Introduction
Intracerebral hemorrhage (ICH) is the most common
human cerebrovascular disease, accounting for 10–15% of strokes
(1), which is characterized by
rupture of one or more blood vessels in the brain and infiltration
of blood into the brain (2).
Hematoma after ICH disrupts neighboring blood vessels, increases
bleeding and hematoma growth, which cause ischemia and impaired
neuronal function (3). Recent
studies have demonstrated that ICH triggers an inflammation cascade
to accelerate the formation of edema, which aggravates the mass
effect around hematoma and amplifies cell death (4,5).
Inflammatory cells, such as blood-derived leukocytes, microglia and
macrophages, are essential for removing the hematoma but also
increase brain damage caused by cerebral hemorrhage (6). Microglia are thought to be the
earliest inflammatory cells of ICH (7), and are the major cell type
responsible for damage after ICH by releasing cytokines,
chemokines, ferrous iron and other immunoactive molecules (8).
MicroRNAs (miRNA) are endogenous small RNA
molecules, which contribute to post-transcriptional gene silencing
and regulate the expression of hundreds of target genes (9). They can control diverse fundamental
biological and pathological processes, such as growth, development
and death (10). Recent studies
found that miRNAs could regulate the transcription of microglia and
the expression levels of related signaling pathways, and
participate in the polarization of microglia (11,12).
For example, it was revealed that overexpression of miRNA-126-3p
could attenuate blood-brain barrier disruption, cerebral edema and
neuronal injury following ICH by regulating PIK3R2 and Akt
(13). In addition, miR-124 could
ameliorate ICH-induced inflammatory injury by modulating microglia
polarization toward the M2 phenotype via C/EBP-α (14).
miR-222 is a component of the miR221/222 cluster,
whose members are located on the X chromosome (XP113) with the same
seed sequence (15). miR-222 is
associated with the pathogenesis and progression of cancer and
functions by affecting cell proliferation, tumor growth and cell
apoptosis (16,17). However, the exact mechanism of
miR-222 on inflammation and brain injury in ICH has not been
elucidated. In the present study, the miR-222 level was detected,
and the potential role of miR-222 in ICH in vivo and in
vitro was further explored. It is expected that miR-222 may
provide new insights into the understanding of cerebral protection
and act as a neuroprotective agent for ICH therapeutics.
Materials and methods
Primary cell cultures
For primary microglia cells, glial cells were
isolated from the brains of rat pups and cultured in DMEM (Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 20% fetal bovine
serum (Invitrogen; Thermo Fisher Scientific, Inc.) in a humidified
atmosphere at 37°C. Microglia was isolated from the mixed glial
population when mixed glial cells were confluent (12–14 days). The
purity of microglia was assessed by immunofluorescence using
microglia-specific antibody CDb11. Microglial cultures with >98%
purity were used for the study.
Animals
In total, 60 male C57BL/6 mice (6–8 weeks, 18–22 g)
were purchased from Model Animal Research Institute of Nanjing
University and bred under specific pathogen-free conditions.
Experiments were performed according to animal care guidelines
approved by The Animal Ethics Committee of Nanjing Medical
University, and animals were treated in accordance with The
Guidelines of the United States National Institutes of Health. Mice
were maintained at constant ambient temperature (22±1°C) under a
12-h light/dark cycle with food and water ad libitum. The
model of mice was established as described (18). Animal health and behavior were
monitored every day.
Preparation of erythrocyte
lysates
Blood was obtained from C57BL/6 mice. Single-cell
suspension of erythrocytes was prepared. Then, 2×105
erythrocytes were incubated with 1 ml red blood cell lysing
solution for 20 min, and centrifuged at 2,000 × g and 4°C for 10
min. Subsequently, the supernatants were used as erythrocyte
lysates.
Cell transfection
The miR-222 mimics, inhibitors, and corresponding
negative control (NC) were all purchased from Shanghai GenePharma
Co., Ltd. The sequence of the miR-222 mimics was
5′-AGCUACAUCUGGCUACUGGGU-3′, the corresponding NC was
5′-UUCUCCGAACGUGUGUCACGUTT-3′, the miR-222 inhibitor was
5′-ACCCAGUAGCCAGAUGUAGCU-3′ and the corresponding NC was
5′-UCUACUCUUUCUAGGAGGUUGUGA-3′. Cells were seeded into 6-well
plates and cultured for 24 h, and then transfected with miR-222
mimics, miR-222 inhibitors or corresponding NC (20 µM) using a
Lipofectamine® 3000 Transfection kit (Invitrogen; Thermo
Fisher Scientific, Inc.). After 48 h, the cells were used for
further assays.
Reverse transcription-quantitative PCR
(RT-qPCR)
For mRNA quantification, total RNA was isolated from
cells with TRIzol® (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocols.
Amplification and quantification of cDNA were carried out with SYBR
Green ROX Mix (ABgene). RT-qPCR was performed using SYBR Premix Ex
Taq™ (Takara Bio, Inc.) on a LightCycler 480 system (Roche
Diagnostics), using the following cycling conditions: 95°C for 10
min, 95°C for 10 sec, 60°C for 20 sec, 72°C for 10 sec, 40-sec
cycles. GAPDH and U6 were used as internal controls to normalize
the expression levels of mRNA and miRNAs (19), respectively. The sequences for the
primers used were as follows: miR-222, 5′-AGCTACATCTGGCTACTGGGT-3′;
U6, 5′-CAAGGATGACACGCAAATTCG-3′; GAPDH forward,
5′-GGTCACCAGGGCTGCTTTTA-3′ and reverse, 5′-GAGGGATCTCGCTCCTGGA-3′.
The primers used were purchased from Generay Biotech. The relative
expression was calculated by the 2−ΔΔCq (20).
Cell Counting Kit-8 (CCK-8) assay
Cell viability of microglia was assessed using a
CCK-8 assay. Cells (1×104) were seeded in 96-well
plates. After being cultured for 24 h, 10 µl erythrocyte lysates or
PBS was added the cell culture medium. After 48 h, the culture
supernatant was removed and the cells were incubated with 10 µl
CCK-8 (Dojindo Molecular Technologies, Inc.) reagent for 4 h at
37°C, according to the manufacturer's protocol. Then, the
absorbance was detected at a wavelength of 490 nm on a microplate
reader (Multiskan MK3; Thermo Fisher Scientific, Inc.).
Cell apoptosis
Apoptosis was evaluated using an apoptosis and
necrosis assay kit (Oncogene Research Products). Briefly, cells
were suspended in binding buffer and incubated at room temperature
in the dark for 15 min. Annexin-V-fluorescein isothiocyanate and
propidium iodide were added to cell suspension for 15 min in the
dark at room temperature according to the manufacturer's protocol.
Then, stained cells were analyzed using a FC500 flow cytometer
equipped with Cell Quest 3.0 software (BD Biosciences).
Western blotting
Briefly, total protein was extracted using RIPA
lysis buffer (Beyotime Institute of Biotechnology) from cells or
tissues and quantified using a commercial bicinchoninic acid kit
(BCA protein Assay kit; Pierce; Thermo Fisher Scientific, Inc.).
Protein (30 µg) was separated by SDS-PAGE on 12% gels and
transferred onto PVDF membranes. Then, the membrane was blocked
with 5% non-fat milk and incubated with primary antibodies
overnight at 4°C, including cleaved caspase-3 (cat. no. 9661;
1:1,000), cleaved caspase-9 (cat. no. 9505; 1:1,000), Bcl-2 (cat.
no. 2872; 1:1,000), Bax (cat. no. 2774; 1:1,000) and GAPDH (cat.
no. 8884; 1:2,000; all from Cell Signaling Technology, Inc.). The
blots were incubated for 2 h at room temperature with horseradish
peroxidase-conjugated secondary antibodies (cat. no. 7076; 1:2,000;
Cell Signaling Technology, Inc.). The signals were detected with an
ECL system (Beyotime Institute of Biotechnology) and analyzed using
the Quantity One software 4.6.6 (Bio-Rad Laboratories, Inc.).
ELISA
The levels of inflammatory factors in cell
supernatants were measured by ELISA according to the manufacturer.
Microglia were plated in 24-well plates and stimulated with
erythrocyte lysates for 48 h, and then culture supernatant was
added and stored at −80°C until analyzed by ELISA. The levels of
IL-6 (cat. no. BMS213HS), TNF-α (cat. no. BMS223HS), MCP-1 (cat.
no. BMS281INST) and IL-1β (cat. no. BMS224-2) were measured using
an ELISA kit (eBioscience; Thermo Fisher Scientific, Inc.).
ICH model
Mice were anesthetized with an intraperitoneal
injection of 350 mg/kg chloral hydrate and fixed on a mouse
stereotaxic frame (Alcott Biotech Co., Ltd.). A 20-µl volume of
autologous non-anticoagulated blood was collected from the tail
vein of the mouse and then implanted into the caudate nucleus at 2
µl/min at the following coordinates relative to bregma: 0.8 mm
anterior, 2 mm left lateral and 3.5 mm deep. Then the needle was
pulled out without blood reflux after 5 min duration and the wound
was sutured. The body temperature remained at 37°C during the
process, and the mice could drink freely after waking up. Only the
mice observed with neurological deficits were regarded as
successful models.
Intracerebroventricular injection
As previously reported (14), the stereotaxic coordinates were 0.5
mm posterior and 1.0 mm lateral to bregma and 2.5–3.0 mm ventral to
the surface of the skull. The miR-222 mimics or miR-222 inhibitor
(1.5 µl, 1 µg/1 µl) were added to Entranster™ in vivo
transfection reagent. The solution was injected
intracerebroventricularly by a micro syringe (Hamilton Company)
under the guidance of the stereotaxic instrument (RWD Life
Science).
Evaluation of neurological scores
The neurological deficits were determined by
neurological severity scores, a composite of motor, sensory,
reflex, and balance tests according to a previous study (21). Neurological function was graded on
a scale of 1 to 18; a score of 1 point was awarded for the
inability to perform the test or for the lack of a tested reflex.
Scoring was conducted by 2 trained investigators, and the mean
score of the subscales was the final score of each mouse.
Brain water content
The mice were decapitated and the brains were
removed at 72 h after ICH. The brain samples were immediately
weighed on an electronic analytical balance to obtain wet weight.
The brain was dried for 24 h at 100°C by an Electric Blast Drying
Oven to obtain dry weight. The water content of the brain (%) was
calculated as follows=(Wet weight-Dry weight)/Wet weight ×100.
Dual luciferase 3′-UTR reporter
assay
The program TargetScan (www.targetscan.org) was used to predict the target of
miR-222. The wild-type and mutant integrin subunit β8 (ITGB8)
3′-UTR dual-luciferase reporter vectors were constructed by
subcloning the human ITGB8 mRNA 3′-UTR and mutant 3′-UTR sequences
into the pGL3 Dual-Luciferase Reporter Vectors (Promega
Corporation). Cells were transfected with 80 ng luciferase reporter
vectors and miR-222 mimics using the Lipofectamine 3000
(Invitrogen; Thermo Fisher Scientific, Inc.). After 24 h,
luciferase activities were assessed using Dual-Luciferase Reporter
System (Berthold Detection Systems GmbH) according to the
manufacturer's instructions.
Statistical analysis
All experiments were performed at least three times.
Data are presented as the mean ± standard deviation and analyzed by
GraphPad Prism 5.0 (GraphPad Software) and SPSS 18 software (SPSS,
Inc.). The differences between the two groups were calculated by
Student's t-test, and the differences among multiple groups were
calculated by one-way ANOVA followed by Scheffe test. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-222 is upregulated in erythrocyte
lysate-induced microglia
ICH can promote microglia activation and release
proinflammatory mediators, which can cause neuronal injury
(22). Based on research,
microglia cells were selected in response to erythrocyte lysis as a
research object in vitro. For primary microglia cells, an
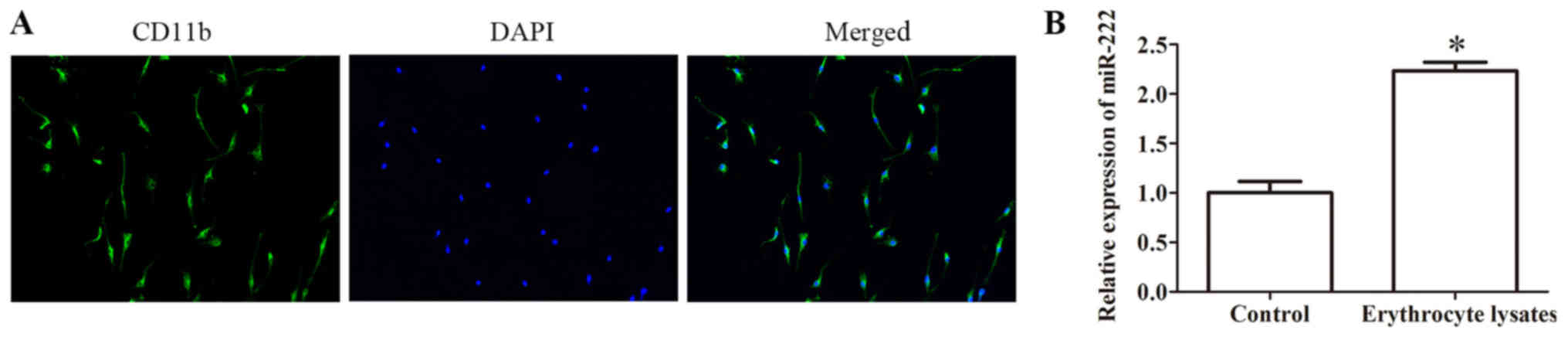
immunofluorescence assay was conducted to identify
microglia-specific antibody CD11b (Fig. 1A). Furthermore, the result of
qRT-PCR revealed that the expression of miR-222 was significantly
increased in the erythrocyte lysate-induced group compared with the
control group, indicating that the expression of miR-222 may be
related to ICH incidence (Fig.
1B).
miR-222 regulates erythrocyte
lysate-induced microglia cell viability and apoptosis
To further investigate the role of miR-222 on
microglia cell viability and apoptosis in vitro, miR-222
mimics or miR-222 inhibitors were transfected into microglia cells
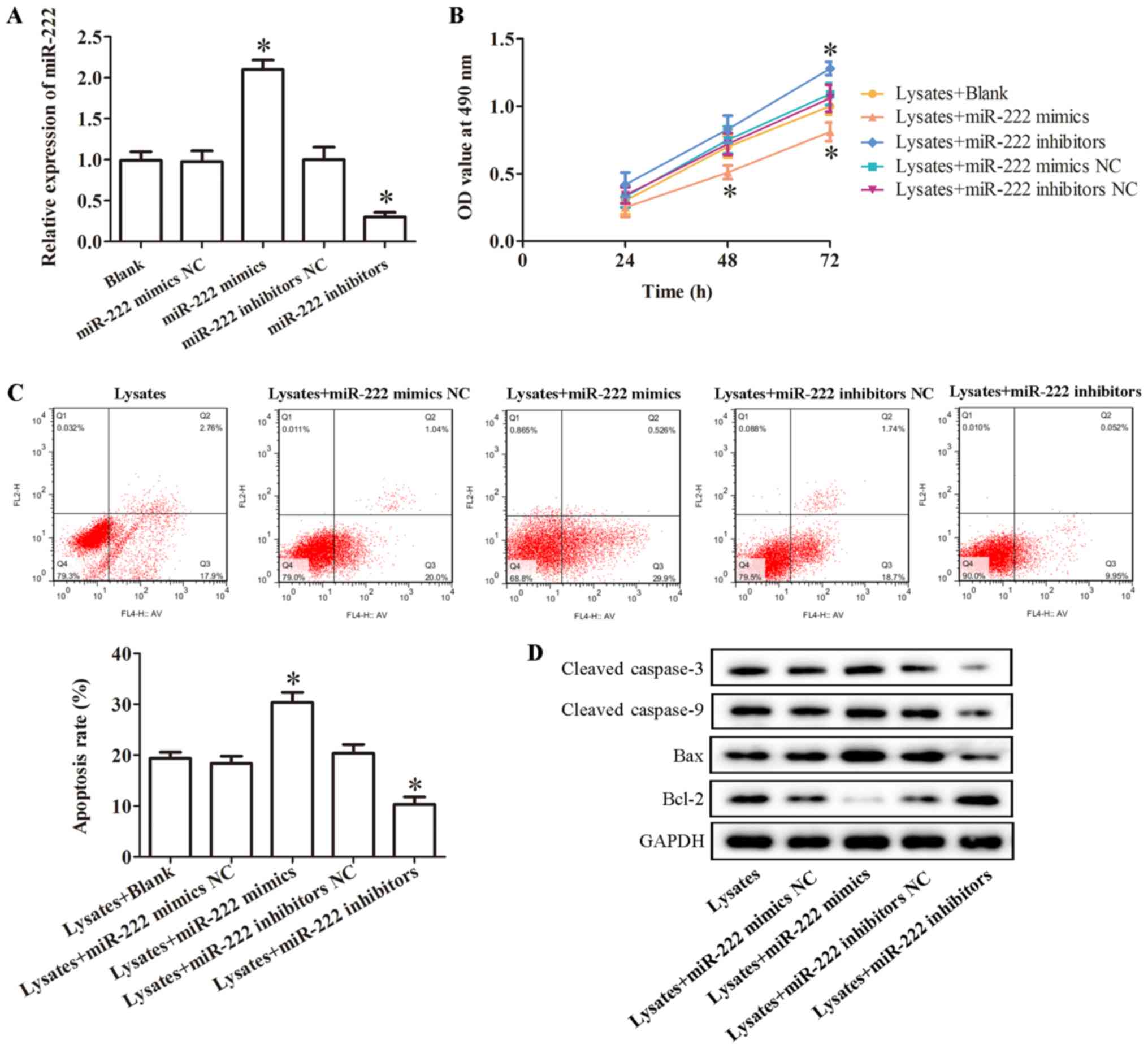
to upregulate or knockdown the level of miR-222. The result of
qRT-PCR revealed miR-222 mimics significantly promoted the
expression of miR-222, while miR-222 inhibitors significantly
inhibited it (Fig. 2A).
Furthermore, a CCK-8 assay was conducted to evaluate
microglia cell viability. As revealed in Fig. 2B, overexpression of miR-222
significantly suppressed erythrocyte lysate-induced microglia cell
viability, while inhibition of miR-222 significantly promoted
erythrocyte lysate-induced microglia cell viability (Fig. 2B). In addition, the effect of
miR-222 on cell apoptosis was evaluated by flow cytometric
analysis. The result demonstrated that the percentage of
erythrocyte lysate-induced apoptotic cells was statistically
increased by overexpression of miR-222, while miR-222 inhibitors
significantly decreased erythrocyte lysate-induced cell apoptosis
(Fig. 2C). In addition, the levels
of apoptosis-related proteins, including cleaved caspase-3, cleaved
caspase-9, Bax and Bcl-2, were evaluated by western blotting. The
result indicated that overexpression of miR-222 significantly
promoted the expression of cleaved caspase-3, cleaved caspase-9 and
Bax and decreased the level of Bcl-2 in erythrocyte lysate-induced
microglia cells, however, miR-222 inhibitors produced the opposite
effect (Fig. 2D).
miR-222 regulates inflammatory
response in erythrocyte lysate-induced microglia cells
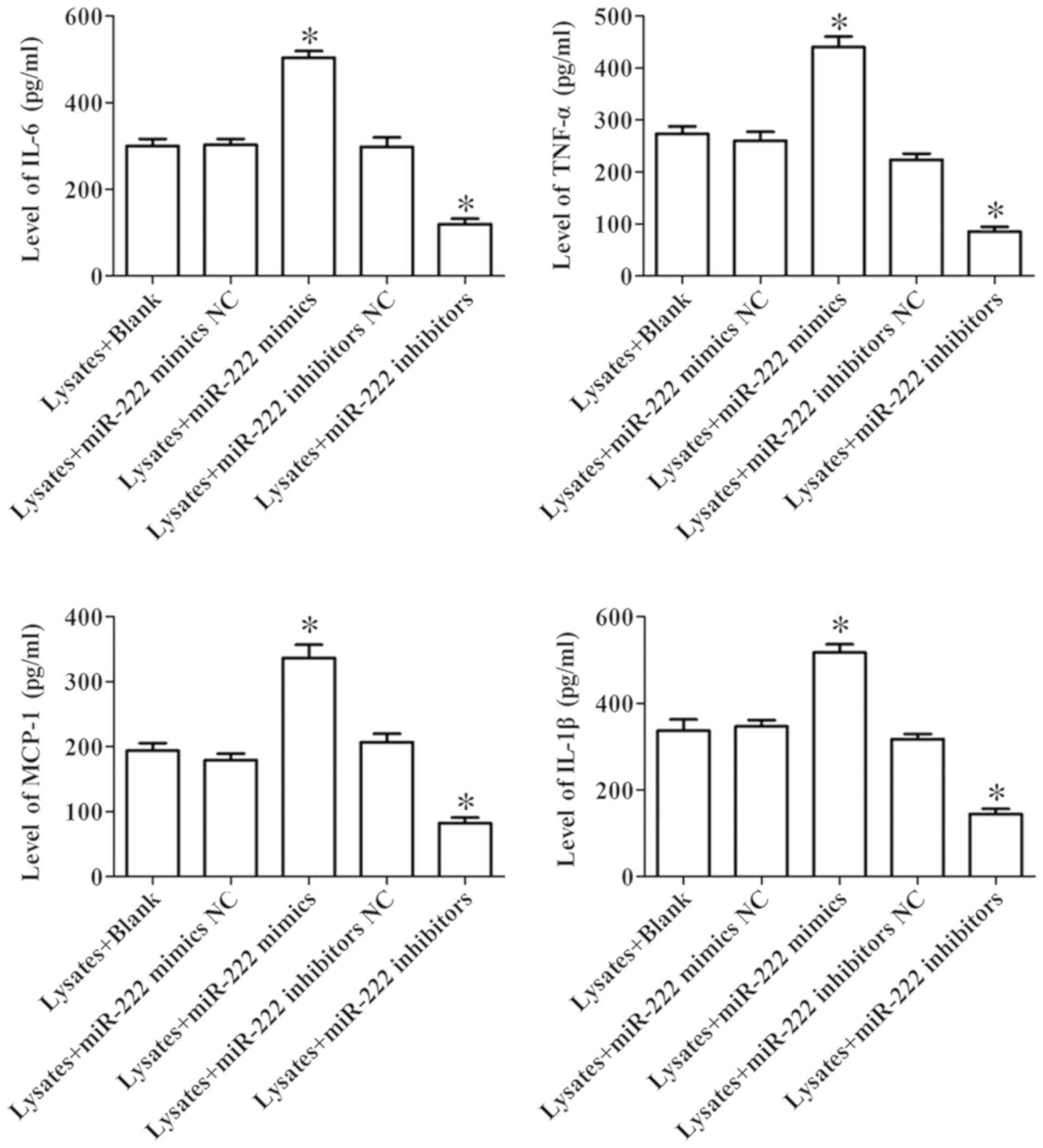
The expression of cytokines in microglia exposed to
erythrocyte lysates was assessed by ELISA. As revealed in Fig. 3, miR-222 mimics significantly
promoted the levels of IL-6, TNF-α, MCP-1 and IL-1β compared to the
NC group in microglia exposed to erythrocyte lysates, while
inhibition of miR-222 significantly suppressed the expression of
these cytokines. These results demonstrated that miR-222 could
regulate inflammatory response in erythrocyte lysate-induced
microglia cells.
miR-222 mediates brain damage in ICH
mice
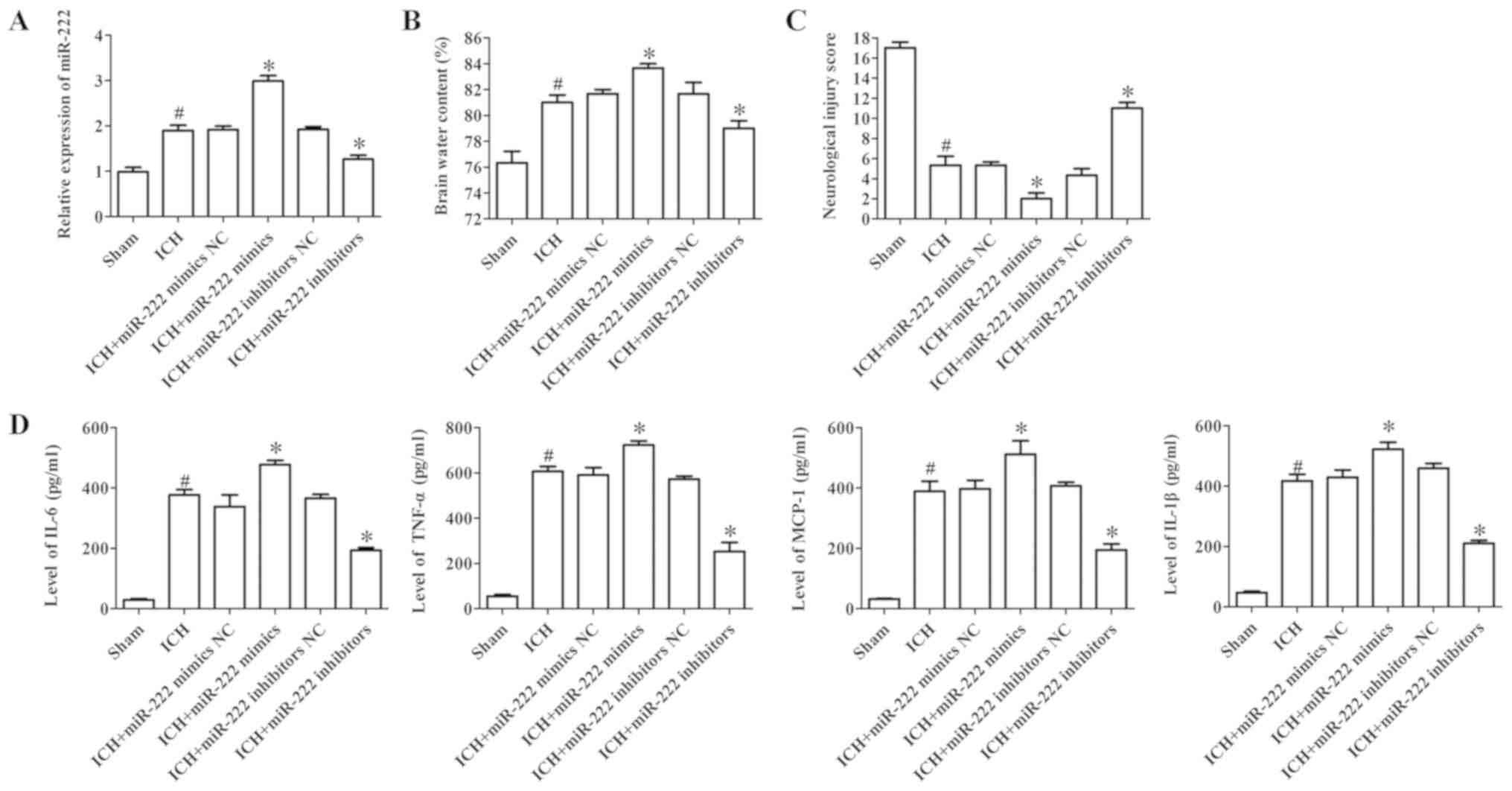
After being subjected to ICH for 72 h, miR-222
expression, water content in the brains of mice, the expression of
inflammatory cytokines and neurological injury were evaluated. The
results demonstrated that the expression of miR-222 subjected to
ICH significantly increased (Fig.
4A). In addition, the water content in the brains of mice and
neurological injury in the ICH group were significantly increased
when compared to the sham group, while inhibition of miR-222
clearly decreased the water content and neurological injury
(Fig. 4B and C). To determine the
contribution of miR-222 to inflammation, ELISA was performed to
assess the levels of inflammatory factors in brain tissues,
including IL-6, TNF-α, MCP-1 and IL-1β. The results demonstrated
that the expression of IL-6, TNF-α, MCP-1 and IL-1β were
significantly increased in the ICH group compared to the sham
group, while miR-222 inhibitors significantly decreased the
expression of these cytokines (Fig.
4D). These results revealed that inhibition of miR-222 could
ameliorate the neurological symptoms and improve brain function
after ICH.
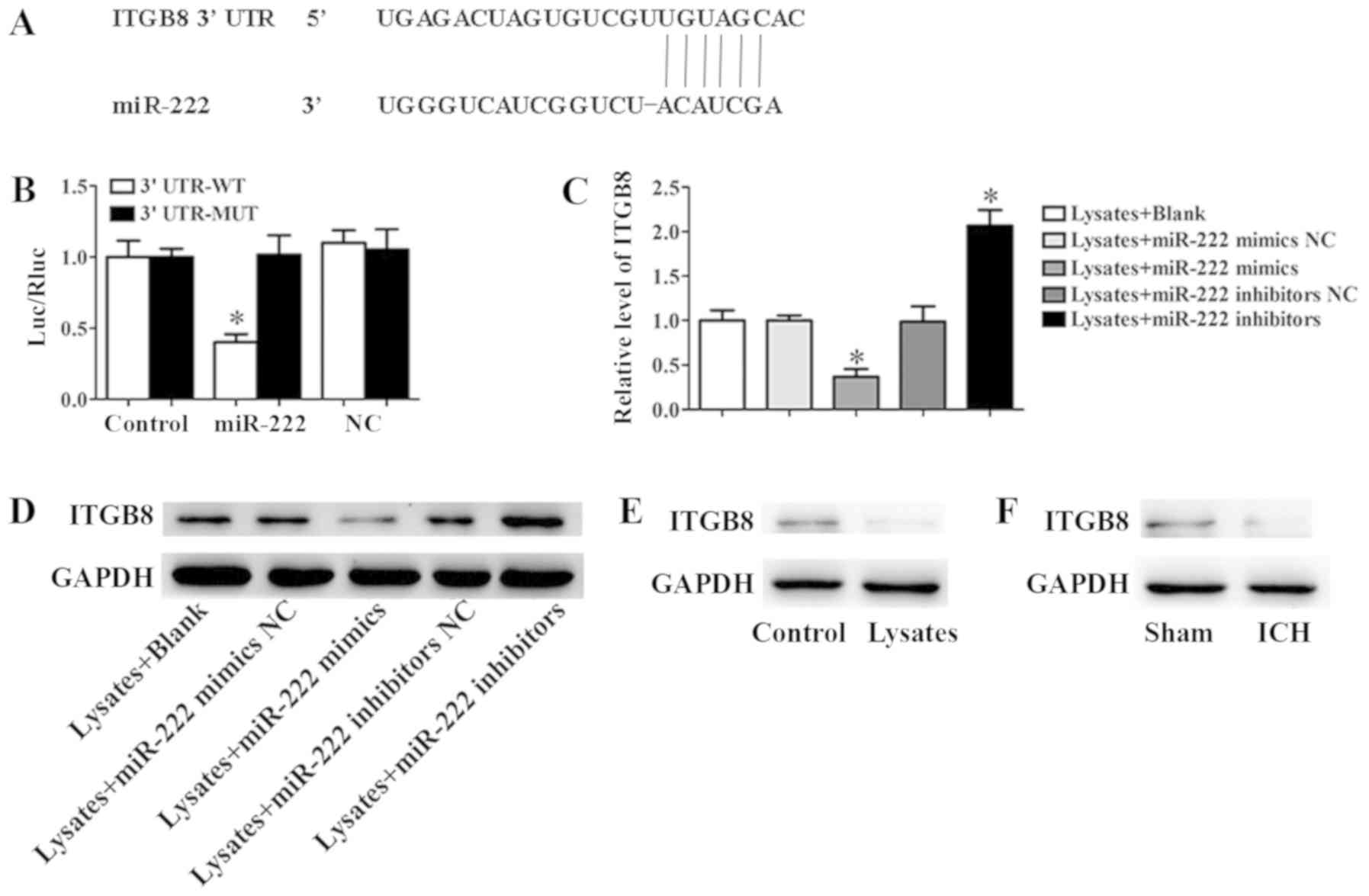
ITGB8 is a direct target of
miR-222
According to the target prediction program
TargetScan, the 3′-UTR of ITGB8 mRNA contains a putative miR-222
target sequence (Fig. 5A). To
demonstrate that ITGB8 was a direct target of miR-222 in microglia,
a dual-luciferase reporter system was performed. The result
revealed that co-expression with miR-222 mimics significantly
inhibited the activity of a firefly luciferase reporter containing
wild-type ITGB8 3′-UTR, while this was not detected on a reporter
with a mutated ITGB8 3′-UTR (Fig.
5B). In addition, the results of qRT-PCR and western blot assay
revealed that miR-222 mimics significantly inhibited the expression
of ITGB8, however, inhibition of miR-222 could significantly
promote this expression (Fig. 5C and
D). In addition, erythrocyte lysates significantly inhibited
the expression of ITGB8 in microglia cells (Fig. 5E). Moreover, in the ICH mice, the
expression of ITGB8 was markedly downregulated compared to the sham
group (Fig. 5F).
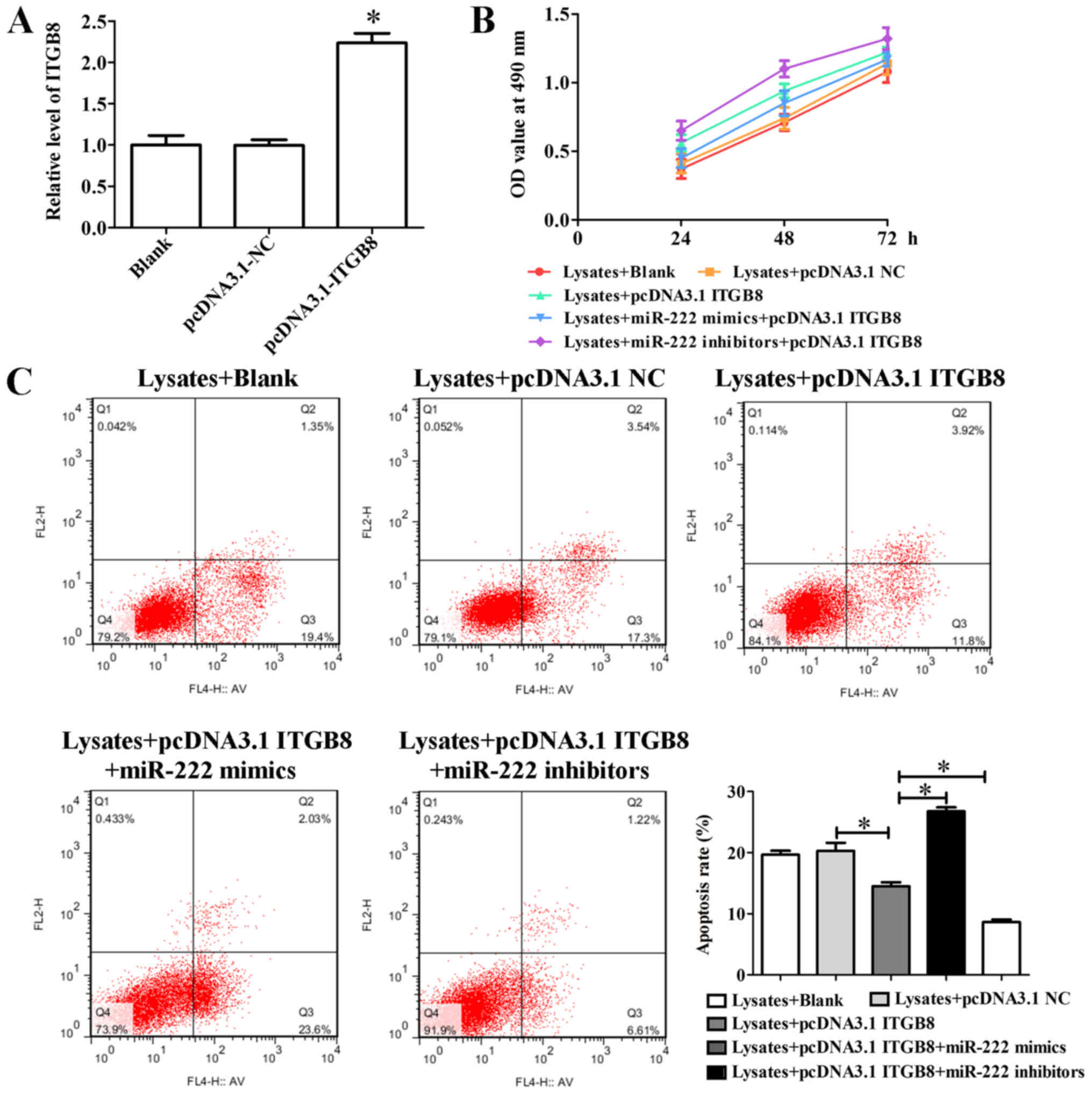
Inhibition of miR-222 promotes
erythrocyte lysate-induced microglia cell viability, reduces
apoptosis and inflammatory response by targeting ITGB8
To identify the role of ITGB8 in the
miR-222-mediated inflammatory response, ITGB8 expression was
promoted by pcDNA3.1. The result of qRT-PCR indicated that
pcDNA3.1-ITGB8 significantly promoted the expression of ITGB8,
while pcDNA3.1-NC did not increase this expression (Fig. 6A). Furthermore, a cell viability
assay revealed that pcDNA3.1-ITGB8 clearly promoted erythrocyte
lysate-induced microglia cell viability, and pcDNA3.1-ITGB8
relieved the decrease of cell viability of the miR-222 mimics
(Fig. 6B). In addition,
overexpression of ITGB8 significantly promoted a reduction of
erythrocyte lysate-induced cell apoptosis and co-transfection with
miR-222 mimics significantly inhibited the decrease of cell
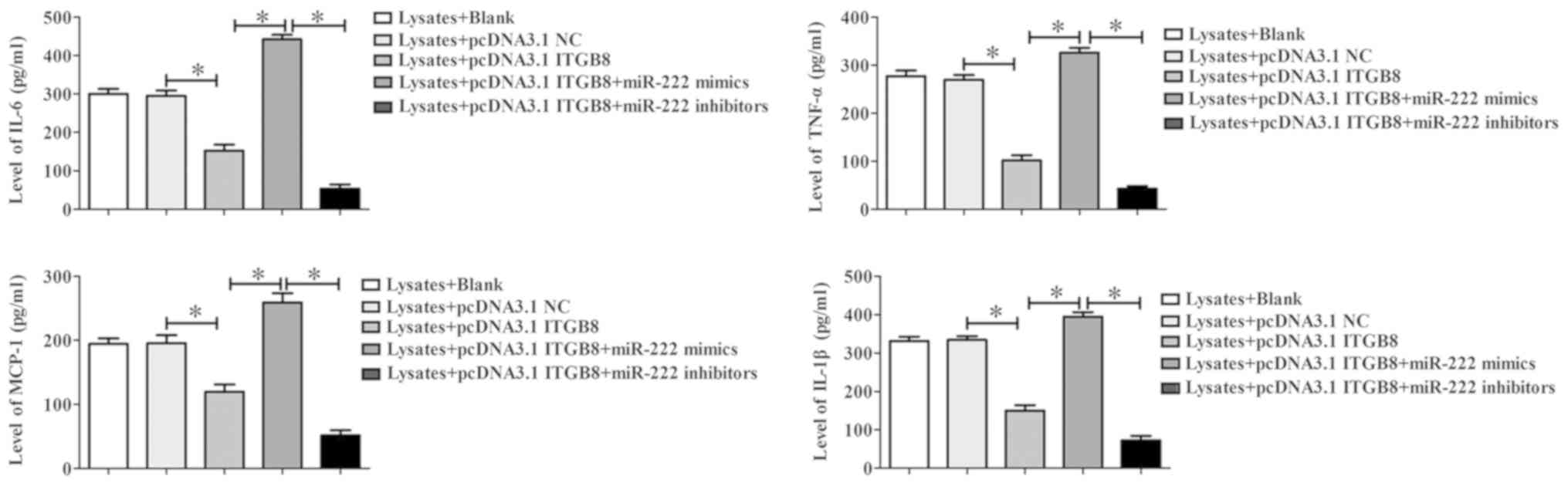
apoptosis cause by overexpression of ITGB8 (Fig. 6C). Furthermore, pcDNA3.1-ITGB8
significantly inhibited the levels of inflammatory factors
including IL-6, TNF-α, MCP-1 and IL-1β, and pcDNA3.1-ITGB8 could
suppress the increase in the levels of inflammatory factors caused
by miR-222 mimics (Fig. 7). These
results indicated that inhibition of miR-222 promoted erythrocyte
lysate-induced microglia cell viability, reduced apoptosis and
inflammatory response by targeting ITGB8.
Discussion
ICH, a common type of stroke with high mortality and
morbidity, is one of the leading causes of death worldwide
(23). Previous studies have
revealed the involvement of miR-222 in the regulation of
angiogenesis and inflammation (24,25).
Therefore, it was hypothesized that miR-222 may play a key role in
the occurrence and development of ICH. In the present study,
erythrocyte lysates could significantly promote the expression of
miR-222 in microglia. Inhibition of miRNA-222 promoted erythrocyte
lysate-induced microglia cell viability and decreased cell
apoptosis. In addition, in ICH mice, miR-222 inhibitors could
significantly decrease water content and neurological injury, as
well as reduce the expression of inflammatory factors. Furthermore,
ITGB8 was identified as a direct target of miR-222, and miR-222
could downregulate ITGB8 to inhibit inflammation, reduce brain
edema and improve neurological functions.
After ICH, microglia are greatly active, and their
reaction stimulated by hemorrhagic brain damage is extremely
complex, which is far beyond the nutrition of neurons and the scope
of denaturation and necrosis (26). Significant necrosis and apoptosis
are often accompanied by neuronal cells around the hemorrhagic
foci, and these necrotic lesions gather around a large number of
activated microglia and can cause the release of a series of
cytokines including NO, TNF and IL-6 (27). Therefore, the role of its mediated
inflammatory response in the progression of ICH has been gradually
recognized. Qureshi et al (28) revealed that there were 10 cases of
apoptosis in tissue specimens in the perihematoma region in 12
patients with ICH. Apoptosis was observed in the specimens obtained
at days 1, 2 and 5 after the onset of symptoms. The average
percentage of apoptotic cells around the hematoma was 38%, while
the average percentage of necrotic cells was only 25%, indicating
that apoptosis was the main form of cell death around the hematoma.
In in vitro experiments, it was revealed that the production
of NO in activated microglia significantly increased and the
inducible NO synthase notably increased in the cells. Concurrently,
NO mediated mitochondrial DNA damage and led to apoptosis (29). In consistence with these findings,
our results revealed that erythrocyte lysates could significantly
suppress cell viability and induce cell apoptosis in microglia, and
promote the expression of cytokines. In addition, erythrocyte
lysates could significantly promote the expression of miR-222 in
microglia. Furthermore, it was revealed that inhibition of
miRNA-222 could reverse erythrocyte lysate-induced damage in
microglia.
The formation of cerebral edema after intracerebral
hemorrhage is the most important pathological change of
intracerebral hemorrhage, which is also the key factor leading to
the deterioration of clinical cerebral hemorrhage (30). Clinical studies have revealed that
the peak of death from cerebral hemorrhage occurs in the first few
days after the onset of symptoms, which may be related to
progressive brain edema formation (31). Cerebral edema develops rapidly
after intracerebral hemorrhage, occurs within 1–2 h after
hemorrhage, progressively aggravates, peaks at 24 h, and begins to
be absorbed for 4–5 days (32).
The study of the mechanism of secondary cerebral edema after
intracerebral hemorrhage can help the clinical treatment of
cerebral hemorrhage patients, reduce secondary injury around the
hematoma, reduce the mortality and disability rate of cerebral
hemorrhage, and improve the quality of life of the patients
(33). In the present study, the
results revealed that miR-222 inhibitors could significantly
decrease the water content and neurological injury, as well as
reduce the expression of inflammatory factors in ICH mice.
ITGB8 is an important member of the integrin family,
members of which are mediators of the interactions between cells
and the matrix (34). Previous
studies have revealed that ITGB8 in perivascular astrocytes plays
an important role in regulating brain vessel homeostasis through
modulation of TGF-β activation and expression of TGF-β-responsive
genes that promote vessel differentiation and stabilization
(35). In addition, Ma et
al (36) revealed that ITGB8
deficiency enhanced the formation of dysplastic vessels and
hemorrhage. Herein, ITGB8 was identified as a direct target of
miR-222, and miR-222 could downregulate ITGB8 to inhibit
inflammation, increase cell viability and decrease cell apoptosis
in erythrocyte lysate-induced microglia.
In conclusion, it was revealed in the present study,
that miR-222 suppressed ITGB8 production by directly binding to its
3′-UTR, which reduced neuronal inflammation and improved neuronal
function.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YYB and JZN performed the experiments and analyzed
the data. JZN was a major contributor in writing the manuscript.
YYB and JZN both read and approved the final manuscript and agree
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Experiments were performed according to animal care
guidelines approved by The Animal Ethics Committee of Nanjing
Medical University, and animals were treated in accordance with The
Guidelines of the United States National Institutes of Health.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang MD, Wang Y, Xia YP, Dai JW, Gao L,
Wang SQ, Wang HJ, Mao L, Li M, Yu SM, et al: High serum MiR-130a
levels are associated with severe perihematomal edema and predict
adverse outcome in acute ICH. Mol Neurobiol. 53:1310–1321. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Min H, Jang YH, Cho IH, Yu SW and Lee SJ:
Alternatively activated braininfiltrating macrophages facilitate
recovery from collagenase-induced intracerebral hemorrhage. Mol
Brain. 9:422016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rodríguez JA, Sobrino T, López-Arias E,
Ugarte A, Sánchez-Arias JA, Vieites-Prado A, de Miguel I, Oyarzabal
J, Páramo JA, Campos F, et al: CM352 reduces brain damage and
improves functional recovery in a rat model of intracerebral
hemorrhage. J Am Heart Assoc. 6(pii): e0060422017.PubMed/NCBI
|
|
4
|
Keep RF, Hua Y and Xi G: Intracerebral
haemorrhage: Mechanisms of injury and therapeutic targets. Lancet
Neurol. 11:720–731. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sheth KN and Rosand J: Targeting the
immune system in intracerebral hemorrhage. JAMA Neurol.
71:1083–1084. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu H, Zhang Z, Li Y, Zhao R, Li H, Song Y,
Qi J and Wang J: Time course of upregulation of inflammatory
mediators in the hemorrhagic brain in rats: Correlation with brain
edema. Neurochem Int. 57:248–253. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang Z, Zhang Z, Lu H, Yang Q, Wu H and
Wang J: Microglial polarization and inflammatory mediators after
intracerebral hemorrhage. Mol Neurobiol. 54:1874–1886. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhao X, Wu T, Chang CF, Wu H, Han X, Li Q,
Gao Y, Li Q, Hou Z, Maruyama T, Zhang J and Wang J: Toxic role of
prostaglandin E2 receptor EP1 after intracerebral hemorrhage in
mice. Brain Behav Immun. 46:293–310. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ge Y, Yan X, Jin Y, Yang X, Yu X, Zhou L,
Han S, Yuan Q and Yang M: fMiRNA-192 and miRNA-204 directly
suppress lncRNA HOTTIP and interrupt GLS1-Mediated glutaminolysis
in hepatocellular carcinoma. PLoS Genet. 11:e10057262015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Martignani E, Miretti S, Accornero P and
Baratta M: miRNAs highlights in stemand cancer cells. Mini Rev Med
Chem. 11:1165–1182. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Veremeyko T, Siddiqui S, Sotnikov I, Yung
A and Ponomarev ED: IL-4/IL-13-dependent and independent expression
of miR-124 and itscontribution to M2 phenotype of monocytic cells
in normal conditions andduring allergic inflammation. PLoS One.
8:e817742013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Moore CS, Rao VT, Durafourt BA, Bedell BJ,
Ludwin SK, Bar-Or A and Antel JP: miR-155 as a multiple
sclerosis-relevant regulator of myeloid cellpolarization. Ann
Neurol. 74:709–720. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xi T, Jin F, Zhu Y, Wang J, Tang L, Wang
Y, Liebeskind DS and He Z: MicroRNA-126-3p attenuates blood-brain
barrier disruption, cerebral edema and neuronal injury following
intracerebral hemorrhage by regulating PIK3R2 and Akt. Biochem
Biophys Res Commun. 494:144–151. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu A, Zhang T, Duan H, Pan Y, Zhang X,
Yang G, Wang J, Deng Y and Yang Z: MiR-124 contributes to M2
polarization of microglia and confers brain inflammatory protection
via the C/EBP-α pathway in intracerebral hemorrhage. Immunol Lett.
182:1–11. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Galardi S, Mercatelli N, Giorda E,
Massalini S, Frajese GV, Ciafre SA and Farace MG: miR-221 and
miR-222 expression affects the proliferation potential of human
prostate carcinoma cell lines by targeting p27Kip1. J Biol Chem.
282:23716–23724. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun T, Wang X, He HH, Sweeney CJ, Liu SX,
Brown M, Balk S, Lee GS and Kantoff PW: miR-221 and miR-222
expression affects the proliferation potential of human prostate
carcinoma cell lines by targeting p27Kip1. Oncogene. 33:2790–2800.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou Z, Zhou L, Jiang F, Zeng B, Wei C,
Zhao W and Yu D: Downregulation of miR-222 induces apoptosis and
cellular migration in adenoid cystic carcinoma cells. Oncol Res.
25:207–214. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kinoshita K, Matsumoto K, Kurauchi Y,
Hisatsune A, Seki T and Katsuki H: A Nurr1 agonist amodiaquine
attenuates inflammatory events and neurological deficits in a mouse
model of intracerebral hemorrhage. J Neuroimmunol. 330:48–54. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen D, Wu X, Zhao J and Zhao X:
MicroRNA-634 functions as a tumor suppressor in pancreatic cancer
via directly targeting heat shock-related 70-kDa protein 2. Exp
Ther Med. 17:3949–3956. 2019.PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu A, Duan H, Zhang T, Pan Y, Kou Z, Zhang
X, Lu Y, Wang S and Yang Z: IL-17A promotes microglial activation
and neuroinflammation in mousemodels of intracerebral haemorrhage.
Mol Immunol. 73:151–157. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wasserman JK, Zhu X and Schlichter LC:
Evolution of the inflammatoryresponse in the brain following
intracerebral hemorrhage and effects of delayedminocycline
treatment. Brain Res. 1180:140–154. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xie WJ, Yu HQ, Zhang Y, Liu Q and Meng HM:
CD163 promotes hematoma absorption and improves neurological
functions in patients with intracerebral hemorrhage. Neural Regen
Res. 11:1122–1127. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Corsten MF, Heggermont W, Papageorgiou AP,
Deckx S, Tijsma A, Verhesen W, van Leeuwen R, Carai P, Thibaut HJ,
Custers K, et al: The microRNA-221/-222 cluster balances the
antiviral and inflammatory response in viral myocarditis. Eur Heart
J. 36:2909–2919. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ribeiro-Rodrigues TM, Laundos TL,
Pereira-Carvalho R, Batista-Almeida D, Pereira R, Coelho-Santos V,
Silva AP, Fernandes R, Zuzarte M, Enguita FJ, et al: Exosomes
secreted by cardiomyocytes subjected to ischaemia promote cardiac
angiogenesis. Cardiovasc Res. 1113:1338–1350. 2017. View Article : Google Scholar
|
|
26
|
Zhao X, Sun G, Ting SM, Song S, Zhang J,
Edwards NJ and Aronowski J: Cleaning up after ICH: The role of Nrf2
in modulating microglia function and hematoma clearance. J
Neurochem. 133:144–152. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang Z, Zhao T, Zou Y, Zhang JH and Feng
H: Curcumin inhibits microglia inflammation and confers
neuroprotection in intracerebral hemorrhage. Immunol Lett.
160:89–95. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Qureshi AI, Suri MF, Ostrow PT, Kim SH,
Ali Z, Shatla AA, Guterman LR and Hopkins LN: Apoptosis as a form
of cell death in intracerebral hemorrhage. Neurosurgery.
52:1041–1047. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Beufler B: Innate immunity: An overview.
Mol Immunol. 40:845–859. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gebel JM, Brott TG, Sila CA, Tomsick TA,
Jauch E, Salisbury S, Khoury J, Miller R, Pancioli A, Duldner JE,
et al: Decreased perihematomal edema in thrombolysis-related
intracerebral hemorrhage compared with spontaneous intrscerebral
hemorrhage. Stroke. 31:596–600. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xi G, Wagner KR, Keep RF, Hua Y, de
Courten-Myers GM, Broderick JP, Brott TG and Hoff JT: Role of blood
clot formation on early edema development after experimental
intracerbral hemorrhage. Stroke. 29:2580–2586. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wei P, You C, Jin H, Chen H and Lin B:
Correlation between serum IL-1beta levels and cerebral edema extent
in a hypertensive intracerebral hemorrhage rat model. Neurol Res.
36:170–175. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hu YL, Wang H, Huang Q, Wang G and Zhang
HB: MicroRNA-23a-3p promotes the perihematomal edema formation
after intracerebral hemorrhagevia ZO-1. Eur Rev Med Pharmacol Sci.
22:2809–2816. 2018.PubMed/NCBI
|
|
34
|
Kumar V, Soni UK, Maurya VK, Singh K and
Jha RK: Integrin beta8 (ITGB8) activates VAV-RAC1 signaling via FAK
in the acquisition of endometrial epithelial cell receptivity for
blastocyst implantation. Sci Rep. 7:18852017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cambier S, Gline S, Mu D, Collins R, Araya
J, Dolganov G, Einheber S, Boudreau N and Nishimura SL: Integrin
alpha(v)beta8-mediated activation of transforming growth
factor-beta by perivascular astrocytes: An angiogenic control
switch. Am J Pathol. 166:1883–1894. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ma L, Shen F, Jun K, Bao C, Kuo R, Young
WL, Nishimura SL and Su H: Integrin β8 deletion enhances vascular
dysplasia and hemorrhage in thebrain of adult alk1 heterozygous
mice. Transl Stroke Res. 7:488–496. 2016. View Article : Google Scholar : PubMed/NCBI
|