Introduction
Non-alcoholic fatty liver disease (NAFLD) is a type
of pathological fat accumulation in the liver and is a major health
condition in the world (1). NAFLD
includes a wide spectrum of conditions, ranging from non-alcoholic
fatty liver (NAFL) to non-alcoholic steatohepatitis (NASH)
(2,3). NAFL is characterized by an
accumulation of hepatocellular lipids, mainly triglycerides,
whereas NASH is identified by the addition of inflammation
(4). It is estimated that 10–20%
of patients with NAFL also develop NASH (5), and NASH is associated with a
>10-fold increased risk (2.8 vs. 0.2%) of liver-associated
mortality and twice the risk of cardiovascular disease (6). Therefore, the increasing prevalence
of NASH and the increased risk of cardiovascular disease and
liver-associated mortality have led to a demand for medical
therapy. However, no pharmacological therapy has been shown to be
effective for long-term use (7).
Tetramethylpyrazine (TMP) is a biologically active
alkaloid extracted from Ephedra sinica (8), which has been widely used in Chinese
herbal medicines for various purposes, including treating
cardiovascular and cerebrovascular defects, and anti-oxidation,
antifibrotic, anti-nociceptive, anti-inflammatory and
anti-neoplastic activities (9,10).
TMP can decrease arsenic-induced reactive oxygen species (ROS)
production, enhance glutathione peroxidase (GSH-pX) levels, prevent
mitochondrial dysfunction, and suppress the activation of
pro-inflammatory signals and the development of autophagy and
apoptosis (11). However, the
effect of TMP on the progression of NAFL to NASH remains to be
fully elucidated. The present study observed the protective effect
of TMP on NASH in mice, examined its therapeutic mechanism and
provided a theoretical basis for its clinical application.
Materials and methods
The present study was approved by The Research
Ethics Committee of the China Academy of Chinese Medical Sciences
(Beijing, China), in accordance with the National Institutes of
Health Guidelines for the Care and Use of Laboratory Animals
(National Institutes of Health, Bethesda, MD, USA). All animals
were treated in accordance with the guidelines and regulations for
the use and care of animals of the Center for Laboratory Animal
Care, China Academy of Chinese Medical Sciences.
Chemicals and reagents
TMP and simvastatin were purchased from Nanjing
Jingzhu Bio-technology Co., Ltd. (Nanjing, China). An Oil Red O
staining kit, 2′,7′-dichlorofluorescin diacetate (DCFH-DA) and
chloral hydrate were obtained from Sigma-Aldrich; Merck KGaA
(Darmstadt, Germany). Superoxide dismutase (SOD), GSH-px and
malondialdehyde (MDA) kits were purchased from Nanjing Jiancheng
Bioengineering Institute (Nanjing, China). Antibodies against tumor
necrosis factor-α (TNF-α, cat. no. 3707), interleukin-6 (IL-6, cat.
no. 12912), nuclear factor-κB (NF-κB, cat. no. 6956), and
phosphorylated (p)-NF-κB (cat. no. 13346) were acquired from Cell
Signaling Technology, Inc. (Danvers, MA, USA). Secondary antibodies
against rabbit (cat. no. 7074) and mouse (cat. no. 7076)
immunoglobulin G were obtained from Cell Signaling Technology, Inc.
An antibody against β-actin (cat. no. sc-81178) was acquired from
Santa Cruz Biotechnology, Inc. (Dallas, TX, USA).
Animals and establishment of an NAFL
model
The C57BL/J mice (n=60, male to female ratio 1:1)
were purchased from Beijing HFK Bioscience Co., Ltd. (Beijing,
China). These mice (aged 8 weeks old, body weight 22–24 g) were
bred in a specific pathogen-free laboratory (temperature, 20–24°C;
humidity, 40–70%) at the China Academy of Chinese Medical Sciences
Laboratory Animal Center (Beijing, China). The mice were acclimated
to the feed for 1 week prior to the initiation of experimental
intervention. In the present study, the murine NAFLD model was
established by feeding mice a high fat diet for 8 weeks. A
proportion of the mice (n=12) were fed a regular diet, whereas
others (n=48) were fed a high fat diet (60 kcal % fat; cat. no.
D12492; Research Diets, Xietong Organism Co., Ltd., Nanjing,
China). The standard used for establishment of the murine model for
NAFLD was the NAFLD activity score. Food and water were provided
ad libitum throughout the study. After 8 weeks, the mice fed
a high fat diet were divided into four groups: Model group (n=12);
4 mg/kg/day simvastatin-treated group (n=12); and 100 (n=12) and
200 (n=12) mg/kg/day TMP-treated groups. From 9 weeks, the mice
received either distilled water (control and model groups), 4
mg/kg/day simvastatin (simvastatin-treated group), or 100 or 200
mg/kg/day TMP (TMP-treated groups) by forced oral ingestion. On day
85, after 12 h of fasting, the mice were anesthetized with chloral
hydrate (500 mg/kg) by intraperitoneal injection and blood samples
were taken from the inferior vena cava. The survival of mice was
confirmed by monitoring their breathing and heartbeat.
Subsequently, the mice were sacrificed by cervical dislocation and
their livers were quickly removed.
Hematological examination
In order to detect the protective effects of TMP on
the liver, serum was collected. The serum levels of glucose,
alanine aminotransferase (ALT), aspartate aminotransferase, total
bilirubin, triglyceride (TG), total cholesterol (TC), low-density
lipoprotein-cholesterol (LDL-C) and high-density
lipoprotein-cholesterol were measured using standard protocols in
the clinical laboratory.
Histology and
immunohistochemistry
For histological analysis, liver tissues fixed with
4% buffered paraformaldehyde were embedded in paraffin, and
3-µm-thick sections were prepared. The sections were then stained
with hematoxylin and eosin. The sections of liver tissues were
evaluated using a light microscope (magnification, ×200; Olympus
Corporation, Tokyo, Japan) for the degree of steatosis (low to
medium-power evaluation of parenchymal involvement by steatosis
<5%, score 0; 5–33%, score 1; >33–66%, score 2; >66% score
3), lobular inflammation (overall assessment of all inflammatory
foci, no foci, score=0; <2 foci per ×200 magnification field,
score 1; 2–4 foci per ×200 magnification field, score 2; >4 foci
per ×200 magnification field, score 3) and hepatocellular
ballooning (none, score 0; few balloon cells, score 1; numerous
cells/prominent ballooning, score 2) by an experienced pathologist
blinded to the clinical data, according to Kleiner et al
(12). The unweighted sum of these
three variables was used to calculate the NAFLD activity score.
Samples with scores ≥5 were diagnosed with NASH (13). Immunohistochemical analyses were
performed using antibodies against TNF-α and IL-6 (rabbit antibody;
dilution 1:200) according to the protocol of our previous study
(14).
Oil Red O staining
For Oil Red O staining, frozen liver samples were
cut into 5-µm-thick sections and subsequently air-dried onto
slides. First, specimens were fixed in 4% paraformaldehyde, and
washed with running tap water for 1–10 min. The specimens were then
rinsed with 60% isopropanol, stained with freshly prepared Oil Red
O working solution for 15 min and rinsed with 60% isopropanol.
Finally, the nuclei were lightly stained by dipping the slides into
hematoxylin solution five times and rinsing with distilled water
for examination using light microscopy (magnification, ×200).
Antioxidant assay
At the end of the experiment, the mice were
anesthetized using chloral hydrate and sacrificed by cervical
dislocation. Subsequently, the livers were quickly harvested and
placed on ice. The antioxidant enzyme activities, including the
activities of SOD and GSH-px, and the MDA content were determined,
according to the protocol of our previous study (14).
Measurement of ROS of liver
tissue
The ROS level of the liver tissue was detected using
the redox-sensitive fluorescent dye DCFH-DA. Briefly, the frozen
liver samples were cut into 5-µm-thick sections and left to air dry
onto slides. First, the specimens were fixed in 4% paraformaldehyde
and washed three times with PBS. The specimens were incubated with
DCFH-DA (10 µmol/l), which was diluted with PBS at 37°C for 30 min,
and then washed five times with PBS. Images of the relative level
of fluorescent product were captured using a fluorescence
microscope connected to an imaging system (BX50-FLA; Olympus
Corporation). ImageJ 1.47i software (National Institutes of Health)
was used to analyze the mean fluorescence intensity of DCFH-DA,
which indirectly detected the level of ROS.
Western blotting
The protein expression levels of TNF-α, IL-6, NF-κB,
p-NF-κB and β-actin in the liver tissues obtained from rats in
different groups were detected by western blot analysis. The liver
tissues were incubated in lysis buffer supplied with a cocktail of
phosphatase inhibitors (Roche Diagnostics, Indianapolis, IN, USA).
Total protein was quantified using the bicinchoninic acid protein
assay kit (Pierce; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). The protocol and semiquantitative analysis were performed
according the protocol of our previous study (14). The following antibodies were used:
TNF-α, IL-6, NF-κB, p-NF-κB (rabbit antibody; dilution 1:1,000) and
β-actin (rabbit antibody; dilution 1:1,000). The secondary
antibodies used were the following: Horseradish
peroxidase-conjugated anti-rabbit immunoglobulin G (IgG; cat. no.
7074; dilution 1:5,000; Santa Cruz Biotechnology, Inc.) or
anti-mouse IgG (cat. no. 7076; dilution 1:5,000; Santa Cruz
Biotechnology, Inc.).
Statistical analysis
SPSS version 13.0 software for Windows (SPSS, Inc.,
Chicago, IL, USA) was used for statistical analysis. All values are
presented as the mean ± standard deviation of the mean for the
indicated number of measurements. For comparisons among multiple
groups, one-way analysis of variance followed by the Least
Significant Difference test was performed. P<0.05 was considered
to indicate a statistically significant difference.
Results
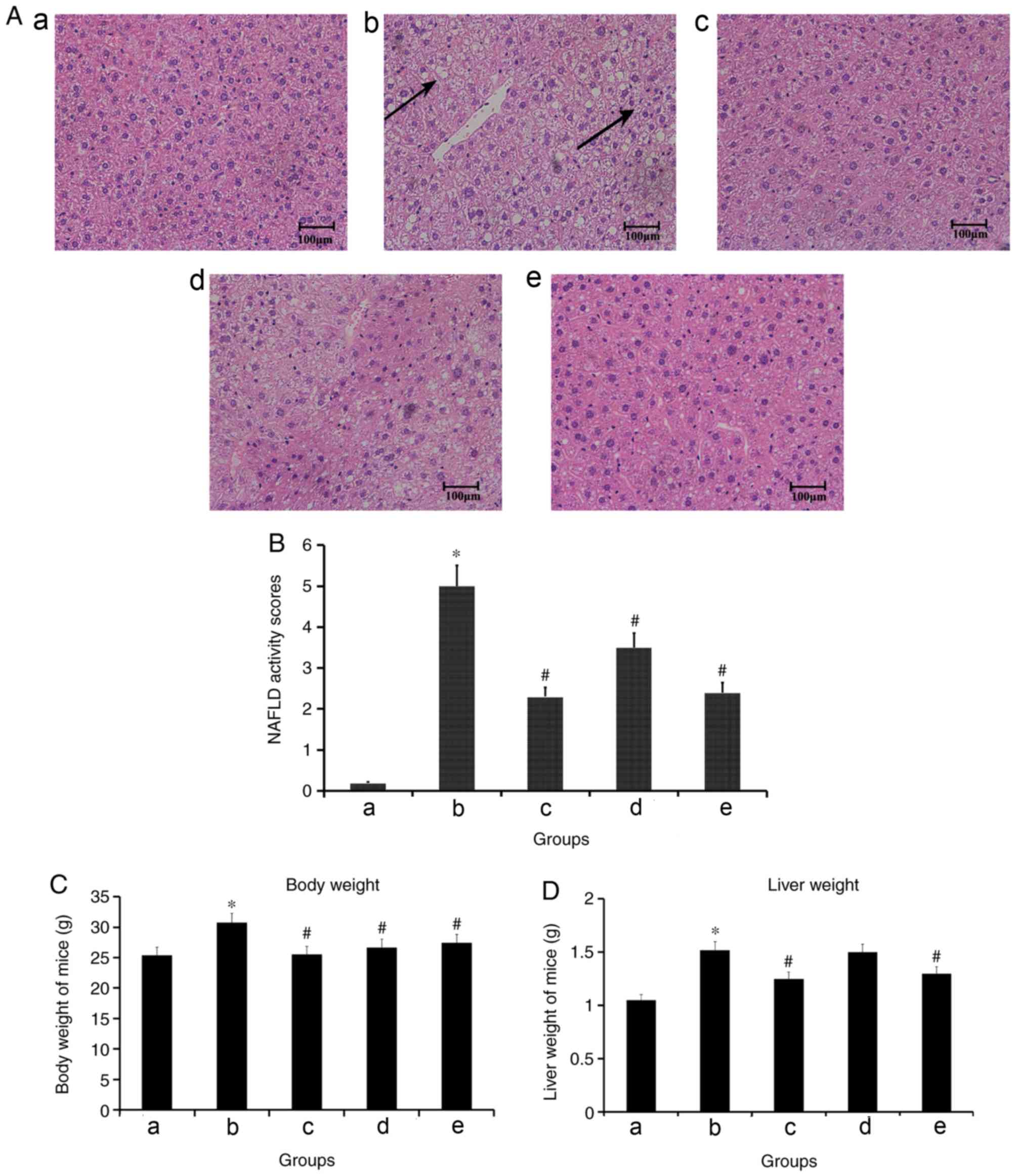
Effects of TMP treatment on liver
injury in a high fat diet-induced model of NASH
The administration of a high fat diet to C57BL/6
mice resulted in a classical pathophysiological model of NASH, with
microvesicular and macrovesicular steatosis, indicative of
disturbed lipid metabolism, and few foci of inflammatory cell
accumulations in the liver. Treatment with simvastatin and TMP
decreased the size and number of macrovesicular steatosis and
inflammation, compared with model mice (Fig. 1A). In addition, compared with the
control group, the NAFLD activity scores of the model group were
significantly increased. However, compared with the model group,
the NAFLD activity scores of 4 mg/kg/day simvastatin-treated, 100
mg/kg/day TMP-treated and 200 mg/kg/day TMP-treated groups were
significantly decreased (Fig. 1B).
Compared with the control group, the body weight and liver weight
of mice in the model group were significantly increased. However,
compared with the model group, the body weight and liver weight of
mice in the simvastatin-treated and 200 mg/kg/day TMP-treated
groups, and the body weight of mice in the 100 mg/kg/day
TMP-treated group, were significantly decreased (Fig. 1C and D).
TMP does not decrease the levels of
serum glucose, ALT, TG, TC or LDL-C
It was demonstrated that, compared with the control
group, the levels of blood glucose, ALT, TG, TC and LDL-C were
increased in the model group. However, compared with the model
group, 100 and 200 mg/kg/day TMP did not decrease the levels of
serum glucose, TC, TG and LDL-C, whereas the levels of ALT were
decreased. Furthermore, compared with the model group, simvastatin
did not decrease the levels of serum glucose, ALT, TG and LDL-C,
whereas the levels of TC were decreased (Table I).
 | Table I.Effect of TMP on levels of SG, TG, TC,
LDL-C and ALT (n=12). |
Table I.
Effect of TMP on levels of SG, TG, TC,
LDL-C and ALT (n=12).
| Group | Dose (mg/kg/day) | SG (mmol/l) | TG (mmol/l) | TC (mmol/l) | LDL-C (mmol/l) | ALT |
|---|
| Control | – | 8.9±1.3 | 0.54±0.07 | 1.91±0.20 | 0.20±0.05 | 39.5±7.9 |
| Model | – | 10.6±1.4a |
0.67±0.09a |
3.72±0.84a |
0.28±0.05a |
53.4±13.3a |
| Low-dose TMP | 100 | 10.4±1.6 | 0.62±0.17 | 3.82±0.99 | 0.30±0.05 | 47.3±6.6 |
| High-dose TMP | 200 | 9.3±1.3 | 0.58±0.04 | 3.76±0.66 | 0.27±0.05 |
40.8±10.2b |
| Simvastatin |
4 | 9.5±1.7 | 0.59±0.10 |
2.77±0.64b | 0.29±0.05 | 43.8±9.6 |
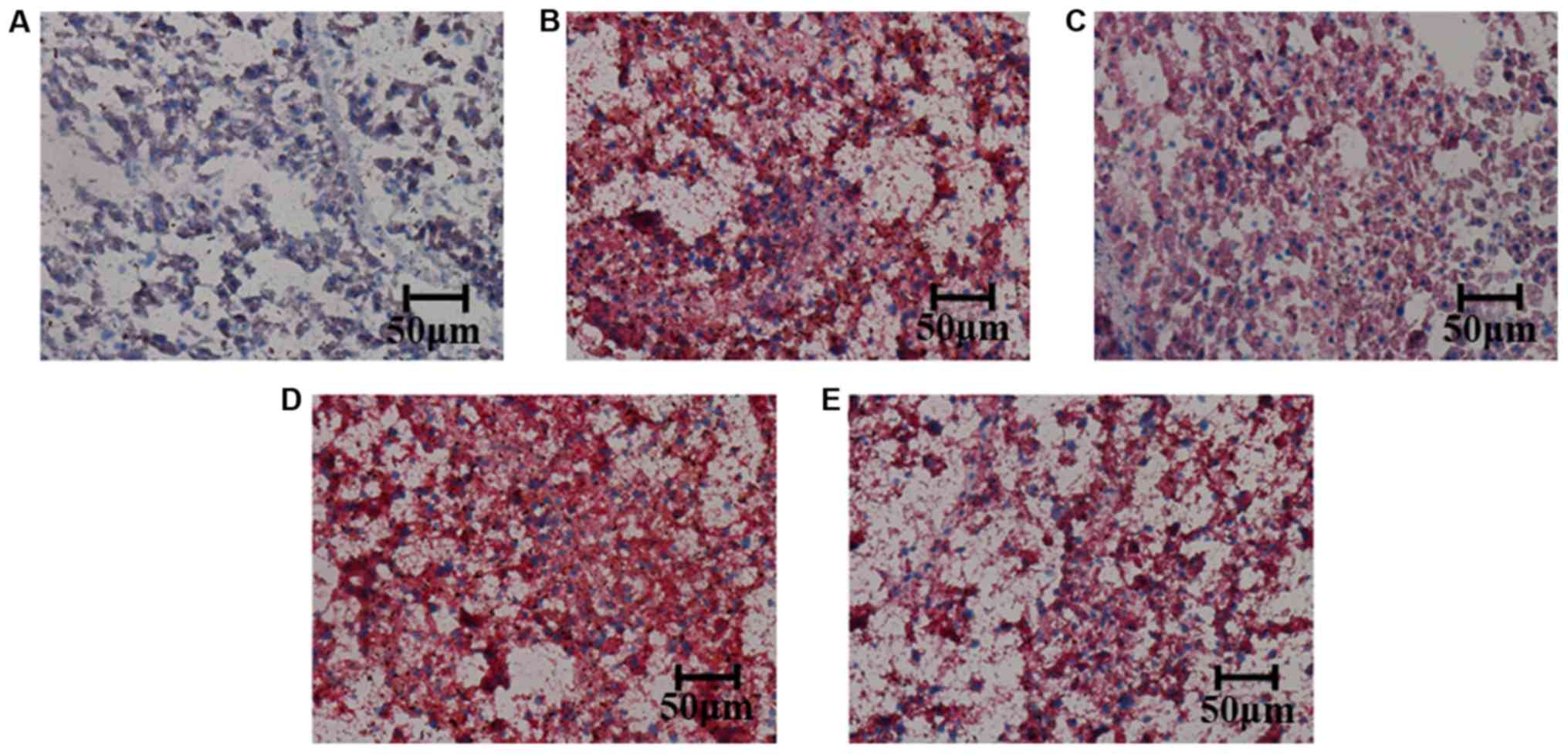
High doses of TMP can improve fat
deposition in the liver
The fat deposition of the liver was detected by Oil
Red O staining. It was observed that, compared with the control
group, the fat deposition in the liver was increased in the model
group. The fat deposition of liver was decreased in the simvastatin
4 and 200 mg/kg/day TMP-treated groups; however, not in the 100
mg/kg/day TMP-treated group, compared with the model group
(Fig. 2).
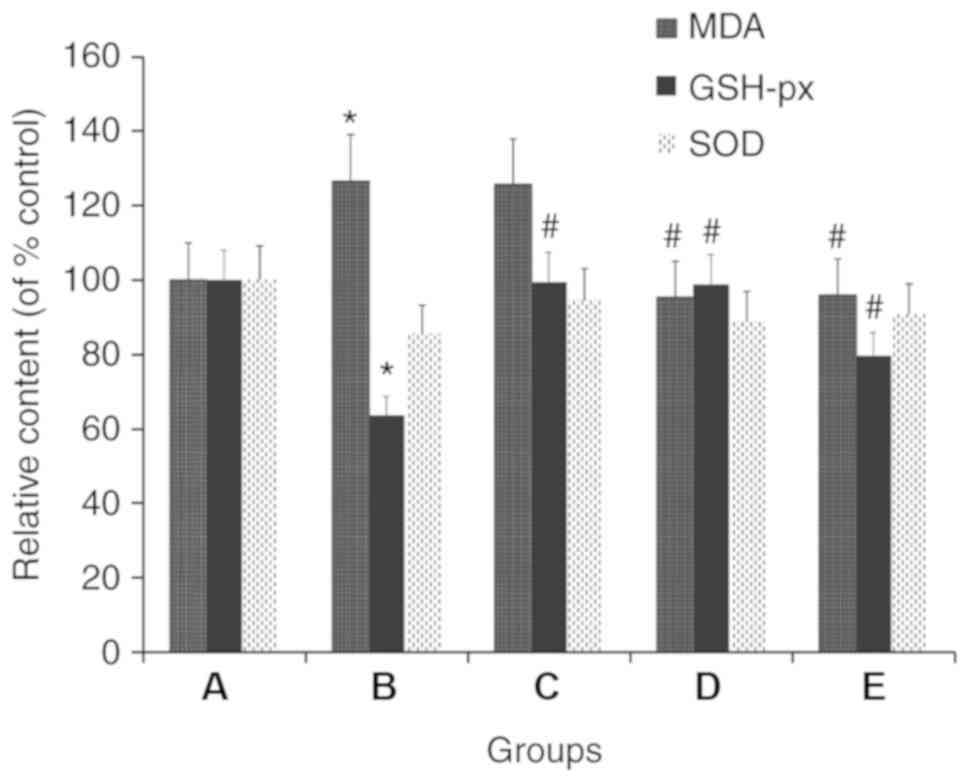
TMP improves the levels of MDA, SOD
and GSH-px in liver tissues
Compared with the control group, the model group
exhibited GSH-px and SOD levels that were 36.2 and 14.5% lower,
respectively, in the liver tissues; however, the MDA level was
26.4% higher in the model group. It was identified that, compared
with the model group, the C57BL/6J mice treated with 100 and 200
mg/kg/day TMP exhibited 24.9 and 55.0% higher GSH-px levels,
respectively, in the liver tissues; however, the SOD levels were
not increased. Additionally, the MDA level was 24.0 and 24.5%
lower, respectively, in the 100 and 200 mg/kg/day TMP groups,
compared with that in the model group. Furthermore, compared with
model group, the C57BL/6J mice treated with 4 mg/kg/day simvastatin
had 55.9 and 10.5% higher GSH-px and SOD levels, respectively, in
the liver tissues, whereas the MDA levels did not increase
(Fig. 3).
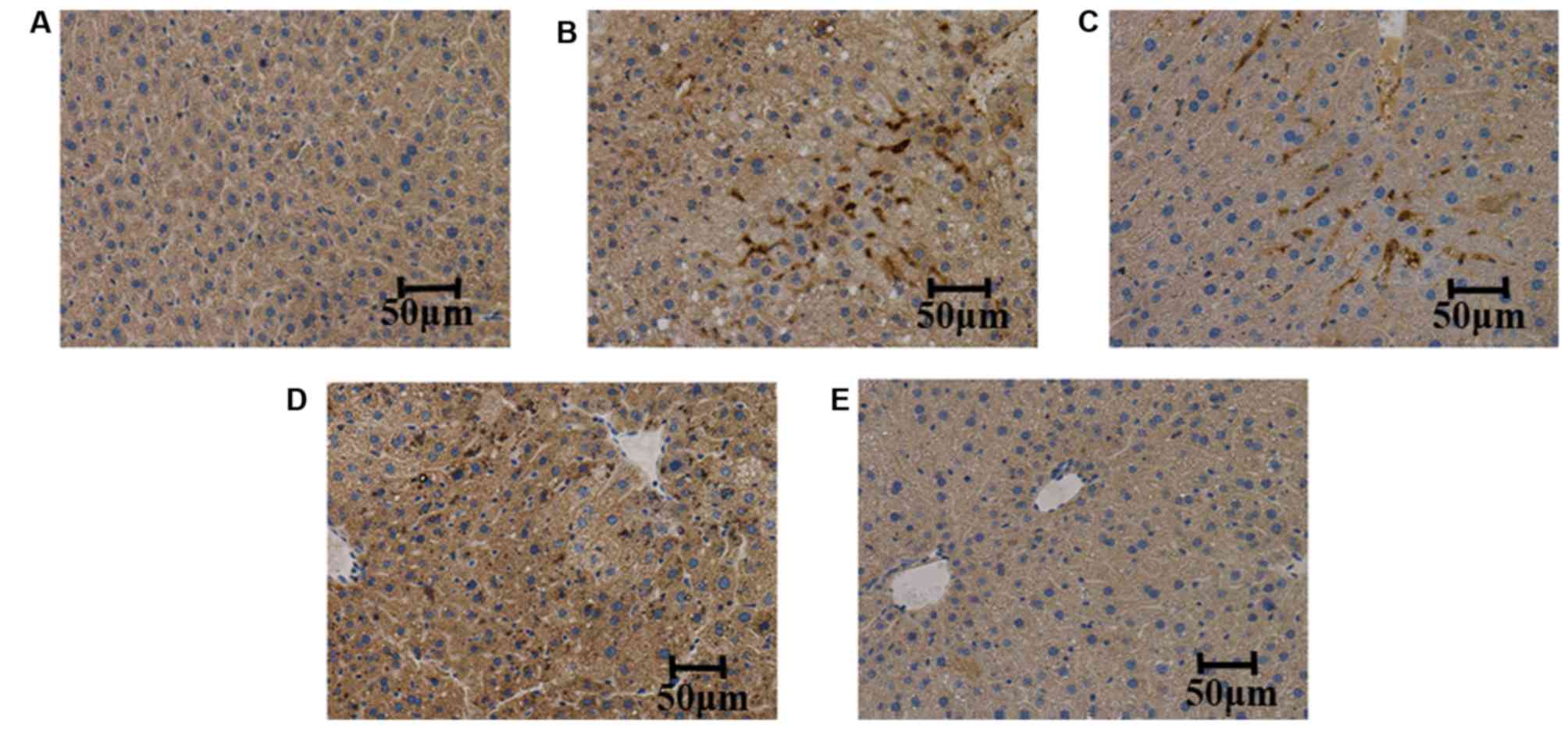
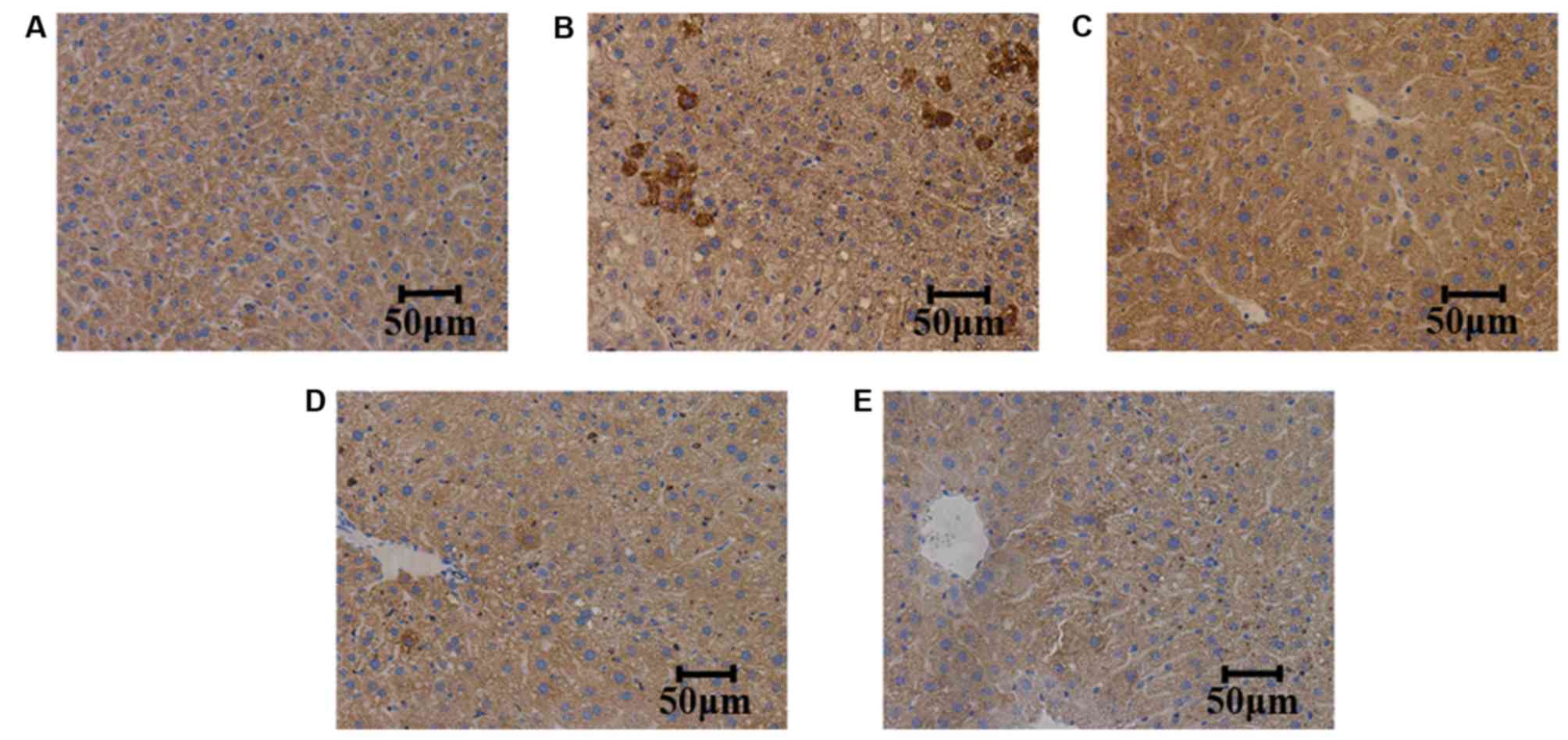
TMP suppresses the expression of TNF-α
and IL-6 in liver tissues
The expression levels of TNF-α and IL-6 were
measured in the liver tissues of the mice. It was observed that,
compared with the control group, the expression levels of TNF-α and
IL-6 were increased in the model group. Furthermore, compared with
the model group, TMP (100 and 200 mg/kg/day) treatment reduced the
expression levels of TNF-α and IL-6; however, simvastatin did not
suppress the expression of TNF-α and IL-6, as determined by
immunohistochemistry (Figs. 4 and
5).
TMP suppresses the expression of
TNF-α, IL-6 and NF-κB, and the phosphorylation of NF-κB in liver
tissues
Western blot analysis was conducted to determine the
expression of TNF-α, IL-6 and NF-κB, and the phosphorylation of
NF-κB in the liver tissues. It was demonstrated that, compared with
the control group, the expression levels of TNF-α and IL-6, and the
ratio of p-NF-κB/NF-κB were increased in the model group. Compared
with the model group, TMP (100 and 200 mg/kg/day) and simvastatin
treatment reduced the expression levels of TNF-α and IL-6, and the
ratio of p-NF-κB/NF-κB (Fig.
6).
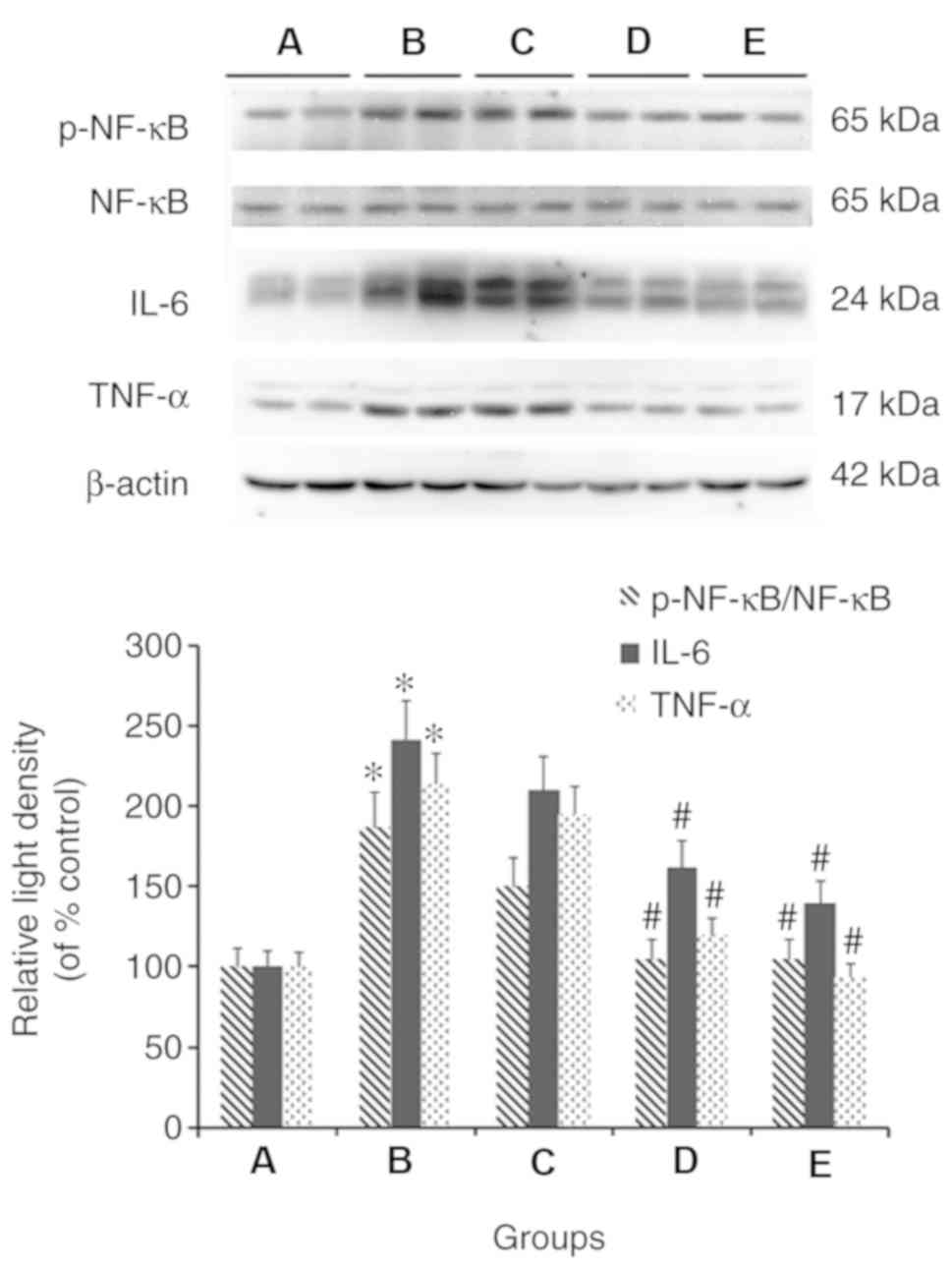 | Figure 6.Effects of TMP on the expression of
TNF-α, IL-6 and the ratio of p-NF-κB/NF-κB in the liver tissues of
mice fed a high fat diet. Western blot analysis of representative
liver tissue samples from the (A) control, (B) model, (C) 4
mg/kg/day simvastatin-treated, (D) 100 mg/kg/day TMP-treated and
(E) 200 mg/kg/day TMP-treated groups are shown. *P<0.05,
compared with the control group; #P<0.05, compared
with the model group. TMP, tetramethylpyrazine; TNF-α, tumor
necrosis factor-α; IL-6, interleukin-6; NF-κB, nuclear factor
κB. |
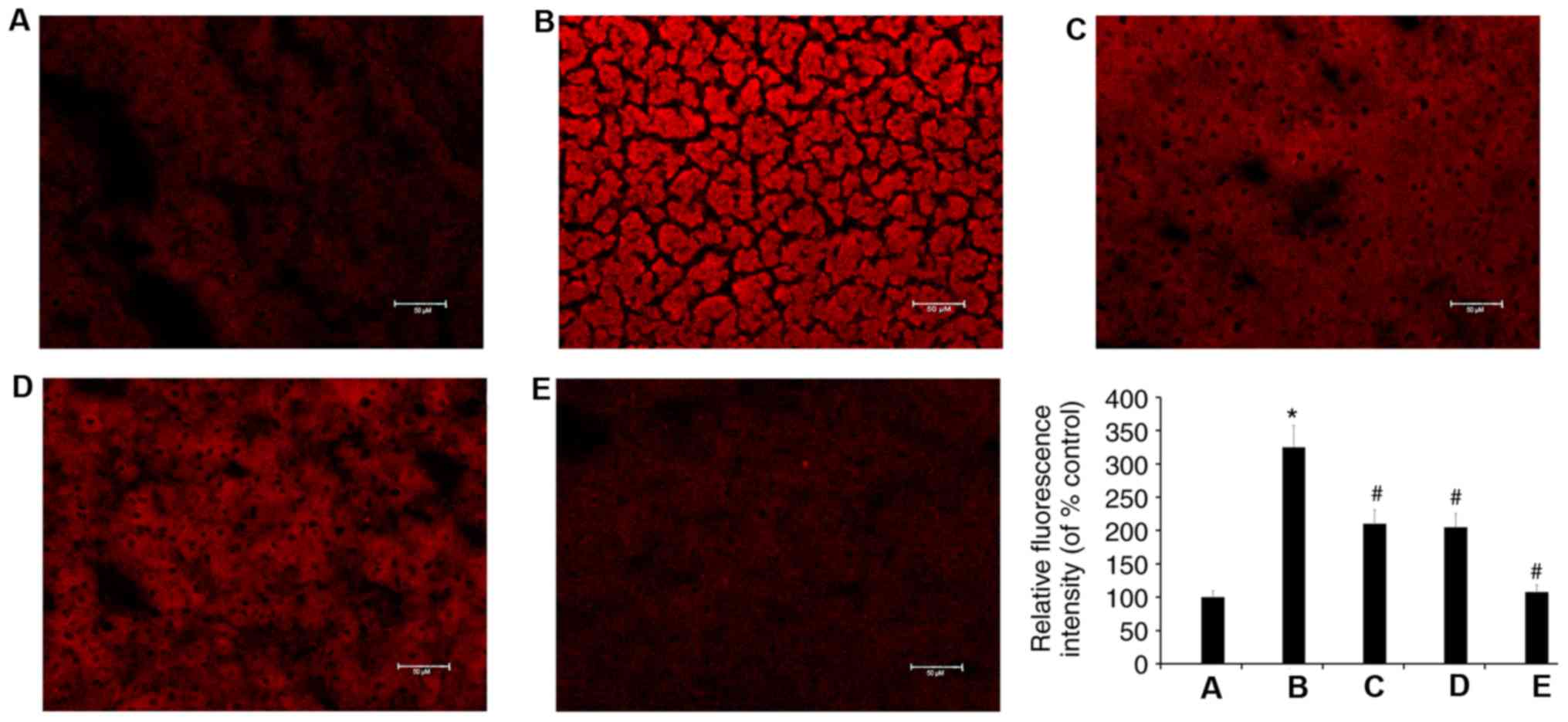
TMP suppresses the production of ROS
induced by a high fat diet
It was found that, compared with the control group,
the production of ROS was increased in the livers of C57BL/6 mice
fed a high fat diet. It was also demonstrated that, compared with
the model group, TMP (100 and 200 mg/kg/day) and simvastatin
treatment reduced the production of ROS (Fig. 7).
Discussion
NAFL is characterized by an accumulation of
hepatocellular lipids, mainly triglycerides, whereas NASH is
characterized by the presence of hepatocellular damage and
inflammation (15), which can
drive the development of liver fibrosis (16), the strongest predictor of
NAFLD-associated mortality (17).
If inflammation is inhibited, the progression of NAFL to NASH may
be delayed or repressed, which may also restrict the development of
liver fibrosis. The present study investigated the effect of TMP on
liver inflammation induced by a high fat diet. At 8 weeks, blood
was collected through the eyeball of a random selection of ~10% of
mice fed a high fat diet (six mice) and 50% of mice fed a regular
diet (six mice), and the levels of AST and ALT were detected. The
results showed that, compared with mice fed a regular diet, the
levels of AST and ALT were increased (data not shown) in the mice
fed a high fat diet. According to this, it was possible to confirm
the presence of liver damage in the mice fed a high fat diet.
Therefore, inflammation of the liver was not examined at 8 weeks,
but at 12 weeks in the mice fed a high fat diet. The murine NAFLD
model was established by feeding mice a high fat diet for 8 weeks.
NAFL induced in C57BL/J mice imitates human NAFLD, and the standard
for the establishment of the murine model for NAFLD is the NAFLD
activity score. Therefore, the NAFLD activity score was considered
to be suitable for the murine NAFLD model. Others have also used
the NAFLD activity score to examine the liver of C57BL/6JRj mice
(18). In the present study, if
there was no intervention of the model group, NAFL progressed to
NASH and the NAFLD activity scores of the model group were >5.
It was observed that simvastatin and 200 mg/kg/day TMP lowered the
increased body weight and liver weight of the mice, which had been
induced by a high fat diet. It was hypothesized that the loss of
weight following treatment with simvastatin and 200 mg/kg/day TMP
is due to the reduction in the weight of the liver. However, 100
mg/kg/day TMP only lowered the body weight of the mice. According
to the manufacturer's instructions, the minimum lethal dose of TMP
in mice is 700–800 mg/kg via intraperitoneal administration.
Therefore, it was hypothesized that TMP also had the effect of
weight loss. The effect of different doses of TMP on the body
weight of mice requires further investigation.
In the present study, it was observed that TMP had
no effect on serum glucose, TC, TG or LDL-C. However, TMP protected
hepatocytes from damage, as it was demonstrated that 200 mg/kg/day
TMP decreased the ALT level. The positive control agent (4
mg/kg/day simvastatin) did not decrease the serum levels of
glucose, ALT, TG or LDL-C; however, it decreased the TC level.
Therefore, compared with simvastatin, TMP had a protective effect
on the liver. Future experiments aim to investigate the molecular
mechanism of TMP in preventing NASH. The pathogenesis of NASH
includes insulin resistance, increased inflammation (TNF-α and
IL-6) and increased oxidative damage (18). The levels of TNF-α and other
TNF-induced cytokines, including IL-6 and IL-8, were higher in
animals with NASH. This is likely associated with the progression
of NASH in liver cirrhosis (19).
In the present study, it was observed that TMP inhibited the liver
inflammation induced by a high fat diet. The expression levels of
TNF-α and IL-6 were increased in C57BL/J mice fed a high fat diet.
Compared with the model group, TMP decreased the expression of
TNF-α and IL-6. However, the positive control agent, simvastatin,
at a dose of 4 mg/kg/day did not have this effect. Therefore, it
can be deduced that TMP, rather than simvastatin, can inhibit liver
inflammation. It was previously reported that TNF-α can activate
NF-κB (20). In the present study,
it was also observed that the phosphorylation of NF-κB was
increased in C57BL/J mice fed a high fat diet, and TMP inhibited
the increased phosphorylation of NF-κB. Therefore, TMP can inhibit
the inflammatory response in the liver induced by a high fat diet
by inhibiting the expression of TNF-α and the phosphorylation of
NF-κB.
It was previously reported that TNF-α is a regulator
of the generation of ROS (21,22).
ROS are essential for normal physiological functions, gene
expression, cell growth, defense against infections and the control
of vascular endothelial cells (23). However, when ROS production exceeds
scavenging abilities, cells are exposed to oxidative stress and are
damaged (24,25). The present study investigated
whether ROS levels were increased in the livers of mice fed a high
fat diet following an increase of TNF-α, and whether TMP inhibits
the increase of ROS. The results demonstrated that the levels of
ROS and TNF-α were increased in the livers of mice fed a high fat
diet. TMP treatment lowered the increased levels of ROS and TNF-α
induced by the high fat diet. It can be deduced that TMP protects
the liver by inhibiting the TNF-α/ROS/NF-κB signaling pathway. The
effect of simvastatin on ROS requires further investigation.
In conclusion, the present study is the first to
demonstrate, to the best of our knowledge, that TMP inhibits the
inflammatory response of the liver by reducing the production of
TNF-α and IL-6, the generation of ROS and subsequently NF-κB
activation. TMP has the potential to become an effective
therapeutic drug to treat liver disease by inhibiting the
inflammatory response. A limitation of the present study is that
time course data on the effect of TMP were absent. Further
investigations are required to investigate the effect of TMP on a
high fat-induced murine NAFLD model in time-dependent manner.
Although it was observed that TMP suppressed the production of
TNF-α and IL-6 in vivo, for a definitive conclusion, an
in vitro investigation of TMP is required.
Acknowledgements
Not applicable.
Funding
This study was supported by grants from the National
Natural Science Foundation of China (grant no. 81671391) and
Beijing Hospital Nova Project (grant no. BJ-2016-033).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BC and YM performed the experiments and analyzed the
data. XX and JW contributed to the conception and design of the
study. GH and YL designed the present study and revised the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by The Research Ethics
Committee of the China Academy of Chinese Medical Sciences
(Beijing, China), in accordance with the National Institutes of
Health Guidelines for the Care and Use of Laboratory Animals
(National Institutes of Health, Bethesda, MD, USA). All animals
were treated in accordance with the guidelines and regulations for
the use and care of animals of the Center for Laboratory Animal
Care, China Academy of Chinese Medical Sciences.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ghaemi A, Hosseini N, Osati S, Naghizadeh
MM, Dehghan A, Ehrampoush E, Honarvar B and Homayounfar R: Waist
circumference is a mediator of dietary pattern in non-alcoholic
fatty liver disease. Sci Rep. 8:47882018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Komazaki R, Katagiri S, Takahashi H,
Maekawa S, Shiba T, Takeuchi Y, Kitajima Y, Ohtsu A, Udagawa S,
Sasaki N, et al: Periodontal pathogenic bacteria, Aggregatibacter
actinomycetemcomitans affect non-alcoholic fatty liver disease by
altering gut microbiota and glucose metabolism. Sci Rep.
7:139502017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Torres DM, Williams CD and Harrison SA:
Features, diagnosis, and treatment of nonalcoholic fatty liver
disease. Clin Gastroenterol Hepatol. 10:837–858. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schröder T, Kucharczyk D, Bär F, Pagel R,
Derer S, Jendrek ST, Sünderhauf A, Brethack AK, Hirose M, Möller S,
et al: Mitochondrial gene polymorphisms alter hepatic cellular
energy metabolism and aggravate diet-induced non-alcoholic
steatohepatitis. Mol Metab. 5:283–295. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schuppan D and Schattenberg JM:
Non-alcoholic steatohepatitis: Pathogenesis and novel therapeutic
approaches. J Gastroenterol Hepatol. 28 (Suppl 1):S68–S76. 2013.
View Article : Google Scholar
|
|
6
|
Ekstedt M, Franzén LE, Mathiesen UL,
Thorelius L, Holmqvist M, Bodemar G and Kechagias S: Long-term
follow-up of patients with NAFLD and elevated liver enzymes.
Hepatology. 44:865–873. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schattenberg JM and Schuppan D:
Nonalcoholic steatohepatitis: The therapeutic challenge of a global
epidemic. Curr Opin Lipidol. 22:479–488. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yeom GG, Min S and Kim SY:
2,3,5,6-Tetramethylpyrazine of Ephedra sinica regulates
melanogenesis and inflammation in a UVA-induced
melanoma/keratinocytes co-culture system. Int Immunopharmacol.
18:262–269. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ran X, Ma L, Peng C, Zhang H and Qin LP:
Ligusticum chuanxiong Hort: A review of chemistry and pharmacology.
Pharm Biol. 49:1180–1189. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shao Z, Wang L, Liu S and Wang X:
Tetramethylpyrazine protects neurons from oxygen-glucose
deprivation-induced death. Med Sci Monit. 23:5277–5282. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gong X, Ivanov VN, Davidson MM and Hei TK:
Tetramethylpyrazine (TMP) protects against sodium arsenite-induced
nephrotoxicity by suppressing ROS production, mitochondrial
dysfunction, pro-inflammatory signaling pathways and programed cell
death. Arch Toxico. 89:1057–1070. 2015. View Article : Google Scholar
|
|
12
|
Kleiner DE, Brunt EM, Van Natta M, Behling
C, Contos MJ, Cummings OW, Ferrell LD, Liu YC, Torbenson MS,
Unalp-Arida A, et al: Design and validation of a histological
scoring system for nonalcoholic fatty liver disease. Hepatology.
41:1313–1321. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Staroń R, Van Swelm RP, Lipiński P,
Gajowiak A, Lenartowicz M, Bednarz A, Gajewska M, Pieszka M,
Laarakkers CM, Swinkels DW and Starzyński RR: Urinary hepcidin
levels in iron-deficient and iron-supplemented piglets correlate
with hepcidin hepatic mRNA and serum levels and with body iron
status. PLoS One. 10:e1366952015. View Article : Google Scholar
|
|
14
|
Wei J, Zhen YZ, Cui J, He FL, Shen T, Hu
G, Ren XH and Lin YJ: Rhein lysinate decreases inflammation and
adipose infiltration in KK/HlJ diabetic mice with non-alcoholic
fatty liver disease. Arch Pharm Res. 39. 960–969. 2016.
|
|
15
|
Rinella ME: Nonalcoholic fatty liver
disease: A systematic review. JAMA. 313:2263–2273. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fujii H and Kawada N: Inflammation and
fibrogenesis in steatohepatitis. J Gastroenterol. 47:215–225. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Morrison MC, Kleemann R, van Koppen A,
Hanemaaijer R and Verschuren L: Key inflammatory processes in human
NASH are reflected in Ldlr−/−. Leiden mice: A
translational gene profiling study. Front Physiol. 9:1322018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Henkel J, Alfine E, Saín J, Jöhrens K,
Weber D, Castro JP, König J, Stuhlmann C, Vahrenbrink M, Jonas W,
et al: Soybean oil-derived poly-unsaturated fatty acids enhance
liver damage in NAFLD induced by dietary cholesterol. Nutrients.
10(pii): E13262018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hebbard L and George J: Animal models of
nonalcoholic fatty liver disease. Nat Rev Gastroenterol Hepatol.
8:35–44. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hayden MS and Ghosh S: Regulation of NF-κB
by TNF family cytokines. Semin Immunol. 26:253–266. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Abd El-Kader SM, Al-Shreef FM and
Al-Jiffri OH: Biochemical parameters response to weight loss in
patients with non-alcoholic steatohepatitis. Afr Health Sci.
16:242–249. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kastl L, Sauer SW, Ruppert T, Beissbarth
T, Becker MS, Süss D, Krammer PH and Gülow K: TNF-α mediates
mitochondrial uncoupling and enhances ROS-dependent cell migration
via NF-κB activation in liver cells. FEBS Lett. 588:175–183. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Blaser H, Dostert C, Mak TW and Brenner D:
TNF and ROS crosstalk in inflammation. Trends Cell Biol.
26:249–261. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Montezano AC and Touyz RM: Reactive oxygen
species and endothelial function-role of nitric oxide synthase
uncoupling and Nox family nicotinamide adenine dinucleotide
phosphate oxidases. Basic Clin Pharmacol Toxicol. 110:87–94. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Togo M, Konari N, Tsukamoto M, Kimoto R,
Yamaguchi T, Takeda H and Kambayashi I: Effects of a high-fat diet
on superoxide anion generation and membrane fluidity in liver
mitochondria in rats. J Int Soc Sports Nutr. 15:132018. View Article : Google Scholar : PubMed/NCBI
|