Introduction
Renal cell carcinoma (RCC), a urinary tract
malignancy originating from the renal epithelium, is among the 10
most common types of cancer globally, accounting for ~100,000
deaths annually (1,2). Partial nephrectomy is generally used
as an effective treatment for patients with early-stage RCC;
however, ~1/3 of patients have already progressed to distant
metastases at the time of initial diagnosis, resulting in a poor
prognosis (3). The 5-year survival
rate is only 10%, and the average survival time is <1 year
(3–6). There are limited options for the
treatment of patients with advanced or metastatic disease;
therefore, it is necessary to identify novel and reliable
biomarkers to target for the development of novel therapeutic
strategies for RCC.
Long noncoding RNAs (lncRNAs), a group of noncoding
RNAs >200 nucleotides in length, are involved in the regulation
of gene expression in addition to diverse biological processes,
including cell growth, migration, invasion and apoptosis (7–9).
Increasing evidence has indicated that lncRNAs exert important
functions in numerous diseases, including tumorigenesis and tumor
progression via epigenetic, transcriptional and
post-transcriptional approaches (10–12).
For example, lnc-ATB has been reported to regulate the development
of tumors (13–15). Lei et al (16) revealed that lnc-ATB is upregulated
in gastric cancer and acts as a pro-oncogene in gastric cancer
tumorigenesis via the regulation of a microRNA
(miRNA/miR)-141-3p/transforming growth factor β2 feedback loop.
lnc-ATB has also been reported to be involved in the growth and
invasion of RCC cells (14).
Additionally, there are numerous lncRNAs that are associated with
the tumorigenesis and progression of RCC; for example, the lncRNA
HOX transcript antisense RNA improves the migratory and invasive
abilities of RCC cells in vitro by upregulating the histone
H3K27 demethylase JMJD3 (5).
Silencing DLX6 antisense RNA 1 inhibits the growth of RCC cells by
regulating the miR-26a/PTEN axis (17).
The lncRNA deleted in lymphocytic leukemia 1 (DLEU1)
is located on chromosome 13q14.3, and is frequently deleted in
hematopoietic malignancies, including chronic lymphocytic leukemia
and multiple myeloma (18–20). Conversely, DLEU1 has also been
reported to be highly expressed in gastric cancer (21), breast cancer (22), epithelial ovarian carcinoma
(23) and endometrial carcinoma
(24), and is proposed to serve an
oncogenic role in tumor progression. At present, to the best of our
knowledge, no study has investigated the biological effects of
DLEU1 on the progression and development of RCC. Therefore, in the
present study, the role of DLEU1 in the tumorigenesis of RCC and
the underlying mechanisms were investigated. The present findings
suggested that knockdown of DLEU1 suppressed the growth, migration
and invasion of RCC cells, and induced cell apoptosis.
Additionally, the protein kinase B (Akt) pathway and
epithelial-mesenchymal transition (EMT) were inhibited by knockdown
of DLEU1. Collectively, this study indicated a pro-oncogenic role
for DLEU1 in the progression of RCC via modulation of the Akt
pathway and EMT.
Materials and methods
Cell culture and transfection
The human RCC cell lines KETR3 and 786-O were
obtained from the Cell Bank of Chinese Academy of Sciences and
cultured in DMEM (HyClone; GE Healthcare Life Sciences)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.)
and antibiotics (penicillin, 100 U/ml; streptomycin, 0.1 mg/ml;
Sigma-Aldrich; Merck KGaA) at 37°C with 5% CO2. In
total, ~6×104 cells were cultured overnight until 60–70%
convergence and cells were subsequently transfected with 50 nM
small interfering RNA (siRNA/si)-DLEU1 (Oligobio) or scrambled
siRNA [as negative control (NC); Oligobio] using Lipo6000 (Beyotime
Institute of Biotechnology) and cultured for 6–8 h at 37°C.
Subsequently, the medium containing the transfection reagent was
removed and the cells were cultured in DMEM for 24–48 h before the
next experiments. The sequence of siRNA-DLEU1 was
5′-CAACGGAAUGUAUCAAUGATT-3′, the sequence of siRNA-control was
5′-TTCTCCGAACGTGTCACGT-3′.
Reverse transcription-quantitative PCR
(RT-qPCR)
A total of 24 h post-transfection, total RNA was
isolated from transfected cells using an Ultrapure RNA kit (CoWin
Biosciences Co., Ltd.) and reverse transcribed into cDNA using a
HiFiScript cDNA Synthesis kit (CoWin Biosciences Co., Ltd.)
according to the manufacturer's protocol. qPCR was performed with a
SYBR Premix Ex Taq II kit (Takara Bio, Inc.). The thermocycling
conditions were as follows: Initial denaturation at 95°C for 30
sec, followed by 40 cycles of 95°C for 3 sec and 60°C for 30 sec.
Data were analyzed according to the sample quantification cycle
(Cq) value from three independent experiments. The relative gene
expression was calculated using the 2−∆∆Cq method
(25). Target gene expression was
normalized to the expression of β-actin. The primer sequences were
as follows: DLEU1, forward 5′-CCAGCCCACAGGCATTTAGT-3′, reverse,
5′-GTTCCGAGGCTTAAGTGCGA-3′; and β-actin, forward
5′-CCCGAGCCGTGTTTCCT-3′ and reverse,
5′-GTCCCAGTTGGTGACGATGC-3′.
Cell Counting kit-8 (CCK-8) assay
A CCK-8 (Beijing Solarbio Science & Technology,
Co., Ltd.) assay was conducted to assess cell viability. Briefly,
cells were seeded in each well of a 96-well plate at a density of
1×103 cells/well; after transfection for 0, 24, 48 and
72 h, cells were treated with CCK-8 reagent (10 µl) for an
additional 1 h at 37°C. The optical density value of excitation was
detected using a microplate reader (BioTek Instruments, Inc.) at a
wavelength of 450 nm. The assay was conducted three times
independently.
Colony forming assay
A total of 24 h post-transfection, cells were seeded
in a 6-cm dish at a density of 200 cells/well and were cultured for
~1 week at 37°C in an atmosphere containing 5% CO2 until
visible colonies were formed. The culture medium was removed, and 5
ml 4% paraformaldehyde was added to fix the colonies for 30 min at
room temperature followed by staining with 0.1% crystal violet for
30 min at room temperature. The colonies were counted under a light
microscope (Nikon Corporation, Inc.) and images were captured. The
assay was conducted three times independently.
Transwell assay
Transwell chambers with 8-µm pore filters (EMD
Millipore) coated with Matrigel (BD Biosciences) in a 24-well plate
were used for the cell invasion assay; chambers were not coated
with Matrigel for the migration assay. A total of 24 h
post-transfection, cells were trypsinized and resuspended in
serum-free culture medium at a concentration of 1×106
cells/ml. Subsequently, a 100- or 5-µl cell suspension was added to
the upper compartment of Transwell chambers for the invasion and
migration assays, respectively. Complete medium (600 µl) with 10%
FBS was added to the bottom chamber to serve as a chemoattractant.
After 48 h at 37°C, the residual cells on the upper surface were
removed, and the cells that had invaded or migrated to the lower
surface of the filter were fixed with 4% paraformaldehyde for 30
min at room temperature, followed by staining with 0.1% crystal
violet for 20 min at room temperature. Images of the invaded and
migrated cells were captured (magnification, ×40) and counted under
a light microscope. The assay was conducted three times
independently.
Apoptosis assay
Cells transfected with siRNA for 24 h were
resuspended in 1X binding buffer [10 mM HEPES/NaOH (pH 7.4), 140 mM
NaCl, 2.5 mM CaCl2] at a density of 1–5×106
cells/ml. Subsequently, the cell suspension (100 µl) was stained
with 5 µl Annexin V-FITC (BD Biosciences) in the dark for 5 min at
room temperature and incubated with 10 µl propidium iodide (PI) for
5 min at room temperature. The rate of apoptosis was analyzed by
flow cytometry (BD FACSCanto II; BD Biosciences) and calculated
using BD FACSDiva™ software (version 6.0; BD Biosciences). The
assay was conducted three times independently.
Western blot analysis
After 48 h of transfection, cells were harvested and
lysed using RIPA lysis buffer (CoWin Biosciences Co., Ltd.) at 4°C
for protein extraction. A bicinchoninic assay kit (Beyotime
Institute of Biotechnology) was used to measure the protein
concentration according to the manufacturer's protocol. Proteins
(20 µg/sample) were separated via 10% SDS-PAGE and were then
transferred onto PVDF membranes (EMD Millipore), which were blocked
with 5% non-fat milk for 1 h at room temperature, followed by
incubation overnight at 4°C with primary antibodies (1:1,000).
After washing with TBS-Tween-20 (20%), the membranes were incubated
with horseradish peroxidase-conjugated secondary antibodies
anti-rabbit immunoglobulin G (IgG; 1:3,000; cat. no. SA00001-2;
ProteinTech Group, Inc.) or anti-mouse IgG (1:3,000; cat. no.
SA00001-1; ProteinTech Group, Inc.) for 1 h at room temperature. An
enhanced chemiluminescence kit (CoWin Biosciences Co., Ltd.) was
used for signal development, and the bands were analyzed using
ImageJ software version 1.44 (National Institutes of Health). The
primary antibodies used were as follows: Anti- Bcl-2 (cat. no.
60178-1-Ig; ProteinTech Group, Inc.), Bax (cat. no. 50599-2-Ig;
ProteinTech Group, Inc.), total caspase-3 (cat. no. 66470-2-Ig;
ProteinTech Group, Inc.), cleaved caspase-3 (cat. no. 25546-1-AP;
ProteinTech Group, Inc.), total caspase-9 (cat. no.9502; Cell
Signaling Technology, Inc.), cleaved caspase-9 (cat. no.
10380-1-AP; ProteinTech Group, Inc.), Akt (cat. no. 9272; Cell
Signaling Technology, Inc.), phosphorylated (p)-Akt (cat. no.
66444-1-Ig; ProteinTech Group, Inc.), cyclin D1 (cat. no.
60186-1-Ig; ProteinTech Group, Inc.), P70S6K (cat. no. 14485-1-AP;
ProteinTech Group, Inc.) and GAPDH (cat. no. cat. no. 60004-1-Ig;
ProteinTech Group, Inc.). The assay was conducted three times
independently.
Statistical analysis
SPSS 18.0 software (SPSS, Inc.) was used for
statistical analysis. The data from triplicate experiments are
presented as the means ± standard deviation. The difference between
two groups was compared with Student's t-test. P<0.05 was
considered to indicate a statistically significant difference.
Results
DLEU1 knockdown inhibits the viability
and proliferation of RCC cells
To investigate the functional role of DLEU1 in the
growth of RCC cells, loss-of-function experiments were performed in
the RCC cell lines KETR3 and 786-O using si-DLEU1; scrambled siRNA
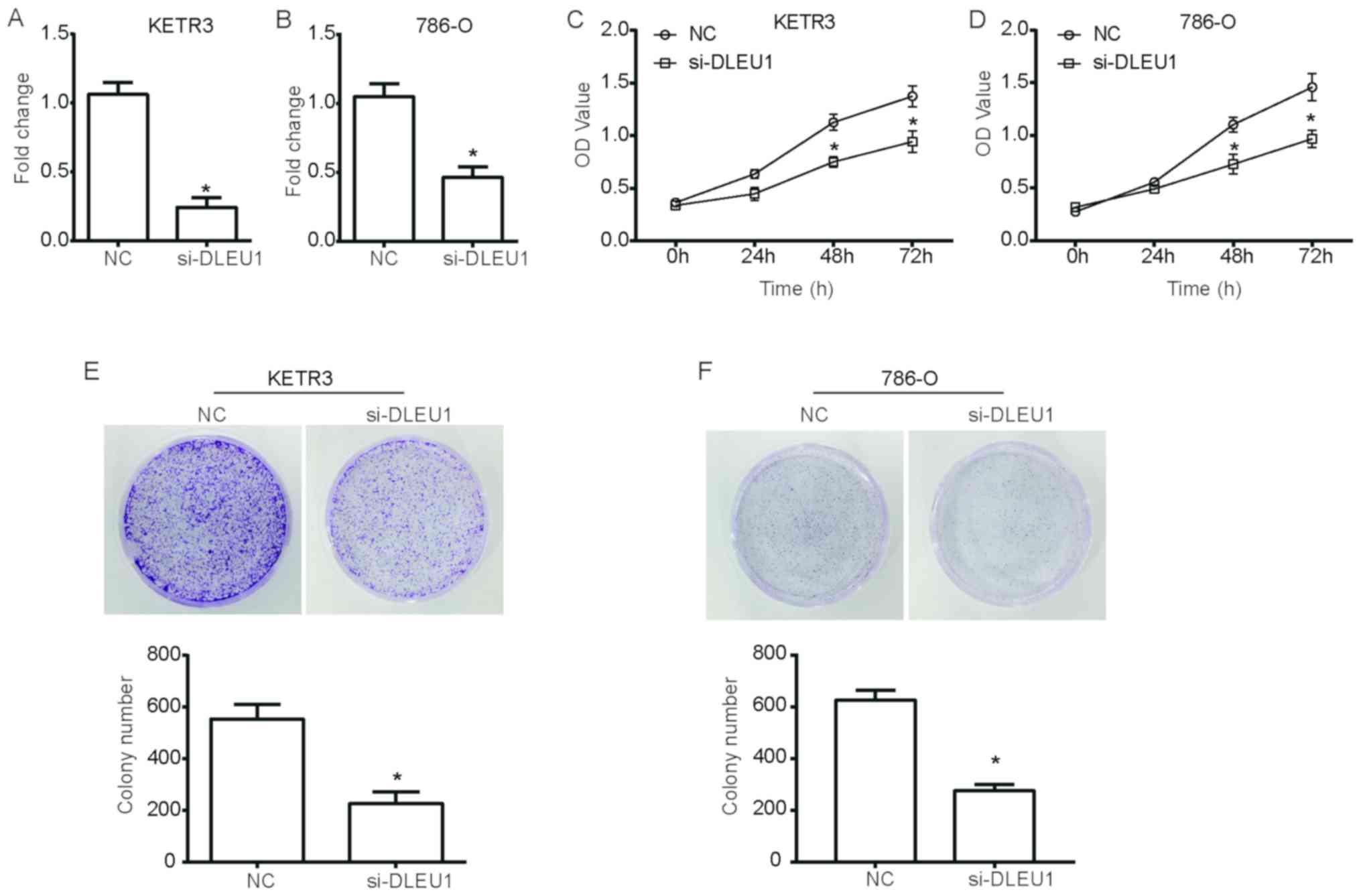
was used as the NC. As presented in Fig. 1A and B, the expression levels of
DLEU1 were significantly downregulated in si-DLEU1-transfected
cells compared with in NC cells (P<0.05). A CCK-8 assay was then
performed using cells transfected with si-DLEU1. Notably, the
growth curves of KETR3 cells demonstrated a significant reduction
in the viability of si-DLEU1-transfected cells compared with NC
cells (P<0.05; Fig. 1C).
Additionally, 786-O cells transfected with si-DLEU1 exhibited a
significant suppression of cell viability (P<0.05; Fig. 1D). Furthermore, a colony forming
assay further revealed that DLEU1 knockdown significantly decreased
the number of KETR3 and 786-O cell colonies compared with in the NC
group (P<0.05; Fig. 1E and F),
thus suggesting that DLEU1 knockdown inhibited the clonogenic
ability of RCC cells.
DLEU1 knockdown inhibits the migratory
and invasive abilities of RCC cells in vitro
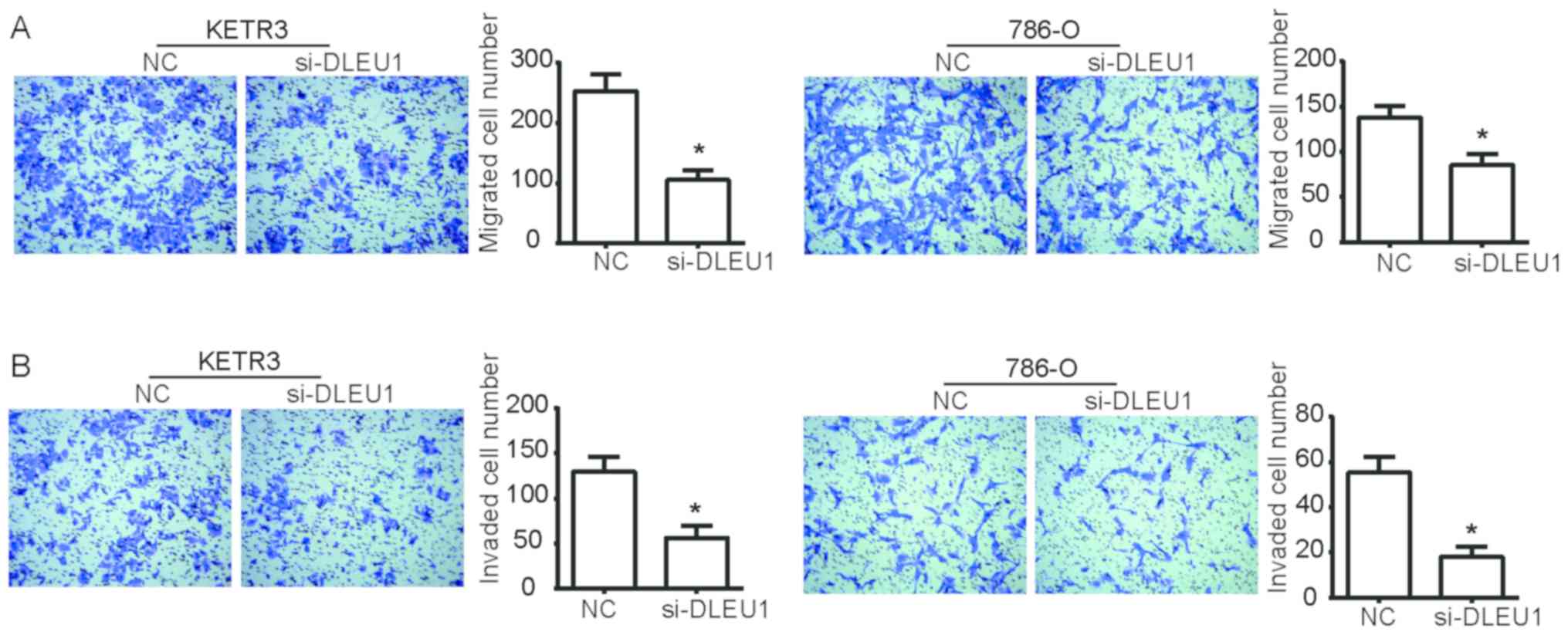
To determine the functional role of DLEU1 in the
progression of RCC, Transwell assays were performed to detect the
migration and invasion of KETR3 and 786-O cells. As presented in
Fig. 2A, si-DLEU1 transfection
significantly decreased the ability of KETR3 and 786-O cells to
migrate into the lower chamber compared with NC cells (P<0.05).
Additionally, compared with the NC, si-DLEU1-transfected cells
exhibited a significant reduction in their ability to invade
through the Matrigel into the lower chamber (P<0.05; Fig. 2B). Collectively, these data
indicated that DLEU1 knockdown reduced the migratory and invasive
abilities of RCC cells in vitro.
DLEU1 knockdown promotes apoptosis of
RCC cells in vitro
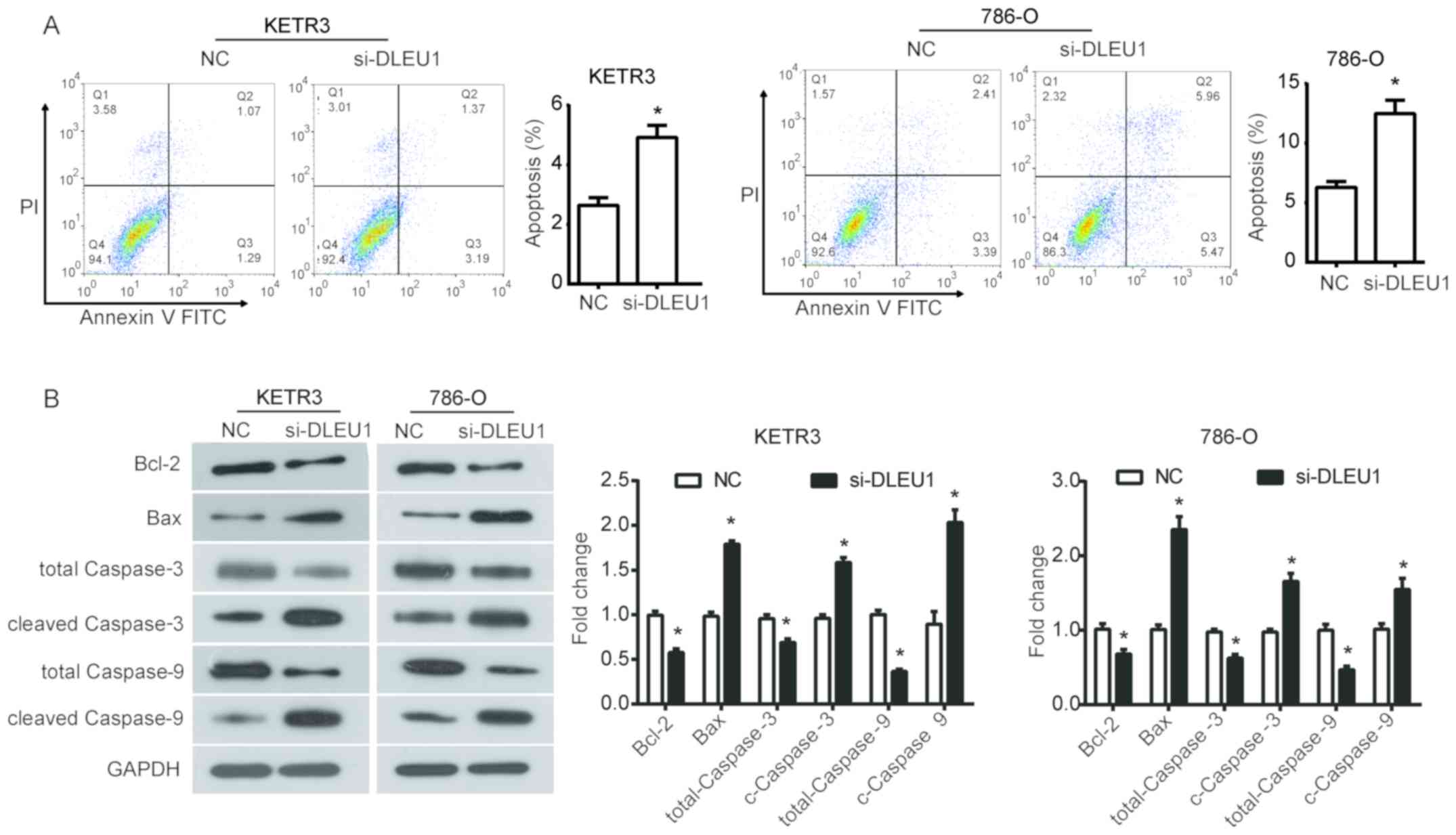
To further assess the effect of DLEU1 on the
survival of RCC cells, cell apoptosis was evaluated using flow
cytometry. It was demonstrated that DLEU1 knockdown significantly
increased the percentage of apoptotic KETR3 cells compared with in
the NC group (P<0.05; Fig. 3A).
Similarly, silencing DLEU1 with siRNA significantly promoted
apoptosis of 786-O cells (P<0.05; Fig. 3A). Subsequently, the expression of
apoptosis-associated proteins was evaluated to further investigate
the mechanisms underlying the increased apoptosis induced by the
downregulation of DLEU1 in RCC cells. From the results of western
blot analysis, it was revealed that DLEU1 downregulation
significantly inhibited the expression of Bcl-2, total caspase-3
and total caspase-9, and increased the expression of Bax, cleaved
caspase-3 and cleaved caspase-9 in KETR3 and 786-O cells compared
with in the NC group (P<0.05; Fig.
3B). Collectively, these findings suggested that the increase
in apoptosis induced by DLEU1 knockdown may be dependent on its
regulation of the Bcl-2/Bax axis and caspase cascade in RCC
cells.
DLEU1 knockdown suppresses the Akt
pathway and disrupts EMT in RCC cells
The phosphatidylinositol-3-kinase (PI3K)/Akt
signaling pathway is an established pathway involved in the
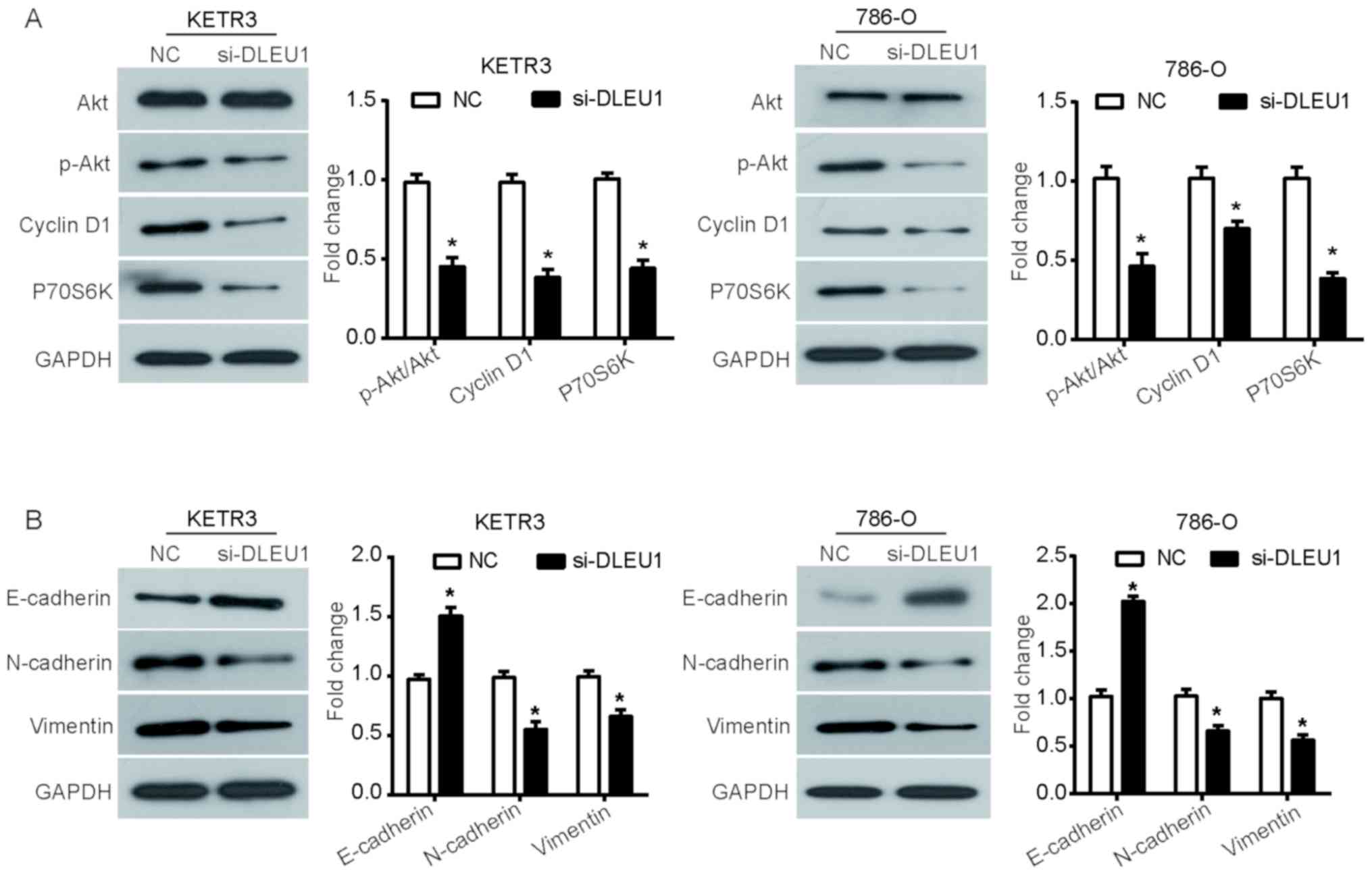
occurrence and development of cancer (26). To determine whether the Akt
signaling pathway was involved in the function of DLEU1, the
phosphorylation levels of Akt, and the expression of downstream
proteins cyclin D1 and P70S6 kinase (P70S6K) were evaluated via
western blot analysis. It was observed that silencing DLEU1
significantly reduced the phosphorylation levels of Akt in KETR3
and 786-O cells compared with the NC (P<0.05), but induced no
notable effects on the expression of total Akt (Fig. 4A). Additionally, the expression
levels of cyclin D1 and P70S6K, proteins involved in the cell
cycle, were similarly downregulated in DLEU1 knockdown cells
(P<0.05; Fig. 4A).
Due to the critical role of EMT in promoting tumor
cell migration and invasion, the expression of EMT marker proteins
was further assessed to investigate whether EMT was involved in the
function of DLEU1. It was revealed that si-DLEU1-transfected KETR3
cells demonstrated a significant increase in the expression of
E-cadherin, an important marker of epithelial cells, with similar
observations in 786-O cells (P<0.05; Fig. 4B). Furthermore, DLEU1 knockdown
also induced a significant downregulation in the expression of
N-cadherin and vimentin, which are important mesenchymal markers,
in KETR3 and 786-O cells (P<0.05; Fig. 4B). Collectively, these results
suggested that DLEU1 knockdown interfered with the mesenchymal
properties of RCC cells.
Discussion
Increasing evidence has indicated that lncRNAs serve
important roles in modulating a wide variety of fundamental
biological processes by regulating gene expression, despite the
fact that lncRNAs do not encode proteins (7–9). In
view of their important regulatory functions, lncRNAs have been
reported to be closely associated with tumorigenesis and metastasis
(5). For example, the lncRNA
metastatic renal cell carcinoma-associated transcript 1 has been
reported to function as a prognostic biomarker and therapeutic
target for clear cell RCC (ccRCC), as it promotes the metastasis of
ccRCC (6). Therefore, further
understanding of the association between lncRNAs and tumor
progression may provide novel targets and therapeutic perspectives
for tumor-targeted therapy (5,27).
It has been demonstrated that there is recurrent deletion of DLEU1
in hematopoietic tumors, and that DLEU1 acts as a tumor suppressor
gene via inhibition of the cell cycle and promotion of programmed
cell death in Burkitt lymphoma (20). Recent studies have confirmed that
DLEU1 is markedly upregulated in certain solid tumor tissues and
cell lines, and serves as an oncogene to promote tumorigenesis in
gastric cancer, ovarian carcinoma and endometrial carcinoma
(21,23,24).
Therefore, the present study aimed to investigate the function of
DLEU1 in the progression of RCC. The present study, to the best of
our knowledge, is the first to demonstrate that silencing DLEU1 in
two RCC cell lines (KETR3 and 786-O cells) inhibited the viability,
migration and invasion of RCC cells. This promoted the
mitochondrial pathway of apoptosis by regulating the Bcl-2/Bax axis
and caspase cascade, suggesting a pro-oncogenic role of DLEU1 in
the progression and development of RCC in vitro. This is
consistent with other findings from ovarian cancer (23).
To provide insight into the relevant mechanisms by
which dysregulated DLEU1 contributes to cell proliferation,
migration, invasion and apoptosis in RCC cells, potentially
associated signaling pathways were further investigated. It is
widely reported that the PI3K/Akt signaling pathway serves a
pivotal role in the regulation of cellular processes, and tumor
progression and development, including cell survival, apoptosis,
cell cycle regulation and invasion (28). Additionally, the PI3K/Akt signaling
pathway is frequently activated in cancer, and targeted inhibition
of this pathway has become the focus for the treatment of various
types of cancer, including RCC (29,30).
Therefore, following silencing of DLEU1, alterations in the
PI3K/Akt signaling pathway and the expression of important
downstream proteins were determined, including cyclin D1 and
P70S6K, which are key regulators in mediating the cell cycle and
proliferation (31–33). In the present study, western
blotting revealed a significant decrease in the expression of
p-Akt, cyclin D1 and P70S6K following DLEU1 knockdown, which is
consistent with results from endometrial cancer cells that DLEU1
knockdown reduces the expression of Akt1, mTOR and P70S6K (24). Based on these results, the Akt
pathway may be involved in the mechanisms underlying the
pro-oncogenic role of DLEU1 in RCC cells.
EMT is a crucial process that promotes cell invasion
and enhances the metastatic potential of tumor cells (34). It has been demonstrated that
decreased expression of E-cadherin, and increased expression of
N-cadherin and vimentin in epithelial cells indicates that the
tumors are susceptible to a metastatic phenotype (35,36).
Therefore, in this study, it was further investigated as to whether
DLEU1 affected the EMT process in RCC cells. It was revealed that
knockdown of DLEU1 resulted in increased expression of E-cadherin,
a classical epithelial cell marker, but reduced the expression of
N-cadherin and vimentin, suggesting that DLEU1 may modulate EMT in
RCC cells in vitro.
In conclusion, to the best of our knowledge, the
present study is the first to report that DLEU1 may serve as an
oncogene in the progression and development of RCC via modulation
of the Akt pathway and EMT phenotype in RCC cells, thus indicating
that DLEU1 may serve as a potential therapeutic target in RCC.
Notably, lncRNAs usually function by binding to miRNAs or genes;
therefore, the target genes of DLEU1 that are involved in the
growth and survival of RCC cells will be investigated in the
future, as will the role of DLEU1 in vivo.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Inner
Mongolia Natural Science Foundation Project (grant. no.
2015MS08115), the Inner Mongolia Autonomous Region Health and
Family Planning Research Project (grant. no. 201701066) and the
Major Project of Affiliated Hospital of Inner Mongolia Medical
University (grant. no. NYFY ZD 2014011).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
GY, CC, YX and HC conceived and designed the
experiments. GY, CC, LB, GW, YH, YW, HC and YX performed the
experiments. GY, YX and HC contributed to the conception of the
study and wrote the manuscript. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
Akt
|
protein kinase B
|
|
lncRNA
|
long noncoding RNA
|
|
RCC
|
renal cell carcinoma
|
References
|
1
|
Hsieh JJ, Purdue MP, Signoretti S, Swanton
C, Albiges L, Schmidinger M, Heng DY, Larkin J and Ficarra V: Renal
cell carcinoma. Nat Rev Dis Primers. 3:170092017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Thomas JS and Kabbinavar F: Metastatic
clear cell renal cell carcinoma: A review of current therapies and
novel immunotherapies. Crit Rev Oncol Hematol. 96:527–533. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Motzer RJ and Molina AM: Targeting renal
cell carcinoma. J Clin Oncol. 27:3274–3276. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xia M, Yao L, Zhang Q, Wang F, Mei H, Guo
X and Huang W: Long noncoding RNA HOTAIR promotes metastasis of
renal cell carcinoma by up-regulating histone H3K27 demethylase
JMJD3. Oncotarget. 8:19795–19802. 2017.PubMed/NCBI
|
|
6
|
Li JK, Chen C, Liu JY, Shi JZ, Liu SP, Liu
B, Wu DS, Fang ZY, Bao Y, Jiang MM, et al: Long noncoding RNA
MRCCAT1 promotes metastasis of clear cell renal cell carcinoma via
inhibiting NPR3 and activating p38-MAPK signaling. Mol Cancer.
16:1112017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Esteller M: Non-coding RNAs in human
disease. Nat Rev Genet. 12:861–874. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shi SJ, Wang LJ, Yu B, Li YH, Jin Y and
Bai XZ: LncRNA-ATB promotes trastuzumab resistance and
invasion-metastasis cascade in breast cancer. Oncotarget.
6:11652–11663. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ong MS, Cai W, Yuan Y, Leong HC, Tan TZ,
Mohammad A, You ML, Arfuso F, Goh BC, Warrier S, et al: ‘Lnc’-ing
Wnt in female reproductive cancers: Therapeutic potential of long
non-coding RNAs in Wnt signalling. Br J Pharmacol. 174:4684–4700.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hosseini ES, Meryet-Figuiere M,
Sabzalipoor H, Kashani HH, Nikzad H and Asemi Z: Dysregulated
expression of long noncoding RNAs in gynecologic cancers. Mol
Cancer. 16:1072017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang F, Li L, Xu H, Liu Y, Yang C, Cowley
AW Jr, Wang N, Liu P and Liang M: Characteristics of long
non-coding RNAs in the Brown Norway rat and alterations in the Dahl
salt-sensitive rat. Sci Rep. 4:71462014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Iguchi T, Uchi R, Nambara S, Saito T,
Komatsu H, Hirata H, Ueda M, Sakimura S, Takano Y, Kurashige J, et
al: A long noncoding RNA, lncRNA-ATB, is involved in the
progression and prognosis of colorectal cancer. Anticancer Res.
35:1385–1388. 2015.PubMed/NCBI
|
|
14
|
Xiong J, Liu Y, Jiang L, Zeng Y and Tang
W: High expression of long non-coding RNA lncRNA-ATB is correlated
with metastases and promotes cell migration and invasion in renal
cell carcinoma. Jpn J Clin Oncol. 46:378–384. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li J, Li Z, Zheng W, Li X, Wang Z, Cui Y
and Jiang X: LncRNA-ATB: An indispensable cancer-related long
noncoding RNA. Cell Prolif. 50:2017. View Article : Google Scholar :
|
|
16
|
Lei K, Liang X, Gao Y, Xu B, Xu Y, Li Y,
Tao Y, Shi W and Liu J: Lnc-ATB contributes to gastric cancer
growth through a MiR-141-3p/TGFβ2 feedback loop. Biochem Biophys
Res Commun. 484:514–521. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zeng X, Hu Z, Ke X, Tang H, Wu B, Wei X
and Liu Z: Long noncoding RNA DLX6-AS1 promotes renal cell
carcinoma progression via miR-26a/PTEN axis. Cell Cycle.
16:2212–2219. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Garding A, Bhattacharya N, Claus R, Ruppel
M, Tschuch C, Filarsky K, Idler I, Zucknick M, Caudron-Herger M,
Oakes C, et al: Epigenetic upregulation of lncRNAs at 13q14.3 in
leukemia is linked to the In Cis downregulation of a gene cluster
that targets NF-κB. PLoS Genet. 9:e10033732013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dowd AA, Homeida S and Elkarem HA:
Detection of chromosome 13 (13q14) deletion among Sudanese patients
with multiple myeloma using a molecular genetics fluorescent in
situ hybridization technique (FISH). Malays J Pathol. 37:95–100.
2015.PubMed/NCBI
|
|
20
|
Lee S, Luo W, Shah T, Yin C, O'Connell T,
Chung TH, Perkins SL, Miles RR, Ayello J, Morris E, et al: The
effects of DLEU1 gene expression in Burkitt lymphoma (BL):
Potential mechanism of chemoimmunotherapy resistance in BL.
Oncotarget. 8:27839–27853. 2017.PubMed/NCBI
|
|
21
|
Li X, Li Z, Liu Z, Xiao J, Yu S and Song
Y: Long non-coding RNA DLEU1 predicts poor prognosis of gastric
cancer and contributes to cell proliferation by epigenetically
suppressing KLF2. Cancer Gene Therapy. 25:58–67. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu Q, Guo L, Jiang F, Li L, Li Z and Chen
F: Analysis of the miRNA-mRNA-lncRNA networks in ER+ and ER- breast
cancer cell lines. J Cell Mol Med. 19:2874–2887. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang LL, Sun KX, Wu DD, Xiu YL, Chen X,
Chen S, Zong ZH, Sang XB, Liu Y and Zhao Y: DLEU1 contributes to
ovarian carcinoma tumourigenesis and development by interacting
with miR-490-3p and altering CDK1 expression. J Cell Mol Med.
21:3055–3065. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Du Y, Wang L, Chen S, Liu Y and Zhao Y:
lncRNA DLEU1 contributes to tumorigenesis and development of
endometrial carcinoma by targeting mTOR. Mol Carcinog.
57:1191–1200. 2018. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen JS, Wang Q, Fu XH, Huang XH, Chen XL,
Cao LQ, Chen LZ, Tan HX, Li W, Bi J and Zhang LJ: Involvement of
PI3K/PTEN/AKT/mTOR pathway in invasion and metastasis in
hepatocellular carcinoma: Association with MMP-9. Hepatol Res.
39:177–186. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ellinger J, Blondeau JJC, Deng M, Syring
I, Schrödter S, Schmidt D, Perner S and Müller SC: 863
Identification and functional analysis of novel long non-coding
RNAs in clear cell renal cell carcinoma. Eur Urol Suppl.
14:e8632015. View Article : Google Scholar
|
|
28
|
Kwong LN and Davies MA: Navigating the
therapeutic complexity of PI3K pathway inhibition in melanoma. Clin
Cancer Re. 19:5310–5319. 2013. View Article : Google Scholar
|
|
29
|
Guo H, German P, Bai S, Barnes S, Guo W,
Qi X, Lou H, Liang J, Jonasch E, Mills GB and Ding Z: The PI3K/AKT
pathway and renal cell carcinoma. J Genet Genomics. 42:343–353.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sathe A and Nawroth R: Targeting the
PI3K/AKT/mTOR pathway in bladder cancer. Methods Mol Biol.
1655:335–350. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Diehl JA: Cycling to Cancer with Cyclin
D1. Cancer Biol Ther. 1:226–231. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Qie S and Diehl JA: Cyclin D1, cancer
progression, and opportunities in cancer treatment. J Mol Med
(Berl). 94:1313–1326. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tavares MR, Pavan IC, Amaral CL,
Meneguello L, Luchessi AD and Simabuco FM: The S6K protein family
in health and disease. Life Sci. 131:1–10. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bates RC and Mercurio A: The
epithelial-mesenchymal tansition (EMT) and colorectal cancer
progression. Cancer Biol Ther. 4:365–370. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nakajima S, Doi R, Toyoda E, Tsuji S, Wada
M, Koizumi M, Tulachan SS, Ito D, Kami K, Mori T, et al: N-cadherin
expression and epithelial-mesenchymal transition in pancreatic
carcinoma. Clin Cancer Res. 10:4125–4133. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mao XW, Xiao JQ, Xu G, Li ZY, Wu HF, Li Y,
Zheng YC and Zhang N: CUL4B promotes bladder cancer metastasis and
induces epithelial-to-mesenchymal transition by activating the
Wnt/β-catenin signaling pathway. Oncotarget. 8:77241–77253. 2017.
View Article : Google Scholar : PubMed/NCBI
|