Introduction
Cerebrovascular disease is a common disease with
high morbidity and mortality worldwide (1). Ischemic stroke as a type of
cerebrovascular disease can lead to cognitive impairment and
neurological deficit (2). At
present, thrombolysis, which effectively reduces infarct size, is
the main treatment for ischemic stroke (3). However, cerebral ischemia/reperfusion
(I/R) injury often occurs irreversibly in ischemic brain tissue
after restoration of blood perfusion, which severely restricts the
recovery of patients with ischemic stroke (4). Although some promising
neuroprotective agents have been identified, such as curcumin,
rosmarinic acid, astragaloside VI and berberine (5,6),
their protective effects are still limited. Therefore,
identification of novel neuroprotective agents for cerebral I/R
injury is highly desired.
In recent years, abundant evidence shows that
oxidative stress plays an important role in the pathogenesis of
cerebral I/R injury (7). Wu et
al found that reactive oxygen species (ROS) and malondialdehyde
(MDA) were significantly increased in rats with middle cerebral
artery occlusion (MCAO), whereas the activity of antioxidants was
markedly reduced (7). The impaired
antioxidant defense system was found to lead to oxidative damage of
brain lipids, proteins and DNA, further leading to cell death and
brain dysfunction (8,9). Nuclear factor (erythroid-derived
2)-related factor 2 (Nrf2) is a key transcription factor that
regulates the antioxidant stress response (10). Many studies have shown that
activation of the Nrf2 pathway can protect against cerebral I/R
injury in vivo and in vitro (11,12).
Neuroprotective agents, such as resveratrol, isoquercetin and
protocatechualdehyde also reportedly protect the brain from
cerebral I/R injury through Nrf2 activation (9,11,13).
Therefore, repair of the endogenous antioxidant system through
activation of the Nrf2 pathway may be an effective strategy to
alleviate cerebral I/R injury.
Many studies have demonstrated that plant extracts
are potential neuroprotective agents in the treatment of ischemic
stroke (14). Theaflavin, which is
derived from black tea, has been widely used as an antioxidant.
Feng et al reported that theaflavin prevents DNA damage by
suppressing oxidative stress and inhibiting cytochrome P450 1A1
in vitro (15). In
addition, theaflavin demonstrated neuroprotective effects by
inhibiting the activity of oxidant enzymes in PC12 cells (16). These findings provide the
foundation for a new field of exploration using theaflavins in the
treatment of cerebrovascular disease caused by oxidative stress.
Cai et al reported that theaflavin ameliorates cerebral I/R
injury in rats through anti-inflammatory effects and modulation of
signal transducer and activator of transcription (STAT)-1 (17). However, the exact molecular
mechanism by which theaflavin attenuates cerebral I/R injury
through antioxidative stress still awaits further
investigation.
The non-regeneration and vulnerability of nerves
suggest that they are extremely fragile and difficult to repair
once damaged. Endogenous neural stem cells (NSCs) with
proliferative ability exist in at least two regions of the central
nervous system of adult mammals, the hippocampus and lateral
ventricle (18,19). NSCs can self-renew, generate
neuroblasts and migrate to damaged areas of the brain to replace
dead nerve cells to induce neurological activity (20,21).
Because of the relatively small number of NSCs, their proliferation
is essential for the recovery of neural function and repair of
cognitive impairment in cerebral I/R injury. Therefore, exploration
of the effects of neuroprotective agents on NSC proliferation has
considerable clinical significance.
Herein, an in vivo experimental animal model
of stroke and in vitro model of NSCs that were subjected to
oxygen-glucose deprivation and reoxygenation (OGD/R) were used to
demonstrate that theaflavin is able to promote neurogenesis and
repair neurological function by inhibiting oxidative stress to
induce brain recovery.
Materials and methods
Cerebral I/R model (7)
In total, 50 Male Sprague-Dawley (SD) rats, 7–8
weeks old (body weight 270–300 g) were obtained from East China
Normal University [Production License: SCXK (Shanghai) 2016-0004].
All animal protocols were approved and regulated by the Animal Care
and Ethics Committee of East China Normal University. All rats were
allowed free access to water and food and maintained at a
controlled temperature range (20±2°C). The intraluminal suture
middle cerebral artery occlusion (MCAO) method was performed to
induce focal cerebral ischemia. Rats were anesthetized with
isoflurane (4%) in N2O gas (70%) with O2
(30%) in all experiments. A midline cervical incision was used to
expose the left-sided carotid arteries. The left external carotid
artery was dissected and isolated distally by coagulating its
branches, on which a distal ligature was placed. A piece of 3-0 tip
rounded monofilament nylon suture was introduced via the lumen of
the left external/internal carotid artery stump to be embedded into
the left anterior cerebral artery. At 1.5 h of cerebral ischemia,
the monofilament nylon suture was withdrawn to permit reperfusion.
During the surgical process, moorVMS-LDF1-HP high power laser
doppler detector (Jiander Technology Co., Ltd., Beijing, China) was
used to monitor blood flow changes. An abrupt reduction in regional
cerebral perfusion to less than 30% of the baseline value was
considered to achieve focal ischemia. For the sham group, the
surgical process was the same as that for the MCAO group but
without suture occlusion.
Primary cultured NSCs
Primary cultured NSCs were harvested from the
hippocampus in embryonic SD rats aged 14–16 days as previously
described (21). In brief, the
fore cortex in rat embryos was isolated and ground into single
cells using nylon mesh (70 µm). Cells were then re-suspended in
Dulbecco's modified Eagle's medium (DMEM)/Nutrient Mixture F-12
(DMEM/F12) (Invitrogen, Thermo Fisher Scientific, Inc., Waltham,
MA, USA) and were plated in 10-cm dishes for culture at
1×104 cells/cm2 in DMEM/F12 medium with fetal
bovine serum (FBS, 10%; Gibco, Thermo Fisher Scientific, Inc.),
epidermal growth factor (EGF, 10 ng/ml; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany), B27 (2%; Invitrogen; Thermo Fisher Scientific,
Inc.), basic fibroblast growth factor (bFGF, 10 ng/ml;
Sigma-Aldrich; Merck KGaA), penicillin (5 IU; Sigma-Aldrich; Merck
KGaA), and streptomycin (5 µg/ml; Sigma-Aldrich; Merck KGaA). Cells
were incubated in a humidified atmosphere containing 21%
O2 and 78% N2 supplied with 5% CO2
at 37°C for 5 days to form a neurosphere (diameter ~100 µm). The
neurospheres were collected and triturated to a single-NSC
suspension. The NSCs were then replanted at 1×104
cells/cm2 and passaged every 4 to 6 days. These NSCs
still possessed self-renewal, proliferation and differentiation
capacity until the 10th passage. The NSCs of the 2nd to 5th
passages were used for subsequent experiments. To establish the
OGD/R cellular model, NSCs were cultured in glucose-free balanced
salt solution under hypoxic conditions (1% O2, 5%
CO2, and 94% N2) at 37°C for 3 h.
Subsequently, NSCs were cultured in normal medium and normoxic
conditions (21% O2, 5% CO2 and 78%
N2).
Theaflavin treatments
Theaflavin [98.0%, high-performance liquid
chromatography (HPLC) grade] was purchased from Shyuanye
Biotechnology (Shanghai, China). In the in vivo animal
study, theaflavin dissolved in saline was intravenously injected
into SD rats (10 mg/kg group and 50 mg/kg group) 2 h after MCAO
ischemia, once a day for 7 days. For the 10 mg/kg group, SD rats
received theaflavin at 10 mg/kg; for the 50 mg/kg group, SD rats
received theaflavin at 50 mg/kg. The same volume of saline was
injected into SD rats (MCAO group). For in vitro
experiments, theaflavin dissolved in phosphate-buffered saline
(PBS) was added to the DMEM/F12 medium. The NSCs were seeded into
6/96-well plates in the DMEM/F12 media with theaflavin. After
incubation for 2 h, NSCs were challenged with OGD/R. The NSCs were
divided into the following: Control group, OGD/R group (same volume
of PBS), 2 µM theaflavin group, and 10 µM theaflavin group. After
incubation for 48 h, a series of following experiments were
conducted.
Tissue preparation and staining
The SD rats were sacrificed at 7 days after
theaflavin treatment. The harvested brain tissues were cut into
five equally-sized coronal sections (2–3 mm thickness) using an EM
UC7 microtome (Leica Microsystems Inc., Wetzlar, Germany). The
brain sections were then incubated with 2%
2,3,5-triphenyltetrazolium chloride (TTC; Sigma-Aldrich; Merck
KGaA) at 37°C for 15 min with gentle shaking. Subsequently, the
brain sections were fixed with paraformaldehyde (4%) overnight. The
stained slices were photographed and the infarct size was
quantified using the Image-Pro Plus 6.0 software (Media
Cybernetics, Inc., Rockville, MD, USA). For terminal
deoxynucleotidyl transferase dUTP nick end labeling (TUNEL)
staining (Beyotime Institute of Biotechnology, Haimen, China), the
harvested brain tissues were cut into 2.5 mm blocks and fixed with
paraformaldehyde overnight. Subsequently, blocks were embedded in
paraffin and cut into 4-µm coronal sections, according to the
manufacturer's protocol. Apoptotic images were obtained using the
Axioplan 2 imaging system (Carl Zeiss AG, Oberkochen, Germany). Six
sections of each sample were selected and five fields of each
section were examined for quantification of apoptosis.
Evaluation of neurological deficits,
learning and memory ability
Neurological deficit testing in each group was
conducted by an investigator who was blinded to the experimental
groups according to the method of Longa et al (22). Neurological findings were scored on
a five-point scale: Depressed levels of consciousness and inability
to walk spontaneously=4; falling to the right=3; circling to the
right=2; failure to extend right forepaw fully=1; and no
neurological deficits=0. The Morris water maze test (9,11)
was conducted to evaluate memory and learning function by an
investigator blinded to the experimental groups. To make the water
opaque, non-toxic ink was added. Rats were brought into the
conducting room 30 min prior to the test. They were subjected to
four trials per day for 5 consecutive days. Rats were given 1 min
to find the platform in each trial. The escape latency was recorded
when experimental rats reached the platform. It was recorded as 1
min even when the rat could not reach the platform within 1 min.
Rats were allowed to stay on the platform for another 15 sec after
locating it. After the test, rats were dried and then returned to
their cages. A 4-min inter-trial rest was allowed. The probe trial
was performed on the sixth day. The round tank was divided into
four equal quadrants. The target circle was considered as an area
20 cm in diameter, as measured from the platform center. The
platform crossing frequency was defined as the number of times the
rat crossed the area within 1.5 min.
Neurosphere assay (6)
Primary cultured NSCs that were subjected to both
theaflavin treatment and OGD/R were fixed with paraformaldehyde
(4%) for 20 min. After being rinsed with PBS, NSCs were incubated
with blocking buffer (PBS, 0.1% Triton-100, and 100% goat serum)
for 2 h. The primary antibody against Nestin (mouse, dilution
1:300; Abcam, Cambridge, UK; cat. no. ab6142) was added at 4°C
overnight. After being washed with PBS, the secondary antibody
(Alexa 568 goat anti-mouse, dilution 1:300; Huabio; cat. no.
HA1017) was added at 25°C in the dark for 2 h. Subsequently, NSCs
were incubated with 4′,6-diamidino-2-phenylindole (DAPI) for 10 min
to identify the neurosphere, after which they were washed with PBS.
The neurosphere images were obtained using a Zeiss Axioskop 50
Trinocular Fluorescence Microscope (Carl Zeiss; magnification,
×200). Fifteen fields were randomly chosen in each well. The
diameter of the neurosphere was quantified using the Image-Pro Plus
6.0 software (Media Cybernetic).
Cell Counting Kit-8 (CCK-8) assay and
flow cytometry
Cell viability of NSCs was evaluated using the CCK-8
assay (Sigma-Aldrich; Merck KGaA). Primary cultured NSCs
(1×104/well) were seeded into 96-well plates and
cultured for 24 h. The NSCs were then subjected to theaflavin
treatment and OGD/R. Subsequently, the NSCs were incubated with 10%
water-soluble tetrazolium dye (WST-8). The optical density (OD) was
then measured using a LUX microplate reader (Thermo Fisher
Scientific) at an absorbance of 450 nm. The apoptotic rate in NSCs
was detected using the Annexin V-FITC Apoptosis kit (Beyotime)
according to the manufacturer's instructions. Primary cultured NSCs
were seeded into 6-well plates and cultured for 24 h. The NSCs were
then subjected to theaflavin treatment and OGD/R. Subsequently, the
NSCs were incubated with Annexin V and then added to propidium
iodide (PI) buffer for 15 min at 25°C in a dark room. The apoptotic
cells were quantified by a FACSAria II flow cytometer (BD
Biosciences, Franklin Lakes, NJ, USA).
Detection of oxidative stress
The supernatant containing the lysates of rat brain
tissues and NSCs was homogenized in ice-cold mitochondria isolation
buffer. The total ROS level in the brain tissue was detected using
the Total ROS Activity Assay kit (Sigma-Aldrich; Merck KGaA). The
supernatant lysate was incubated with Total ROS Green in an
incubator (5% CO2) for 6 h at 37°C. Fluorescence
intensity was then detected at 525 nm emission and 490 nm
excitation. The levels of malondialdehyde (MDA), glutathione
peroxidase (GSH-Px), and superoxide dismutase (SOD) in the rat
brain tissues and NSCs were measured using standard assay kits,
including the Total SOD Activity Detection kit (Solarbio, Beijing,
China), Malondialdehyde (MDA) Content Detection kit (Solarbio), and
Glutathione Peroxidase (GSH-Px) Detection kit (R&D Systems,
Inc., Minneapolis, MN, USA), according to the manufacturer's
instructions. The OD value was then measured using a LUX microplate
reader (Thermo Fisher Scientific).
RNA extraction and quantitative
real-time PCR (qRT-PCR)
According to the manufacturer's instructions, total
RNA was extracted and complementary DNA (cDNA) was synthesized
using the Animal Total RNA Isolation kit (Sangon Biotech Co., Ltd.,
Shanghai, China) and M-MuLV First Strand cDNA Synthesis kit (Sangon
Biotech Co., Ltd.). qRT-PCR was performed using SYBR Real-Time PCR
kit (Genepharma, Shanghai, China) under the following conditions:
95°C for 600 sec; followed by 45 cycles at 95°C for 10 sec; 60°C
for 20 sec; and 72°C for 30 sec. The relative fold expression was
calculated using the 2−ΔΔCq method (23). Each analysis was performed in
triplicate. In addition, U6 served as an internal reference
gene for miRNAs, and β-actin, for mRNA. The sequences were as
follows: miRNA-128-3p forward, 5′-GGTCACAGTGAACCGGTC-3′ and
reverse, 5′-GTGCAGGGTCCGAGGT-3′; U6 forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′;
Nrf2 forward, 5′-AGCACACCCAGTCAGAAACCAG-3′ and reverse,
5′-TCTACAAACGGGAATGTCG-3′; β-actin forward,
5′-GGTCACAGTGAACCGGTC-3′ and reverse, 5′-GTGCAGGGTCCGAGGT-3′.
Cell transfection
For cell transfection, NSCs were seeded at a density
of 5.5×105/well in a 6-well plate 12 h before
transfection. For miRNA-128-3p downregulation, NSCs were
transfected with 100 nM negative control miRNA
5′-CAGUACUUUUGUGUAGUACAAA-3′ (NC miRNA) or 100 nM miRNA-inhibitor
5′-AAAGAGACCGGUUCACUGUGA-3′ (GenePharma, Shanghai, China) using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific),
according to the manufacturer's instructions, and incubated for an
additional 48 h. For miRNA-128-3p upregulation, NSCs were
transfected with 100 nM negative control miRNA (NC miRNA) sense:
5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense:
5′-ACGUGACACGUUCGGAGAATT-3′ or 100 nM miRNA-mimics, sense:
5′-UCACAGUGAACCGGUCUCUUUAGU-3′ and antisense:
5′-UAAAGAGACCGGUUCACUGUGAUU-3′ (GenePharma, Shanghai, China) using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific),
according to the manufacturer's instructions, and incubated for an
additional 48 h. After transfection, the NSCs were harvested for
further analysis. Lentivirus-mediated Nrf2 plasmids (OE-Nrf2) were
constructed by GenePharma Co. For Nrf2 overexpression, cells were
transfected with lentivirus-mediated Nrf2 plasmids. Cells
transfected with an empty vector were also employed and
investigated. The transfected cells were subsequently harvested for
further analysis.
Dual-luciferase reporter assay
The 3′-untranslated region (3′UTR) of Nrf2 was
amplified by Sangon, and the sequences were inserted into a
pMIR-reporter luciferase vector (Ambion, Rockville, MD, USA). The
mutated 3′UTR was designed, constructed, and then inserted into the
luciferase vector. The mutated-type (MUT) and wild-type (WT) 3′UTR
vectors, NC miRNA, and miRNA mimics were co-transfected into 293
cells, alternately. The Dual-Luciferase Reporter Assay
(Activemotif, USA) was performed to measure luciferase intensity 2
days after co-transfection.
Western blot analysis
Rat brain tissues and NSCs were harvested using a
lysis buffer solution [radioimmunoprecipitation assay (RIPA)
buffer, Beyotime Biotechnology], protease inhibitor cocktail
(Thermo Fisher Scientific), and phosphatase inhibitor cocktail
(Beyotime Biotechnology). After protein quantification using the
bicinchoninic acid (BCA) protein concentration assay kit (Thermo
Fisher Scientific), Total protein (50 µg) was separated by 10%
sodium dodecyl sulfate polyacrylamide gel electrophoresis
(SDS-PAGE). After being transferred onto a polyvinylidene
difluoride (PVDF) membrane, the protein was probed with primary
antibodies for Nrf2 (rat, dilution 1:300; Abcam; cat. no. ab137550)
and glyceraldehyde 3-phosphate dehydrogenase (GAPDH; rat, dilution
1:500; Huabio; cat. no. ET1601-4). After being washed, the
membranes were incubated with secondary antibody goat anti-mouse
IgG (dilution, 1:1,000; Millipore; cat. no. 401211). Band intensity
was quantified by Image-Pro Plus 6.0 software (Media
Cybernetic).
Statistical analysis
All experiments were performed in triplicate. All
data are expressed as mean ± SD. One-way analysis of variance
(ANOVA) was used for multiple-group experiments, followed by the
Tukey's test, using SPSS 22.0 software (IBM Corp., Armonk, NY,
USA). Significance was set at a probability value of P<0.05.
Results
Theaflavin shows neuroprotective
effects in rats subjected to cerebral I/R injury
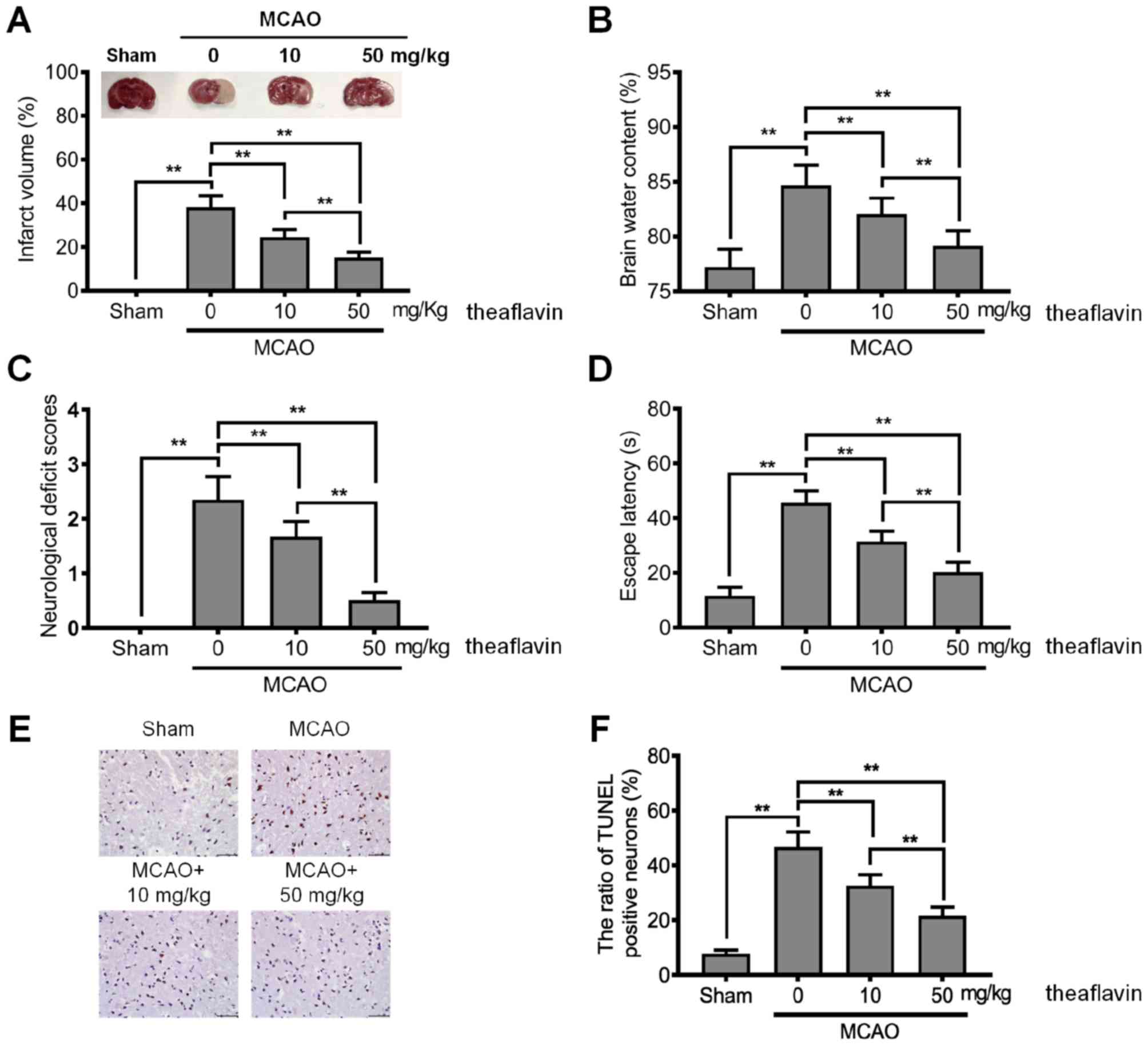
To explore the neuroprotective effects of
theaflavin, its effects in regards to reducing neurological
deficits in rats of the MCAO model were investigated. As shown in
Fig. 1A, the MCAO model rats had a
significantly higher infarct volume than those in the sham group
(P<0.01), indicating that the MCAO model was successfully
constructed. The mean infarct volume of rats treated with
theaflavin was significantly smaller than that of the untreated (0
mg/kg) MCAO model rats and also showed a significant declining
trend as the dose of theaflavin increased (Fig. 1A). Furthermore, the results in
Fig. 1B revealed that theaflavin
treatment significantly reduced brain water content with in a
dose-dependent manner. Brain water content in the MCAO model rats
that received theaflavin at 50 mg/kg was significantly lower than
that in rats that received theaflavin at 10 mg/kg (P<0.01). We
further assessed the effects of theaflavin on the neurological
deficit score, learning and memory ability. As shown in Fig. 1C and D, theaflavin treatment
markedly reduced the neurological deficit score and escape latency.
However, severe neurological deficits and escape latency were
significantly enhanced in the MCAO model (0 mg/kg) relative to the
sham group (P<0.01). The TUNEL results in Fig. 1E and F demonstrated that the number
of TUNEL-positive cells was significantly increased in the MCAO
model rats (0 mg/kg) compared with rats in the sham group
(P<0.01). In contrast to rats in the MCAO group (0 mg/kg), rats
treated with theaflavin 7 days after MCAO showed a significantly
reduced number of TUNEL-positive cells in a dose-dependent manner
(P<0.01) (Fig. 1E and F). The
above data indicate that theaflavin has neuroprotective effects
against cerebral I/R injury.
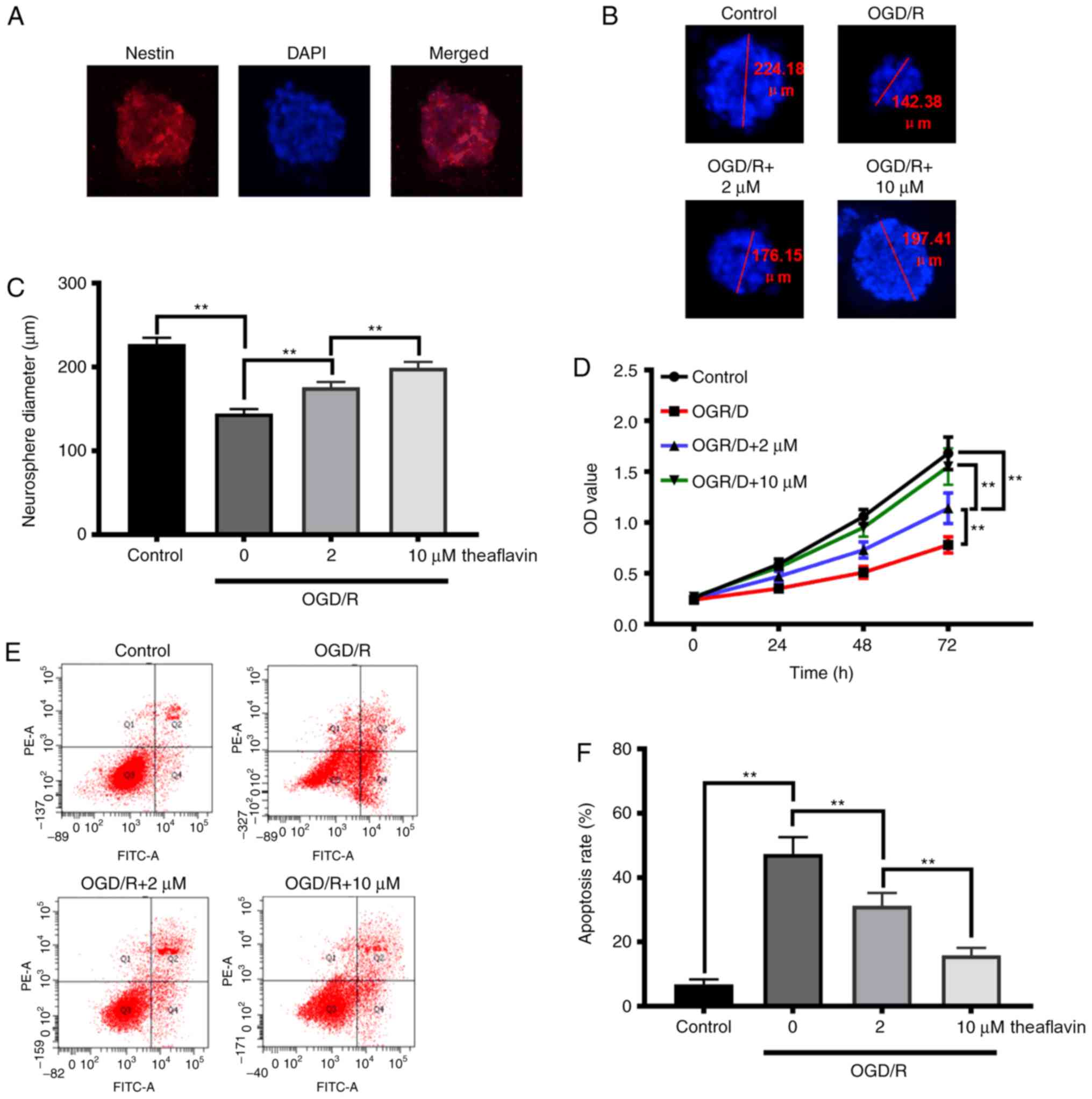
Theaflavin promotes the proliferation
of NSCs and inhibits apoptosis
To elucidate the repair function of theaflavin in
neural function and cognitive impairment, we investigated its
effects on the proliferation and apoptosis of NSCs. The NSCs from
the hippocampus were isolated as shown in Fig. 2A, and results revealed that
theaflavin treatment (2 and 10 µM) significantly increased the
neurosphere diameters of the NSCs subjected to OGD/R (P<0.01)
(Fig. 2B and C). The CCK-8 results
(Fig. 2D) also demonstrated that
theaflavin treatment (2 and 10 µM) significantly enhanced the cell
viability of NSCs subjected to OGD/R in a time-dependent manner.
Moreover, with an increasing dose of theaflavin, NSC viability was
significantly increased (P<0.01) (Fig. 2D). After OGD/R, the apoptotic rate
in NSCs was significantly higher than that in the control group
(P<0.01). Notably, theaflavin treatment (2 and 10 µM)
significantly reduced the apoptotic rate in NSCs subjected to OGD/R
(P<0.01) (Fig. 2E and F). These
data indicate that theaflavin treatment promotes NSC proliferation
and inhibits NSC apoptosis, which would favor the recovery of
neural function and repair of cognitive impairment.
Theaflavin alleviates oxidative stress
in MCAO model rats and NSCs subjected to OGD/R
To investigate the underlying mechanism of
theaflavin in exerting neuroprotective effects, we detected the
antioxidative effects of theaflavin. As illustrated in Table I, the production of ROS and MDA was
evidently increased in the MCAO model rats. Following theaflavin
treatment, the elevated levels of ROS and MDA in the MCAO model
rats were significantly attenuated in a dose-dependent manner
(Table I), suggesting that
theaflavin is capable of suppressing ROS generation and lipid
peroxidation. We then investigated the effects of theaflavin on
antioxidant enzymes in MCAO model rats. The results in Table I show that the activities of SOD
and glutathione (GSH-Px) in the brain were markedly suppressed.
Seven days after theaflavin administration, the activities of SOD
and GSH-Px were significantly elevated. The activities of
antioxidants in the MCAO model rats receiving a theaflavin dose of
50 mg/kg were significantly higher than those receiving a dose of
10 mg/kg (Table I). We further
explored the underlying mechanism of theaflavin in alleviating
oxidative stress in vitro. We found that the production of
ROS and MDA was also markedly increased in the OGD/R cellular model
(Table II), and the activities of
SOD and GSH-Px were significantly suppressed in the OGD/R cellular
model (Table II). The elevated
ROS and MDA levels and the reduced SOD and GSH-Px activities were
significantly attenuated in the OGD/R cellular model after the
administration of theaflavin. Taken together, these results suggest
that theaflavin alleviates oxidative stress in both MCAO ischemic
brains and NSCs subjected to OGD/R.
 | Table I.Theaflavin inhibits oxidative stress
in the rat brain. |
Table I.
Theaflavin inhibits oxidative stress
in the rat brain.
| Group | ROS (F/F0) | MDA (mmol/mg) | SOD (U/mg) | GSH-Px (U/mg) |
|---|
| Sham | 1.38±0.14 | 6.78±0.86 | 125.73±14.38 | 17.71±3.35 |
| MCAO + theaflavin
(0 mg/kg) |
2.34±0.27a |
15.66±1.38a |
58.44±7.83a |
8.63±1.02a |
| MCAO + theaflavin
(10 mg/kg) |
1.97±0.21a,b |
13.05±1.15a,b |
87.59±9.57a,b |
12.58±1.69a,b |
| MCAO + theaflavin
(50 mg/kg) |
1.58±0.17a–c |
9.04±1.02a–c |
103.26±13.21a–c |
14.36±2.07a–c |
| Table II.Theaflavin inhibits oxidative stress
in NSCs. |
Table II.
Theaflavin inhibits oxidative stress
in NSCs.
| Group | ROS (F/F0) | MDA (mmol/mg) | SOD (U/mg) | GSH-Px (U/mg) |
|---|
| Control | 1.21±0.13 | 1.45±0.26 | 16.49±2.36 | 1.52±0.18 |
| OGD/R+ theaflavin
(0 mg/kg) |
2.69±0.30a |
2.93±0.34a |
6.38±1.05a |
0.63±0.09a |
| OGD/R + theaflavin
(2 mg/kg) |
2.12±0.24a,b |
2.16±0.18a,b |
8.97±1.25a,b |
0.91±0.11a,b |
| OGD/R + theaflavin
(10 mg/kg) |
1.64±0.19a–c |
1.74±0.21a–c |
13.26±1.68a–c |
1.34±0.15a–c |
Theaflavin increases the expression of
Nrf2 by downregulating miRNA-128-3p
To further delineate the molecular mechanism by
which theaflavin alleviates oxidative stress, we detected the
expression of Nrf2 in vivo and in vitro. As evident
in Fig. 3A-D, the expression of
Nrf2 was markedly inhibited in MCAO model rats and NSCs subjected
to OGD/R (P<0.01). Seven days after theaflavin administration,
the expression of Nrf2 was significantly enhanced (P<0.01).
Particularly, theaflavin at 50 mg/kg had a stronger ability to
increase Nrf2 levels than that at 10 mg/kg. Moreover, the miRNAs
that target Nrf2 were investigated using bioinformatics methods.
The miRNA-128-3p was predicted to regulate Nrf2 expression using
the TargetScan tool (Fig. 3E). The
results in Fig. 3F demonstrated
that the miRNA-128-3p level was significantly upregulated in the
MCAO model rats (0 mg/kg theaflavin) compared to the Sham group
(P<0.01). After transfection of NSCs with the miRNA mimics or
miRNA inhibitor, the change in relative expression levels of
miRNA-128-3p were significantly increased or decreased,
respectively (Fig. 3G). A
significant increase in miRNA-128-3p levels was also found in the
NSCs subjected to OGD/R (P<0.01) (Fig. 4H). Notably, we found that
theaflavin reduced the level of miRNA-128-3p levels significantly
in both the MCAO and OGD/R NSC models in a dose-dependent manner
(P<0.01) (Fig. 3F and H). The
results presented in Fig. 3H also
indicated that miRNA mimics + (10 µM) theaflavin treatment
increased the level of miRNA-128-3p, in contrast to our
observations in NSCs subjected to OGD/R, indicating that theaflavin
induced the downregulation of miRNA-128-3p. To confirm whether
miRNA-128-3p inhibited Nrf2 expression, a dual-luciferase reporter
assay was employed using the wild-type (WT) site or the mutant
(MUT) site. A considerable decline in luciferase activity was
evident with co-transfection of the miRNA-128-3p mimic and reporter
plasmid at the Nrf2 WT site (Fig.
3I) (P<0.01). However, a slight reduction in luciferase
activity was observed after co-transfection with the miRNA-128-3p
mimic and reporter plasmid at the Nrf2 MUT site (Fig. 3I). To further demonstrate the
effect of miRNA-128-3p on Nrf2, the levels of Nrf2 mRNA and
protein were measured using qRT-PCR and western blot analysis. As
shown in Fig. 3J and K, ectopic
expression of miRNA-128-3p induced a significant decline in the
levels of Nrf2 mRNA and protein. These results indicate that
theaflavin downregulates miRNA-128-3p to increase Nrf2 levels to
alleviate oxidative stress.
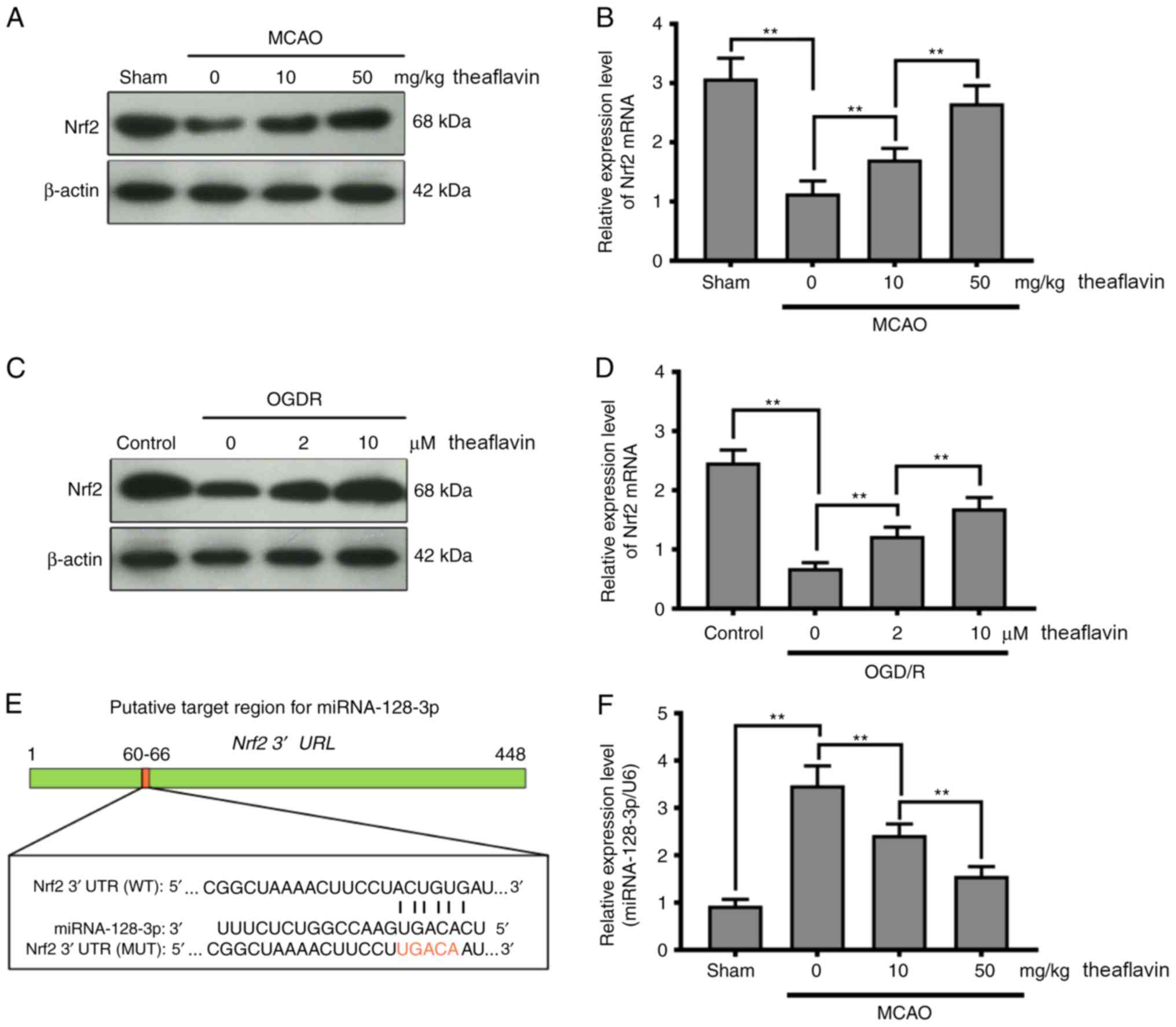 | Figure 3.Direct inhibition of Nrf2 expression
by miRNA-128-3p via targeting of the 3′ UTR in NSCs. (A) Western
blot analysis shows the Nrf2 levels 24 h after cerebral I/R in the
MCAO model with or without theaflavin treatment (0, 10 and 50
mg/kg). β-actin was used as the internal control for normalization.
(B) RT-qPCR analysis shows Nrf2 mRNA levels 24 h after cerebral I/R
in the MCAO model with or without theaflavin treatment (0, 10 and
50 mg/kg). GAPDH mRNA was used as the internal control for
normalization. (C) Western blot analysis shows Nrf2 levels in NSCs
subjected to OGD/R with or without theaflavin treatment (0, 2 and
10 µM). β-actin was used as the internal control for normalization.
(D) RT-qPCR analysis shows Nrf2 mRNA levels in NSCs subjected to
OGD/R with or without theaflavin treatment (0, 2 and 10 µM).
GAPDH mRNA was used as the internal control for
normalization. (E) Sequence alignment between miRNA-128-3p and the
3′UTR of Nrf2 mRNA. (F) RT-qPCR analysis shows miRNA-128-3p levels
24 h after cerebral I/R in the MCAO model with or without
theaflavin treatment (0, 10 and 50 mg/kg). U6 was used as the
internal control for normalization. (G) RT-qPCR analysis shows
miRNA-128-3p levels after transfection of miRNA mimics or miRNA
inhibitor. (H) RT-qPCR analysis shows miRNA-128-3p levels in NSCs
subjected to OGD/R with or without theaflavin treatment (0, 2 and
10 µM) or with 10 µM theaflavin and miRNA mimic. U6 was used as the
internal control for normalization. (I) The luciferase reporter
gene assay shows the effect of miRNA-128-3p on luciferase activity.
(J) RT-qPCR analysis shows Nrf2 mRNA levels in NSCs subjected to
OGD/R after transfection of mimics (K) Western blot analysis shows
the Nrf2 levels in NSCs subjected to OGD/R after transfection of
mimics. All assays were performed in triplicate. Data are expressed
as mean ± SD. **P<0.01. Nrf2, nuclear factor (erythroid-derived
2)-related factor 2; UTR, untranslated region; NSCs, neural stem
cells; I/R, ischemia-reperfusion; MCAO, middle cerebral artery
occlusion; OGD/R, oxygen-glucose deprivation and reoxygenation. |
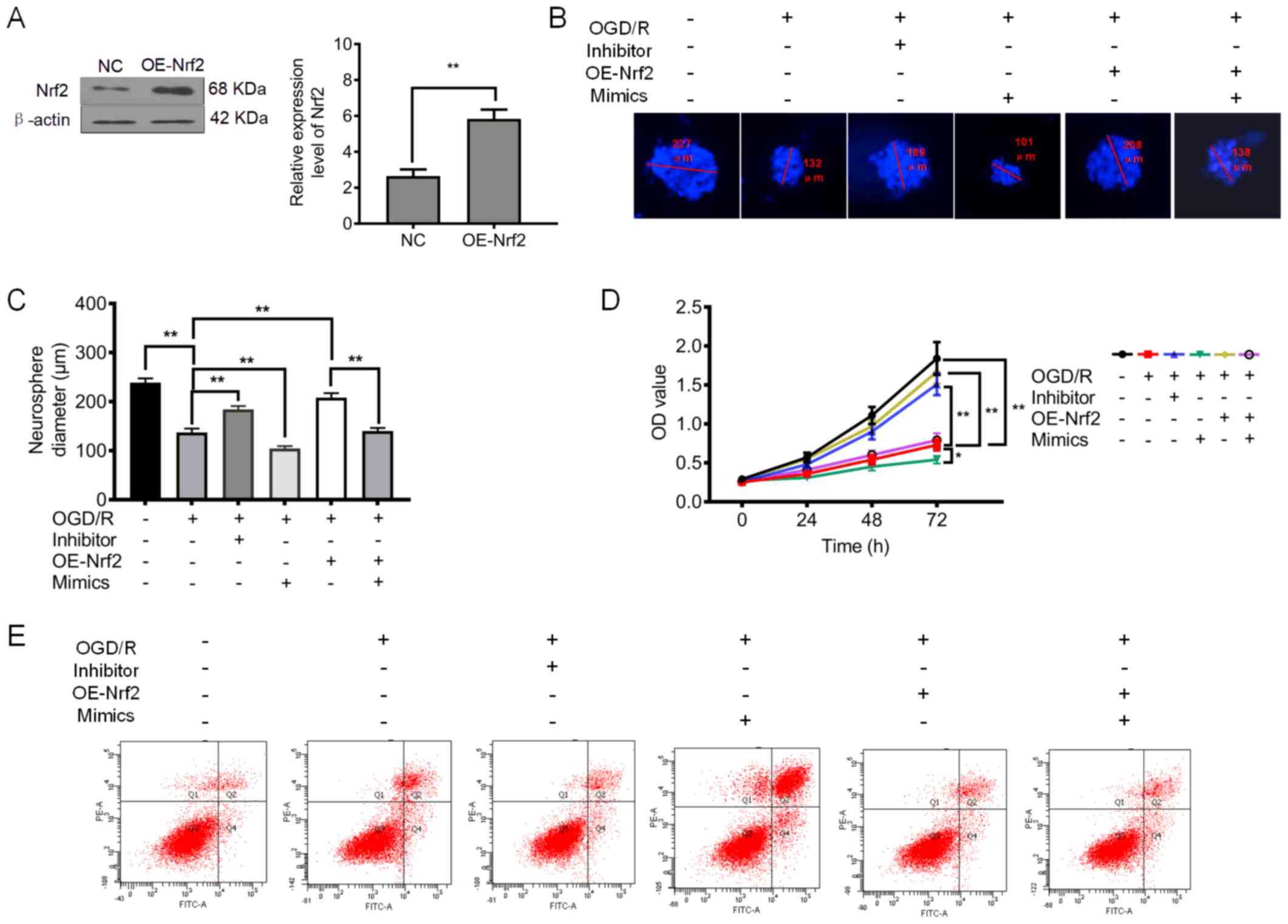 | Figure 4.Downregulation of miRNA-128-3p and
upregulation of Nrf2 promote cell proliferation and inhibit
apoptosis and oxidative stress. (A) Relative expression level of
Nrf2 after upregulation of Nrf2 (OE-Nrf2). (B) Representative
neurosphere images in NSCs subjected to OGD/R after downregulation
or upregulation of miRNA-128-3p (with inhibitor and mimics,
respectively), or upregulation of Nrf2 (OE-Nrf2). (C) Quantitative
analysis of neurosphere diameter by Image J software. (D) Cell
Counting Kit-8 (CCK-8) assay shows the cell viability of NSCs
subjected to OGD/R after downregulation or upregulation of
miRNA-128-3p (with inhibitor and mimics, respectively), or
upregulation of Nrf2 (OE-Nrf2). (E) Flow cytometry shows the
apoptosis of NSCs subjected to OGD/R after downregulation of
miRNA-128-3p (Inhibitor) or upregulation of Nrf2 (OE-Nrf2). (F)
Quantitative analysis of the apoptotic rate. (G) ROS levels in NSCs
subjected to OGD/R after after downregulation or upregulation of
miRNA-128-3p (with inhibitor and mimics, respectively), or
upregulation of Nrf2 (OE-Nrf2). (H) MDA content in NSCs subjected
to OGD/R after downregulation of miRNA-128-3p (Inhibitor) or
upregulation of Nrf2 (OE-Nrf2). (I) GSH-Px levels in NSCs subjected
to OGD/R after downregulation or upregulation of miRNA-128-3p (with
inhibitor and mimics, respectively), or upregulation of Nrf2
(OE-Nrf2). (J) SOD activity in NSCs subjected to OGD/R after
downregulation of miRNA-128-3p (Inhibitor) or upregulation of Nrf2
(OE-Nrf2). All assays were performed in triplicate. Data are
expressed as the mean ± SD. *P<0.05, **P<0.01. Nrf2, nuclear
factor (erythroid-derived 2)-related factor 2; NSCs, neural stem
cells; OGD/R, oxygen-glucose deprivation and reoxygenation; MDA,
malondialdehyde; GSH-Px, glutathione peroxidase; SOD, superoxide
dismutase. |
Inhibition of NSC proliferation and
enhancement of oxidative stress by miRNA-128-3p
To further confirm the regulatory function of
miRNA-128-3p, we examined its effects on NSC proliferation.
Furthermore, we also upregulated the expression of Nrf2 (Fig. 4A). The results presented in
Fig. 4B-D demonstrated that the
application of the miRNA-128-3p inhibitor significantly increased
the neurosphere diameter and cell viability of the NSCs subjected
to OGD/R (P<0.01). Notably, both the ectopic expression of
miRNA-128-3p and Nrf2 slightly increased the neurosphere diameter
and cell viability of NSCs subjected to OGD/R (P<0.01).
Furthermore, it was found that upregulation of Nrf2 (OE-Nrf2) was
capable of increasing the neurosphere diameter and cell viability,
an effect that was abolished after treatment with miRNA-128-3p
mimics (Fig. 4B-D). As shown in
Fig. 4E and F, the elevated
apoptotic rate in NSCs was significantly attenuated by the
miRNA-128-3p inhibitor (P<0.01). Whereas the apoptotic rate in
NSCs receiving both miRNA-128-3p mimics and overexpression of Nrf2
was slightly reduced compared with that in NSCs subjected to OGD/R
(P>0.05). In addition, upregulation of Nrf2 showed a stronger
ability to reduce the rate of apoptosis, an effect which was
abolished after treatment with miRNA-128-3p mimics (P<0.01).
These findings indicate that downregulation of miRNA-128-3p
promoted NSC proliferation by upregulating Nrf2 expression.
We investigated the effect of miRNA-128-3p on
oxidative stress in NSCs subjected to OGD/R. The levels of ROS and
MDA were significantly reduced after treatment with miRNA-128-3p
inhibitors (P<0.01) (Fig. 4G and
H), and were slightly reduced after treatment with miRNA-128-3p
mimics and theaflavin, the elevated ROS and MDA levels were
slightly attenuated. As expected, upregulation of Nrf2
significantly inhibited the production of ROS and MDA, an effect
which was abolished after treatment with miRNA-128-3p mimics
(Fig. 4G and H). These data
suggest that reducing miRNA-128-3p levels is beneficial for the
inhibition of ROS generation and lipid peroxidation. Further
studies showed that the miRNA-128-3p inhibitor and elevated Nrf2
levels significantly upregulated the levels of antioxidant enzymes,
including GSH-Px and SOD (Fig. 4I and
J). Particularly, treatment with miRNA-128-3p mimics and Nrf2
overexpression were unable to increase the levels of the
antioxidant enzymes (P>0.05) (Fig.
4I and J). Collectively, these results indicate that reduced
miRNA-128-3p levels are beneficial to the restoration of an
impaired antioxidant defense system.
Discussion
Nowadays, ischemic stroke presents a considerable
social and family burden. Although the reperfusion of ischemic
cerebral tissue is the most common way to rescue ischemic stroke,
CI/RI can further lead to brain damage and even death (24,25).
Therefore, new strategies must be developed to prevent CI/RI. In
recent years, plant extracts have garnered increasing attention
from academia and the public. Previously, studies have shown that
theaflavin is a promising active compound from black tea that can
regulate inflammation and oxidative stress (17,26).
However, the biological function and the molecular mechanism of
theaflavin in ameliorating ischemic damage has not yet been fully
elucidated. In the present study, we explored the biological
function of theaflavin in ameliorating CI/RI and investigated its
potential molecular mechanism. We found that theaflavin exerts
neuroprotective effects by reducing infarct volume and improving
learning and memory in rats subjected to CI/RI. Furthermore, we
provided convincing evidence that theaflavin treatment can reduce
apoptosis in NSCs and promote NSC proliferation. These findings
were supported by the results of the MCAO injury rat model, as well
as those of the cultured NSCs subjected to OGD/R. Importantly, we
proved that theaflavin could be a Nrf2 promoter by downregulating
miRNA-128-3p, which effectively inhibited oxidative stress in rats
subjected to CI/RI and NSCs subjected to OGD/R. Our results
collectively indicate that theaflavin could be a promising natural
compound for the attenuation of CI/RI. To our knowledge, we found a
new molecular mechanism for the neuroprotective function of
theaflavin downregulating miRNA-128-3p to inhibit oxidative stress
to further promote repair following ischemic stroke.
Most of the biological functions of tea polyphenols,
including theaflavins and catechins, have been ascribed to their
anti-oxidative, anti-tumor, anti-bacterial, anti-inflammatory,
anti-viral, and cardiovascular protective activities (27–29).
For example, Zhang et al found that theaflavin showed
neuroprotective effects against oxidative stress in PC12 cells,
which was derived from the suppression of oxidant enzyme activity
(16). In the present study,
theaflavin treatment commenced during the development of CI/RI in
rats. Theaflavin was evidently capable of reducing infarct volume
and ameliorating symptoms related to CI/RI. Importantly, these
inhibitory effects of theaflavin were observed in a dose-dependent
manner. Our results indicate that theaflavin has a neuroprotective
function in CI/RI. To further investigate the role of theaflavin in
its neuroprotective effects in CI/RI, TUNEL staining was used in
the present study. Theaflavin decreased the rate of apoptosis in
the brain tissue of rats subjected to CI/RI. Recently, several
research studies have shown that neurogenesis is insufficient for
functional recovery in ischemic stroke disability (30,31).
Therefore, targeting endogenous NSCs to promote adult neurogenesis
is a feasible strategy for the recovery of nervous function and
repair of cognitive impairment in ischemic stroke. We isolated NSCs
from the hippocampus, which is responsible for long-term memory
storage conversion and orientation functions. Following theaflavin
treatment, the cell viability of NSCs subjected to OGD/R was
significantly increased and the apoptosis rate showed a significant
decline. These results further indicate that theaflavin has a
neuroprotective function.
Oxidative stress is a critically important
pathological process in CI/RI that contributes to neurocyte injury
(32). Neural tissue is vulnerable
to oxidative attack, owing to its high consumption of oxygen,
relatively low antioxidant capacity, and high polyunsaturated fatty
acid content (33). Moreover,
oxidative injury of mitochondrial DNA would lead to impaired energy
regulation, which is critical in high-energy-requiring neurons
(8,34). Current research has shown that
plant extracts exert strong neuroprotective effects by inhibiting
oxidative stress. As a result of the anti-oxidative function of
theaflavin, we explored its role in inhibiting oxidative stress in
CI/RI. Seven days after theaflavin treatment, the activities of
antioxidants in both the MCAO model rats and NSCs subjected to
OGD/R were significantly elevated. In addition, the elevated ROS
and MDA levels were significantly attenuated in both the MCAO model
rats and NSCs subjected to OGD/R after theaflavin treatment,
indicating that theaflavin also exhibits a stronger anti-oxidative
capacity in CI/RI. Our findings are largely consistent with a
recent report, in which theaflavins showed significant protective
effects in erythrocytes against oxidative stress induced by
tert-butyl hydroperoxide by increasing GSH-Px and reducing MDA
(35). The alleviation of
oxidative stress by theaflavin is responsible for NSC proliferation
and anti-apoptosis, which aids in the recovery of nervous function
and the repair of cognitive impairment. The Nrf2-dependent pathway
plays a critical role in oxidative stress. Recent research reported
that theaflavin could ameliorate ionizing radiation-induced
hematopoietic stem cell injury via the Nrf2 pathway (36). Thus, the expression of Nrf2 in
CI/RI was investigated. We found that the Nrf2 protein and
Nrf2 mRNA levels were obviously up-regulated by theaflavin
in both the MCAO rat model and NSCs subjected to OGD/R. Thus, we
believe that theaflavin may attenuate CI/RI by inhibiting oxidative
stress via the Nrf2-dependent pathway. These results suggest the
feasibility of theaflavin as a treatment for ischemic stroke and
neurological recovery.
The miRNAs are crucial regulators in spontaneous
neurogenesis and sustainability of neuronal phenotype by
post-transcriptional regulation. Furthermore, miRNAs, including
miRNA-23a-3p, miRNA-424, and miRNA-125b reportedly reduce CI/RI by
targeting the oxidative stress pathway (37–39).
Based on these functions of miRNA, Nrf2 mRNA/miRNA interaction was
predicted using the miRNA target prediction tool, TargetScan, and
Nrf2 mRNA/miRNA-128-3p interaction information was also predicted.
Our results showed that the significant increase in miRNA-128-3p
was eliminated following theaflavin treatment in both the MCAO
ischemic rats and NSCs subjected to OGD/R. Interestingly, ectopic
expression of miRNA-128-3p resulted in a significant decline in the
levels of Nrf2 mRNA and protein, which further confirmed the
regulatory effect of miRNA-128-3p on Nrf2. Thus, theaflavin
improved the expression of Nrf2 by downregulating miRNA-128-3p.
Several studies have shown that abnormal expression of miRNA-128-3p
is closely related to I/RI, contextual learning, and DNA damage
response (40,41). For example, Chen et al found
that inhibition of miR-128-3p could protect human cardiomyocytes
from I/RI (42). Loss-of-function
experiments in our study showed that downregulation of miRNA-128-3p
promoted NSC proliferation and reduced the rate of apoptosis, which
is consistent with previous reports that antagonism of miR-128-3p
could remarkably improve the proliferation of lymphatic endothelial
cells (43). Furthermore,
miRNA-128-3p exhibited opposite regulatory effects to Nrf2 in
oxidative stress. Thus, downregulation of miRNA-128-3p expression
along with theaflavin treatment contribute to the attenuation of
CI/RI. Chen et al demonstrated that the downregulation of
miRNA-128-3p in mouse hippocampi promoted the differentiation of
NSCs in the performance of the water maze task (41). However, the research of Mao et
al showed the opposite conclusion, as they reported that an
increase in miRNA-128-3p levels contributed to neuronal survival in
ischemia-induced brain injury (44). Such inconsistency may be that due
to different animal models and experimental conditionsled to a
disparity in observations. Our findings present a comprehensive
understanding of the underlying mechanism of theaflavin in ischemic
stroke.
In conclusion, theaflavin could effectively
attenuate CI/RI and improve learning and memory through
miRNA-128-3p-mediated anti-oxidative actions in rats subjected to
ischemia-induced brain injury and NSCs subjected to OGD/R. Further
studies can focus on the regulatory mechanism of theaflavin on
miRNA-128-3p expression, high-energy-requiring neurons and the
effect of theaflavin on NSC differentiation. In all, theaflavin
holds the potential to be a new drug candidate for ischemic stroke
treatment.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant (grant
no. 201640129) from Shanghai Municipal Planning Commission of
science and Research Fund grant (grant no. 2018YFA0107900) from
National Key R&D Program of China; grant (grant no. 81200936)
from the National Nature Science Foundation, and grant (grant no.
134119a8500) from Shanghai Committee of Science and Technology.
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
RL and JZ concieved of and designed the study. HW
provided administrative support. XL and ZY collected and assembled
the data. LF analysed and interpreted the data. RL wrote the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Della-Morte D, Pacifici F and Rundek T:
Genetic susceptibility to cerebrovascular disease. Curr Opin
Lipidol. 27:187–195. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hu X, De Silva TM, Chen J and Faraci FM:
Cerebral vascular disease and neurovascular injury in ischemic
stroke. Circ Res. 120:449–471. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gerschenfeld G, Muresan IP, Blanc R,
Obadia M, Abrivard M, Piotin M and Alamowitch S: Two paradigms for
endovascular thrombectomy after intravenous thrombolysis for acute
ischemic stroke. JAMA Neurol. 74:549–556. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ji K, Xue L, Cheng J and Bai Y:
Preconditioning of H2S inhalation protects against cerebral
ischemia/reperfusion injury by induction of HSP70 through
PI3K/Akt/Nrf2 pathway. Brain Res Bull. 121:68–74. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xu J, Kong X, Xiu H, Dou Y, Wu Z and Sun
P: Combination of curcumin and vagus nerve stimulation attenuates
cerebral ischemia/reperfusion injury-induced behavioral deficits.
Biomed Pharmacother. 103:614–620. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen X, Wu H, Chen H, Wang Q, Xie XJ and
Shen J: Astragaloside VI promotes neural stem cell proliferation
and enhances neurological function recovery in transient cerebral
ischemic injury via activating EGFR/MAPK signaling cascades. Mol
Neurobiol. 56:3053–3067. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu J, Chen Y, Yu S, Li L, Zhao X, Li Q,
Zhao J and Zhao Y: Neuroprotective effects of sulfiredoxin-1 during
cerebral ischemia/reperfusion oxidative stress injury in rats.
Brain Res Bull. 132:99–108. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang M, Li YJ, Ding Y, Zhang HN, Sun T,
Zhang K, Yang L, Guo YY, Liu SB, Zhao MG and Wu YM: Silibinin
prevents autophagic cell death upon oxidative stress in cortical
neurons and cerebral ischemia-reperfusion injury. Mol Neurobiol.
53:932–943. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Guo C, Wang S, Duan J, Jia N, Zhu Y, Ding
Y, Guan Y, Wei G, Yin Y, Xi M and Wen A: Protocatechualdehyde
protects against cerebral ischemia-reperfusion-induced oxidative
injury via protein kinase Cε/Nrf2/HO-1 pathway. Mol Neurobiol.
54:833–845. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Khajevand-Khazaei MR, Azimi S, Sedighnejad
L, Salari S, Ghorbanpour A, Baluchnejadmojarad T, Mohseni-Moghaddam
P, Khamse S and Roghani M: S-allyl cysteine protects against
lipopolysaccharide-induced acute kidney injury in the C57BL/6 mouse
strain: Involvement of oxidative stress and inflammation. Int
Immunopharmacol. 69:19–26. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen M, Dai LH, Fei A, Pan SM and Wang HR:
Isoquercetin activates the ERK1/2-Nrf2 pathway and protects against
cerebral ischemia-reperfusion injury in vivo and in
vitro. Exp Ther Med. 13:1353–1359. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang W, Wei R, Zhang L, Tan Y and Qian C:
Sirtuin 6 protects the brain from cerebral ischemia/reperfusion
injury through NRF2 activation. Neuroscience. 366:95–104. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chang C, Zhao Y, Song G and She K:
Resveratrol protects hippocampal neurons against cerebral
ischemia-reperfusion injury via modulating JAK/ERK/STAT signaling
pathway in rats. J Neuroimmunol. 315:9–14. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jiao S, Zhu H, He P and Teng J: Betulinic
acid protects against cerebral ischemia/reperfusion injury by
activating the PI3K/Akt signaling pathway. Biomed Pharmacother.
84:1533–1537. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Feng Q, Torii Y, Uchida K, Nakamura Y,
Hara Y and Osawa T: Black tea polyphenols, theaflavins, prevent
cellular DNA damage by inhibiting oxidative stress and suppressing
cytochrome P450 1A1 in cell cultures. J Agric Food Chem.
50:213–220. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang J, Cai S, Li J, Xiong L, Tian L, Liu
J, Huang J and Liu Z: Neuroprotective effects of theaflavins
against oxidative stress-induced apoptosis in PC12 cells. Neurochem
Res. 41:3364–3372. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cai F, Li CR, Wu JL, Chen JG, Liu C, Min
Q, Yu W, Ouyang CH and Chen JH: Theaflavin ameliorates cerebral
ischemia-reperfusion injury in rats through its anti-inflammatory
effect and modulation of STAT-1. Mediators Inflamm. 2006:304902006.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liang Q, Luo Z, Zeng J, Chen W, Foo SS,
Lee SA, Ge J, Wang S, Goldman SA, Zlokovic BV, et al: Zika virus
NS4A and NS4B proteins deregulate Akt-mTOR signaling in human fetal
neural stem cells to inhibit neurogenesis and induce autophagy.
Cell Stem Cell. 19:663–671. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li M, Liao YJ, Hou GH, Yang ZB and Zuo ML:
Monosialotetrahexosylganglioside protect cerebral
ischemia/reperfusion injury through upregulating the expression of
tyrosine hydroxylase by inhibiting lipid peroxidation. Biomed
Pharmacother. 84:1923–1929. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hodge RD, Nelson BR, Kahoud RJ, Yang R,
Mussar KE, Reiner SL and Hevner RF: Tbr2 is essential for
hippocampal lineage progression from neural stem cells to
intermediate progenitors and neurons. J Neurosci. 32:6275–6287.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen T, Yu Y, Tang LJ, Kong L, Zhang CH,
Chu HY, Yin LW and Ma HY: Neural stem cells over-expressing
brain-derived neurotrophic factor promote neuronal survival and
cytoskeletal protein expression in traumatic brain injury sites.
Neural Regen Res. 12:433–439. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Longa EZ, Weinstein PR, Carlson S and
Cummins R: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Benakis C, Brea D, Caballero S, Faraco G,
Moore J, Murphy M, Sita G, Racchumi G, Ling L, Pamer EG, et al:
Commensal microbiota affects ischemic stroke outcome by regulating
intestinal γδ T cells. Nat Med. 22:516–523. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Choudhury GR and Ding S: Reactive
astrocytes and therapeutic potential in focal ischemic stroke.
Neurobiol Dis. 85:234–244. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Imran A, Arshad MU, Arshad MS, Imran M,
Saeed F and Sohaib M: Lipid peroxidation diminishing perspective of
isolated theaflavins and thearubigins from black tea in arginine
induced renal malfunctional rats. Lipids Health Dis. 17:1572018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tipoe GL, Leung TM, Hung MW and Fung ML:
Green tea polyphenols as an anti-oxidant and anti-inflammatory
agent for cardiovascular protection. Cardiovasc Hematol Disord Drug
Targets. 7:135–144. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fan FY, Sang LX and Jiang M: Catechins and
their therapeutic benefits to inflammatory bowel disease.
Molecules. 22(pii): E4842017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bernatoniene J and Kopustinskiene DM: The
role of catechins in cellular responses to oxidative stress.
Molecules. 23(pii): E9652018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang Y, Xu D, Qi H, Yuan Y, Liu H, Yao S,
Yuan S and Zhang J: Enriched environment promotes post-stroke
neurogenesis through NF-κB-mediated secretion of IL-17A from
astrocytes. Brain Res. 1687:20–31. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang J, Fu X, Zhang D, Yu L, Li N, Lu Z,
Gao Y, Wang M, Liu X, Zhou C, et al: ChAT-positive neurons
participate in subventricular zone neurogenesis after middle
cerebral artery occlusion in mice. Behav Brain Res. 316:145–151.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li P, Shen M, Gao F, Wu J, Zhang J, Teng F
and Zhang C: An antagomir to MicroRNA-106b-5p ameliorates cerebral
ischemia and reperfusion injury in rats via inhibiting apoptosis
and oxidative stress. Mol Neurobiol. 54:2901–2921. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tao X, Sun X, Xu L, Yin L, Han X, Qi Y, Xu
Y, Zhao Y, Wang C and Peng J: Total flavonoids from Rosa
laevigata Michx fruit ameliorates hepatic ischemia/reperfusion
injury through inhibition of oxidative stress and inflammation in
rats. Nutrients. 8(pii): E4182016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li K, Ding D and Zhang M: Neuroprotection
of osthole against cerebral ischemia/reperfusion injury through an
Anti-apoptotic pathway in rats. Biol Pharm Bull. 39:336–342. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yoshino K, Hara Y, Sano M and Tomita I:
Antioxidative effects of black tea theaflavins and thearubigin on
lipid peroxidation of rat liver homogenates induced by tert-butyl
hydroperoxide. Biol Pharm Bull. 17:146–149. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Han X, Zhang J, Xue X, Zhao Y, Lu L, Cui
M, Miao W and Fan S: Theaflavin ameliorates ionizing
radiation-induced hematopoietic injury via the NRF2 pathway. Free
Radic Biol Med. 113:59–70. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu P, Zhao H, Wang R, Wang P, Tao Z, Gao
L, Yan F, Liu X, Yu S, Ji X and Luo Y: MicroRNA-424 protects
against focal cerebral ischemia and reperfusion injury in mice by
suppressing oxidative stress. Stroke. 46:513–519. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhao H, Tao Z, Wang R, Liu P, Yan F, Li J,
Zhang C, Ji X and Luo Y: MicroRNA-23a-3p attenuates oxidative
stress injury in a mouse model of focal cerebral
ischemia-reperfusion. Brain Res. 1592:65–72. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liang Y, Xu J, Wang Y, Tang JY, Yang SL,
Xiang HG, Wu SX and Li XJ: Inhibition of MiRNA-125b decreases
cerebral ischemia/reperfusion injury by targeting CK2α/NADPH
oxidase signaling. Cell Physiol Biochem. 45:1818–1826. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang R, Liu C, Niu Y, Jing Y, Zhang H,
Wang J, Yang J, Zen K, Zhang J, Zhang CY and Li D: MicroRNA-128-3p
regulates mitomycin C-induced DNA damage response in lung cancer
cells through repressing SPTAN1. Oncotarget. 8:58098–58107. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen J, Li W, Li Y, He S, Li L, Liang L,
Song Y, Qin D and Zheng H: MicroRNA-128-3p impaired water maze
learning by suppressing Doublecortin expression in both wild type
and Aβ-42 infused mice. Neurosci Lett. 626:79–85. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen GH, Xu CS, Zhang J, Li Q, Cui HH, Li
XD, Chang LP, Tang RJ, Xu JY, Tian XQ, et al: Inhibition of
miR-128-3p by tongxinluo protects human cardiomyocytes from
Ischemia/reperfusion injury via Upregulation of p70s6k1/p-p70s6k1.
Front Pharmacol. 8:7752017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhou J, He Z, Guo L, Zeng J, Liang P, Ren
L, Zhang M, Zhang P and Huang X: MiR-128-3p directly targets
VEGFC/VEGFR3 to modulate the proliferation of lymphatic endothelial
cells through Ca2+ signaling. Int J Biochem Cell Biol.
102:51–58. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mao G, Ren P, Wang G, Yan F and Zhang Y:
MicroRNA-128-3p protects mouse against cerebral ischemia through
reducing p38α mitogen-activated protein kinase activity. J Mol
Neurosci. 61:152–158. 2017. View Article : Google Scholar : PubMed/NCBI
|