Introduction
Radiotherapy (RT) is widely used to treat cancer
because of its ability to control tumor growth; RT can destroy
cancerous tissue and cause tumor cell necrosis (1). RT induces changes in the tumor
microenvironment and promotes the expression, and release of,
tumor-related antigens, thereby activating the antitumor immune
response (2). RT also induces
tumor cells to produce cytokines, such as tumor necrosis factor
(TNF)-α, which attract effector T cells to the target region,
increasing the expression of damage-associated molecular patterns
on the surface of tumor cell, which induces ‘immunogenic cell
death’ in tumor cells (3,4). Previous studies have reported that RT
can upregulate the expression of CD70 on dendritic cells (DCs) and
induce T cells to produce interferon (IFN)-γ (5,6).
CD8+ T cells are activated after RT combined with
immunotherapy (7). Together, RT
and immunotherapy increase the antigen presenting ability of DCs
and increase the number of antigens on the surface of tumor cells,
inducing T cell-mediated clearance (8,9).
However, the upregulation of regulatory T cells
(Tregs), and some inhibitory receptors and ligands, including
programmed cell death 1 (PD-1), programmed cell death ligand 1
(PD-L1), T cell immunoglobulin domain and mucin domain 3 (TIM-3),
B- and T-lymphocyte attenuator (BTLA) and galectin-9, prevents RT
from achieving the optimal therapeutic effect (10). These molecules are major biological
markers of T cell exhaustion, which leads to reduced T cell
proliferation and cytotoxicity, and increased apoptosis. The
antitumor cytotoxic effect and the adaptive immunotherapy effect of
T cells is reduced, which leads to the occurrence, development and
metastasis of tumors (11).
Therefore, RT combined with immune checkpoint blockade has
attracted considerable attention and has already achieved some
success. PD-1 blockade combined with RT is among the most effective
immunotherapies for cancer (12,13);
however, a substantial proportion of patients gradually develop
treatment resistance (14–16). Currently, the mechanism of drug
resistance remains uncertain, reflecting the need for a new
strategy (17).
One potential explanation for tumor immune escape
and resistance involves the immaturity of DCs, and the exhaustion
of T cells (18,19). Indoleamine 2,3-dioxygenase (IDO) is
an important regulatory factor in tumor-mediated immunosuppression
that has a negative effect on DCs and T cells (20).
As the tryptophan catabolic enzyme involved in the
initiation and rate-limiting steps, IDO is highly expressed in
various types of cancer, as well as in immune cells, and plays an
important role in immune cell suppression (15,21).
In humans and murine models, the expression of IDO has been found
to be a prominent predictor of a poor prognosis (19), which may be related to its negative
regulatory effect on DCs and T cells. IDO overexpression in DCs can
affect their maturation, resulting in a decreased
antigen-presenting ability and increased negative co-stimulatory
molecule expression. These effects also allow IDO to indirectly
influence local T cells (20).
Additionally, IDO supplementation induces Tregs, which contribute
to the immunosuppressive effects (22). IDO can mediate the depletion of the
essential amino acid tryptophan, causing tryptophan starvation
(11). In the middle of the G1
phase, T cells are extremely sensitive to tryptophan deficiency,
which causes protein insufficiency and blockade in the G1 phase.
This effect can increase the susceptibility of T cells to apoptosis
(14). Together, the negative
effects of IDO on DCs and T cells leads to immunosuppression in
tumor cells.
Researchers have expressed interest in developing
IDO inhibitors as new immunotherapies to treat cancer (23). Although 1-methyl-tryptophan (1MT)
has not achieved the desired anticancer effect in recent clinical
trials (23), as a general
therapeutic approach, IDO inhibitors still have potential and
research value. IDO inhibition combined with RT may provide a
better therapeutic effect, and may partially solve the problem of
tumor immunosuppression. The present study aimed to investigate
whether an IDO inhibitor synergized with RT can further inhibit
tumor growth by activating DCs and antagonizing T cell
exhaustion.
Materials and methods
Cell line and animals
Lewis lung cancer (LLC) cells, a murine tumor cell
line, were obtained from the American Type Culture Collection and
maintained in RPMI 1640 medium (HyClone; GE Healthcare Life
Sciences) with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.), 100
U/ml penicillin (Gibco; Thermo Fisher Scientific, Inc.) and 100
µg/ml streptomycin (Gibco; Thermo Fisher Scientific, Inc.) at 37°C
in 5% CO2. The media was changed every other day and the
cells were digested with 0.25% trypsin for 2–3 min in the
logarithmic growth phase from passages 1–4. Mycoplasma testing was
completed for the cell line.
C57BL/6 female mice (age, 6–8 weeks old; weight,
20–25 g) were purchased from the animal breeding facility of Hunan
Slack Jingda Experimental Animal Co., Ltd. (cat. no.
43004700049723). The mice were maintained under controlled
conditions (20-23°C, 40–70% humidity and a 12-h light/dark cycle).
Mice ate ~5 g chow per day and drank ~7 ml water. All animal
experiments were performed in accordance with the Animal Research
Reporting of in vivo Experiments and the National Institutes
of Health guidelines for animal welfare, and the study was approved
by the Laboratory Animal Ethics Committee of Nanchang University
(permit no. NCDXSYDWFL-2015097). All animal experiments complied
with the ARRIVE guidelines and the 2013 AVMA euthanasia guidelines
(24).
Tumor models and treatment
protocols
Tumors were generated on the backs of mice by
subcutaneously injecting 1×105 LLC cells in PBS. Tumor
size was measured using a caliper every day after the tumors
appeared, and the tumor volume was calculated using the following
formula: Tumor volume=0.5× (tumor length) × (tumor
width)2. The mice were monitored daily and euthanized
using CO2 inhalation (the flow rate of CO2
was 20% of the chamber volume/minute). Mice were euthanized if any
of the following conditions were observed: A body condition score
of 1/5 (mouse is emaciated: Skeletal structure is extremely
prominent; little or no flesh cover, or vertebrae distinctly
segmented), a body condition score of 2/5 (mouse is under
conditioned: Segmentation of the vertebral column evident, or
dorsal pelvic bones are readily palpable) and profound lethargy, a
tumor affecting the gait, normal posture or ability to eat,
urinate, or defecate independently of the size of the tumor or the
determination by a University Laboratory Animal Resources
veterinarian that the animal should be euthanized. Animal death was
verified by pupil dilation and the absence of a heartbeat. The mice
were divided into 4 groups: i) The control group (CON); ii) the RT
group; iii) the 1MT group; and iv) the combined group (1MT + RT).
On the first day after tumor inoculation, 1MT was diluted to 400
mg/kg in saline and was administered by gavage twice daily. On day
13 after inoculation of the tumor, RT was given locally on the mice
in the RT group with a fractionation scheme of 10 Gy [the mice were
anesthetized with chloral hydrate (4% solution, 400 mg/kg) by
intraperitoneal injection before receiving radiation].
The mice were sacrificed on day 28
after inoculation
Tumors were recovered and weighed, and tumors and
spleens from each group were quickly prepared for flow cytometry.
Tumor-bearing mouse spleens were disaggregated with the flat end of
a syringe in 5 ml of RPMI 1640 medium in a tissue culture dish.
Dispersed cells were filtered through a 40-µm Falcon cell
strainer.
Flow cytometry analysis
Phenotypic analysis and characterization of DCs or T
cells was performed using a FACSCanto II flow cytometer (BD
Biosciences). Antibodies were purchased from eBioscience; Thermo
Fisher Scientific, Inc. Individual spleens were isolated and
pressed through a 40-µm Falcon Cell Strainer. Dissociated cells
were treated with Ammonium-Chloride-Potassium lysis buffer (Lonza
Group Ltd.) to lyse red blood cells. The resulting cell suspension
was then centrifuged for 5 min at 500 × g and 4°C, before removal
of the supernatant and re-suspension of the cells in PBS for
further FACS analysis. DC and T cell subsets were analyzed with
two- or three-color staining with various combinations of mouse
antibodies. DCs were stained with FITC-CD11C (cat. no. 11-0041-85),
phycoerythrin (PE)-CD80 (cat. no. 12-0081-82), PE-CD86 (cat. no.
12-0861-83) and PE-major histocompatibility complex (MHC) II (cat.
no. 1895-09) monoclonal antibodies. For T cells, FITC-CD4 (cat. no.
11-0041-82), allophycocyanin (APC)-CD25 (cat. no. 12-0251-83),
PE-FOXP3 (cat. no. 563101), PE-CD8 (cat. no. 12-0081-82) and
APC-IFN-γ (cat. no. 17-7311-82) conjugated anti-mouse monoclonal
antibodies were used for staining (3×105 cells in 150 µl
full medium with 0.2 µg antibodies; all antibodies and isotype
controls were diluted using this method). FOXP3 and IFN-γ
expression was assessed using intracellular staining with
Fixation/Permeabilization Concentrate and Fixation/Permeabilization
Diluent (eBioscience; Thermo Fisher Scientific, Inc.). Isotype
controls (mouse IgG2a K isotype control-PE, cat. no. 43912-60-100;
mouse IgG1 K isotype control-APC, cat. no. 44212-80-100; mouse
IgG2a K isotype control FITC, cat. no. 43912-50-100) were used to
discriminate positive cell staining from nonspecific background
staining.). Results were analyzed using FlowJo version 10.5.2
(FlowJo LLC) software.
Reverse transcription-quantitative
(RT-q) PCR
Total RNA was isolated using TRIzol®
(Invitrogen; Thermo Fisher Scientific, Inc.) and complementary
(c)DNA synthesis was performed using the Vazyme Reverse
Transcription System (Vazyme). HiScript II qRTSuperMix II was added
to RNA, which was reverse transcribed using the following
temperature protocol: 10 min at 25°C, 30 min at 42°C and 5 min at
85°C. The mRNAs of mouse IDO, PD-1, PD-L1, TIM-3, BTLA and
galectin-9 were determined using RT-qPCR. Using the RT reaction as
a template, SYBR-Green I dye (Vazyme) was used for qPCR. The
reaction conditions were 10 min at 95°C, followed by 15 sec at 95°C
and 1 min at 60°C for 40 cycles for IDO, or 1 min at 58°C for 40
cycles for PD-1, or 1 min at 63°C for 40 cycles for PD-L1, or 1 min
at 60°C for 40 cycles for TIM-3, BTLA and galectin-9. The primer
sequences for RT-qPCR are listed in Table I. The relative mRNA expression
levels of target genes were calculated using the 2−ΔΔCq
method (25) compared with GAPDH
expression.
 | Table I.Sequences of primers used for reverse
transcription-quantitative PCR. |
Table I.
Sequences of primers used for reverse
transcription-quantitative PCR.
| Gene | Primer | Product length |
|---|
| IDO | F
5′-GGGCTTTGCTCTACCACATCCACT-3′ | 234 bp |
|
| R
5′-ACATCGTCATCCCCTCGGTTCC-3′ |
|
| BTLA | F
5′-TGCAGGAGCCAGAAGAGAAAGTCA-3′ | 306 bp |
|
| R:
5′-CAATGTGGGGGTCAGGGATGG-3′ |
|
| GAL-9 | F
5′-GTTGTCCGAAACAACTCAGAT-3′ | 315 bp |
|
| R
5′-ATATGATCCACACCGAGAAG-3′ |
|
| PD-1 | F
5′-GGCCGCCTTCTGTAATGGTTTGA-3′ | 279 bp |
|
| R
5′-AGGGGCTGGGATATCTTGTTGAGG-3′ |
|
| PD-L1 | F
5′-GACCAGCTTTTGAAGGGAAATG-3′ | 385 bp |
|
| R
5′-CTGGTTGATTTTGCGGTATGG-3′ |
|
| TIM3 | F
5′-AGTGGGAGTCTCTGCTGGGTTGA-3′ | 279 bp |
|
| R
5′-AGGATGGCTGCTGGCTGTTGA-3′ |
|
| GAPDH | F
5′-TGATGACATCAAGAAGGTGGTGAA-3′ | 345 bp |
|
| R
5′-TGGGATGGAAATTGTGAGGGAGAT-3′ |
|
Statistical analysis
Experiments were repeated three times. Data are
expressed as the mean ± SD. Comparisons among groups were performed
using One-way ANOVA followed by Tukey's post hoc test. Data
analysis was performed using SPSS 17.0 software (SPSS, Inc.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
IDO inhibition combined with RT
downregulates IDO expression and suppresses tumor growth
To examine whether IDO inhibition affects tumor
growth, mice were divided into four groups and subjected to various
treatments. The longest diameter of a single subcutaneous tumor was
1.9 cm and no animal presented with multiple tumors. The growth
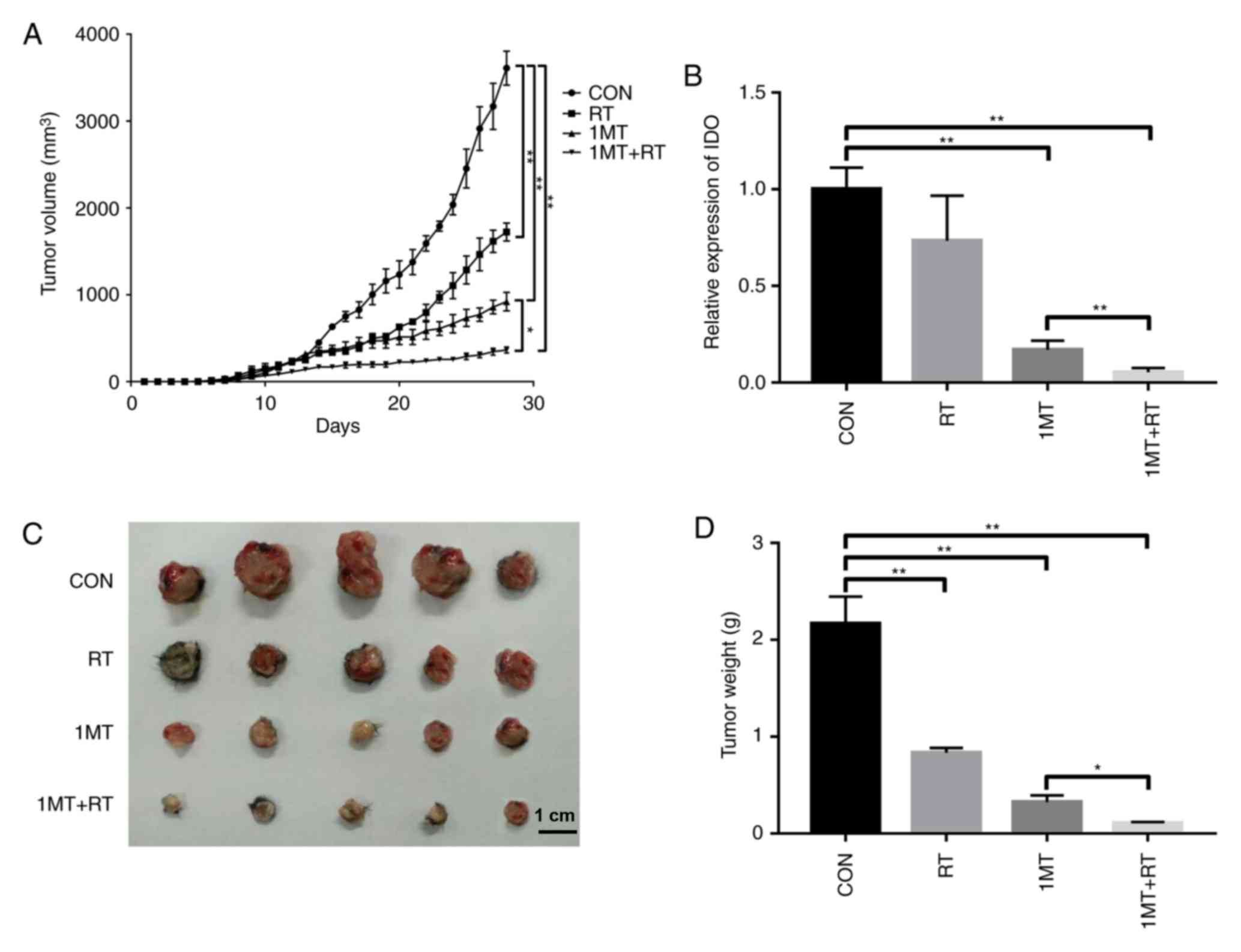
curve (Fig. 1A) suggested that
tumor progression was significantly slower in both the RT group and
the 1MT group compared with the CON group. Furthermore, 1MT
combined with RT had a larger therapeutic effect than either
treatment alone. RT-qPCR showed that the tumors in the 1MT and the
combined groups had lower IDO expression levels than those in the
CON group (Fig. 1B), suggesting
that differences in the therapeutic outcome may be due to
differences in IDO expression. On the 28th day after tumor
inoculation, the mice were euthanized using carbon dioxide
inhalation and the tumor tissue was weighed. The tumor weights and
data are shown in Fig. 1C and D.
As shown in Fig. 1D, the average
tumor weights were as follows: 2.165 g in the CON group, 0.834 g in
the RT group, 0.324 g in the 1MT group and 0.107 g in the combined
group.
The slower tumor progression in the 1MT-treated
group compared with the CON group suggested that IDO is
responsible, at least in part, for tumor immunosuppression and that
using the 1MT inhibitor can reverse this effect. The combined group
showed a significantly better therapeutic effect than the other
groups, indicating that 1MT and RT can act synergistically. Some of
the mechanisms driving this effect are discussed below.
IDO inhibition synergizes with RT to
promote the maturation of DCs and downregulate the inhibitory
receptor ligand PD-L1 in the tumor microenvironment
As shown in Fig. 1,
compared to the control condition, combined therapy delays tumor
progression. One potential explanation for tumor immune escape and
resistance is that IDO may be involved in DC immaturity and T cell
anergy, suggesting that the inhibition of IDO expression induced by
1MT may lead to the tumor suppressive effects shown in the Fig. 1. To investigate the influences of
changes in IDO on DCs, maturity markers, including CD80, CD86 and
MHC II, were examined. Based on flow cytometry analysis (Fig. 2A-C), the results indicated that
these co-stimulatory factors were highly expressed in the combined
treatment group, with CD80+ expression in 80.1% of
CD11c+ cells and CD86+ expression in 71.0% of
CD11c+ cells. Additionally, the DCs in the 1MT group and
RT group also showed a high degree of maturity, indicating that RT
and IDO inhibition increased the maturity of DCs, and that these
therapies can synergize to produce greater effects.
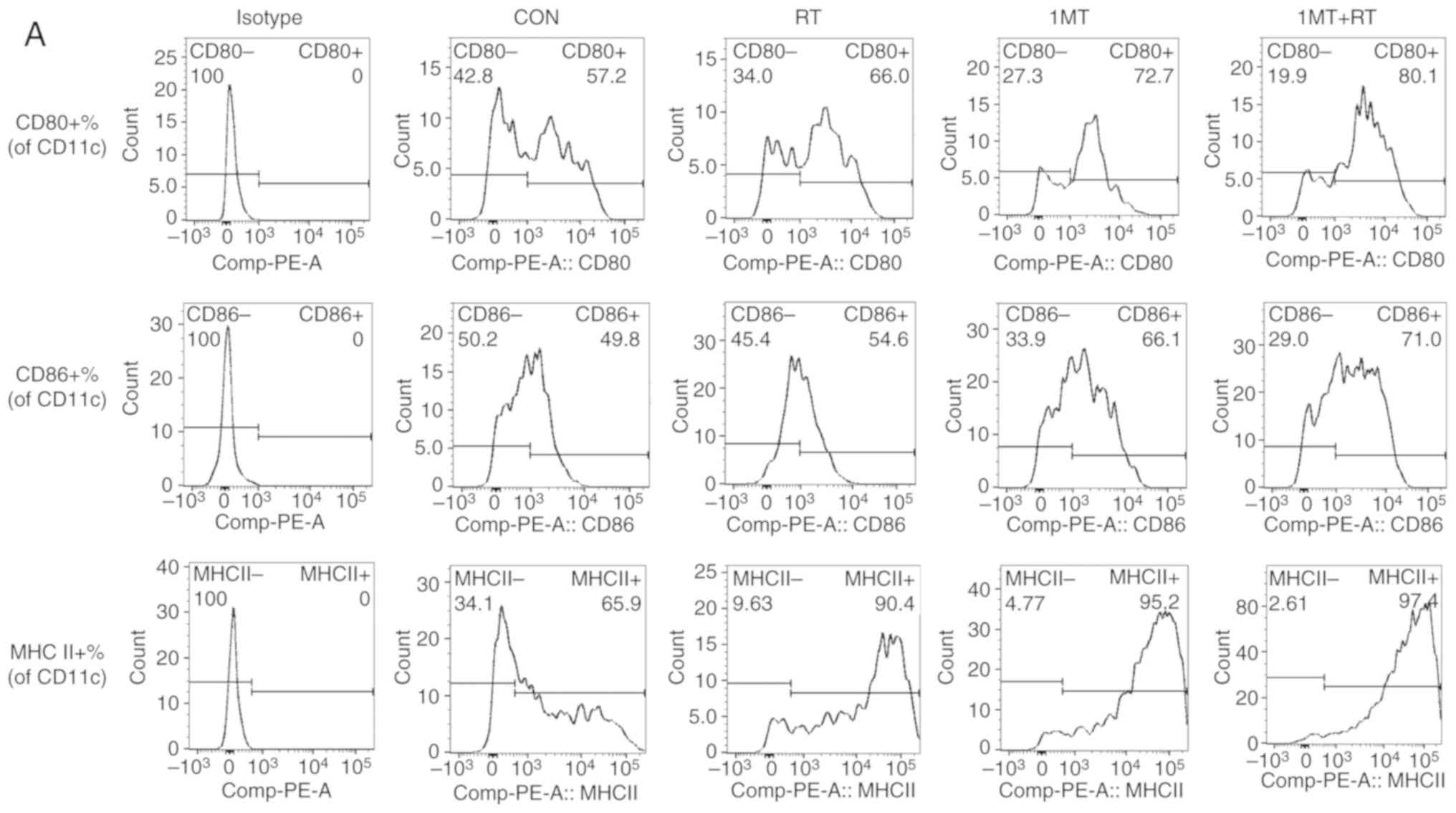 | Figure 2.IDO inhibition and RT in combination
promote the maturation of DCs and downregulate the inhibitory
receptor ligand PD-L1 in the tumor microenvironment. (A) The
results of flow cytometry. To validate the influences of changes in
IDO expression on DCs, maturity markers, including CD80, CD86 and
MHC II, were examined. Comparison of the maturation of DCs under
various treatments based on flow cytometry. RT and IDO inhibitor
treatment upregulated the expression of (B) CD80 and (C) CD86. The
combined therapy of 1MT+RT showed the strongest effect, which
significantly increased the expression of the cell surface
co-stimulatory factors CD80 and CD86 compared with the single
treatments. (D) Evaluation of the ability to initiate T cell
activation and provide a second stimulatory signal was performed by
measuring the expression level of MHC II on DCs. The results
indicated that RT and IDO inhibitor treatment significantly
upregulated the expression of MHC II compared with the CON group;
the combined therapy showed the strongest effect. (E) PD-L1 in the
tumor microenvironment, as an inhibitory receptor ligand expressed
on DCs, was measured. The results suggested that RT and IDO
inhibition significantly downregulated PD-L1 expression compared
with the CON group; the n=5, experiments were repeated three times.
combined therapy had the strongest effect. **P<0.001, *P<0.01
vs. CON. IDO, indoleamine 2,3-dioxygenase; RT, radiotherapy; 1MT,
1-methyl-tryptophan; CON, control; DCs, dendritic cells; MHC II,
major histocompatibility complex II; PD-L1, programmed cell death
ligand 1. |
Mature DCs must express high levels of MHC II and
downregulate co-inhibitory molecules to initiate T cell activation,
and provide a secondary stimulatory signal. Based on flow cytometry
analysis (Fig. 2A and D), DCs in
the RT, 1MT and combined groups showed high MHC II expression, with
the mean values of 89.27±1.71, 95.00±0.50 and 96.63±0.54% of
CD11c+ cells expressing MHC II, respectively. These
results indicated that RT and IDO inhibition upregulate the
expression of MHC II, which may increase the antigen-presenting
ability of DCs. These treatments also showed increased therapeutic
effects when combined compared with their use alone. RT-qPCR showed
that PD-L1 in the tumor microenvironment, as an inhibitory receptor
ligand, was significantly downregulated after RT and/or 1MT
treatment compared with the corresponding level in the CON group
(Fig. 2E), which reduces the
inhibitory effect of PD-L1 on T cells. These results indicated that
RT and IDO inhibition can lead to the progression of DC maturation,
enhancing their ability to activate T cells and to initiate the
immune response. The IDO inhibitor synergized with RT to produce an
increase in immune activation effects compared with the use of the
treatments individually.
IDO inhibition synergizes with RT to
reduce the expression of inhibitory receptors and Tregs, and
promote T cell activation
Through stimulation by antigen peptide-MHC II
complexes and co-stimulatory factors, DCs can activate T cells and
thus facilitate the clearance of antigens, and regulate the immune
response (26). Therefore, through
direct or indirect regulation of T cells, the IDO inhibitor can
active the immune response against a tumor. As shown in the
previous results, the IDO inhibitor can increase the maturity of
DCs and downregulate the expression of inhibitory receptor ligands,
which increases the ability of DCs to activate T cells. To
determine whether this phenomenon can enhance the activity of T
cells, IFN-γ, Tregs and inhibitory receptors were examined.
IFN-γ is a dimeric soluble cytokine with
immuno-activating and antitumor properties (27). Based on the results of flow
cytometry (Fig. 3A and B),
compared with the CON group, IDO inhibition increased the
activation and secretion of IFN-γ, suggesting that 1MT can enhance
the differentiation of T cells, which can result in an enhanced
antitumor ability. RT can also significantly upregulate the
secretion of IFN-γ compared with the CON group. The combined group,
which was treated with both RT and the IDO inhibitor, showed a
better therapeutic outcome than either individual treatment
group.
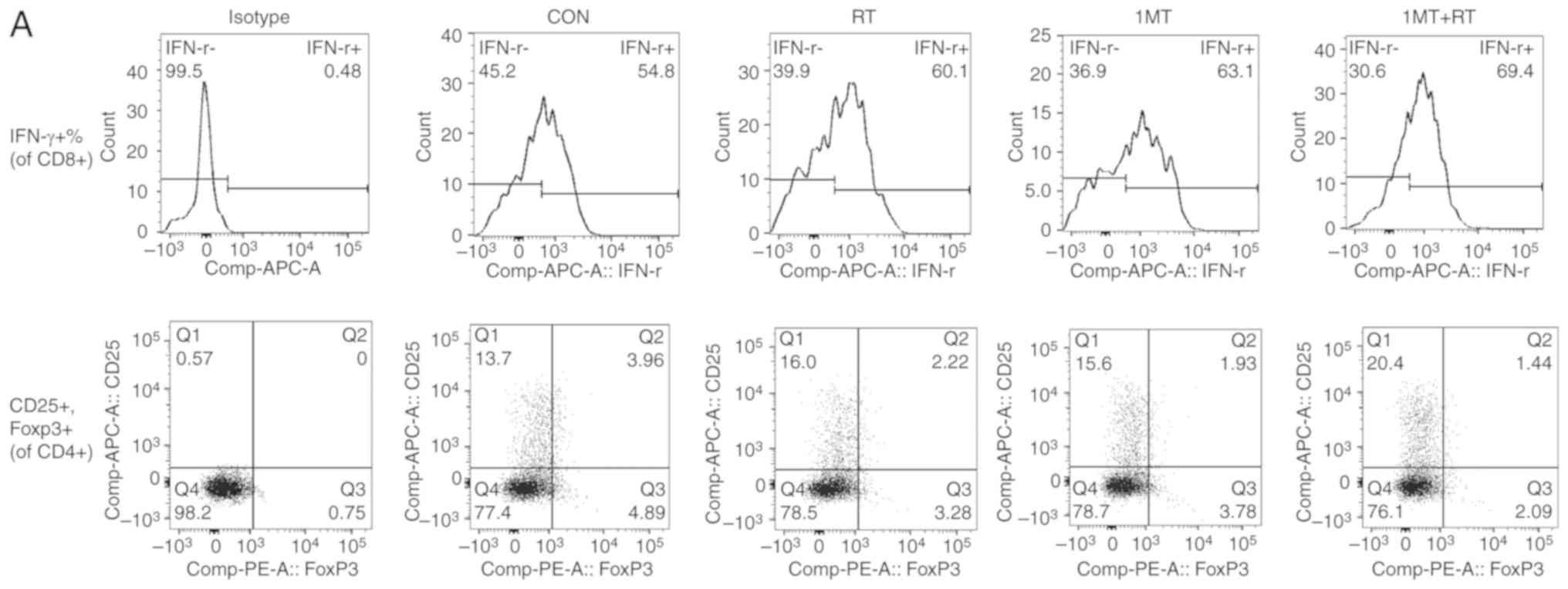 | Figure 3.IDO inhibition and RT in combination
reduce the expression of inhibitory receptors and Tregs, and
promote T cell activation. (A) The results of flow cytometry.
IFN-γ, a dimeric soluble cytokine that promotes the proliferation
and activation of cytotoxic lymphocytes, was examined to determine
whether the activity of T cells was enhanced following treatment.
CD4+CD25+FOXP3+ Tregs are also
measured to determine the inhibitory effect on T cells. (B) The
results showed that IDO inhibition synergized with RT to produce
the most significant effect among all the treatments. The combined
therapy of 1MT+RT had the stronger therapeutic effect. (C) The
percentage of CD4+CD25+FOXP3+
cells (CD25+FOXP3+ cells of CD4+
cells) was measured to evaluate the inhibitory effect on Tregs. The
results indicated that combined therapy showed the strongest
inhibitory effect on Tregs. PD-1, TIM-3 and BTLA are negative
regulators expressed on T cells and together lead to T cell anergy
and exhaustion. The levels of (D) PD-1, (E) TIM-3 and (F) BTLA were
measured, the results indicated that there was no significant
difference between the CON and RT groups, however, 1MT and 1MT
combined with RT significantly decreased the expression of these
markers compared with the CON group; the combined group showing the
strongest effect. (G) Galectin-9 is expressed on cancer cells and
is responsible for inhibition of T cell proliferation, and
apoptosis. The level of galectin-9 was measured and the results
showed no significant difference between the CON and RT groups,
however, 1MT significantly downregulated galectin-9 expression
compared with the CON group; IDO inhibition synergized with RT to
produce the stronger effect on Galecton-9 expression. n=5,
experiments were repeated three times. **P<0.001, *P<0.01 vs.
CON group. IDO, indoleamine 2,3-dioxygenase; RT, radiotherapy; 1MT,
1-methyl-tryptophan; CON, control; PD-1, programmed cell death 1;
TIM-3, T cell immunoglobulin domain and mucin domain 3; BTLA, B-
and T-lymphocyte attenuator; IFN-γ, interferon-γ; FOXP3, Forkhead
box P3; Tregs, regulatory T cells. |
Tregs, as a subpopulation of T cells, regulate the
immune system by suppressing T cell activation and proliferation
(28). Based on the results of
flow cytometry analysis (Fig. 3A and
C), compared with the CON group, IDO inhibition significantly
decreased the number of Tregs, suggesting that 1MT can relieve the
negative regulatory effect caused by Tregs and boost T cell
activity. Thus, IDO inhibition can enhance T cell function and
antitumor immunity. IDO inhibition synergized with RT to yield
better therapeutic effects than either therapy alone.
PD-1 is a protein receptor that regulates the immune
response by promoting self-tolerance and downregulating immunity
(29). Based on the RT-qPCR
analysis (Fig. 3D), no significant
difference in PD-1 expression was found between the RT and CON
groups, however, IDO inhibition significantly downregulated the
expression of PD-1, and when IDO inhibition synergized with RT, the
inhibitory effect was greater. This suggested that the IDO
inhibitor synergized with RT, and that this relieves the inhibitory
effects of PD-1 on T cells to a greater extent than either therapy
alone. Therefore, combined therapy can activate T cells and boost
antitumor immunity to a greater degree than either treatment
alone.
TIM-3 is an immune checkpoint protein that promotes
immune suppression together with other inhibitory molecules. TIM-3
is often upregulated in tumor-infiltrating lymphocytes, strategies
targeting TIM-3 can be used to boost T cell activity and activate
antitumor immunity (30). Based on
the RT-qPCR analysis (Fig. 3E),
compared with the CON group, RT alone had no effect on the
expression of TIM-3, however, IDO inhibition significantly
downregulated the expression of TIM-3, indicating that 1MT can
relieve the inhibitory effects of TIM-3 on T cells and prevent T
cell depletion. These results suggested that IDO inhibition boosted
antitumor immunity. The IDO inhibitor synergized with RT to yield a
better therapeutic effect than either therapy alone.
BTLA is a protein expressed on T helper (Th) 1 cells
during T cell activation and a negative regulatory ligand that
inhibits T cells (31). Based on
the RT-qPCR analysis (Fig. 3F), no
significant difference in BTLA expression was found between the RT
and CON groups, however, IDO inhibition decreased the expression
level of BTLA compared with the CON group, which may result in
enhanced T cell activity. Therefore, IDO inhibitors can enhance T
cell activity and antitumor immunity. IDO inhibition synergized
with RT and yielded better therapeutic effects than either
treatment alone.
Galectin-9 is a ligand expressed on the surface of
multiple types of cancer cells, and as the most studied ligand for
TIM-3. Galectin-9 can lead to T cell exhaustion by inhibiting T
cell proliferation and inducing apoptosis (32). Based on the RT-qPCR analysis
(Fig. 3G), RT alone had no effect
on the expression of galectin-9, however, IDO inhibition
significantly downregulated the expression of galectin-9 compared
with the CON group, indicating that 1MT relieves the inhibitory
effects of galectin-9 on T cells and boosts antitumor immunity. IDO
inhibition synergized with RT and yielded better therapeutic effect
than either treatment alone.
Discussion
The aim of the present study was to examine the
feasibility and efficacy of RT combined with IDO inhibition in
treating cancer.
The role of the inflammatory response in RT remains
difficult to predict. The efficacy of RT is closely related to the
immune status of the patient. RT can not only trigger innate and
adaptive immune responses, leading to tumor regression, but can
also cause tumor cells to develop a variety of resistance
mechanisms that promote tumor immune escape (33–35),
which is why RT can synergize with immunotherapy to produce greater
effects on the inhibition of tumor growth, IDO and negative
regulators. By analyzing the results of combination therapy, the
positive effects of IDO inhibition can be observed. IDO is an
intracellular enzyme that degrades the essential amino acid
L-tryptophan to N-kynurenine (36). IDO exerts immunosuppressive effects
by reducing the local concentration of tryptophan and increasing
the production of immunomodulatory tryptophan metabolites (37). Immunomodulatory tryptophan
metabolites have different effects on immune cells. For example,
the metabolites can inhibit proliferation and promote the apoptosis
of T lymphocytes, and can induce naive T cells to differentiate
into Tregs. IDO overexpression in DCs can affect cell maturation,
which results in decreased antigen presentation and the increased
expression of negative co-stimulatory molecules (36). To alleviate the inhibitory effect
of IDO on the immune system, the present study used an IDO
inhibitor. Treatment with 1MT has been shown to enhance the
function of cytotoxic lymphocytes in vitro following an
allogeneic mixed lymphocyte reaction by reducing the expression of
IDO (38). In addition,
significant reductions in the levels of the associated negative
regulators PD-1, TIM-3 and BTLA are also associated with the
effects of RT and IDO inhibitors (39–41).
PD-1 and its ligands, PD-L1 and PD-L2, are immunological checkpoint
proteins whose primary function is to limit inflammatory reactions
in peripheral tissues (42).
However, when these proteins are expressed in the tumor
microenvironment, this process represents an effective mechanism
for tumor-induced immunosuppression and evasion. TIM-3 is a member
of the TIM gene family, which includes TIM-1, TIM-3, and TIM-4 in
humans and TIM-1–8 in mice. TIM-3 is expressed on Th1, Th17 and
CD8+ T cells of the mouse bone marrow lineage (43). Binding between TIM-3 and its ligand
has been found to inhibit T cell responses and induce peripheral
immune tolerance. BTLA is an inhibitory molecule expressed by T
cells, B cells, DCs and natural killer cells. Herpes Virus Entering
Medium is a known ligand for BTLA. The cytoplasmic domain of BTLA
is required for BTLA to fully inhibit T cell proliferation and
cytokine production, including IFN-γ, IL-2 and IL-10. The reduction
of these cytokines may be related to the reduction of IDO. These
molecules have similar functions in promoting T cell activation and
DC-mediated activation of T cells, and enhancing immunity (44), which is consistent with the results
of the tumor growth curve in the present study showing that tumor
growth was significantly reduced.
The present study found that 1MT combined with RT
caused greater reductions in tumor growth, decreases in inhibitory
receptors and ligands, and activation of DCs and T cells in
vivo compared with either therapy alone. The killing activity
of the activated cytotoxic T lymphocytes may further indicate the
immune activation ability of this therapeutic strategy. In future
experiments, additional measurements will be made during in
vitro experiments, including measurement of the killing
activity of cytotoxic T lymphocytes.
Jiang and Chan (45) summarized the effects of RT and
immune checkpoint blockade, supporting the results of the present
study. Other in vitro experiments with tumor cells treated
with RT or IDO inhibitors also showed similar effects, which also
support the findings of the present study (46).
In conclusion, the present study demonstrated that
IDO inhibition combined with RT can significantly decrease the
expression level of IDO in tumor cells compared with the CON group.
Furthermore, DCs tend to be mature and show a greater
antigen-presenting ability following combination therapy.
Inhibitory receptors and ligands were downregulated in both DCs and
T cells following combined therapy compared with the CON group,
which partially overcomes the problem of T cell exhaustion.
Together, these changes facilitate the activation of antitumor
immunity. In addition, IDO inhibition and RT synergized to provide
a more effective inhibition of tumor growth than either therapy
alone.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from
National Natural Science Foundation of China (grant no. 81860504)
and the Key Research and Development Program of Jiangxi Province
(grant no. 20171BBG70120).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
HW, ML, ZL, WY, XZ, LW, JC, BM, RZ and WM were
involved in the conception and supervision of the project. ML and
ZL were involved in the design of the study. ML and ZL performed
the experiments, analyzed the results, and prepared the paper. All
the authors read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were performed in accordance
with the Animal Research Reporting of in vivo Experiments
and the National Institutes of Health guidelines for animal
welfare, and the study was approved by the Institutional Animal
Care and Use Committee of Nanchang University, China. All animal
experiments complied with the ARRIVE guidelines and the AVMA
euthanasia guidelines of 2013.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
O'Donnell JS, Smyth MJ and Teng MW:
Acquired resistance to anti-PD1 therapy: Checkmate to checkpoint
blockade? Genome Med. 8:1112016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Weinlich R, Oberst A, Beere HM and Green
DR: Necroptosis in development, inflammation and disease. Nat Rev
Mol Cell Biol. 18:127–136. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Postow MA, Callahan MK, Barker CA, Yamada
Y, Yuan J, Kitano S, Mu Z, Rasalan T, Adamow M, Ritter E, et al:
Immunologic correlates of the abscopal effect in a patient with
melanoma. N Engl J Med. 366:925–931. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Levy A, Chargari C, Marabelle A,
Perfettini JL, Magné N and Deutsch E: Can immunostimulatory agents
enhance the abscopal effect of radiotherapy? Eur J Cancer.
62:36–45. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Golden EB, Chhabra A, Chachoua A, Adams S,
Donach M, Fenton-Kerimian M, Friedman K, Ponzo F, Babb JS, Goldberg
J, et al: Local radiotherapy and granulocyte-macrophage
colony-stimulating factor to generate abscopal responses in
patients with metastatic solid tumours: A proof-of-principle trial.
Lancet Oncol. 16:795–803. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Seyedin SN, Tang C and Welsh JW: Author's
view: Radiation and immunotherapy as systemic therapy for solid
tumors. Oncoimmunology. 4:e9864022015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ungaro A, Orsi F, Casadio C, Galdy S,
Spada F, Cella CA, Tonno CD, Bonomo G, Vigna PD, Murgioni S, et al:
Successful palliative approach with high-intensity focused
ultrasound in a patient with metastatic anaplastic pancreatic
carcinoma: A case report. Ecancermedicalscience. 10:6352016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Espenel S, Vallard A, Rancoule C, Garcia
MA, Guy JB, Chargari C, Deutsch E and Magné N: Melanoma: Last call
for radiotherapy. Crit Rev Oncol Hematol. 110:13–19. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lu CS and Liu JH: Pneumonitis in cancer
patients receiving anti-PD-1 and radiotherapies: Three case
reports. Medicine (Baltimore). 96:e57472017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ye Q, Wang C, Xian J, Zhang M and Cao Y
and Cao Y: Expression of programmed cell death protein 1 (PD-1) and
indoleamine 2,3-dioxygenase (IDO) in the tumor microenvironment and
in tumor-draining lymph nodes of breast cancer. Hum Pathol.
75:81–90. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liang C, Peng L, Zeng S, Zhao Q, Tang L,
Jiang X, Zhang J, Yan N and Chen Y: Investigation of indoleamine
2,3-dioxygenase 1 expression in uveal melanoma. Exp Eye Res.
181:112–119. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Brahmer J, Reckamp KL, Baas P, Crinó L,
Eberhardt WE, Poddubskaya E, Antonia S, Pluzanski A, Vokes EE,
Holgado E, et al: Nivolumab versus docetaxel in advanced
squamous-cell non-small-cell lung cancer. N Engl J Med.
373:123–135. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Younes A, Brody J, Carpio C,
Lopez-Guillermo A, Ben-Yehuda D, Ferhanoglu B, Nagler A, Ozcan M,
Avivi I, Bosch F, et al: Safety and activity of ibrutinib in
combination with nivolumab in patients with relapsed non-Hodgkin
lymphoma or chronic lymphocytic leukaemia: A phase 1/2a study.
Lancet Haematol. 6:e67–e78. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Prendergast GC, Malachowski WJ, Mondal A,
Scherle P and Muller AJ: Indoleamine 2,3-dioxygenase and its
therapeutic inhibition in cancer. Int Rev Cell Mol Biol.
336:175–203. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kozuma Y, Takada K, Toyokawa G, Kohashi K,
Shimokawa M, Hirai F, Tagawa T, Okamoto T, Oda Y and Maehara Y:
Indoleamine 2,3-dioxygenase 1 and programmed cell death-ligand 1
co-expression correlates with aggressive features in lung
adenocarcinoma. Eur J Cancer. 101:20–29. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li A, Barsoumian HB, Schoenhals JE,
Cushman TR, Caetano MS, Wang X, Valdecanas DR, Niknam S, Younes AI,
Li G, et al: Indoleamine 2,3-dioxygenase 1 inhibition targets
anti-PD1-resistant lung tumors by blocking myeloid-derived
suppressor cells. Cancer Lett. 431:54–63. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tanizaki Y, Kobayashi A, Toujima S, Shiro
M, Mizoguchi M, Mabuchi Y, Yagi S, Minami S, Takikawa O and Ino K:
Indoleamine 2,3-dioxygenase promotes peritoneal metastasis of
ovarian cancer by inducing an immunosuppressive environment. Cancer
Sci. 105:966–973. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Moreno ACR, Porchia B, Pagni RL, Souza
PDC, Pegoraro R, Rodrigues KB, Barros TB, Aps LRMM, de Araújo EF,
Calich VLG and Ferreira LCS: The combined use of melatonin and an
indoleamine 2,3-dioxygenase-1 inhibitor enhances vaccine-induced
protective cellular immunity to HPV16-associated tumors. Front
Immunol. 9:19142018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Alahdal M, Xing Y, Tang T and Liang J:
1-Methyl-D-tryptophan reduces tumor CD133+ cells,
Wnt/β-catenin and NF-kappaβp65 while enhances lymphocytes NF-κβ2,
STAT3, and STAT4 pathways in murine pancreatic adenocarcinoma. Sci
Rep. 8:98692018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Spranger S, Spaapen RM, Zha Y, Williams J,
Meng Y, Ha TT and Gajewski TF: Up-regulation of PD-L1, IDO, and
T(regs) in the melanoma tumor microenvironment is driven by CD8(+)
T cells. Sci Transl Med. 5:200ra1162013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Williams DK, Markwalder JA, Balog AJ, Chen
B, Chen L, Donnell J, Haque L, Hart AC, Mandal SK, Nation A, et al:
Development of a series of novel o-phenylenediamine-based
indoleamine 2,3-dioxygenase 1 (IDO1) inhibitors. Bioorg Med Chem
Lett. 28:732–736. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vatner RE, Cooper BT, Vanpouille-Box C,
Demaria S and Formenti SC: Combinations of immunotherapy and
radiation in cancer therapy. Front Oncol. 4:3252014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jia H, Ren W, Feng Y, Wei T, Guo M, Guo J,
Zhao J, Song X, Wang M, Zhao T, et al: The enhanced antitumour
response of pimozide combined with the IDO inhibitor L-MT in
melanoma. Int J Oncol. 53:949–960. 2018.PubMed/NCBI
|
|
24
|
Leary S, Underwood W, Anthony R, Cartner
S, Corey D, Grandin T, Greenacre C, Gwaltney-Brant S, McCrackin MA,
Meyer R, et al: AVMA Guidelines for the Euthanasia of Animals: 2013
edition. University of Alaska Anchorage; 2013. 2013, PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Holling TM, Schooten E and van Den Elsen
PJ: Function and regulation of MHC class II molecules in
T-lymphocytes: Of mice and men. Hum Immunol. 65:282–290. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kursunel MA and Esendagli G: The untold
story of IFN-γ in cancer biology. Cytokine Growth Factor Rev.
31:73–81. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gondek DC, Lu LF, Quezada SA, Sakaguchi S
and Noelle RJ: Cutting edge: Contact-mediated suppression by
CD4+CD25+ regulatory cells involves a granzyme B-dependent,
perforin-independent mechanism. J Immunol. 174:1783–1786. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fife BT and Pauken KE: The role of the
PD-1 pathway in autoimmunity and peripheral tolerance. Ann N Y Acad
Sci. 1217:45–59. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Blackburn SD, Shin H, Haining WN, Zou T,
Workman CJ, Polley A, Betts MR, Freeman GJ, Vignali DA and Wherry
EJ: Coregulation of CD8+ T cell exhaustion by multiple inhibitory
receptors during chronic viral infection. Nat Immunol. 10:29–37.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Haymaker CL, Wu RC, Ritthipichai K,
Bernatchez C, Forget MA, Chen JQ, Liu H, Wang E, Marincola F, Hwu P
and Radvanyi LG: BTLA marks a less-differentiated
tumor-infiltrating lymphocyte subset in melanoma with enhanced
survival properties. Oncoimmunology. 4:e10142462015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sakuishi K, Apetoh L, Sullivan JM, Blazar
BR, Kuchroo VK and Anderson AC: Targeting Tim-3 and PD-1 pathways
to reverse T cell exhaustion and restore anti-tumor immunity. J Exp
Med. 207:2187–2194. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Vanpouille-Box C, Alard A, Aryankalayil
MJ, Sarfraz Y, Diamond JM, Schneider RJ, Inghirami G, Coleman CN,
Formenti SC and Demaria S: DNA exonuclease Trex1 regulates
radiotherapy-induced tumour immunogenicity. Nat Commun.
8:156182017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gong X, Li X, Jiang T, Xie H, Zhu Z, Zhou
F and Zhou C: Combined radiotherapy and Anti-PD-L1 antibody
synergistically enhances antitumor effect in non-small cell lung
cancer. J Thorac Oncol. 12:1085–1097. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Herter-Sprie GS, Koyama S, Korideck H, Hai
J, Deng J, Li YY, Buczkowski KA, Grant AK, Ullas S, Rhee K, et al:
Synergy of radiotherapy and PD-1 blockade in Kras-mutant lung
cancer. JCI Insight. 1:e874152016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Brown ZJ, Yu SJ, Heinrich B, Ma C, Fu Q,
Sandhu M, Agdashian D, Zhang Q, Korangy F and Greten TF:
Indoleamine 2,3-dioxygenase provides adaptive resistance to immune
checkpoint inhibitors in hepatocellular carcinoma. Cancer Immunol
Immunother. 67:1305–1315. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lim JY, Lee SE, Park G, Choi EY and Min
CK: Inhibition of indoleamine 2,3-dioxygenase by stereoisomers of
1-methyl tryptophan in an experimental graft-versus-tumor model.
Exp Hematol. 42:862–866.e3. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xiao B, Liu B, Song Y, Yu Z and Guo S:
Local cytotoxic T-lymphocyte-associated antigen-4 immunoglobulin
inhibition of rejection response is dependent on indoleamine
2,3-dioxygenase activities in the allograft. Transplant Proc.
46:3637–3640. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ostrand-Rosenberg S, Horn LA and
Ciavattone NG: Radiotherapy both promotes and inhibits
myeloid-derived suppressor cell function: Novel strategies for
preventing the tumor-protective effects of radiotherapy. Front
Oncol. 9:2152019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Oweida A, Hararah MK, Phan A, Binder D,
Bhatia S, Lennon S, Bukkapatnam S, Van Court B, Uyanga N, Darragh
L, et al: Resistance to radiotherapy and PD-L1 blockade is mediated
by TIM-3 upregulation and regulatory T-cell infiltration. Clin
Cancer Res. 24:5368–5380. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xu JF, Zhao K, Huang BJ, Xiong P, Fang M,
Shen GX and Gong FL: Clone and expression of murine BTLA
extracellular domain gene and its effect on the expression of B7 on
dendritic cells. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 22:413–416.
2006.(In Chinese). PubMed/NCBI
|
|
42
|
Shukuya T and Carbone DP: Predictive
markers for the efficacy of Anti-PD-1/PD-L1 antibodies in lung
cancer. J Thorac Oncol. 11:976–988. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Du W, Yang M, Turner A, Xu C, Ferris RL,
Huang J, Kane LP and Lu B: TIM-3 as a target for cancer
immunotherapy and mechanisms of action. Int J Mol Sci. 18(pii):
E6452017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ritthipichai K, Haymaker CL, Martinez M,
Aschenbrenner A, Yi X, Zhang M, Kale C, Vence LM, Roszik J,
Hailemichael Y, et al: Multifaceted role of BTLA in the control of
CD8+ T-cell Fate after antigen encounter. Clin Cancer
Res. 23:6151–6164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jiang W, Chan CK, Weissman IL, Kim BYS and
Hahn SM: Immune priming of the tumor microenvironment by radiation.
Trends Cancer. 2:638–645. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hirata E and Sahai E: Tumor
microenvironment and differential responses to therapy. Cold Spring
Harb Perspect Med. 7(pii): a0267812017. View Article : Google Scholar : PubMed/NCBI
|