Introduction
β-thalassemia is an inherited disease that affects
several organ systems, leading to a relatively short lifespan
(1,2). β-thalassemia is particularly
prevalent in the Mediterranean, Africa and Southeast Asia (3). Pathophysiologically, β-thalassemia is
characterized by insufficient hemoglobin β-chain expression and
excess α-chain accumulation, resulting in ineffective
erythropoiesis and chronic hemocytolysis (4). Patients with β-thalassemia are
dependent on frequent blood transfusions, which leave them at risk
of transfusion-transmitted infections and iron overload-related
tissue damage (5). Currently,
allogeneic bone marrow transplantation can cure the disease;
however, the limited availability of suitable bone marrow donors,
high cost and potential graft-versus-host responses limit its
clinical application (6). Gene
editing and therapy may be promising approaches for treatment of
β-thalassemia, but they still are at an early stage (6). Therefore, the development of novel
therapies for intervention of β-thalassemia is urgently
required.
Epidemiological surveys have demonstrated that high
levels of fetal hemoglobin (HbF, α2γ2) are inversely associated
with the severity of clinical symptoms in patients with
β-thalassemia (7). Therefore,
reactivation of γ-globin gene expression is a therapeutic strategy
for the treatment of β-thalassemia (8). There are three categories of HbF
inducers: Ribonucleotide reductase inhibitors, short-chain fatty
acids and cytotoxic agents (9).
Hydroxyurea (HU), a ribonucleotide reductase inhibitor, has been
approved by the United States Food and Drug Administration (FDA)
for the treatment of sickle cell disease (SCD) (7). However, its therapeutic efficacy in
β-thalassemia remains contentious (8,10,11).
Several compounds have been discovered to induce HbF or γ-globin
gene expression in human erythroleukemic K562 cells, erythroid
cells, primates and even in patients (12–15).
However, these substances either have a short half-life in
vivo, cytotoxicity, apparent carcinogenicity or induce
hematopoietic suppression (12).
Therefore, the discovery of new safe reagents for inducing HbF or
γ-globin gene expression may be of significance in the management
of patients with β-thalassemia.
Previous studies have reported that some natural
compounds from medical herbs can induce γ-globin gene expression
with little toxicity (16–19). Oridonin (ORI) is a tetracycline
diterpenoid from the traditional Chinese medicine Rabdosia
rubescens, which has anti-tumor, immunomodulatory and
neuroprotective activities (20–26).
Previous studies have demonstrated that induction of HbF expression
is associated with activation of p38 mitogen-activated protein
kinase (MAPK) (27–29) and activated p38 MAPK can further
activate cAMP response element binding 1 (CREB1), which can bind to
specific sites in the γ-globin gene promoters to induce its
expression (30,31). In addition, histone modification is
a known regulator of gene expression (32). Given that ORI can activate p38 MAPK
in some types of cancer cells (20,25),
it was hypothesized that ORI may also activate p38 MAPK and CREB1
to induce histone modification and γ-globin gene expression in
differentiating human erythroid precursor cells.
The present study purified and enriched human
erythroid precursor cells from patients with β-thalassemia.
Two-phase culture was employed to induce the differentiation of
erythroid precursor cells, and the effects of ORI on γ-globin
expression and the potential underlying mechanisms were
analyzed.
Materials and methods
Subjects and cell culture
A total of 12 patients with β-thalassemia were
recruited at the outpatient service of the Guangdong Women and
Children Hospital between January 2015 and January 2016. The
demographic and clinical characteristics of subjects are presented
in Table I. Patients with
β-thalassemia were diagnosed by laboratory blood tests and genetic
examination. Patients were excluded from the study if they had
received a blood transfusion during the past 3 months. Written
informed consent was obtained from the patients' guardians and the
experimental protocol was approved by the Ethical Committee of
Guangdong Women and Children Hospital.
 | Table I.Demographic and clinical
characteristics of subjects. |
Table I.
Demographic and clinical
characteristics of subjects.
| No. | Sex | Age (years) | Genotype | Subgroup |
|---|
| 1 | M | 7.2 |
CD41-42/IVS-II-654 | Major |
| 2 | F | 13.0 | CD41-42/CD17 | Major |
| 3 | F | 12.3 |
CD17/IVS-II-654 | Intermedia |
| 4 | M | 2.5 |
CD41-42/IVS-I-1 | Major |
| 5 | F | 8.0 |
CD41-42/CD41-42 | Major |
| 6 | F | 2.3 | IVS-II-654/-28 | Intermedia |
| 7 | F | 8.0 | IVS-II-654/-28 | Intermedia |
| 8 | F | 6.5 |
CD17/IVS-II-654 | Intermedia |
| 9 | M | 10.0 |
CD41-42/CD41-42 | Major |
| 10 | M | 3.5 | βE/-28 | Intermedia |
| 11 | M | 10.5 | −28/-28 | Intermedia |
| 12 | F | 6.6 | βE/IVS-II-654 | Intermedia |
Peripheral venous blood samples were collected from
the subjects. Blood mononuclear cells were isolated by
Ficoll-Hypaque density gradient (GE Healthcare) centrifugation for
20 min at 400 × g and room temperature, and CD34+ cells
were purified using immunomagnetic beads, as previously described
(33). The purified
CD34+ cells were subjected to two-phase liquid culture,
as previously described (34).
Briefly, CD34+ cells were cultured for 7 days (phase I
culture) in minimal essential media (MEM; Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum (FBS;
Biological Industries), 1 µg/ml cyclosporin A (Sandoz AG) and 10%
conditioned medium from cultures of bladder carcinoma 5,637 cells
(American Type Culture Collection) at 37°C in an atmosphere
containing 5% CO2 and 95% humidity. The non-adherent
cells were harvested, washed and cultured (phase II culture) for 7
days in fresh medium supplemented with 30% FBS, 10−5 M
β-mercaptoethanol, 10−6 M dexamethasone, 300 µg/ml
holo-transferrin (Sigma-Aldrich; Merck KGaA), 10 ng/ml human
recombinant stem cell factor and 1 U/ml human recombinant
erythropoietin (Ortho Pharmaceutical Corp.).
On day 6 of phase II culture, the cells were treated
in triplicate with vehicle (0.1% DMSO), ORI (Chengdu Must
Bio-Technology Co., Ltd.) at 0.1, 0.2, 0.5 or 1.0 µM, or 500 µM
sodium butyrate (NaB, a histone deacetylase inhibitor;
Sigma-Aldrich; Merck KGaA). The number of viable cells in the
different groups was counted by trypan blue dye exclusion assay
daily from day 6 to day 14 in a blinded manner. Some cells were
collected on day 12 and the relative mRNA expression levels of
γ-globin and protein expression levels of HbF were determined by
reverse transcription-quantitative PCR (RT-qPCR) and western blot
analysis, respectively. For analysis of p38 MAPK signaling, some
cells were pretreated with vehicle (control), 0.5 µM ORI for 12,
24, 48, 72 or 96 h, or 500 µM NaB for 48 h (as a positive control)
to analyze the time-course of p38 MAPK activation. Some cells were
pretreated with 10 µΜ SB203580 (a p38 MAPK inhibitor; Promega
Corporation) (35) for 30 min at
room temperature and treated with ORI (0.5 µM) or NaB (500 µM) for
48 h. To evaluate the levels of CREB1, cells were treated with
vehicle (control) or ORI (0.5 µM) for 24, 48, 72 or 96 h, or with
500 µM NaB for 72 h (as a positive control) to analyze CREB1
time-dependent activation. Some cells were pretreated with 10 µΜ
SB203580 for 30 min at room temperature and treated with ORI (0.5
µM) or NaB (500 µM) for 72 h. In addition, western blot analysis
and chromatin immunoprecipitation (ChIP) were performed in cells
treated with ORI (0.5 µM) and NaB (500 µM) for 72 h, alone or in
combination with SB203580 pre-treatment.
Trypan blue dye exclusion assay
The number of viable cells was determined by trypan
blue dye exclusion assay. Briefly, human erythroid progenitor cells
during phase II culture were treated with the indicated agents
between days 6 and 14, and cell samples were collected daily or
every other day. The cells were stained with 0.5% trypan blue at
room temperature for 2 min and the number of unstained viable cells
in the different groups was counted in a blinded manner.
RT-qPCR
Total RNA was extracted from different groups of
cells using the RNeasy kit (Qiagen, Inc.). Following qualification
and quantification, each RNA sample (1 µg) was reverse transcribed
into cDNA using oligo(dT)18 and Moloney murine leukemia
virus reverse transcriptase (Promega Corporation). The RT reactions
were performed at 42°C for 60 min, followed by 95°C for 5 min and
0–5°C for 5 min. The relative levels of target gene mRNA
transcripts normalized to control GAPDH were determined by RT-qPCR
using the SYBR Green PCR Master Mix (Thermo Fisher Scientific,
Inc.) and specific primers in a RT-PCR system (Applied Biosystems;
Thermo Fisher Scientific, Inc.). The primer sequences were as
follows: GAPDH, forward 5′-GCACCGTCAAGGCTGAGAAC-3′, reverse
5′-TGGTGAAGACGCCAGTGGA-3′; α-globin, forward
5′-TCCCCACCACCAAGACCTAC-3′, reverse 5′-CCTTAACCTGGGCAGAGCC-3′;
β-globin, forward 5′-CTCATGGCAAGAAAGTGCTCG-3′, reverse
5′-AATTCTTTGCCAAAGTGATGGG-3′; and γ-globin, forward
5′-GGCAACCTGTCCTCTGCCTC-3′ and reverse 5′-GAAATGGATTGCCAAAACGG-3′.
PCR was performed in triplicate at 95°C for 10 min, followed by 40
cycles at 95°C for 10 sec, 60°C for 20 sec and 72°C for 15 sec. PCR
was conducted using an ABI7500 PCR system (cat. no. 4351104;
Applied Biosystems; Thermo Fisher Scientific, Inc.). The data were
normalized to GAPDH and analyzed by 2−ΔΔCq (36).
Western blot analysis
The harvested cells were lysed on ice for 20 min in
50 mmol/l Tris-HCl (pH 8), 150 mmol/l NaCl, 2% Nonidet P-40, 0.5%
sodium deoxycholate, 0.02% sodium azide and 0.1% SDS supplemented
with 10 mmol/l PMSF. This was followed by centrifugation at 12,000
× g for 15 min at 4°C. Following determination of protein
concentrations using a Bradford assay (Bio-Rad Laboratories, Inc.),
the cell lysate samples (30–50 µg/lane) were separated by SDS-PAGE
on 15% gels and transferred to nitrocellulose membranes (EMD
Millipore). The membranes were blocked with 5% skimmed milk powder
in TBS buffer containing 0.1% Tween for 2 h at room temperature and
were incubated with the following primary antibodies: Sheep
polyclonal anti-HbF (cat. no. ab19364; 1:1,000; Abcam), rabbit
monoclonal anti-total (t)-p38 MAPK (cat. no. ab170099; 1:1,000;
Abcam), mouse monoclonal anti-phosphorylated (p)-p38 MAPK (cat. no.
ab45381; 1:1,000; Abcam), rabbit monoclonal anti-p-CREB1 (Ser 133;
cat. no. 9198; 1:1,000; Cell Signaling Technology, Inc.), rabbit
monoclonal anti-t-CREB1 (cat. no. 9197; 1:1,000; Cell Signaling
Technology, Inc.) and mouse monoclonal anti-β-actin (cat. no.
TA336770; 1:1,000; OriGene Technologies, Inc.) overnight at 4°C.
The bound antibodies were detected with horseradish
peroxidase-conjugated rabbit anti-sheep IgG (H+L) (cat. no.
14-23-06; 1:5,000; KPL, Inc.), goat anti-rabbit IgG (H+L) (cat. no.
ZB 2301; 1:5,000; OriGene Technologies, Inc.) and goat anti-mouse
IgG (H+L) (cat. no. ZB 2305; 1:5,000; OriGene Technologies, Inc.)
and visualized using an enhanced chemiluminescent kit (Merck &
Co., Inc.), followed by densitometric analysis in the Bio-BEST-140E
gel image system (SIM Company). The data were normalized to the
control β-actin and the relative levels of HbF, p38 MAPK and CREB1
expression and phosphorylation were determined using Glyko BandScan
(Version 5.0; Glyko, Inc.) in the Bio-BEST-140E gel image system
(SIM International).
ChIP
The impact of ORI on histone modification in the
promoter regions of the Gγ- and Aγ-globin genes was determined by
ChIP using the EZ-ChIP kit (cat. no. 17-371; EMD Millipore),
according to the manufacturer's protocol (Upstate Biotechnology,
Inc.; Thermo Fisher Scientific, Inc.). Briefly, 1×107
cells in the different groups were cross-linked with 1%
formaldehyde for 10 min at room temperature under gentle agitation;
cross-linking was terminated by the addition of 150 mM Glycine for
5 min. After the addition of SDS Lysis Buffer (cat. no. 20-163; EMD
Millipore) containing Protease Inhibitor Cocktail II (cat. no.
20-238; EMD Millipore), according to the manufacturer's protocol,
the cross-linked chromatins were sonicated five times at 100 W
power in an ice bath (10 sec/sonication with 30 sec between them),
in order to obtain DNA fragments averaging 200–1,000 bp in size.
One part of the supernatant fraction of chromatin without primary
antibody treatment was saved as the ‘input sample’. According to
the manufacturer's protocol, in order to minimize the risk of
contamination, the ‘input sample’ was divided into three samples.
These samples were incubated with the antibody of interest [input;
against acetyl-histone H3 (cat. no. 06-599; 1:100; EMD Millipore)
or H4 (cat. no. 06-866; 1:300; EMD Millipore)], anti-RNA polymerase
(positive control; 1:1,000; cat. no. 05-623; EMD Millipore) or
normal mouse IgG (negative control; 1:1,000; cat. no. 12-371; EMD
Millipore) overnight at 4°C with agitation. One tube without DNA
sample was referred to as the ‘No DNA’ PCR control. These purified
DNA samples and the ‘No DNA’ PCR control then underwent PCR using
Control Primers (included in the EZ-ChIP kit; cat. no. 22-004; EMD
Millipore), which are specific for the human GAPDH gene. The size
of the GAPDH PCR product was 166 bp. After verification, the
chromatins were probed with antibodies against acetyl-histone H3
(cat. no. 06-599; 1:100; EMD Millipore) and H4 (cat. no. 06-866;
1:300; EMD Millipore) overnight at 4°C with agitation. The relative
levels of the Gγ- and Aγ-globin promoter DNA fragments to the
control Necdin were determined by RT-qPCR. The primer sequences
were as follows: Necdin (control), forward
5′-GTCCTCTGCCTCTGCCATCA-3′, reverse 5′-ATACAGGGCACTGGCCACTC-3′;
Gγ-globin, forward 5′-GTCCTCTGCCTCTGCCATCA-3′, reverse
5′-ATACAGGGCACTGGCCACTC-3′; and Aγ-globin, forward
5′-TGTGGAAGATGCTGGAGGAG-3′ and reverse
5′-ATACACAGGGCACTGGCCACTG-3′. The PCR products were visualized on a
2% agarose gel with ethidium bromide staining (Thermo Fisher
Scientific, Inc.) and were semi-quantified; densitometric scanning
was conducted using the Bio-BEST-140E gel image system (SIM
International) and data were analyzed using Gel-Pro Analyzer 4.0
software (Media Cybernetics, Inc.).
Statistical analysis
Data are presented as the mean ± standard deviation.
The difference among groups was analyzed by one-way ANOVA, or
repeated ANOVA where appropriate, followed by the least significant
difference post hoc test using SPSS 13.0 (SPSS, Inc.). P<0.05
was considered to indicate a statistically significant difference.
When a P-value of 0.000 was achieved using the software it was
designated as P<0.001.
Results
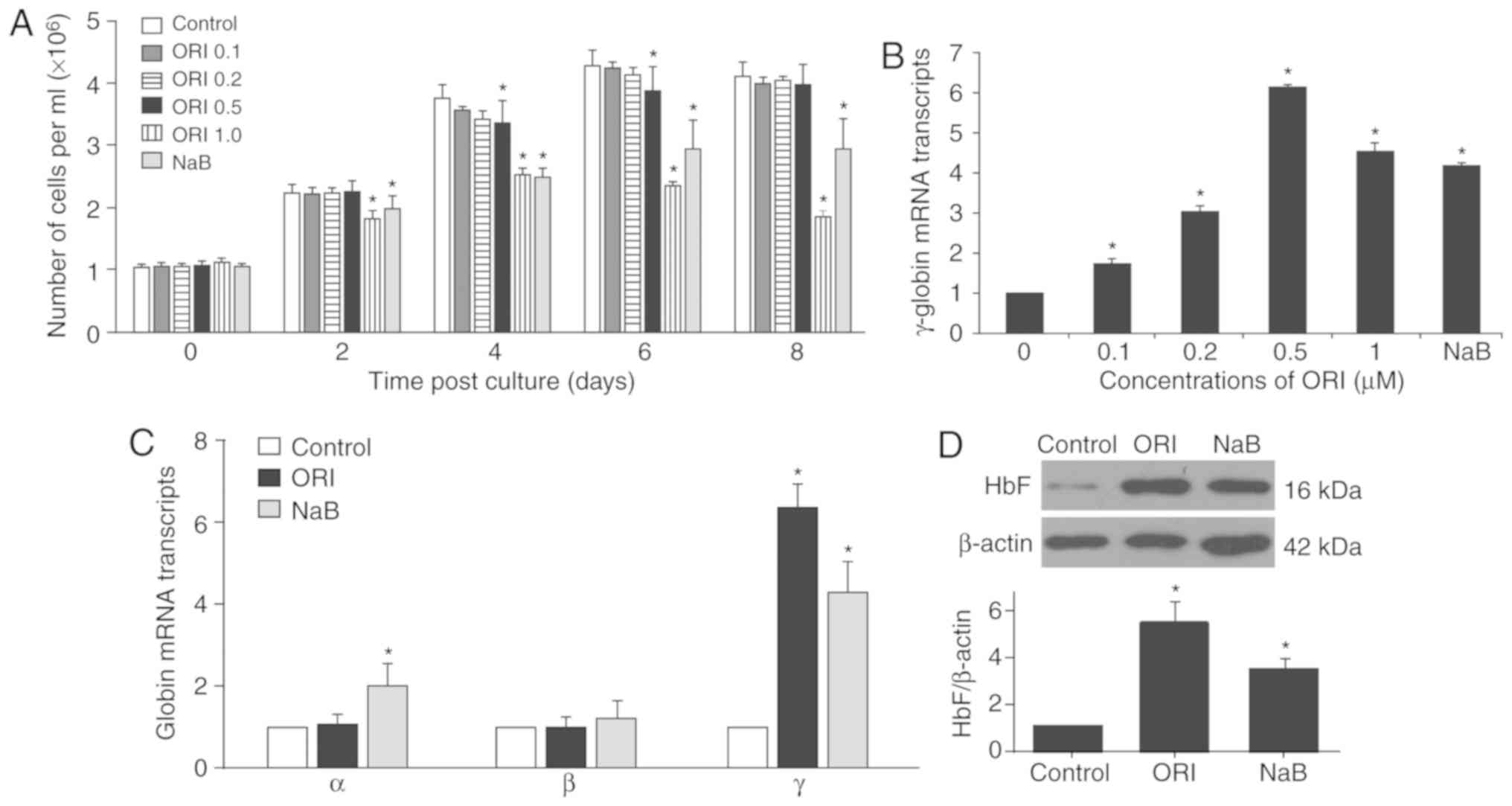
ORI preferably enhances γ-globin
expression during the maturation of erythroid precursor cells
To investigate the potential effect of ORI on the
expression of γ-globin, human CD34+ erythroid precursor
cells were purified from patients with β-thalassaemia and subjected
to two-phase culture. On day 6 of phase II culture, the cells were
treated with vehicle alone (control), different doses of ORI or 500
µM NaB (classic inducer of HbF expression, positive control).
Subsequently, the number of viably cultured cells was counted
longitudinally. As demonstrated in Fig. 1A, treatment with 0.5 µM ORI
temporarily reduced the number of cultured cells on day 4 (from
3.77±0.21×106 to 3.36±0.37×106 cells/ml) and
day 6 (from 4.30±0.24×106 to 3.89±0.39×106
cells/ml) post-culture, whereas treatment with 1 µM ORI, similar to
that of NaB, further decreased the number of cultured cells 2 days
after phase II culture. Cells were harvested on day 12, when the
cells matured into orthochromatic normoblasts and enucleated
erythrocytes (33). The relative
levels of γ-globin mRNA transcripts normalized to the control GAPDH
were determined by RT-qPCR (Fig.
1B). Treatment with different doses of ORI significantly
increased the relative levels of γ-globin mRNA transcripts, which
peaked following treatment with 0.5 µM ORI (6.18±0.04-fold;
P<0.05) and were slightly reduced following treatment with 1.0
µM ORI (4.57±0.20-fold; P<0.05) in orthochromatic normoblasts
and enucleated erythrocytes. Treatment with NaB also significantly
increased the levels of γ-globin mRNA transcripts (4.22±0.05-fold;
P<0.05); its effects were significantly stronger than those of
0.2 µM ORI (3.04±0.16-fold; P<0.05), but less than those of 1.0
µM ORI in erythroid precursor cells. These findings indicated that
ORI treatment enhanced γ-globin expression during the maturation of
erythroid precursor cells.
Given that treatment with 0.5 µM ORI strongly
enhanced γ-globin expression and exerted little cytotoxicity, this
dose was selected for further experiments. Further RT-qPCR analysis
revealed that treatment with ORI did not significantly affect the
relative mRNA expression levels of α-globin (1.10±0.20-fold;
P=0.419) or β-globin (1.00±0.24-fold; P=0.971), but did
significantly increase the relative mRNA expression levels of
γ-globin (6.42±0.57-fold; P<0.05) in the cultured orthochromatic
normoblasts and enucleated erythrocytes (Fig. 1C). However, treatment with NaB
significantly increased the relative mRNA expression levels of
α-globin (2.02±0.50-fold; P<0.05) and γ-globin (4.27±0.76-fold;
P<0.05), but not β-globin (1.22±0.43-fold; P=0.061) in the
cultured cells. Western blotting indicated that treatment with ORI
significantly increased the relative protein expression levels of
HbF by 5.55±0.84-fold (P<0.05), which was significantly higher
than the 3.54±0.40-fold increase in HbF induced by NaB in the
cultured cells (P=0.021; Fig. 1D).
Further western blot analysis indicated that treatment with NaB,
but not ORI, significantly increased the levels of α-globin
expression in the cultured cells (data not shown). Together, such
data demonstrated that ORI treatment preferably enhanced γ-globin
expression during the maturation of cultured erythroid precursor
cells in vitro.
ORI enhances γ-globin expression,
which is partially dependent on activation of p38 MAPK signaling in
erythroid precursor cells
Previous studies have demonstrated that induction of
HbF expression is associated with activation of p38 MAPK (26–29).
To understand the molecular mechanisms underlying the action of
ORI, erythroid precursor cells 1 day post-phase II culture were
treated with ORI for 4 days or NaB for 2 days, and the relative
levels of p38 MAPK expression and phosphorylation in the cells were
determined by western blotting. As demonstrated in Fig. 2A, 12 h post-treatment with ORI, the
relative ratios of p38 MAPK phosphorylation to its expression were
significantly increased (2.69±0.10; P<0.05). The relative ratios
of p38 MAPK phosphorylation to its expression peaked at 48 h
post-treatment (6.19±0.09; P<0.05) and declined gradually at
later time points. Similarly, treatment with NaB for 48 h
significantly increased the relative ratios of p-p38 MAPK to t-p38
MAPK in cultured cells (3.83±0.47; P<0.05). Conversely,
pretreatment with 10 µΜ SB203580, a p38-specific inhibitor,
significantly mitigated ORI- and NaB-induced p38 activation
(Fig. 2B) and reduced γ-globin
mRNA expression (Fig. 2C) and HbF
protein expression in cultured cells (Fig. 2D). The levels of p38
phosphorylation, γ-globin mRNA expression and HbF protein
expression in the SB203580-pretreated cells decreased to 34.82,
42.07 and 48.14% that of ORI-treated cells, respectively. Such data
indicated that ORI-enhanced γ-globin expression was particularly
dependent on activation of p38 MAPK in orthochromatic normoblasts
and enucleated erythrocytes.
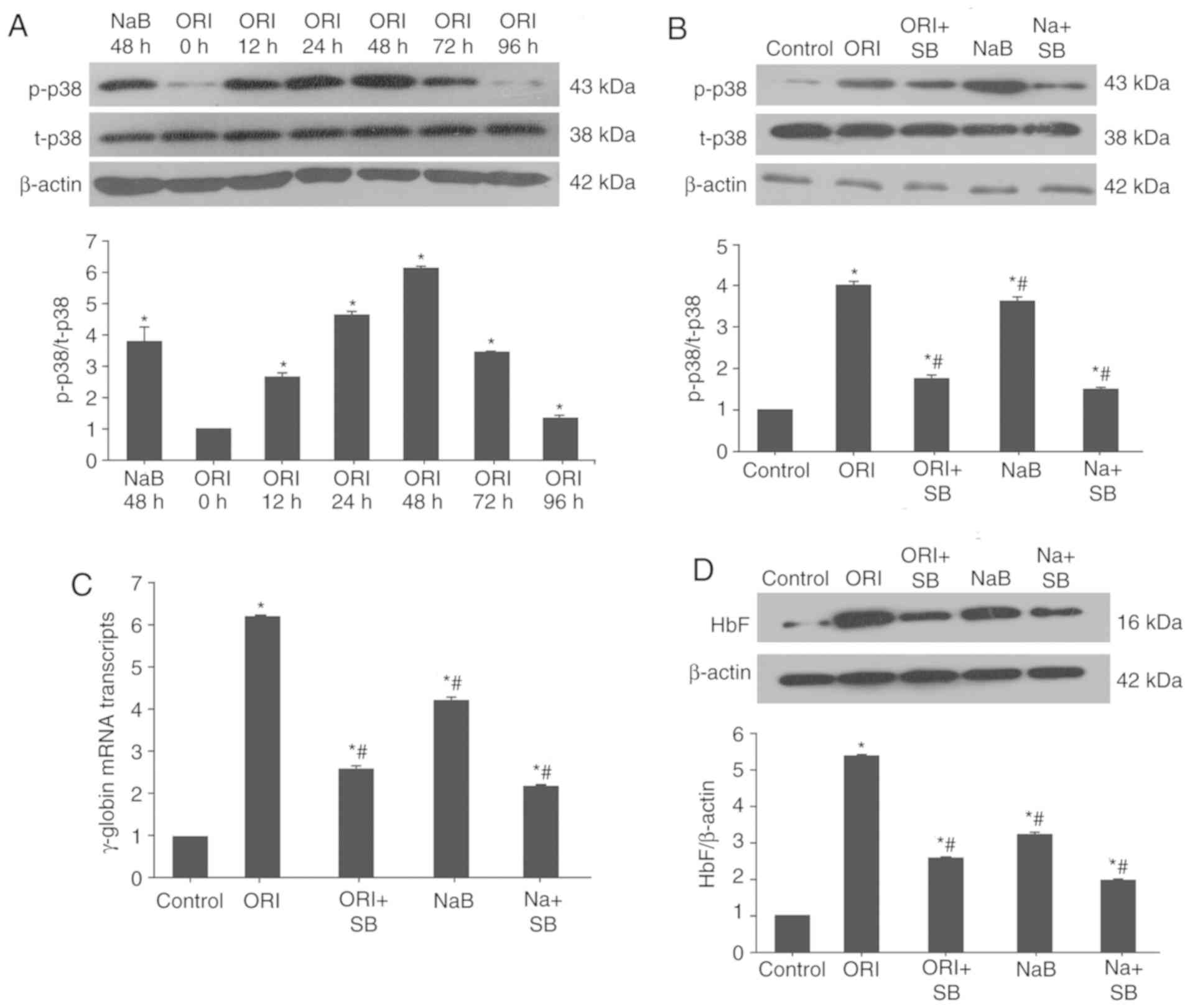 | Figure 2.ORI treatment activates p38 MAPK to
induce γ-globin expression in cultured erythroid precursor cells.
(A) On day 6 of the phase II culture, the cells were treated with
vehicle (control), 0.5 µM ORI for the indicated time periods or 500
µM NaB for 48 h. (B-D) Some cells were pretreated with, or without,
SB203580 for 30 min and treated with 0.5 µM ORI or 500 µM NaB for
48 h. The relative levels of p38 MAPK expression and
phosphorylation, γ-globin mRNA expression and HbF protein
expression were determined by western blot analysis and reverse
transcription-quantitative PCR. (B) ORI or NaB activated p38 MAPK,
which was mitigated by SB203580. (C) ORI induced γ-globin and (D)
HbF expression, which was partially dependent on p38 MAPK
activation. Data are representative images or expressed as the mean
± standard deviation of each group of cells from at least five
biological samples from three separate experiments. *P<0.05 vs.
the control group. #P<0.05 vs. 0.5 µM ORI alone.
MAPK, mitogen-activated protein kinase; NaB, sodium butyrate; ORI,
Oridonin; SB, SB203580. |
ORI treatment induces histone
modification in γ-globin gene promoter regions by enhancing p38
MAPK signaling in erythroid precursor cells
Histone modification is a regulator of gene
expression (31). To delineate the
role of p38 MAPK-dependent epigenetic modifications in ORI-induced
γ-globin gene expression, the cultured cells at 6 days post-phase
II culture were treated with ORI or NaB in the presence or absence
of SB23580 for 72 h. The different groups of cells were harvested
and cross-linked, and DNA fragments were prepared by sonication.
The DNA fragments were identified by ChIP, using
anti-acetyl-histone H3 and 4 for targeting and anti-RNA polymerase
as a control. Subsequently, the precipitated DNA fragments in the
Gγ- and Aγ-globin promoters were amplified by PCR. As demonstrated
in Fig. 3A, purified DNA was then
analyzed by PCR using Control Primers specific for the GAPDH
promoter. The PCR product was observed in the positive control
group (lane 3) and scarcely in the negative control group (lane 2).
GAPDH promoter-specific DNA was also observed in the Input (lane 4)
but not in the ‘No DNA’ PCR control (lane 1). Quantitative analysis
indicated that the levels of acetylated histone H3 (acH3) and
acetylated histone H4 (acH4) in the Gγ- and Aγ-globin promoter
regions in untreated cells were significantly higher than those of
Necdin, a hallmark of specificity (Fig. 3B). The levels of Gγ-acH3 and
Gγ-acH4 increased by 3.13±0.35- and 3.33±0.40-fold, respectively
(P<0.05), whereas Aγ-acH3 and Aγ-acH4 increased by 3.06±0.36-
and 3.28±0.41-fold, respectively (P<0.05; Fig. 3C and D). Conversely, the levels of
acH3 and acH4 at the Gγ- and Aγ-globin gene promoter regions in
SB203580-pretreated cells were decreased by 54.31 and 50.45% for
the Gγ-globin gene, and 51.93 and 47.88% for the Aγ-globin gene,
respectively, relative to cells treated with ORI alone. These
findings indicated that ORI-enhanced γ-globin expression was
partially mediated by p38 MAPK activation-dependent histone
acetylation in cultured orthochromatic normoblasts and enucleated
erythrocytes.
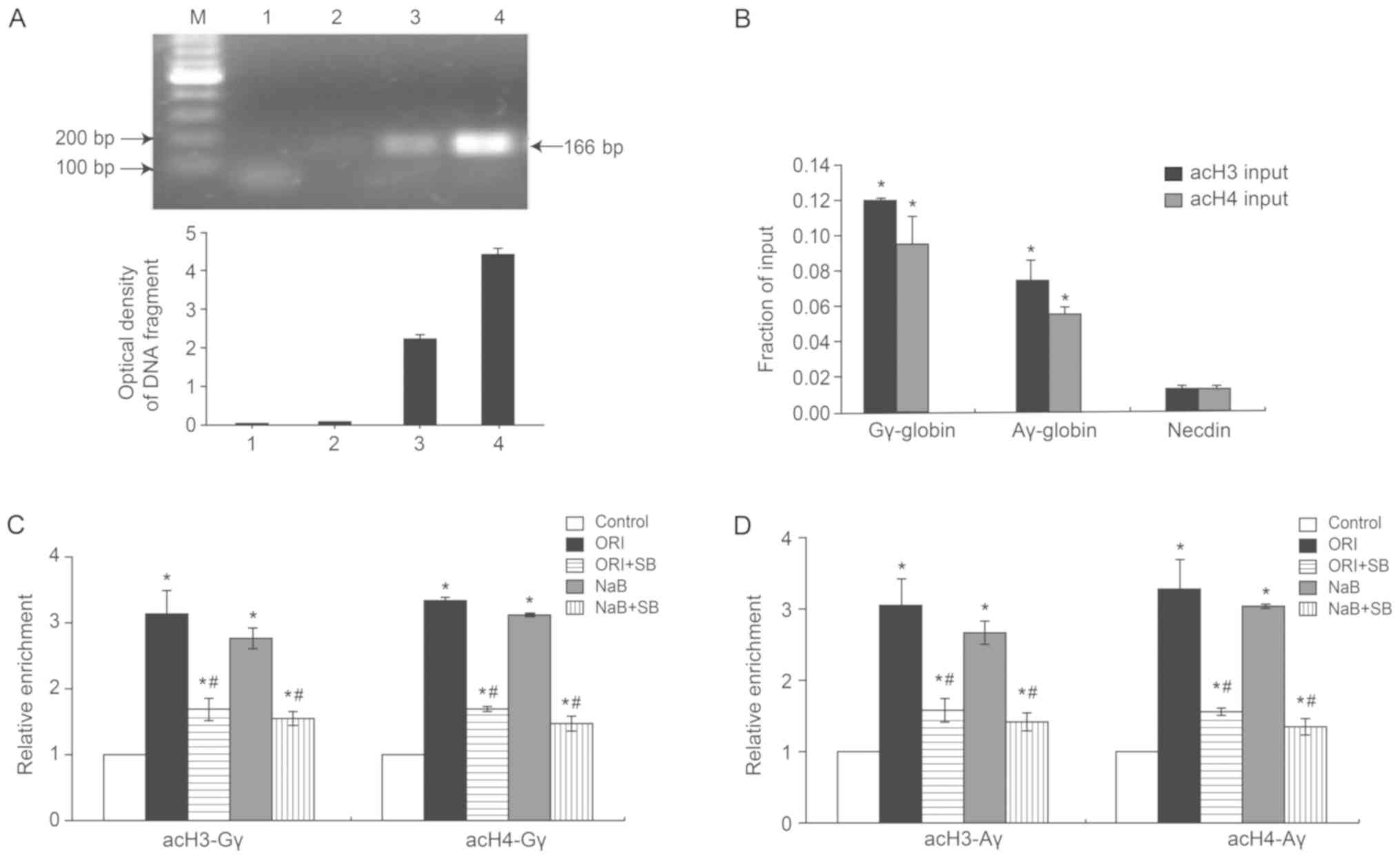 | Figure 3.ORI induces hyperacetylation in the
γ-globin promoter, which is partially dependent on p38 MAPK. On day
6 of the phase II culture, cells were pretreated with, or without,
SB203580 for 30 min and treated with 0.5 µM ORI or 500 µM NaB for
72 h. Their chromatins were prepared and subjected to ChIP using
acH3 and acH4 antibodies. The obtained DNA fragments were amplified
by PCR using specific primers for the Gγ-globin and Aγ-globin
promoters. Data are representative images or expressed as the mean
± standard deviation of each group of cells from at least five
biological samples from three separate experiments. (A)
Identification of PCR products by agarose gel electrophoresis. 1:
‘no DNA’ PCR control; 2: normal mouse IgG ChIP; 3: anti-RNA
polymerase II ChIP; 4: Input DNA. (B) Amplification of the
Gγ-globin and Aγ-globin and control in the input samples.
Amplification of the AcH3 and Ach4 in the (C) Gγ-globin and (D)
Aγ-globin promoters. *P<0.05 vs. the control group.
#P<0.05 vs. 0.5 µM ORI alone. acH3,
anti-acetyl-histone; acH4, anti-acetyl-histone H4; ChIP, chromatin
immunoprecipitation; MAPK, mitogen-activated protein kinase; NaB,
sodium butyrate; ORI, Oridonin; SB, SB203580. |
ORI induces CREB1 phosphorylation by
activating p38 MAPK signaling in erythroid precursor cells
Previous studies have demonstrated that activation
of p38 MAPK signaling by NaB augments γ-globin gene transcription
through CREB1 (29,30). The effects of ORI treatment on
CREB1 activation in the cultured orthochromatic normoblasts and
enucleated erythrocytes were analyzed by western blotting in this
study. Treatment with ORI significantly increased the relative
ratios of p-CREB1 to t-CREB1 in the cultured orthochromatic
normoblasts and enucleated erythrocytes at 24 h post-treatment
(2.13±0.07-fold; P<0.05); its effects peaked at 72 h
post-treatment (4.44±0.07-fold; P<0.05) and then markedly
declined to a level similar to the untreated control
(1.10±0.03-fold; P=0.12; Fig. 4A).
Similarly, treatment with NaB for 72 h significantly increased
CREB1 activation in the cultured cells (4.35±0.11-fold; P<0.05).
However, treatment with SB23580 significantly reduced ORI or
NaB-mediated CREB1 activation by 51.65 and 29.16% in the cultured
orthochromatic normoblasts and enucleated erythrocytes (Fig. 4B). These findings suggested that
ORI treatment activated p38 MAPK and CREB1, contributing to histone
modification, and enhanced γ-globin expression in the cultured
orthochromatic normoblasts and enucleated erythrocytes in
vitro.
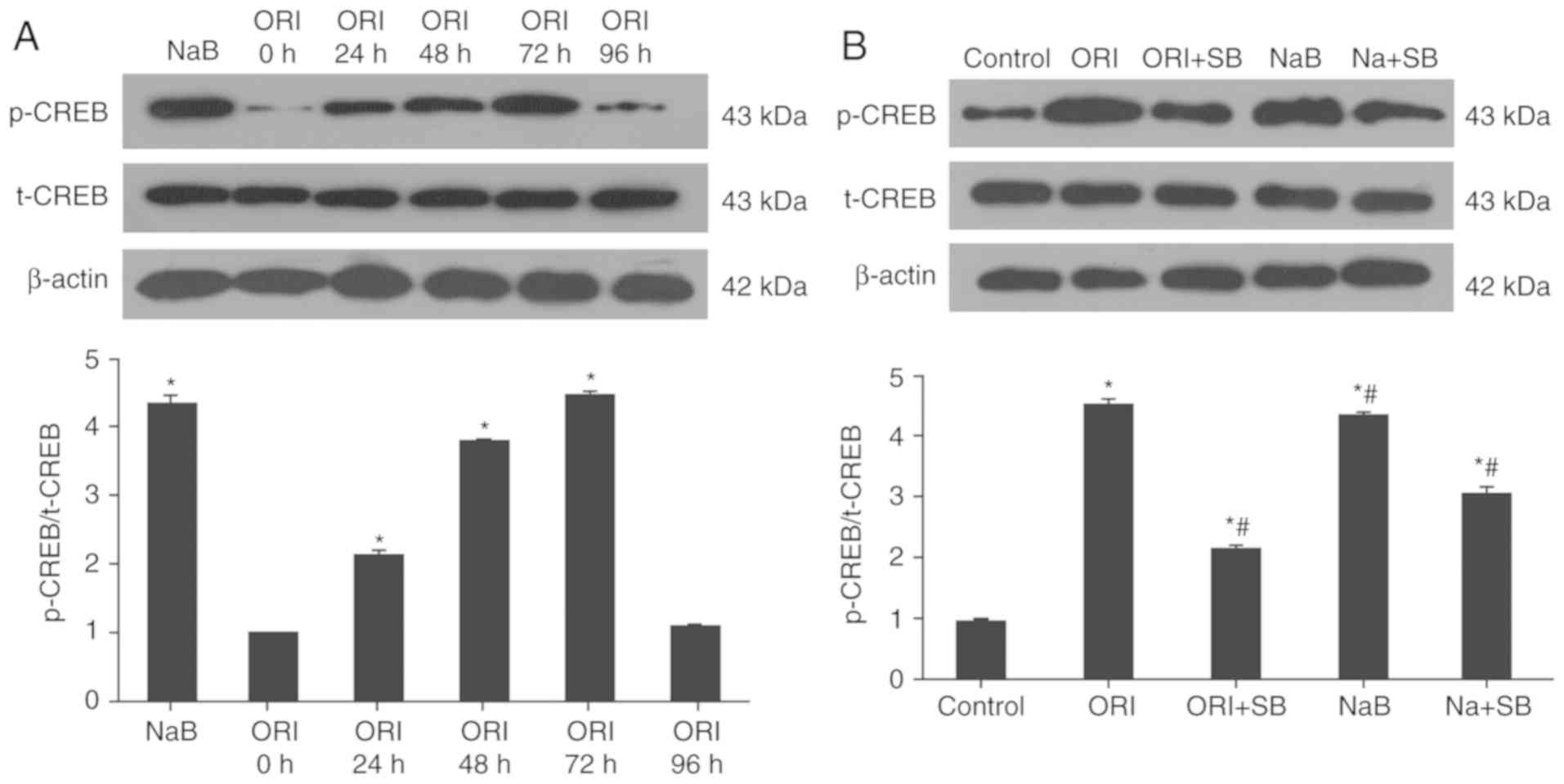 | Figure 4.ORI activates CREB1, which is
partially dependent on p38 MAPK activation. On day 6 of phase II
culture, cells were pretreated with, or without, SB203580 for 30
min and treated with 0.5 µM ORI for 96 h or 500 µM NaB for 72 h.
The relative levels of CREB1 phosphorylation and expression were
determined longitudinally or tested after treatment by western
blotting. Data are representative images or expressed as the mean ±
standard deviation of each group of cells from at least five
biological samples from three separate experiments. (A) ORI
activated CEB1 in a time-dependent manner. (B) ORI activated CREB1,
which was partially dependent on p38 MAPK activation at 72 h
post-treatment. *P<0.05 vs. the control group.
#P<0.05 vs. 0.5 µM ORI alone. CREB1, cAMP-response
element binding protein 1; MAPK, mitogen-activated protein kinase;
NaB, sodium butyrate; ORI, Oridonin; p-, phosphorylated; SB,
SB203580; t-, total. |
Discussion
Several natural products have been demonstrated to
induce HbF expression in vitro and in vivo (15–17,19);
however, the mechanisms underlying the action of these compounds
have yet to be elucidated. The present study revealed that
treatment with ORI significantly increased γ-globin expression with
little cytotoxicity in cultured orthochromatic normoblasts and
enucleated erythrocytes in vitro. In addition, treatment
with ORI activated p38 MAPK and CREB1 signaling, which led to
histone hyperacetylation and γ-globin expression in cultured
orthochromatic normoblasts and enucleated erythrocytes. To the best
of our knowledge, the present study was the first to demonstrate
that ORI effectively induced γ-globin expression in cultured
orthochromatic normoblasts and enucleated erythrocytes. This,
together with little cytotoxicity, suggests that ORI may be a
promising candidate for intervention in β-thalassemia.
An ideal HbF inducer should effectively induce HbF
production and have no cytotoxicity against the differentiated
erythrocytes (37). HU is a
ribonucleotide reductase inhibitor, which has been approved by the
FDA for treatment of SCD (7).
However, this treatment fails to achieve a significant increase in
the levels of HbF to prevent complications in patients with
β-thalassemia (38,39). The notable hematopoietic
suppression induced by HU has hindered the management of
β-thalassemia (12). The present
study demonstrated that treatment with ORI at 0.1–0.5 µM
significantly increased the expression levels of γ-globin in
cultured primary erythroid progenitor cells obtained from patients
with β-thalassemia in a dose-dependent manner. However, treatment
with 1 µM ORI decreased its effect on inducing γ-globin expression.
This, together with a reduction of ~50% in the number of viable
cells suggested that treatment with a higher dose of ORI may
trigger apoptosis of erythroid progenitor cells. In addition, ORI
treatment did not enhance α- and β-globin expression in cultured
primary erythroid progenitor cells, and this preference may prevent
the counteracting effect of some HbF inducers (15,40).
Notably, ORI at 0.5 µM exhibited little inhibition on the viability
of cultured primary erythroid progenitor cells. Rabdosia
rubescens, the herb from which ORI is extracted, has an
excellent safety profile (41–44).
Therefore, the present study suggested that ORI may be safe for
intervention in patients with β-thalassemia in the clinic.
Previous studies have demonstrated that activation
of the p38 MAPK signaling pathway is crucial for inducing γ-globin
expression (27–30,45,46).
MAPK kinase (MKK)3 and MKK6 are p38 MAPK activators, which can also
independently induce HbF production (44). The present study revealed that ORI
activated p38 MAPK signaling in a time-dependent manner, and
inhibition of p38 MAPK signaling mitigated or abrogated ORI-induced
increases in γ-globin expression in the cultured erythroid
progenitors. Such data indicated that ORI-induced γ-globin
expression partially depended on p38 MAPK activation in the
cultured erythroid progenitors. Previous studies have demonstrated
that some γ-globin inducers, such as HU, thalidomide, butyrate and
trichostatin A can promote reactive oxygen species (ROS) production
to activate p38 MAPK signaling, increasing γ-globin expression
(27,47,48).
ORI has been reported to induce the apoptosis of some cancer cells
by enhancing the ROS-mediated p38 MAPK signaling (49,50).
Accordingly, it is possible that ORI may enhance ROS production to
activate p38 MAPK and enhance γ-globin expression. More studies are
required to determine the precise mechanisms underlying the
pharmacological action of ORI.
It is notable that histone modification,
particularly hyperacetylation, is important for enhancing gene
expression. The present study demonstrated that ORI treatment
significantly increased the levels of acetyl H3 and H4 in the Aγ-
and Gγ-globin promoter regions and activated CREB1 in cultured
orthochromatic normoblasts and enucleated erythrocytes; these
effects were abrogated by the inhibition of p38 MAPK signaling. The
findings of the present study extended previous observations and
indicated that enhancement of histone acetylation or inhibition of
histone deacetylases can promote γ-globin expression by activating
p38 MAPK signaling (29,31,46).
It is notable that activated CREB1 is crucial for γ-globin
expression (29,30) and that the motif sequence for
activated CREB1 binding is located in the Gγ-globin promoter
(G-CRE, 5′-TGACGTCA-3′, −1,222 to −1,229) (35,40).
Accordingly, activation of CREB1 by ORI suggests that ORI may
activate p38 MAPK, which subsequently activates CREB1 to induce
histone hyperacetylation, increasing γ-globin expression in
cultured orthochromatic normoblasts and enucleated erythrocytes.
Therefore, the findings of the present study may provide novel
insights into regulation of the γ-globin expression in human
orthochromatic normoblasts and enucleated erythrocytes. Further
studies may investigate whether treatment with ORI can direct or
indirectly increase the interaction of CREB1 with the G-CRE in the
Gγ-globin promoter.
In summary, the results of the present study
indicated that ORI treatment selectively induced γ-globin
expression with little cytotoxicity. ORI treatment induced
epigenetic histone modification at the γ-globin promoter by
activating p38 MAPK and CREB1. Therefore, ORI may be considered a
promising therapeutic agent for intervention of β-thalassemia.
Notably, the present study had limitations, including a small
sample size, and a lack of in vivo studies and morphological
investigation; in particular, morphological analysis was not
conducted to confirm that cells developed into orthochromatic
normoblasts and enucleated erythrocytes on day 12, and this study
did not assess whether the addition of ORI arrested erythrocyte
maturation. Although the present study provided data to indicate
the therapeutic potentials of ORI, further studies with a larger
population are warranted to validate the findings and explore the
mechanisms underlying the pharmacological action of ORI in
enhancing γ-globin expression.
Acknowledgements
Not applicable.
Funding
This work was supported by grants from the Medical
Scientific Research Foundation of Guangdong Province of China
(grant no. 2019085) and Presidential Foundation of the Nanfang
Hospital, Southern Medical University (grant no. 2014C013).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LSG conceived and designed the study and wrote the
manuscript. JC performed the experiments, collected the data and
wrote the manuscript. QYW, JLZ and WMH collected and analyzed the
data. All of the authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Written informed consent was obtained from the
patients' guardians. The experimental protocol was approved by the
Ethical Committee of Guangdong Maternal and Child Health
Hospital.
Patient consent for publication
The patients' guardians provided consent for
publication.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Musallam KM, Cappellini MD, Wood JC, Motta
I, Graziadei G, Tamim H and Taher AT: Elevated liver iron
concentration is a marker of increased morbidity in patients with β
thalassemia intermedia. Haematologica. 96:1605–1612. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
La Nasa G, Argiolu F, Giardini C, Pession
A, Fagioli F, Caocci G, Vacca A, De Stefano P, Piras E, Ledda A, et
al: Unrelated bone marrow transplantation for beta-thalassemia
patients: The experience of the Italian Bone Marrow Transplant
Group. Ann N Y Acad Sci. 1054:186–195. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Galanello R and Origa R: Beta-thalassemia.
Orphanet J Rare Dis. 5:112010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yuan J, Bunyaratvej A, Fucharoen S, Fung
C, Shinar E and Schrier SL: The instability of the membrane
skeleton in thalassemic red blood cells. Blood. 86:3945–3950. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Berger K, Ajani UA, Kase CS, Gaziano JM,
Buring JE, Glynn RJ and Hennekens CH: Light-to-moderate alcohol
consumption and the risk of stroke among U.S. male physicians. N
Engl J Med. 341:1557–1564. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rachmilewitz EA and Giardina PJ: How I
treat thalassemia. Blood. 118:3479–3488. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ley TJ: The pharmacology of hemoglobin
switching: Of mice and men. Blood. 77:1146–1152. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lettre G and Bauer DE: Fetal haemoglobin
in sickle-cell disease: From genetic epidemiology to new
therapeutic strategies. Lancet. 387:2554–2564. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Atweh G and Fathallah H: Pharmacologic
induction of fetal hemoglobin production. Hematol Oncol Clin North
Am. 24:1131–1144. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Banan M: Hydroxyurea treatment in
β-thalassemia patients: To respond or not to respond? Ann Hematol.
92:289–299. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ronchi A and Ottolenghi S: To respond or
not to respond to hydroxyurea in thalassemia: A matter of stress
adaptation? Haematologica. 98:657–659. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Paikari A and Sheehan VA: Fetal
haemoglobin induction in sickle cell disease. Br J Haematol.
180:189–200. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Inati A, Kahale M, Perrine SP, Chui DH,
Taher AT, Koussa S, Abi Nasr T, Abbas HA and Ghalie RG: A phase 2
study of HQK-1001, an oral fetal haemoglobin inducer, in
β-thalassaemia intermedia. Br J Haematol. 164:456–458. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ronzoni L, Sonzogni L, Fossati G, Modena
D, Trombetta E, Porretti L and Cappellini MD: Modulation of gamma
globin genes expression by histone deacetylase inhibitors: An in
vitro study. Br J Haematol. 165:714–721. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Musallam KM, Taher AT, Cappellini MD and
Sankaran VG: Clinical experience with fetal hemoglobin induction
therapy in patients with β-thalassemia. Blood. 121:2199–2212. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bianchi N, Zuccato C, Lampronti I,
Borgatti M and Gambari R: Fetal Hemoglobin Inducers from the
Natural World: A novel approach for identification of drugs for the
treatment of {beta}-thalassemia and sickle-cell Anemia. Evid Based
Complement Alternat Med. 6:141–151. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fibach E, Bianchi N, Borgatti M, Prus E
and Gambari R: Mithramycin induces fetal hemoglobin production in
normal and thalassemic human erythroid precursor cells. Blood.
102:1276–1281. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ng NY and Ko CH: Natural remedies for the
treatment of beta-thalassemia and sickle cell Anemia-current status
and perspectives in fetal hemoglobin reactivation. Int Sch Res
Notices. 2014:1232572014.PubMed/NCBI
|
|
19
|
Theodorou A, Phylactides M, Forti L,
Cramarossa MR, Spyrou P, Gambari R, Thein SL and Kleanthous M: The
investigation of resveratrol and analogs as potential inducers of
fetal hemoglobin. Blood Cells Mol Dis. 58:6–12. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bu HQ, Liu DL, Wei WT, Chen L, Huang H, Li
Y and Cui JH: Oridonin induces apoptosis in SW1990 pancreatic
cancer cells via p53- and caspase-dependent induction of p38 MAPK.
Oncol Rep. 31:975–982. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Owona BA and Schluesener HJ: Molecular
insight in the multifunctional effects of oridonin. Drugs R D.
15:233–244. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang S, Zhang Y, Saas P, Wang H, Xu Y,
Chen K, Zhong J, Yuan Y, Wang Y and Sun Y: Oridonin's therapeutic
effect: Suppressing Th1/Th17 simultaneously in a mouse model of
Crohn's disease. J Gastroenterol Hepatol. 30:504–512. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang X, Chen LX, Ouyang L, Cheng Y and
Liu B: Plant natural compounds: Targeting pathways of autophagy as
anti-cancer therapeutic agents. Cell Prolif. 45:466–476. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhao G, Zhang T, Ma X, Jiang K, Wu H, Qiu
C, Guo M and Deng G: Oridonin attenuates the release of
pro-inflammatory cytokines in lipopolysaccharide-induced RAW264.7
cells and acute lung injury. Oncotarget. 8:68153–68164.
2017.PubMed/NCBI
|
|
25
|
Wu QX, Yuan SX, Ren CM, Yu Y, Sun WJ, He
BC and Wu K: Oridonin upregulates PTEN through activating p38 MAPK
and inhibits proliferation in human colon cancer cells. Oncol Rep.
35:3341–3348. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang S, Yu L, Yang H, Li C, Hui Z, Xu Y
and Zhu X: Oridonin attenuates synaptic loss and cognitive deficits
in an Aβ1-42-induced mouse model of Alzheimer's disease. PLoS One.
11:e01513972016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Aerbajinai W, Zhu J, Gao Z, Chin K and
Rodgers GP: Thalidomide induces gamma-globin gene expression
through increased reactive oxygen species-mediated p38 MAPK
signaling and histone H4 acetylation in adult erythropoiesis.
Blood. 110:2864–2871. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pace BS, Liu L, Li B and Makala LH: Cell
signaling pathways involved in drug-mediated fetal hemoglobin
induction: Strategies to treat sickle cell disease. Exp Biol Med
(Maywood). 240:1050–1064. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wei GH, Zhao GW, Song W, Hao DL, Lv X, Liu
DP and Liang CC: Mechanisms of human gamma-globin transcriptional
induction by apicidin involves p38 signaling to chromatin. Biochem
Biophys Res Commun. 363:889–894. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ramakrishnan V and Pace BS: Regulation of
gamma-globin gene expression involves signaling through the p38
MAPK/CREB1 pathway. Blood Cells Mol Dis. 47:12–22. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sangerman J, Lee MS, Yao X, Oteng E, Hsiao
CH, Li W, Zein S, Ofori-Acquah SF and Pace BS: Mechanism for fetal
hemoglobin induction by histone deacetylase inhibitors involves
gamma-globin activation by CREB1 and ATF-2. Blood. 108:3590–3599.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Marx V: Epigenetics: Reading the second
genomic code. Nature. 491:143–147. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Traycoff CM, Abboud MR, Laver J, Brandt
JE, Hoffman R, Law P, Ishizawa L and Srour EF: Evaluation of the in
vitro behavior of phenotypically defined populations of umbilical
cord blood hematopoietic progenitor cells. Exp Hematol. 22:215–222.
1994.PubMed/NCBI
|
|
34
|
Fibach E and Prus E: Differentiation of
human erythroid cells in culture. Curr Protoc Immunol. Chapter 22:
Unit 22F.7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cuenda A, Rouse J, Doza YN, Meier R, Cohen
P, Gallagher TF, Young PR and Lee JC: SB 203580 is a specific
inhibitor of a MAP kinase homologue which is stimulated by cellular
stresses and interleukin-1. FEBS Lett. 364:229–233. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Thein SL: The emerging role of fetal
hemoglobin induction in non-transfusion-dependent thalassemia.
Blood Rev. 26 (Suppl 1):S35–S39. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pourfarzad F, von Lindern M, Azarkeivan A,
Hou J, Kia SK, Esteghamat F, van Ijcken W, Philipsen S, Najmabadi H
and Grosveld F: Hydroxyurea responsiveness in β-thalassemic
patients is determined by the stress response adaptation of
erythroid progenitors and their differentiation propensity.
Haematologica. 98:696–704. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rigano P, Pecoraro A, Calzolari R, Troia
A, Acuto S, Renda D, Pantalone GR, Maggio A and Di Marzo R:
Desensitization to hydroxycarbamide following long-term treatment
of thalassaemia intermedia as observed in vivo and in primary
erythroid cultures from treated patients. Br J Haematol.
151:509–515. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fathallah H, Taher A, Bazarbachi A and
Atweh GF: Differences in response to fetal hemoglobin induction
therapy in beta-thalassemia and sickle cell disease. Blood Cells
Mol Dis. 43:58–62. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ma Z: Study on immune regulation of
Rabdosia rubescens and its safety assessment (unpublished PhD
thesis). Central South University, Public Health. 2010.(In
Chinese).
|
|
42
|
Zang K, Du L, Liu X, Ma J and Ren Y:
Therapeutic effect of oridonin tablet on acetic acid-induced
ulcerative colitis in mice. Chin J Modern Applied Pharmacy.
29:781–785. 2012.(In Chinese).
|
|
43
|
Ma Z, Hu C and Zhang Y: Therapeutic effect
of Rabdosia rubescens aqueous extract on chronic pharyngitis and
its safety. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 36:170–173.
2011.(In Chinese). PubMed/NCBI
|
|
44
|
Deng Xu-xia and Lyu Xiang: Efficency
compirson of Herba Rabidosiae Rubescentis dripping pills and Herba
Rabdosiae Rubescentis buccal tablets in the treatment of recurrent
oral ulcer. Henan Traditional Chin Med. 37(4)2017.(In Chinese).
|
|
45
|
Mabaera R, West RJ, Conine SJ, Macari ER,
Boyd CD, Engman CA and Lowrey CH: A cell stress signaling model of
fetal hemoglobin induction: What doesn't kill red blood cells may
make them stronger. Exp Hematol. 36:1057–1072. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Pace BS, Qian XH, Sangerman J,
Ofori-Acquah SF, Baliga BS, Han J and Critz SD: p38 MAP kinase
activation mediates gamma-globin gene induction in erythroid
progenitors. Exp Hematol. 31:1089–1096. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Cokic VP, Andric SA, Stojilkovic SS,
Noguchi CT and Schechter AN: Hydroxyurea nitrosylates and activates
soluble guanylyl cyclase in human erythroid cells. Blood.
111:1117–1123. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hsiao CH, Li W, Lou TF, Baliga BS and Pace
BS: Fetal hemoglobin induction by histone deacetylase inhibitors
involves generation of reactive oxygen species. Exp Hematol.
34:264–273. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Oh HN, Seo JH, Lee MH, Yoon G, Cho SS, Liu
K, Choi H, Oh KB, Cho YS, Kim H, et al: Oridonin induces apoptosis
in oral squamous cell carcinoma probably through the generation of
reactive oxygen species and the p38/JNK MAPK pathway. Int J Oncol.
Mar 16–2018.doi: 10.3892/ijo.2018.4319 (Epub ahead of print).
View Article : Google Scholar :
|
|
50
|
Huang J, Wu LS, Tashiro S, Onodera S and
Ikejima T: Reactive oxygen species mediate oridonin-induced HepG2
apoptosis through p53, MAPK, and mitochondrial signaling pathways.
J Pharmacol Sci. 107:370–379. 2008. View Article : Google Scholar : PubMed/NCBI
|