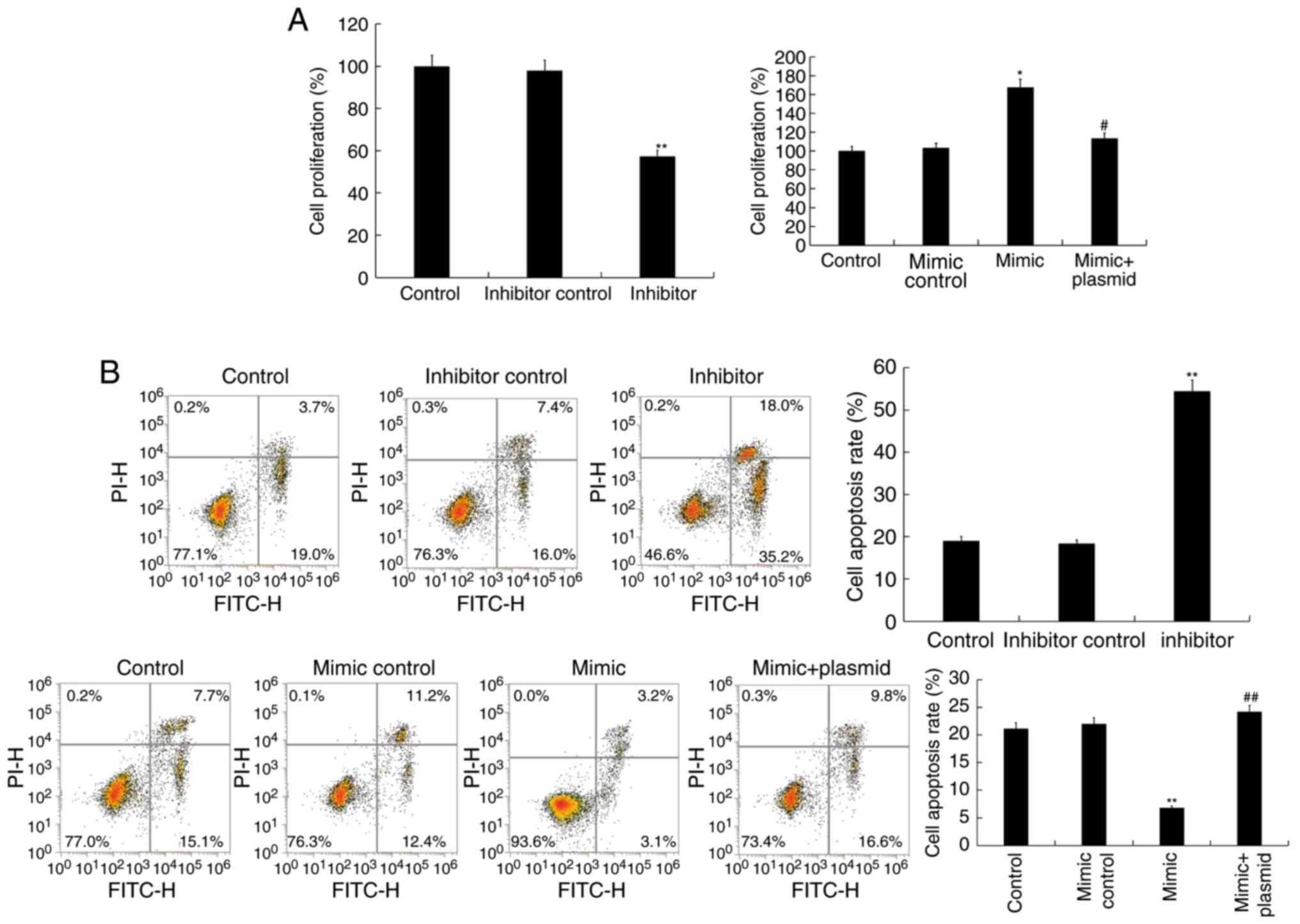 | Figure 5.Effect of miR-130a-3p on cell
proliferation and cell apoptosis of Jurkat T cells. (A) A CCK-8
assay was used to detect cell proliferation ability 48 h after
Jurkat T cells were transfected with inhibitor control, miR-130a-3p
inhibitor, mimic control, miR-130a-3p mimic, or miR-130a-3p
mimic+HOXB1-plasmid. (B) A flow cytometric assay was used to detect
cell apoptosis 48 h after Jurkat T cells were transfected with
inhibitor control, miR-130a-3p inhibitor, mimic control,
miR-130a-3p mimic, or miR-130a-3p mimic+HOXB1-plasmid. Data are
displayed as the mean ± SD. *P<0.05 and **P<0.01 vs. the
control group; #P<0.05 and ##P<0.01 vs.
the mimic group. CCK-8, Cell Counting Kit-8; SD, standard
deviation. |
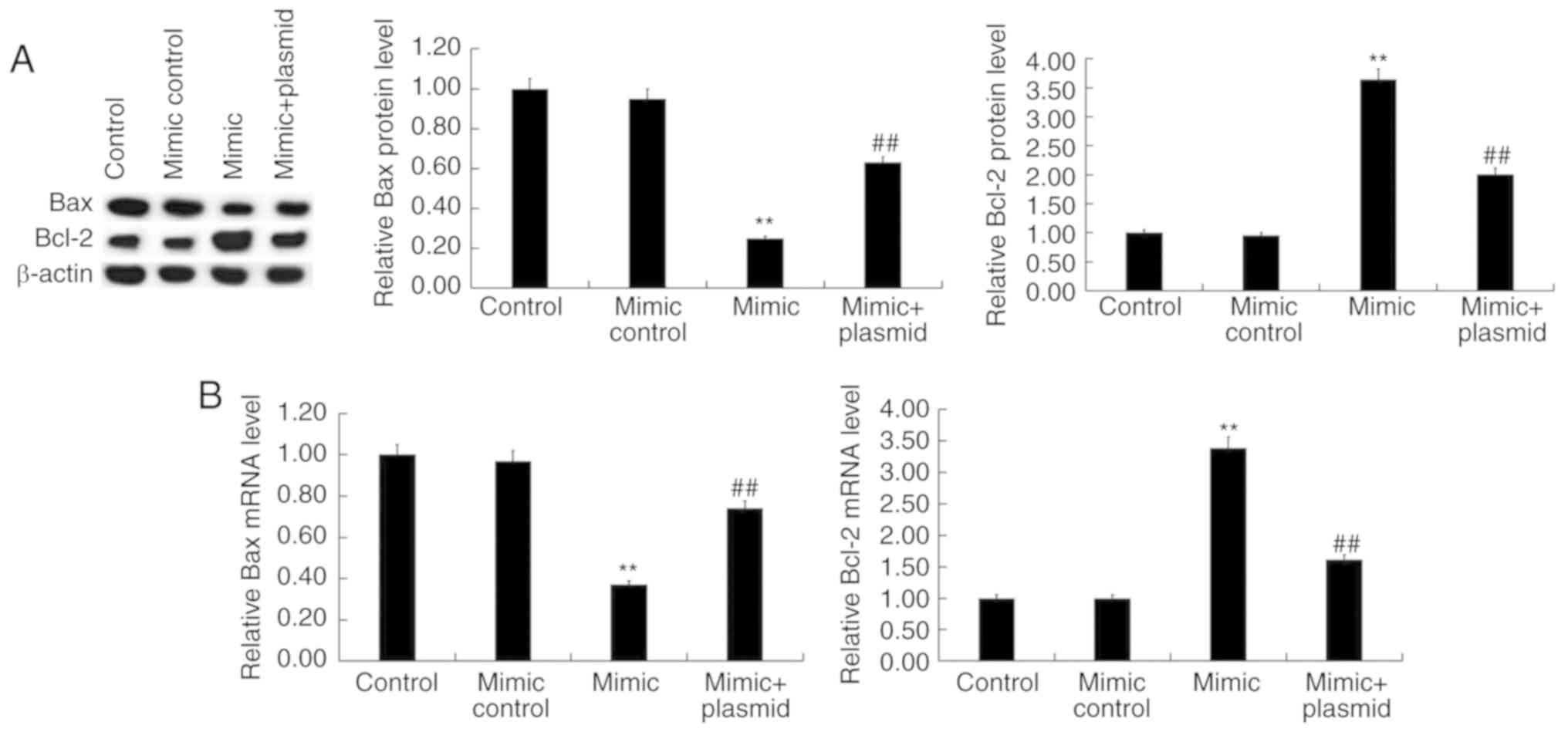
In addition, western blot analysis and RT-qPCR
revealed that miR-130a-3p mimic significantly decreased Bax
expression and increased B-cell lymphoma 2 (Bcl-2) expression at
both the protein (Fig. 6A) and
mRNA (Fig. 6B) level, which were
reversed by HOXB1-plasmid.
Discussion
It has been confirmed that the pathogenesis of
ankylosing spondylitis (AS) is closely related to HLA-B27.
Furthermore, AS is a genetically heritable disease. There is
increasing research suggesting the involvement of T cells in AS
(21,22). At present, there is still no cure
method for AS treatment, thus it is urgent to find an effective
target to cure AS clinically.
Wang et al demonstrated that miR-199a-5p
could inhibit the pathogenesis of AS by targeting Ras homolog
enriched in brain (Rheb) (23). In
addition, miR-199a-5p was expressed at a low level in T cells from
AS patients and could induce autophagy of T cells. Hou et al
indicated that miR-let-7i could induce cell autophagy to protect T
cells from apoptosis by targeting IGF1R (24). In the present study, it was
revealed that miR-130a-3p was downregulated in T cells in
HLA-B27-positive patients compared with the healthy controls.
In the study, it was determined that HOXB1 was a
direct target gene of miR-130a-3p, and HOXB1 was upregulated in AS.
Next, to investigate the relationship between miR-130a-3p and
HOXB1. Jurkat T cells were transfected with inhibitor control,
miR-130a-3p inhibitor, mimic control, miR-130a-3p mimic, or
miR-130a-3p mimic+HOXB1-plasmid. The results revealed that
miR-130a-3p negatively regulated the expression of HOXB1 in Jurkat
T cells.
Numerous T cells have been revealed to be required
for immune response in AS (25).
There are increasing findings suggesting the involvement of T cells
in AS (17,19). Recent studies revealed that
disordered T cells were observed in peripheral blood and
inflammatory joints from patients with AS (26,27).
A previous study indicated that let-7i played an important role in
AS development by controlling T-cell fates by targeting IGF1R
(22). In the present study, it
was thus determined whether miR-130a-3p could affect T-cells fates,
and cell proliferation and cell apoptosis was then analyzed. The
findings of our present study indicated that miR-130a-3p
downregulation inhibited cell proliferation ability and induced
cell apoptosis of Jurkat T cells, while miR-130a-3p overexpression
significantly promoted cell proliferation and inhibited cell
apoptosis. The B-cell lymphoma 2 gene (Bcl-2) is a member of a
family of proteins whose major function is their involvement in the
initiation phase of the intrinsic pathways of apoptosis (28,29).
Bcl-2 has been demonstrated to be associated with apoptosis
(30). Bcl-2-like protein 4 (Bax)
is a pro-apoptotic protein of the Bcl-2 family of proteins
(31). However, HOXB1 has been
reported to regulate the Bax/Bcl-2 ratio thus participating in the
regulation of cell apoptosis (32). The effect of miR-130a-3p on Bcl-2
and Bax expression was then studied in Jurkat T cells. The results
revealed that miR-130a-3p mimic significantly reduced Bax
expression and increased Bcl-2 expression at both the protein and
mRNA levels. It is worth mentioning that all the effects of
miR-130a-3p mimic on Jurkat T cells were eliminated by HOXB1
overexpression. In recent years, miRNA target protectors have been
investigated for more in-depth research of the physiological roles
of specific miRNA-mRNA pairs (33,34).
In the present study, the HOXB1 plasmid was used to increase the
expression of HOXB1, which was decreased by the miR-130a-3p mimic.
Whether miRNA target protectors can specifically block the
inhibition of HOXB1 by miR130a-3p requires further
investigation.
In summary, our data revealed that miR-130a-3p was
decreased in T cells of HLA-B27-positive AS patients. Furthermore,
it was revealed that HOXB1 was a direct target gene of miR-130a-3p,
and HOXB1 was upregulated in T cells of HLA-B27-positive AS
patients. In addition, our data suggested that miR-130a-3p
overexpression could improve Jurkat T cell proliferation and
inhibit cell apoptosis, which were reversed by HOXB1-plasmid.
Collectively, the present study suggested that miR-130a-3p may
affect T-cell fate in AS, and thus participate in AS development.
However, this is a preliminary study of the role of miR-130a-3p in
AS. In addition, there were some limitations in the present study:
i) The expression of miR-130a-3p in HLA-B27-positive patients and
HLA-B-27-negative patients with was not compared; the role of HOXB1
in T-cell survival was not investigated. To ascertain the role of
miR-130a-3p in AS, more research is still required. For example,
the expression of miR-130a-3p in HLA-B27-negative AS patients and
HLA-B27 positive healthy donors should be determined. The role of
HOXB1 alone in T-cell survival should also be addressed. The
underlying mechanism of how T-cell reduction contributes to AS
development requires further study. In the future, these issues
will be addressed.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The data sets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
FL designed the study, drafted the manuscript and
performed literature search. FL, DS and XG collected the data and
performed statistical analysis. NG, DL, LZ, XJ and JM performed
statistical analysis and interpreted the data. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by The Ethics
Committee of Puyang Oilfield General Hospital (Puyang, China) and
written informed consent was obtained from each patient.
Patient consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests.
References
|
1
|
Dougados M and Baeten D:
Sphodyloarthritis. Lancet. 377:2127–2137. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Evans DM, Spencer CC, Pointon JJ, Su Z,
Harvey D, Kochan G, Oppermann U, Dilthey A, Pirinen M, Stone MA, et
al: Interaction between ERAP1 and HLA-B27 in ankylosing spondylitis
implicates peptide handling in the mechanism for HLA-B27 in disease
susceptibility. Nat Genet. 43:761–767. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hou ZD, Xiao ZY, Gong Y, Zhang YP and Zeng
QY: Arylamine N-acetyltransferase polymorphisms in Han Chinese
patients with ankylosing spondylitis and their correlation to the
adverse drug reactions to sulfasalazine. BMC Pharmacol Toxicol.
15:64–80. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu Z, Zhang P and Dong J: Genetic
variants of STAT4 are associated with ankylosing spondylitis
susceptibility and severity in a Chinese Han population. Int J Clin
Exp Med. 7:5877–5881. 2014.PubMed/NCBI
|
|
5
|
Dean LE, Jones GT, MacDonald AG, Downham
C, Sturrock RD and Macfarlane GJ: Global prevalence of ankylosing
spondylitis. Rheumatology (Oxford). 53:650–657. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Haywood KL, Packham JC and Jordan KP:
Assessing fatigue in ankylosing spondylitis: The importance of
frequency and severity. Rheumatology. 53:552–556. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gan FY, Fei YY, Li MT, Wang Q, Xu D, Hou
Y, Zeng XF and Zhang FC: The characteristics of patients having
ankylosing spondylitis associated with Takayasu's arteritis. Clin
Rheumatol. 33:355–358. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li X, Liu F, Lin B, Luo H, Liu M, Wu J, Li
C, Li R, Zhang X, Zhou K and Ren D: miR-150 inhibits proliferation
and tumorigenicity via retarding G1/S phase transition in
nasopharyngeal carcinoma. Int J Oncol. 50:1097–1108. 2017.
View Article : Google Scholar :
|
|
9
|
Ro S, Park C, Young D, Sanders KM and Yan
W: Tissue-dependent paired expression of miRNAs. Nucleic Acids Res.
35:5944–5953. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mallory AC and Vaucheret H: MicroRNAs:
Something important between the genes. Curr Opin Plant Biol.
7:120–125. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Garzon R, Calin GA and Croce CM: MicroRNAs
in cancer. Annu Rev Med. 60:167–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gaur N, Karouzakis E, Gluck S, Bagdonas E,
Jüngel A, Michel BA, Gay RE, Gay S, Frank-Bertoncelj M and Neidhart
M: Micrornas interfere with DNA methylation in rheumatoid arthritis
synovial fibroblasts. RMD Open. 2:e0002992016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Freiesleben S, Hecker M, Zettl UK, Fuellen
G and Taher L: Analysis of microRNA and gene expression profiles in
multiple sclerosis: Integrating interaction data to uncover
regulatory mechanisms. Sci Rep. 6:345122016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lai NS, Yu HC, Chen HC, Yu CL, Huang HB
and Lu MC: Aberrant expression of microRNAs in T cells from
patients with ankylosing spondylitis contributes to the
immunopathogenesis. Clin Exp Immunol. 173:47–57. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jiang Y and Wang L: Role of histone
deacetylase 3 in ankylosing spondylitis via negative feedback loop
with microRNA-130a and enhancement of tumor necrosis factor-1α
expression in peripheral blood mononuclear cells. Mol Med Rep.
13:35–40. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shah N and Sukumar S: The Hox genes and
their roles in oncogenesis. Nat Rev Cancer. 10:361–371. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Han L, Liu D, Li Z, Tian N, Han Z, Wang G,
Fu Y, Guo Z, Zhu Z, Du C and Tian Y: HOXB1 Is a Tumor Suppressor
Gene Regulated by miR-3175 in Glioma. PLoS One. 10:e01423872015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the (-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Su JR, Kuai JH and Li YQ: Smoc2
potentiates proliferation of hepatocellular carcinoma cells via
promotion of cellcycle progression. World J Gastroenterol.
22:10053–10063. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang Y, Wang W, Liu ZY, Xie Y, Qian Y and
Cai XN: Overexpression of miR-130a-3p/301a-3p attenuates high
glucose-induced MPC5podocyte dysfunction through suppression of
TNF-α signaling. Exp Ther Med. 15:1021–1028. 2018.PubMed/NCBI
|
|
21
|
Appel H, Maier R, Bleil J, Hempfing A,
Loddenkemper C, Schlichting U, Syrbe U and Sieper J: In situ
analysis of interleukin-23-and interleukin-12-positive cells in the
spine of patients with ankylosing spondylitis. Arthritis Rheum.
65:1522–1529. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Syrbe U, Scheer R, Wu P and Sieper J:
Differential synovial Th1 cell reactivity towards Escherichia coli
antigens in patients with ankylosing spondylitis and rheumatoid
arthritis. Ann Rheum Dis. 71:1573–1576. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Y, Luo J, Wang X, Yang B and Cui L:
MicroRNA-199a-5p induced autophagy and inhibits the pathogenesis of
ankylosing spondylitis by modulating the mTOR signaling via
directly targeting ras homolog enriched in brain (Rheb). Cell
Physiol Biochem. 42:24812017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hou C, Zhu M, Sun M and Lin Y: MicroRNA
let-7i induced autophagy to protect T cell from apoptosis by
targeting IGF1R. Biochem Biophys Res Commun. 453:728–734. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zou J, Appel H, Rudwaleit M, Thiel A and
Sieper J: Analysis of the CD8+ T cell response to the G1 domain of
aggrecan in ankylosing spondylitis. Ann Rheum Dis. 64:722–729.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wu Y, Ren M, Yang R, Liang X, Ma Y, Tang
Y, Huang L, Ye J, Chen K, Wang P and Shen H: Reduced
immunomodulation potential of bone marrow-derived mesenchymal stem
cells induced CCR4+CCR6+ Th/Treg cell subset imbalance in
ankylosing spondylitis. Arthritis Res Ther. 13:R292011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nistala K, Moncrieffe H, Newton KR,
Varsani H, Hunter P and Wedderburn LR: Interleukin-17-producing T
cells are enriched in the joints of children with arthritis, but
have a reciprocal relationship to regulatory T cell numbers.
Arthritis Rheum. 58:875–887. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Reed JC: Bcl-2 and the regulation of
programmed cell death. J Cell Biol. 124:1–6. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hatok J and Racay P: Bcl-2 family
proteins: Master regulators of cell survival. Biomol Concepts.
7:259–270. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Thomas S, Quinn BA, Das SK, Dash R, Emdad
L, Dasgupta S, Wang XY, Dent P, Reed JC, Pellecchia M, et al:
Targeting the Bcl-2 family for cancer therapy. Expert Opin Ther
Targets. 17:61–75. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gross A, Jockel J, Wei MC and Korsmeyer
SJ: Enforced dimerization of BAX results in its translocation,
mitochondrial dysfunction and apoptosis. EMBO J. 17:3878–3885.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Petrini M, Felicetti F, Bottero L, Errico
MC, Morsilli O, Boe A, De Feo A and Carè A: HOXB1 restored
expression promotes apoptosis and differentiation in the HL60
leukemic cell line. Cancer Cell Int. 13:1012013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Staton AA and Giraldez AJ: Use of target
protector morpholinos to analyze the physiological roles of
specific miRNA-mRNA pairs in vivo. Nat Protoc. 6:2035–2049. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Knauss JL, Bian S and Sun T: Plasmid-based
target protectors allow specific blockade of miRNA silencing
activity in mammalian developmental systems. Front Cell Neurosci.
7:1632013. View Article : Google Scholar : PubMed/NCBI
|