Introduction
Choline is a water-soluble and vitamin-like
nutrient. In humans, cellular absorption of choline occurs via
choline transporters. It is subsequently metabolized and used for
various functions, such as in the regulation of osmotic pressure,
as a methyl donor precursor in epigenetics, and for phospholipid
synthesis. Choline metabolism is divided into three major pathways:
i) Acetylation, ii) phosphorylation, and iii) oxidation, which are
associated with the synthesis of acetylcholine, phosphatidylcholine
(PC), and methyl group donors, respectively. To date, three major
choline transporters have been identified (1). The high-affinity choline transporter,
CHT1/SLC5A7, is believed to be unique to cholinergic neurons
(2). Second, polyspecific organic
cation transporters (OCT1-3/SLC22A1-3) exhibit a low affinity
toward choline (3). Lastly,
choline transporter-like proteins (CTL1-5/SLC44A1-5) are present in
various human tissues (4).
Previous reports have demonstrated that CTL1 is often functionally
expressed on the cell membrane and is responsible for extracellular
choline transportation (1,5). However, modulation of the choline
uptake system in human homeostasis as well as the signaling
pathways involved remain unclear. Protein kinase C (PKC) regulates
the expression of various transporters on the cell surface, such as
gamma-aminobutyric acid, serotonin, and dopamine transporters
(6). PKC signaling is also
reportedly associated with the repression of choline uptake in
differentiated monocytes (7).
Choline metabolism depends on the cellular environment, by which it
proceeds via various available metabolic pathways. Previous studies
have revealed a major role played by the liver in choline
metabolism (8). The liver is the
major PC-metabolizing organ; therefore, it is strongly associated
with choline metabolism (9),
highlighting the considerable demand for choline by the liver for
its optimal functioning. This suggests that hepatic cell function
is at least partly modulated via PKC signaling. The liver cannot
participate in the active synthesis of phospholipids without
choline; thus, a choline shortage induces various liver-related
diseases (10–12). A better understanding of the
relationship between the liver, choline, and choline metabolites
may lead to an improved understanding of the pathogenesis of
various liver-related diseases. However, the relationship between
the liver, choline uptake system and functional expression of
choline transporters expressed on hepatic cells remains to be
clarified. Therefore, we undertook structural, functional, and
molecular analyses of the choline uptake system in the human
immortalized hepatic cell line, Fa2N-4. Furthermore, we examined
whether choline-deficient conditions and the inhibition of choline
uptake affected cell viability, and caspase-3 and −7 activities. In
addition, we investigated whether treatment of Fa2N-4 cells with a
known PKC activator, phorbol-12-myristate 13-acetate (PMA),
affected modulation of the choline uptake system.
Materials and methods
Cell culture
The immortalized human hepatic cell line, Fa2N-4,
was acquired from Sekisui XenoTech LLC and cultured in RPMI 1640
medium supplemented with 10% fetal bovine serum (Gibco; Thermo
Fisher Scientific, Inc.) and 20 mg/l kanamycin (Gibco; Thermo
Fisher Scientific, Inc.) in collagen-coated flasks. The flasks were
kept in an atmosphere comprising 5% CO2 and 95% air at
37°C. The culture medium was replaced every 2–3 days.
[Methyl-3H]choline uptake
by Fa2N-4 cells
[Methyl-3H]choline chloride (specific
activity: 3182 GBq/mmol) was acquired from PerkinElmer Life
Sciences, Inc.. A DC protein assay kit was acquired from Bio-Rad
Laboratories. PMA and 4α-phorbol 12-myristate 13-acetate (4α-PMA)
were acquired from Wako Pure Chemical Industries, Ltd., and
hemicholinium-3 (HC-3) was obtained from Sigma-Aldrich, Merck KGaA.
We assessed the properties of [methyl-3H]choline
chloride uptake according to a previous study (13). Radioactivity was measured using a
liquid scintillation counter (Tri-Carb® 2100TR, Packard)
and [methyl-3H]choline specific uptake was calculated as
the difference between total [methyl-3H]choline uptake
in the presence and absence of 30 mM of unlabeled choline. Protein
concentrations were determined using a DC protein assay kit
according to the manufacturer's instructions.
RNA extraction and real-time
polymerase chain reaction assay
The cells were first washed using Dulbecco's
phosphate buffered saline (D-PBS). Total RNA was extracted from the
cells using a QIAshredder and RNeasy Mini kit (Qiagen Inc.)
according to the manufacturer's instructions. TaqMan probes for
target mRNAs (CHT1, OCT1-3, CTL1-5, and the housekeeping gene,
glyceraldehyde-3-phosphate dehydrogenase [GAPDH]) were
designed based on its human mRNA sequence using TaqMan®
Gene Expression Assays (Applied Biosystems; Thermo Fisher
Scientific, Inc.). Detailed information on primers for real-time
polymerase chain reaction (PCR) assays is outlined in Table I. Since the sequences related to
the primers and probes of the TaqMan® Gene Expression
Assay are not disclosed, the assay ID is shown in Table I. For one-step real-time PCR, 50 ng
total RNA was added to a master mix using a TaqMan®
RNA-to-CT™ 1-Step kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
Next, real-time PCR analysis was conducted using a StepOne Plus™
Real-Time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.). Relative levels of the mRNAs of target genes were determined
using a comparative cycle time (Ct) method, whereas levels of mRNA
expression relative to GAPDH for each target PCR were
determined as follows: Relative mRNA expression = 2−(Ct target
- Ct GAPDH) × 100%.
 | Table I.TaqMan® gene expression
assay. |
Table I.
TaqMan® gene expression
assay.
| Gene | Accession
number | Assay ID | Exon boundary | Assay location | Amplicon
length |
|---|
| CHT1 (SLC5A7) | NM_021815 | Hs00222367_m1 | 4–5 |
749 | 66 |
| CTL1 (SLC44A1) | NM_080546 | Hs00223114_m1 | 5–6 |
759 | 66 |
| CTL2 (SLC44A2) | NM_020428 | Hs01105936_m1 | 20–21 | 2,060 | 67 |
| CTL3 (SLC44A3) | NM_001114106 | Hs00537043_m1 | 5–6 |
625 | 64 |
| CTL4 (SLC44A4) | NM_001178044 | Hs00228901_m1 | 12–13 | 1,177 | 58 |
| CTL5 (SLC44A5) | NM_152697 | Hs01120485_m1 | 9–10 |
673 | 73 |
| OCT1 (SLC22A1) | NM_003057 | Hs00427554_m1 | 8–9 | 1,493 | 66 |
| OCT2 (SLC22A2) | NM_003058 | Hs01010723_m1 | 10–11 | 1,771 | 120 |
| OCT3 (SLC22A3) | NM_021977 | Hs01009568_m1 | 10–11 | 1,657 | 73 |
| GAPDH | NM_002046 | Hs99999905_m1 | 3–3 |
243 | 122 |
Immunoblotting
A RIPA lysis buffer system (sc-24948) and β-ME
Sample Treatment for Tris-SDS were acquired from Santa Cruz
Biotechnology, Inc. and Cosmo Bio Corporation, respectively.
Protein Multicolor III was acquired from BioDynamics Laboratory
Inc.. A Mini-PROTEAN® TGX™ Gel and
Trans-Blot® Turbo™ Transfer Pack were acquired from
Bio-Rad Laboratories, Inc. Anti-CTL1 polyclonal antibody
(ab110767), anti-NaK-ATPase monoclonal antibody (ab76020) as an
internal control for the cell membrane, and anti-cytochrome
c oxidase (COX) IV antibody (ab16056) were acquired from
Abcam. Anti-CTL2 monoclonal antibody (clone 3D11) was obtained from
Abnova Corporation. Anti-β-actin pAb-HRP-DirecT antibody (PM053-7)
was obtained from MBL. Fa2N-4 cell culture was performed according
to a previously published procedure (13). In brief, membranes were incubated
with rabbit anti-CTL1 polyclonal (ab110767) and anti-CTL2
monoclonal (clone 3D11) antibodies. Protein bands were separated by
SDS-polyacrylamide gel electrophoresis, blotted onto a PVDF
membrane and visualized using an ECL Prime Western Blotting
Detection system (GE Healthcare Life Sciences). Luminescent images
were acquired using a ChemiDoc XRS Plus system (Bio-Rad
Laboratories, Inc.). A Mitochondria/Cytosol Fractionation kit
(ab65320) was acquired from Abcam plc and used to isolate proteins
from the mitochondrial fraction. A Trident Membrane Protein
Extraction kit (Genetex, Inc.) was used to obtain proteins from the
membrane fraction.
Immunofluorescence staining
Wash solution, detector blocking solution, and
horseradish peroxidase-conjugated anti-rabbit and anti-mouse IgG
were acquired from Kirkegaard and Perry Laboratories Inc.
Vectashield mounting medium containing
4′,6-diamidino-2-phenylindole (DAPI) was acquired from Vector
Laboratories, Inc.. In addition, Alexa Fluor 488 goat anti-rabbit,
anti-mouse IgG, 568 goat anti-rabbit, and anti-mouse IgG were
acquired from Molecular Probes Inc. Fa2N-4 cells cultured on a
35-mm glass base dish (Iwaki Glass Co.) were washed twice with
D-PBS and fixed with 100% methanol for 20 min at room temperature.
Consequently, the cells were treated with iBind Flex Solution
(Thermo Fisher Scientific, Inc.) for 1 h. Co-localization of CTL1
with the cell membrane was examined using a NaK-ATPase antibody and
that of CTL2 within the mitochondrial membrane using a
mitochondrial marker, COX IV antibody. Antibody staining was
performed according to a previously published protocol (13). Immunofluorescence images were
obtained using a confocal laser scanning microscope FV10i-DOC
(Olympus).
Cell viability assay
Choline chloride, and RPMI 1640 medium, with and
without choline chloride were acquired from Wako Pure Chemical
Industries, Ltd. Fa2N-4 cells were plated at a density of
5×104 cells/well in 24-well plates. Inhibitors were
added 24 h after cell plating, and the final volume of the medium
in each well was maintained at 1.0 ml. Cell numbers were measured
using an ATPLite™ luminescence ATP detection assay system
(PerkinElmer Life and Analytical Sciences) according to the
manufacturer's instructions. A FilterMax F5 Multi-Mode Microplate
Reader was used to measure luminescence (Molecular Devices,
LLC).
Measurement of caspase-3 and −7
activities
Caspase-3 and −7 activities were measured using a
Caspase-Glo® 3/7 Assay kit (Promega Corporation)
according to the manufacturer's instructions. In brief, this kit is
based on cleavage of the DEVD sequence of a luminogenic substrate
by caspase-3 and −7, emitting a luminescence signal. Fa2N-4 cells
were seeded at a density of 5×104 cells/well in 24-well
plates. HC-3 was added or choline-deficient medium was replaced
after cell plating for 24 h. Each well had a final volume of 1.0 ml
medium. Caspase-3 and −7 activities were measured after the
addition of HC-3 or replacement of choline-deficient medium using a
Caspase-Glo®3/7 Assay kit. A FilterMax F5 Multi-Mode
Microplate Reader was used to measure luminescence.
Apoptotic/necrotic/healthy cell
detection assay
Apoptotic, necrotic, and healthy cells were detected
using an apoptotic/necrotic/healthy cell detection kit (Promocell)
according to the manufacturer's instructions. Briefly, cells were
initially seeded and cultured at 80 to 90% confluency. For
experiments, cells were cultured in defined RPMI-1640 with normal
saline for 48 and 72 h as a control, without choline chloride
(choline deficiency: CD) for 48 h, or with 1 mM of HC-3 (HC-3) for
72 h, and then stained with Hoechst 33342 (blue, nucleus),
fluorescein isothiocyanate (FITC)-Annexin V (green) and ethidium
homodimer III (EthD-III; red) for 15 min. A Confocal Laser Scanning
Biological Microscope (FV10i-DOC; Olympus) was used to acquire
fluorescent images.
Statistical analysis
Data are presented as mean ± standard deviation
(SD). Statistical analyses were conducted using Dunnett's multiple
comparison test and unpaired t-test using GraphPad Prism 8 software
(GraphPad Software, Inc.). Statistical significance was assigned
for P-values <0.05. GraphPad Prism 6 was used to calculate
kinetic parameters with Michaelis-Menten kinetics. We also applied
saturation (hyperbola) kinetics to the values. Ki values
were derived using Ki = IC50/[1 +
(L)/Km], where (L) represents the radiolabeled ligand
concentration from half maximal inhibitory concentration
(IC50) values as previously described (14).
Results
Choline transporter mRNA and protein
in Fa2N-4 cells
CHT1, CTL1-5, and OCT1-3 mRNA levels were examined
using real-time PCR. CTL1 and CTL2 mRNAs were expressed at higher
levels, whereas those of CHT1, CTL4, CTL5, OCT1, and OCT2 were
expressed below measurable limits in Fa2N-4 cells. Furthermore,
CTL3 and OCT3 mRNAs were expressed at low levels (Fig. 1A). Both CTL1 and CTL2 expression
was subsequently measured at the protein level using western blot
analysis. Immunoblotting with CTL1 and CTL2 antibodies revealed
bands corresponding to 70 and 60 kDa, respectively (Fig. 1B). The subcellular distribution of
CTL1 and CTL2 in Fa2N-4 cells was determined using
immunofluorescence staining. CTL1 immunoreactivity was
predominantly identified on the cell surface and overlapped with
the cell surface marker, NaK-ATPase (Fig. 1C). In contrast, CTL2
immunoreactivity was identified in intracellular compartments and
overlapped with that of COX IV, a mitochondrial marker (Fig. 1C). Thus CTL1 and CTL2 mRNA and
protein were expressed in Fa2N-4 cells, with CTL1 localized to the
cell surface and CTL2 localized in mitochondria.
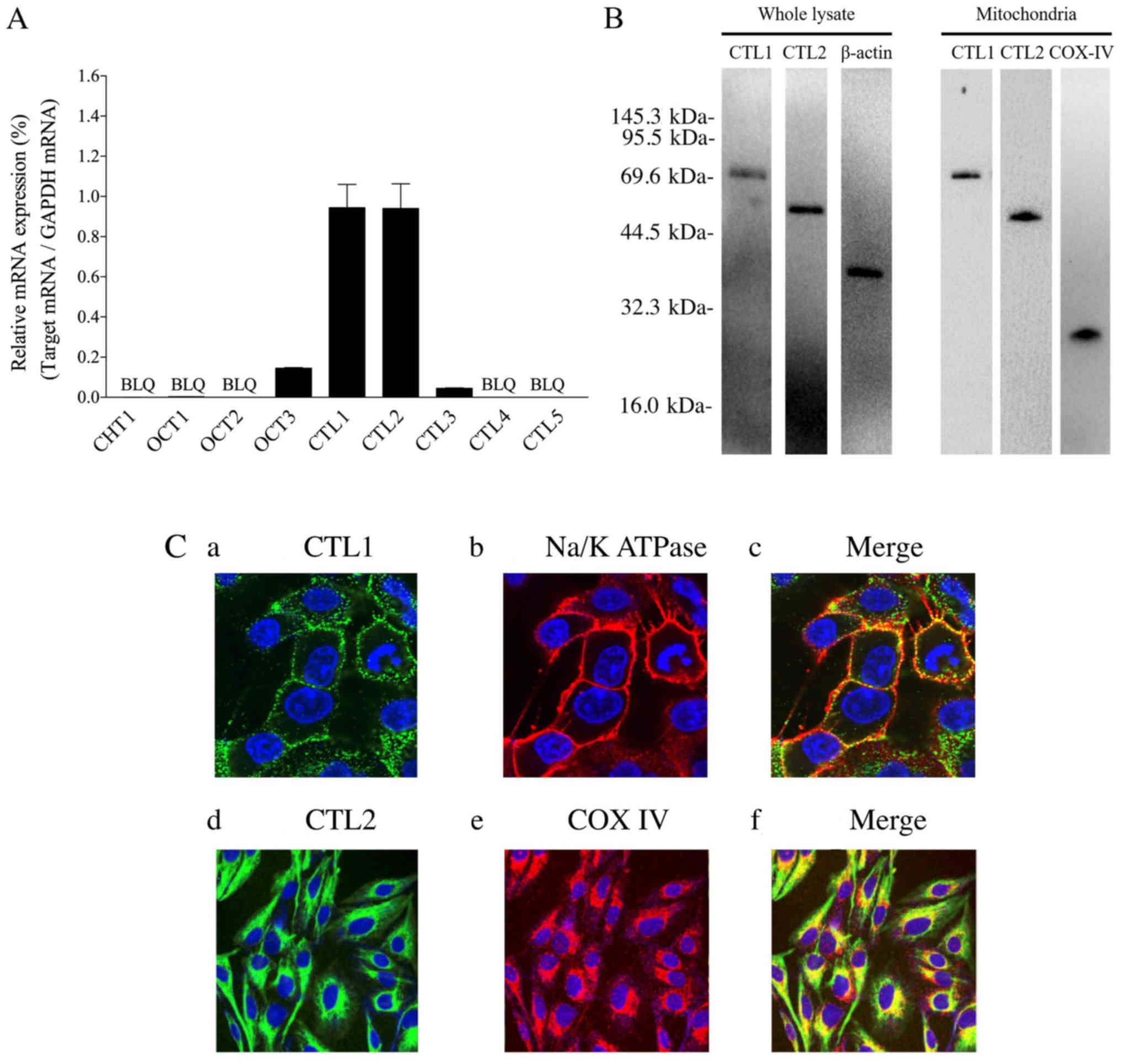 | Figure 1.CTL1 and CTL2 mRNA and protein
expression in Fa2N-4 cells. (A) Relative mRNA expression is
presented as a ratio of target mRNA to GAPDH mRNA (n=3). Each
column represents mean ± standard deviation. BLQ indicates values
below the limit of quantification. (B) Fa2N-4 whole cell and
mitochondrial lysates were used. With a whole cell protein lysate,
choline transporter-like protein 1 (CTL1), CTL2 and β-actin
proteins were detected, whereas CTL1, CTL2, and COX–IV proteins
were detected within mitochondrial lysate. (C) Localization of CTL1
and CTL2 proteins in Fa2N-4 cells were analyzed using the cell
membrane and mitochondrial markers, NaK-ATPase and COX IV,
respectively. The presence of CTL1 is indicated by a green signal
(a), and a merged image of CTL1 and NaK-ATPase (b) is presented in
panel (c). The presence of CTL2 is represented a green signal (d),
and merged images of CTL2 and COX IV (e) are shown in panel (f).
4′,6-diamidino-2-phenylindole (DAPI)-stained nuclei are shown as
blue signals in each panel. Images in (a-c) have a magnification,
×120 whereas (d and e) have a magnification, ×60. CTL, choline
transporter-like protein; COX–IV, cytochrome c oxidase; BLQ,
below the limit of quantification. |
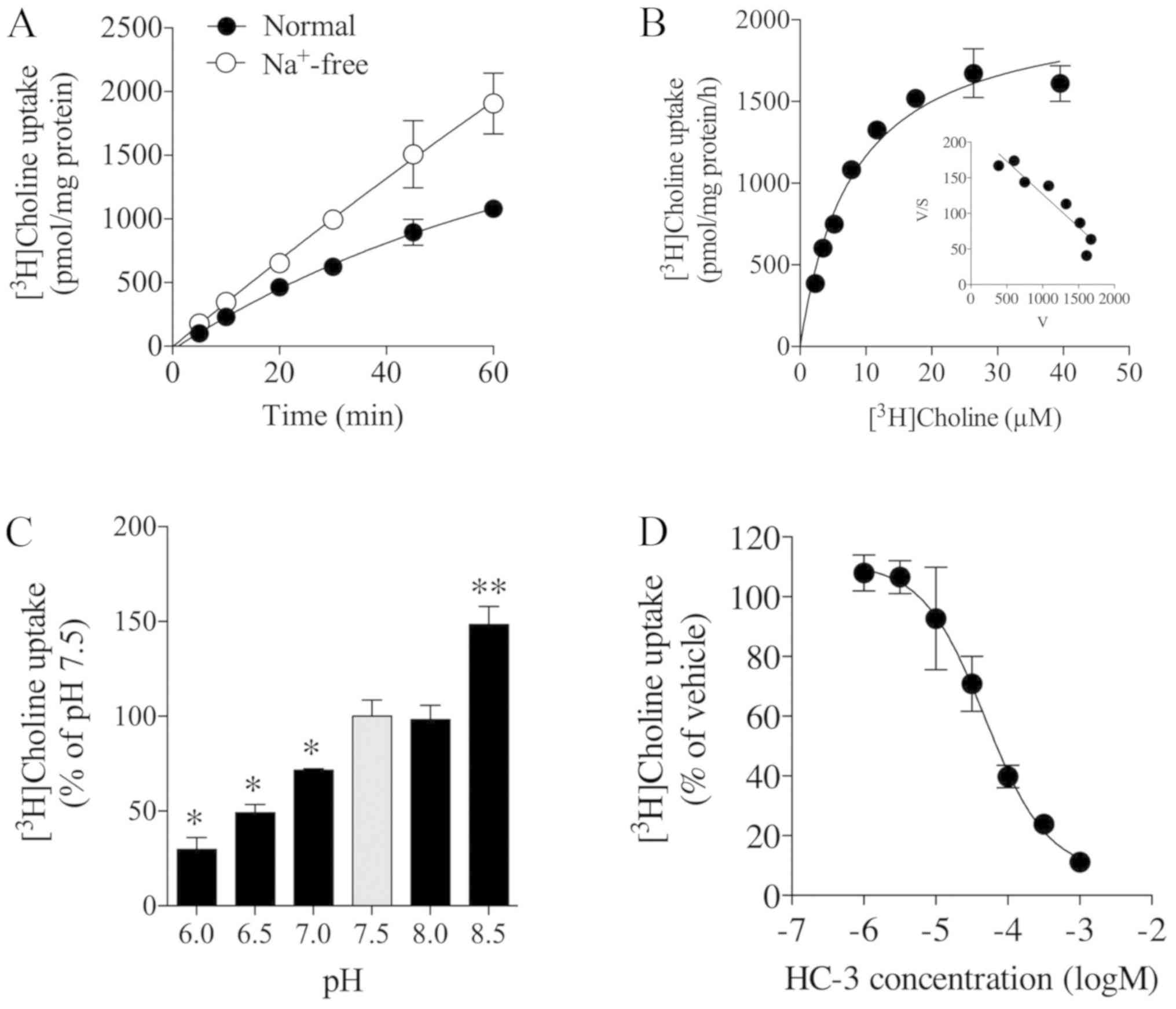
[Methyl-3H]choline uptake
in Fa2N-4 cells
We next examined the time course of
[methyl-3H]choline uptake in Fa2N-4 cells at a
concentration of 10 µM in the presence and absence of extracellular
Na+. [Methyl-3H]choline uptake linearly
increased in a time-dependent manner for at least 20 min (Fig. 2A). Upon the replacement of NaCl
with N-methyl-D-glucamine chloride (NMDG-Cl) in the uptake buffer,
[methyl-3H]choline uptake increased under
Na+-free conditions compared with that of the control
under normal conditions (Fig. 2A).
The kinetic analysis of [methyl-3H]choline uptake
computed by saturation kinetics analysis generated a
Michaelis-Menten constant (Km) of 8.0±0.1 µM and maximal
velocity (Vmax) of 2,100±107.1 pmol/mg protein/h
(Fig. 2B). We also applied
saturation (hyperbola) kinetics to the values; however, the
respective Km and Vmax were the same. An
Eadie-Hofstee plot showed a single process in
[methyl-3H]choline uptake based on linear regression
analysis (Fig. 2B). The effect of
extracellular pH on [methyl-3H]choline uptake in Fa2N-4
cells was examined by varying the pre-incubation medium pH between
6.0 and 8.5. [Methyl-3H]choline uptake was significantly
decreased and increased when extracellular pH was reduced from 7.5
to 6.0 and increased from 7.5 to 8.5, respectively (Fig. 2C). Furthermore, the effect of HC-3,
an inhibitor of choline uptake, on [methyl-3H]choline
uptake in Fa2N-4 cells was examined. It was observed that the
uptake was inhibited by HC-3 in a concentration-dependent manner,
with a Ki value of 48.2 µM (Fig. 2D). Thus,
[methyl-3H]choline uptake by Fa2N-4 cells increased in a
time-and pH-dependent manner, and was increased in the absence of
Na+ and inhibited by HC-3.
Choline deficiency and HC-3 decreased
cell viability, increased caspase-3 and −7 activities, and induced
apoptotic change in Fa2N-4 cells
The effect of choline deficiency and 1 mM HC-3
treatment on caspase-3 and −7 activities in Fa2N-4 cells was
evaluated. It was observed that HC-3 treatment and choline
deficiency significantly inhibited cell viability and increased
caspase-3 and −7 activities in these cells (Fig. 3A). We further examined whether cell
death by choline deficiency and treatment with HC-3 occurs through
apoptosis or necrosis (Fig. 3B).
In this experiment, immune cells were stained by Hoechst 33342
(blue, nuclei) only, and not by FITC-Annexin V (green) or EthD-III
(red) as observed in normal saline treated cells, after 48 and 72
h. Early apoptotic cells were stained both blue and green, whereas
necrotic cells were stained blue and red. Cells that stained blue,
green and red represented dead cells progressing from an apoptotic
cell change (late apoptotic cells), or necrotic cell change. In
Fig. 3B, choline deficiency and 1
mM HC-3 treatment induced marked changes as shown by blue and green
staining, which was interpreted as an apoptotic change. Thus, HC-3
treatment and choline deficiency significantly inhibited cell
viability, increased caspase-3 and −7 activities and induced
apoptosis in Fa2N-4 cells.
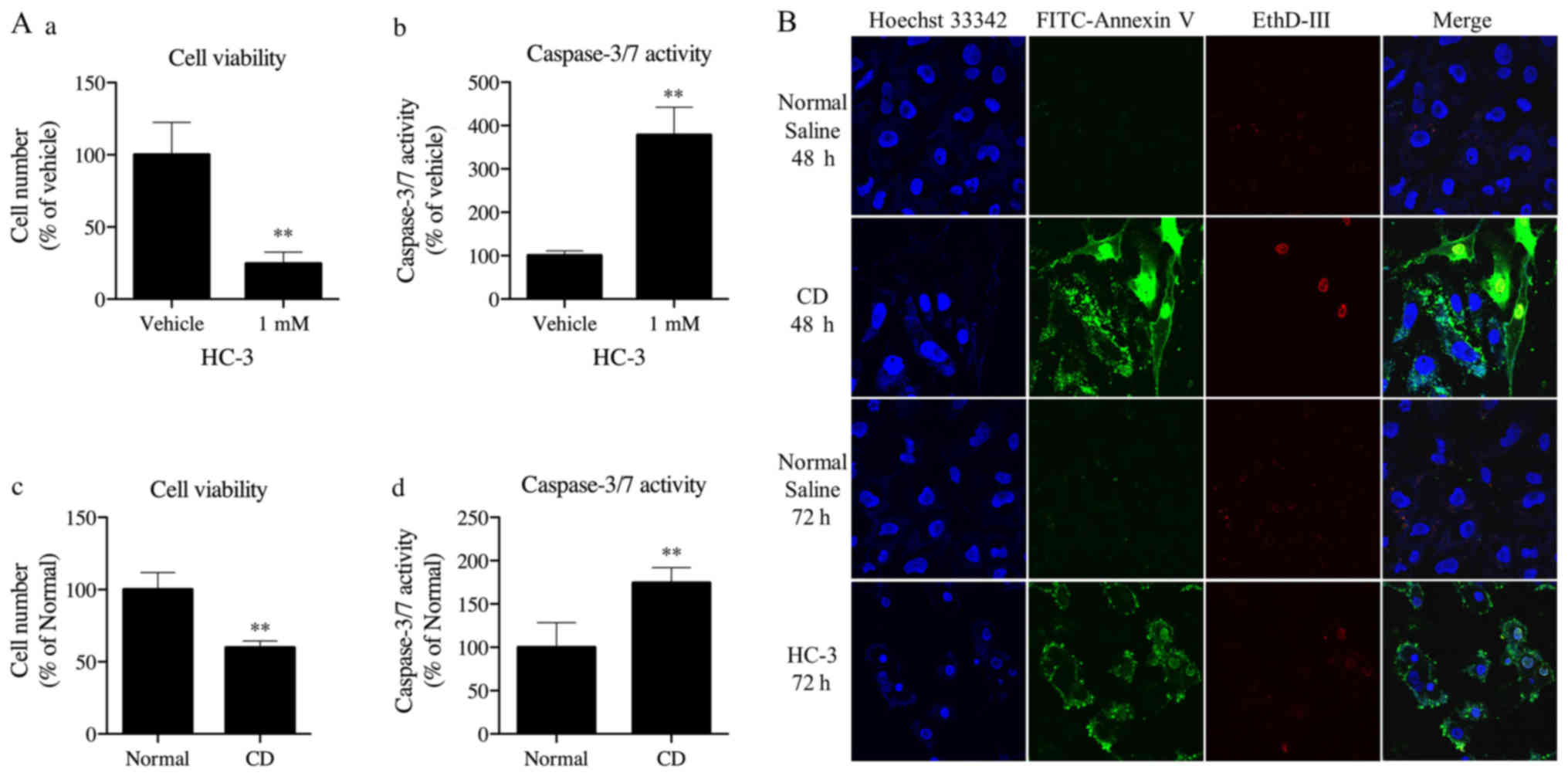 | Figure 3.(A) Effect of HC-3 and choline
deficiency on cell viability and caspase-3 and −7 activities in
Fa2N-4 cells. (a) Treatment of Fa2N-4 cells with 1 mM HC-3
significantly inhibited cell viability and (b) significantly
increased caspase-3 and −7 activities. (c) Fa2N-4 cells were
cultured in RPMI 1640 medium with (normal) or without 30 µM choline
chloride (CD). CD treatment significantly inhibited cell viability
and (d) increased caspase-3 and −7 activities in Fa2N-4 cells. Each
value represents mean ± standard deviation. (A and B, n=4; C and D,
n=6). **P<0.01 compared with the vehicle or normal group. (B)
Choline deficiency and HC-3 treatment induced apoptosis in Fa2N-4
cells. Cells were initially seeded and cultured at 80 to 90%
confluency, and then in defined RPMI-1640 with normal saline for 48
and 72 h as a control, without choline chloride (CD) for 48 h, or
with 1 mM of HC-3 (HC-3) for 72 h. Cells were stained with Hoechst
33342 (blue, nucleus), FITC-Annexin V (green) and ethidium
homodimer V (EthD-V, red) for 15 min. Immune cells were stained
using Hoechst 33342 (blue, nuclei), and not by FITC-Annexin V
(green) or EthD-V (red). Early apoptotic cells were stained blue
and green, whereas necrotic cells were stained blue and red. Cells
stained blue, green and red represented dead cells progressing from
an apoptotic cell change (late apoptotic cells), or necrotic cell
change. Magnification, ×60. HS-3, hemicholinium-3; CD,
choline-deficient. |
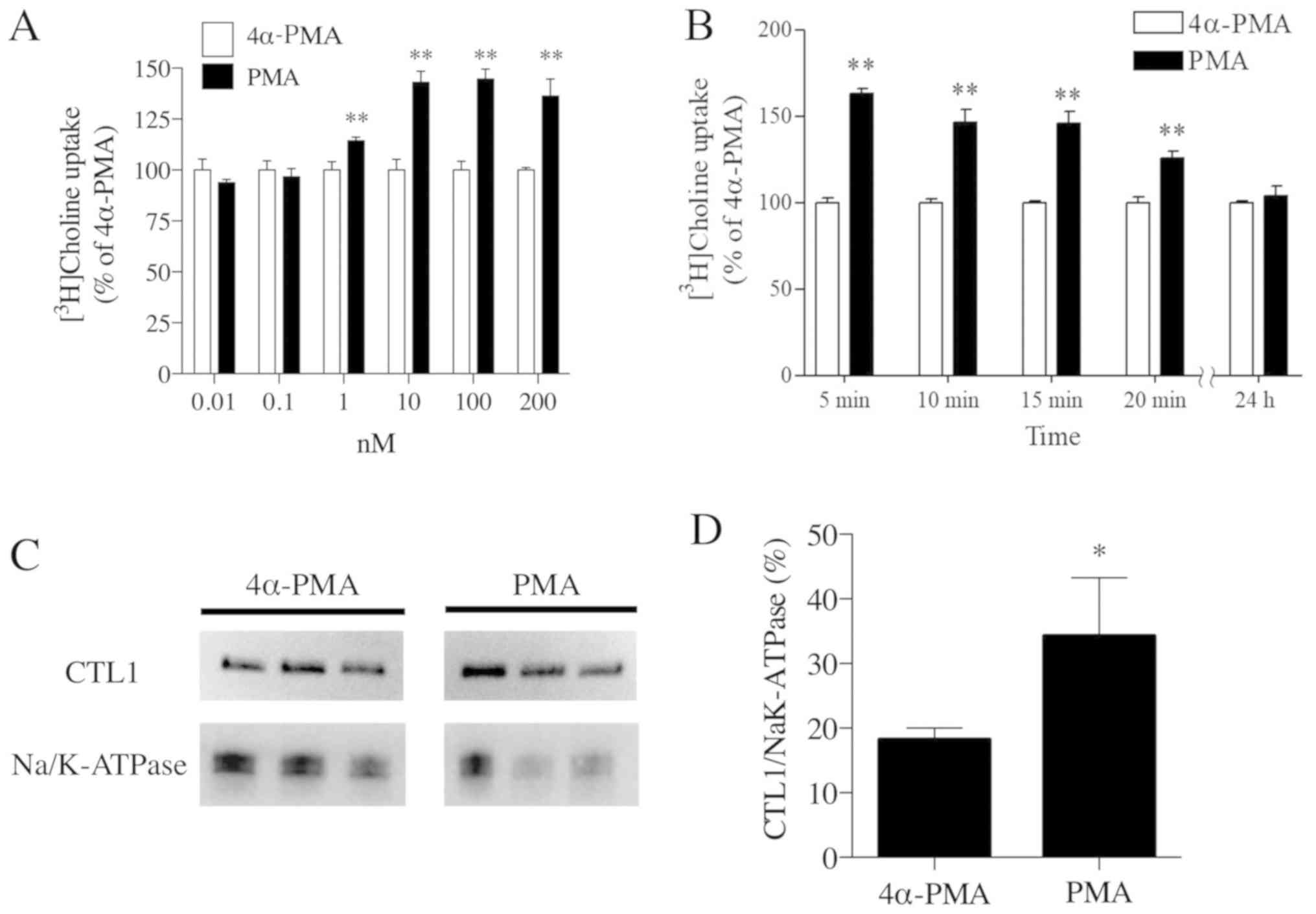
PKC stimulation of the choline
transport system and CTL1
Compared with the 4α-PMA negative control, PMA
treatment increased [methyl-3H]choline uptake in Fa2N-4
cells. The highest uptake of [methyl-3H]choline occurred
when PMA was used at a concentration of 100 nM (Fig. 4A). Using 100 nM as the optimal
concentration of PMA, the time course of choline uptake was
analyzed, with the highest uptake of [methyl-3H]choline
occurring 5 min after PMA treatment (Fig. 4B). However, chronic treatment (24
h) with PMA did not increase or decrease choline uptake compared
with 4α-PMA. Following stimulation by PMA, an activator of PKC, and
using 4α-PMA as a negative control, cell membrane protein
components were extracted using a Trident Membrane Protein
Extraction kit. The extracted proteins were electrophoresed and
underwent western blot analysis to detect CTL1 (Fig. 4C). The band intensities were
quantified and the relative expression levels of CTL1 were
standardized using NaK-ATPase. PMA-treated cells were found to have
a significantly higher CTL1 band intensity than 4α-PMA-treated
cells (Fig. 4D). Thus, PKC
activation induced [methyl-3H]choline uptake in Fa2N-4
cells and increased CTL1 in cell membranes.
Discussion
The analysis of choline transporter expression in
Fa2N-4 cells indicated the high expression of CTL1 and CTL2 mRNAs,
and negligible or low expression of CTL3 and OCT3 mRNAs.
Functionally, a single choline transportation system, which
operates independently of Na+, is responsible for
choline uptake in Fa2N-4 cells. The Km value of choline
chloride for the choline transporter was 8.0±0.1 µM, which is
within the choline concentration range in the human liver (5–30 µM)
(15), whereas the Ki
value using HC-3 was 48.2 µM. CTL1 functions in a pH- and membrane
potential-dependent manner, can operate independently of
Na+, is sensitive to organic cations, and has a proven
affinity for choline (Km value of 10–30 µM) and HC-3
(Ki> 10 µM). These characteristics of CTL1 overlap
with the results of the current study. In contrast, because CHT1
operates in a Na+-dependent manner toward choline
transportation and is inhibited by HC-3 at very low concentrations
(Ki value, 50–100 nM) (16), CHT1 mRNA is expressed below
quantifiable levels in Fa2N-4 cells; this does not support the
concept of CHT1 predominantly contributing to the choline transport
system of Fa2N-4 cells. Furthermore, choline transportation by OCTs
is known to be independent of Na+ ions. OCT1 and OCT2
have Km values of 620 and 210 µM, respectively, whereas
OCT3 does not recognize choline as a substrate (17,18).
These results do not support the theory that OCTs play a
predominant role in choline transportation in Fa2N-4 cells.
To date, it has been suggested that a CTL1-mediated
choline uptake system functions in primary human tumor cell
cultures, such as colon carcinoma cells (19), keratinocytes (20), esophageal cancer cells (21), small cell lung carcinoma cells
(22), tongue cancer cells
(13), and trophoblastic cells
(23). Likewise, CTL1 is
ubiquitously expressed in mammalian tissues, suggesting an
important role in choline transportation. Excluding the prospect
that CHT1 and OCT1-3 operate during choline transportation in
Fa2N-4 cells, we hypothesize that CTL1 is solely responsible for
extracellular choline transportation associated with Fa2N-4 cells.
However, it may be that CTL2-5 are also involved in choline
transportation in such cells. Nevertheless, because the functional
characteristics of CTL2-5 have not been completely investigated, it
is difficult to determine their roles in choline transportation. To
further support our hypothesis that CTL1 is the primary
extracellular choline transporter in Fa2N-4 cells, we examined CTL1
expression and localization. A major band of approximately 70 kDa
was observed in the immunoblots of Fa2N-4 cell-derived proteins,
which corresponded with the estimated size of human CTL1 of 73.3
kDa. Immunofluorescence staining and visualization of CTL1 showed
this to overlap with NaK-ATPase, clearly indicating CTL1 expression
in the cell membrane. Although we did not directly show a
biochemical association of CTL1 with extracellular choline
transportation, we sought to support our hypothesis that CTL1 is
responsible for choline transportation by investigating
[methyl-3H]choline uptake in Fa2N-4 cells. However,
because immunofluorescence staining of CTL2 overlapped with COX IV,
a known mitochondrial marker, CTL2 was believed to play an
essential role in transporting choline to mitochondria in Fa2N-4
cells. Furthermore, choline oxidation occurs in the mitochondria;
in particular, it is elevated in the liver and kidneys,
corresponding to the high energy demand of hepatocytes (24). Therefore, it follows that choline
transporters are abundantly expressed at these locations. Based on
mRNA expression analysis, western blot analysis and
immunofluorescence staining, we hypothesized that CTL2 may be
responsible for choline transportation in the mitochondria.
However, we cannot eliminate the possibility that a novel, yet
unidentified transporter, distinct from CTL2, has a function in
choline transportation in the mitochondria. Moreover, our
hypothesis is only based on mRNA and protein expression analysis.
Further functional studies are necessary to draw a conclusion on a
role for CTL2 in choline transportation in the mitochondria.
Next, we assessed the effects of choline deficiency
and the inhibition of choline uptake on Fa2N-4 cells. Because the
two treatments reduced cell viability, upregulated caspase-3 and −7
activities and markedly increased FITC-Annexin V immunofluorescence
staining, we attributed these changes to apoptosis. Previous
reports have described how choline deficiency or the inhibition of
choline uptake induced apoptosis via the p53 pathway by increasing
the synthesis of ceramide, a known apoptosis-inducing substance
(25). Sphingomyelin, a principal
component of the cell membrane, can be separated into ceramide and
phosphorylcholine by sphingomyelinase (PC); consequently, PC can be
used as a donor in phospholipid synthesis (9). In addition, PC after hydrolysis
mediates Raf activation in response to mitogenic growth factors
(26). These mechanisms indicate
that the choline transportation system, via CTL1, is well
associated with cell survival.
Next, we explored the modulation of choline uptake
via CTL1. PKC operates as one of the key signals in modulating the
trafficking of transporters to the cell membrane, e.g., GABA
transporter (27), serotonin
transporter (28), dopamine
transporter, and excitatory amino acid carrier (28,29).
Except in trafficking, neuronal PKC activity can regulate CHT1
protein and its surface expression, suggesting that CHT is a
phosphoprotein (30). CTL1
modulation and trafficking by PKC signaling have been reported in
human THP-1 monocytic cells, while PMA treatment has been
demonstrated to induce impaired CTL1 trafficking at the plasma
membrane level and to inhibit choline transportation (7). However, it was concluded that PMA
treatment induced THP-1 monocytic cell differentiation in
macrophages, resulting in altered choline uptake. In the current
study, we used Fa2N-4, an established immortalized human hepatocyte
cell line that has proven to be useful for liver-based in
vitro testing and liver research (31,32).
Regarding the PKC-dependent pathway in Fa2N-4 cells, treatment with
palmitic acid has been associated with PKC signaling and has led to
the induction of CYP3A4 activity (33). However, investigations regarding
the effect of PKC signaling on CTL1 modulation and trafficking in
Fa2N-4 or HepG2 cells are lacking. PKC is a known key regulator of
cell growth, and its hyperactivation is believed to play a major
role in tumor progression (34).
Because the HepG2 cell line is derived from a hepatoblastoma, the
expectation was that its progression would be easily affected by
PKC stimulation (35). Therefore,
we selected the Fa2N-4 cell line for this study. We showed a
proportional increase of choline uptake by Fa2N-4 cells with
increasing PMA concentrations and also showed that PKC stimulation
induced CTL1 translocation into the cell membrane. Accordingly, we
conclude that PKC stimulation induced CTL1 to translocate to the
cell membrane, resulting in improved choline uptake. However, our
study was limited in that we only investigated whether PMA
treatment resulted in changes in choline uptake and did not
consider the possibility that such changes also reflect the total
outcome of PKC-dependent functions. Moreover, it is known that PKC
delivers insulin signals at the same time, and that the
hyperinsulinemia state is considerably associated with
non-alcoholic fatty liver disease (NAFLD). However, we did not
examine the pathological role of the CTL1/PKC axis in
hyperinsulinemia in this current study. We intend to explore this
relationship in future. To dissect the phenomenon of PKC signaling
overall, it is necessary to examine the role of each PKC
phosphorylation site of CTL1 in the choline uptake system.
We believe that the current investigation on the
choline uptake system in human immortalized hepatic cells will help
in providing a fundamental understanding of its contribution to the
pathogenesis of hepatic diseases. For instance, choline deficiency
in non-alcoholic steatohepatitis (NASH) model mice induces fat
accumulation and hepatic inflammation, leading to fatty liver
disease. The mechanism underlying this disease is associated with
PKC activation (36,37). Moreover, it is known that despite
the availability of sufficient choline, inhibition of
phosphatidylethanolamine methyltransferase leads to the
pathogenesis of lean NASH (38).
Exogenous choline deficiency has been shown to be related to the
pathogenesis of NAFLD, NASH, and hepatocarcinogenesis (10–12).
Therefore, it is obvious that choline is essential for liver
homeostasis, and that a higher dietary choline intake is associated
with a lower risk of NAFLD and primary liver cancer (39,40).
Then, the current study placed one of major focuses on lean state
conditions by inducing choline shortage to Fa2N-4 cells. It is
logical that researchers have focused on, or are likely to focus
on, the metabolites of choline and its related enzymes. Primarily,
considering the pathogeneses of NAFLD and NASH, choline uptake via
CTL1 may be limited in hepatocytes. It has been demonstrated that
patients with NAFLD and NASH have lower accumulated choline in the
liver on positron emission tomography-computed tomography (41). However, regarding the pathogenesis
of hepatocellular carcinoma (HCC) and lean NASH being induced under
choline-deficient conditions, we believe that a possible mechanism
is that a choline-deficient condition causes an increase in
ceramide concentration via sphingomyelin activation, consequently
inducing cell damage and apoptosis. Hence, we believe that
exploring the modulation of the choline uptake system via CTL1 by
PKC signaling may provide a novel strategy to treat NAFLD, NASH,
and HCC in future.
Although we present novel findings that facilitate
our understanding of hepatic choline transportation, the major
limitation of this study is that in vitro only studies were
undertaken. In addition, we did not examine whether PKC directly
regulates CTL1 to translocate to the cell membrane and induce
increased extracellular choline uptake, or whether this occurred by
phosphorylation signals to CTL1 proteins. It is necessary to
explore these relationships in future. Moreover, it is imperative
to further investigate our hypothesis using in vivo
conditions and clinical trials to gain a better understanding of
hepatic disorders, such as NAFLD, NASH, and HCC.
In summary, we conclude that extracellular choline
is transported via CTL1 in Fa2N-4 cells. Moreover, we hypothesize
that CTL1 and the choline uptake system are associated with cell
survival, and that the choline uptake system is modulated by PKC
signaling via increased cell surface CTL1 expression. These
findings may provide further insights into the pathogenesis of
liver disease.
Acknowledgements
Not applicable.
Funding
No funding was received.
Authors' contributions
TI, MO and MI made substantial contributions to the
conception of the present study. TI and MI performed the
experiments, analyzed and interpreted the data. TI was involved in
writing and revising the manuscript and MI assisted with writing
and revising the manuscript. HS, JS and RS assisted with the
analysis, interpretation of data and revised the manuscript
critically. TY was involved in the experimental work and reviewed
the manuscript critically. RS, MO and MI supervised the research.
All authors read and approved the final version of the manuscript.
All authors agreed to be accountable for all aspects of the work in
ensuring that questions related to the accuracy or integrity of the
work are appropriately investigated and resolved.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
GAPDH
|
glyceraldehyde-3-phosphate
dehydrogenase
|
|
HCC
|
hepatocellular carcinoma
|
|
NAFLD
|
non-alcoholic fatty liver disease
|
|
NASH
|
non-alcoholic steatohepatitis
|
|
PCR
|
polymerase chain reaction
|
|
PMA
|
phorbol 12-myristate 13-acetate
|
|
4α-PMA
|
4α-phorbol 12-myristate 13-acetate
|
|
PC
|
phosphatidylcholine
|
References
|
1
|
Michel V, Yuan Z, Ramsubir S and Bakovic
M: Choline transport for phospholipid synthesis. Exp Biol Med
(Maywood). 231:490–504. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Okuda T, Haga T, Kanai Y, Endou H,
Ishihara T and Katsura I: Identification and characterization of
the high-affinity choline transporter. Nat Neurosci. 3:120–125.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Koepsell H, Lips K and Volk C:
Polyspecific organic cation transporters: Structure, function,
physiological roles, and biopharmaceutical implications. Pharm Res.
24:1227–1251. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Traiffort E, Ruat M, O'Regan S and Meunier
FM: Molecular characterization of the family of choline
transporter-like proteins and their splice variants. J Neurochem.
92:1116–1125. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nakamura T, Fujiwara R, Ishiguro N, Oyabu
M, Nakanishi T, Shirasaka Y, Maeda T and Tamai I: Involvement of
choline transporter-like proteins, CTL1 and CTL2, in
glucocorticoid-induced acceleration of phosphatidylcholine
synthesis via increased choline uptake. Biol Pharm Bull.
33:691–696. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Robinson MB: Regulated trafficking of
neurotransmitter transporters: Common notes but different melodies.
J Neurochem. 80:1–11. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fullerton MD, Wagner L, Yuan Z and Bakovic
M: Impaired trafficking of choline transporter-like protein-1 at
the plasma membrane and inhibition of choline transport in THP-1
monocyte-derived macrophages. Am J Physiol Cell Physiol.
290:C1230–C1238. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schenkel LC, Sivanesan S, Zhang J, Wuyts
B, Taylor A, Verbrugghe A and Bakovic M: Choline supplementation
restores substrate balance and alleviates complications of Pcyt2
deficiency. J Nutr Biochem. 26:1221–1234. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Michel V and Bakovic M: The ubiquitous
choline transporter SLC44A1. Cent Nerv Syst Agents Med Chem.
12:70–81. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Butler LM, Arning E, Wang R, Bottiglieri
T, Govindarajan S, Gao YT and Yuan JM: Prediagnostic levels of
serum one-carbon metabolites and risk of hepatocellular carcinoma.
Cancer Epidemiol Biomarkers Prev. 22:1884–1893. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ghoshal AK and Farber E: The induction of
liver cancer by dietary deficiency of choline and methionine
without added carcinogens. Carcinogenesis. 5:1367–1370. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rinella ME, Elias MS, Smolak RR, Fu T,
Borensztajn J and Green RM: Mechanisms of hepatic steatosis in mice
fed a lipogenic methionine choline-deficient diet. J Lipid Res.
49:1068–1076. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nishiyama R, Nagashima F, Iwao B, Kawai Y,
Inoue K, Midori A, Yamanaka T, Uchino H and Inazu M: Identification
and functional analysis of choline transporter in tongue cancer: A
novel molecular target for tongue cancer therapy. J Pharmacol Sci.
131:101–109. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Haubrich DR and Gerber NH: Choline
dehydrogenase. Assay, properties and inhibitors. Biochem Pharmacol.
30:2993–3000. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wiedeman AM, Dyer RA, Green TJ, Xu Z, Barr
SI, Innis SM and Kitts DD: Variations in plasma choline and
metabolite concentrations in healthy adults. Clin Biochem.
60:77–83. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Apparsundaram S, Ferguson SM, George AL Jr
and Blakely RD: Molecular cloning of a human,
hemicholinium-3-sensitive choline transporter. Biochem Biophys Res
Commun. 276:862–867. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Burckhardt G and Wolff NA: Structure of
renal organic anion and cation transporters. Am J Physiol Renal
Physiol. 278:F853–F866. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gründemann D, Liebich G, Kiefer N, Köster
S and Schömig E: Selective substrates for non-neuronal monoamine
transporters. Mol Pharmacol. 56:1–10. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kouji H, Inazu M, Yamada T, Tajima H, Aoki
T and Matsumiya T: Molecular and functional characterization of
choline transporter in human colon carcinoma HT-29 cells. Arch
Biochem Biophys. 483:90–98. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Uchida Y, Inazu M, Takeda H, Yamada T,
Tajima H and Matsumiya T: Expression and functional
characterization of choline transporter in human keratinocytes. J
Pharmacol Sci. 109:102–109. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nagashima F, Nishiyama R, Iwao B, Kawai Y,
Ishii C, Yamanaka T, Uchino H and Inazu M: Molecular and functional
characterization of choline transporter-like proteins in esophageal
cancer cells and potential therapeutic targets. Biomol Ther.
26:399–408. 2018. View Article : Google Scholar
|
|
22
|
Inazu M, Yamada T, Kubota N and Yamanaka
T: Functional expression of choline transporter-like protein 1
(CTL1) in small cell lung carcinoma cells: A target molecule for
lung cancer therapy. Pharmacol Res. 76:119–131. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yara M, Iwao B, Hara N, Yamanaka T, Uchino
H and Inazu M: Molecular and functional characterization of choline
transporter in the human trophoblastic cell line JEG-3 cells.
Placenta. 36:631–637. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kaplan CP, Porter RK and Brand MD: The
choline transporter is the major site of control of choline
oxidation in isolated rat liver mitochondria. FEBS Lett. 321:24–26.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang XH, Zhao C and Ma ZA: The increase
of cell-membranous phosphatidylcholines containing polyunsaturated
fatty acid residues induces phosphorylation of p53 through
activation of ATR. J Cell Sci. 120:4134–4143. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cai H, Erhardt P, Troppmair J, Diaz-Meco
MT, Sithanandam G, Rapp UR, Moscat J and Cooper GM: Hydrolysis of
phosphatidylcholine couples Ras to activation of Raf protein kinase
during mitogenic signal transduction. Mol Cell Biol. 13:7645–7651.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Beckman ML, Bernstein EM and Quick MW:
Multiple G protein-coupled receptors initiate protein kinase C
redistribution of GABA transporters in hippocampal neurons. J
Neurosci. 19:RC91999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jayanthi LD, Samuvel DJ, Blakely RD and
Ramamoorthy S: Evidence for biphasic effects of protein kinase C on
serotonin transporter function, endocytosis, and phosphorylation.
Mol Pharmacol. 67:2077–2087. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Daniels GM and Amara SG: Regulated
trafficking of the human dopamine transporter. Clathrin-mediated
internalization and lysosomal degradation in response to phorbol
esters. J Biol Chem. 274:35794–35801. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gates J Jr, Ferguson SM, Blakely RD and
Apparsundaram S: Regulation of choline transporter surface
expression and phosphorylation by protein kinase C and protein
phosphatase 1/2A. J Pharmacol Exp Therap. 310:536–545. 2004.
View Article : Google Scholar
|
|
31
|
Hariparsad N, Carr BA, Evers R and Chu X:
Comparison of immortalized Fa2N-4 cells and human hepatocytes as in
vitro models for cytochrome P450 induction. Drug Metab Dispos.
36:1046–1055. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ramboer E, Vanhaecke T, Rogiers V and
Vinken M: Immortalized human hepatic cell lines for in vitro
testing and research purposes. Methods Mol Biol. 1250:53–76. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hu N, Hu M, Duan R, Liu C, Guo H, Zhang M,
Yu Y, Wang X, Liu L and Liu X: Increased levels of fatty acids
contributed to induction of hepatic CYP3A4 activity induced by
diabetes-in vitro evidence from HepG2 cell and Fa2N-4 cell lines. J
Pharmacol Sci. 124:433–444. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Rumsby M, Schmitt J, Sharrard M, Rodrigues
G, Stower M and Maitland N: Human prostate cell lines from normal
and tumourigenic epithelia differ in the pattern and control of
choline lipid headgroups released into the medium on stimulation of
protein kinase C. Br J Cancer. 104:673–684. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bouma ME, Rogier E, Verthier N, Labarre C
and Feldmann G: Further cellular investigation of the human
hepatoblastoma-derived cell line HepG2: Morphology and
immunocytochemical studies of hepatic-secreted proteins. In Vitro
Cell Dev Bio. 25:267–275. 1989. View Article : Google Scholar
|
|
36
|
Greene MW, Burrington CM, Lynch DT,
Davenport SK, Johnson AK, Horsman MJ, Chowdhry S, Zhang J, Sparks
JD and Tirrell PC: Lipid metabolism, oxidative stress and cell
death are regulated by PKC delta in a dietary model of nonalcoholic
steatohepatitis. PLoS One. 9:e858482014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Greene MW, Burrington CM, Ruhoff MS,
Johnson AK, Chongkrairatanakul T and Kangwanpornsiri A: PKC{delta}
is activated in a dietary model of steatohepatitis and regulates
endoplasmic reticulum stress and cell death. J Biol Chem.
285:42115–42129. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Nakatsuka A, Matsuyama M, Yamaguchi S,
Katayama A, Eguchi J, Murakami K, Tesgigawara S, Ogawa D, Wada N,
Yasunaka T, et al: Insufficiency of phosphatidylethanolamine
N-methyltransferase is a risk for lean non-alcoholic
steatohepatitis. Sci Rep. 6:217212016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yu D, Shu XO, Xiang YB, Li H, Yang G, Gao
YT, Zheng W and Zhang X: Higher dietary choline intake is
associated with lower risk of nonalcoholic fatty liver in
normal-weight Chinese women. J Nutr. 144:2034–2040. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhou RF, Chen XL, Zhou ZG, Zhang YJ, Lan
QY, Liao GC, Chen YM and Zhu HL: Higher dietary intakes of choline
and betaine are associated with a lower risk of primary liver
cancer: A case-control study. Sci Rep. 7:6792017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Roppongi M, Izumisawa M, Terasaki K,
Muraki Y and Shozushima M: 18F-FDG and
11C-choline uptake in proliferating tumor cells is
dependent on the cell cycle in vitro. Ann Nucl Med. 33:237–243.
2018. View Article : Google Scholar : PubMed/NCBI
|