Introduction
Pulmonary arterial hypertension (PAH) is a fatal
syndrome characterized by elevated pulmonary arterial resistance,
which can cause right ventricular insufficiency with high mortality
(1,2). Previous studies have indicated that
the primary pathogenesis of PAH is pulmonary vascular remodeling,
which is associated with excessive migration of smooth muscle
cells, oxidative stress, extracellular matrix (ECM) deposition and
perivascular inflammation (3–6).
Particularly, ECM deposition and perivascular inflammation have
been demonstrated to exert great influence in the pathogenesis of
PAH (4,6). Furthermore, multiple reports have
demonstrated that certain pathways, including ERK and NF-κB, are
associated with ECM deposition and inflammation in PAH, providing
potential therapeutic targets for PAH (7,8).
ECM is a basic component of peripheral connective
tissues. It contains numerous structural proteins including
collagen, elastin and fibronectin, among which the relative
contents of collagen and elastin determine the biological
activities of blood vessels and play important roles in cell
signaling pathway regulation and intercellular communications
(9–11). Previous studies have indicated that
the ECM proteins can be modulated by matrix metalloproteinases
(MMPs), particularly, MMP2 and MMP9 can maintain the stability of
ECM (12–14). Thus, the integrality of ECM
components is critical to normal pulmonary function, knowledge of
which contributes to comprehension of the pathogenesis of PAH.
As previously reported, the progression of PAH is
closely related to inflammation; lymphocytes and macrophages
existing around re-modeled pulmonary vessels, and the inflammatory
cytokines in PAH patients increase markedly (15). A previous study also reported that
monocrotaline (MCT)-induced PAH in rats is associated with chronic
pulmonary inflammation (16).
Therefore, suppressing inflammation may become a valid therapy for
PAH.
Formononetin (FMN) is a natural phytoestrogen
isolated from red clover (Trifolium pratense) and has
various biological functions, including proapoptotic,
anti-inflammatory and anti-tumor activities (17). Previous studies have suggested that
FMN can improve various cardiovascular diseases (18,19).
FMN also exhibits strong inhibitory effects on human prostate
cancer cells and nasopharyngeal carcinoma cells (20,21).
Other studies have indicated that reduction of FMN-mediated ECM
deposition and suppression of inflammatory responses are related to
the inactivation of ERK and NF-κB signaling in various cells
(22–24). However, the inhibitory effects of
FMN on PAH and their possible mechanisms are unclear.
Therefore, the objective of the present study was to
explore the therapeutic effectiveness of FMN on MCT-induced PAH and
its effects on ECM deposition and perivascular inflammation in
rats.
Materials and methods
Animals and reagents
In the present study 46 male Sprague-Dawley rats
weighing 230–250 g (7-weeks-old) were purchased from the
experimental animal center of Zhejiang Province. The experimental
procedure was approved by the Ethics Review of Animal Use
Application of the Fifth Affiliated Hospital of Wenzhou Medical
University. All animals were housed at 20–26°C, with 45–55%
humidity and a 12-h light/dark cycle, and had free access to food
and water.
FMN with 98% purity was obtained from MedChem
Express. Bovine serum albumin (BSA) and MCT were provided by
Sigma-Aldrich (Merck KGaA). The primary TGFβ1 (cat. no. sc146)
antibody was provided by Santa Cruz Biotechnology, Inc. The primary
phosphorylated (p-)ERK (cat. no. 9101S), ERK (cat. no. 9102S),
NF-κB (cat. no. 8242S) and GAPDH (cat. no. 5174S) antibodies, and
anti-rabbit (cat. no. 7074S) and anti-mouse (cat. no. 7076S)
HRP-conjugated secondary antibodies were provided by Cell Signaling
Technology, Inc. The primary p-NF-κB (cat. no. ab86299), MMP2 (cat.
no. ab86607), MMP9 (cat. no. ab38898), collagen I (cat. no.
ab34710), collagen III (cat. no. ab7778), fibronectin (FN; cat. no.
ab6328), monocyte chemoattractant protein (MCP)-1 (cat. no.
ab25124), interleukin (IL)1β (cat. no. ab9722), and tumor necrosis
factor (TNF) α (cat. no. ab6671) antibodies were provided by Abcam.
BCA (cat. no. p0012s) and enhanced chemiluminescence (ECL; cat. no.
p0018s) kits were purchased from Beyotime Institute of
Biotechnology. swere obtained from Beijing Solarbio Science &
Technology Co., Ltd.
Experimental design
All rats were randomly assigned into five groups: i)
Control group (n=6); ii) MCT group (n=10); iii) low-FMN group
(n=10; 10 mg/kg/day FMN); iv) medium-FMN group (n=10; 30 mg/kg/day
FMN); and v) high-FMN group (n=10; 60 mg/kg/day FMN). MCT was
dissolved in 1 mol/l hydrochloric acid neutralized with 1 mol/l
sodium hydroxide and diluted with normal saline. The pH was
adjusted to 7.35–7.45. FMN was dissolved in DMSO and diluted with
olive oil. According to a previous study, the rats in the MCT group
received a single subcutaneous injection of 60 mg/kg MCT at day 0
(25), and the control rats were
injected with the equivalent volume of fresh saline. According to a
previous study, echocardiography suggests that PAH can be
established after 2 weeks of MCT injection, and right ventricular
systolic pressure (RVSP) significantly increases (26). Therefore, after 2 weeks of MCT
injections, all rats from these three FMN groups were
intraperitoneally administered with the corresponding doses of FMN
daily for 2 weeks.
Hemodynamic measurement
After 4 weeks of the single MCT injections,
including 2 FMN injections, all rats were weighed and then
anesthetized with pentobarbital sodium (50 mg/kg). Subsequently,
RVSP was measured as reported in our previous study (25). In brief, after the rat was
anesthetized, the right neck tissue was cut open and the right
external jugular vein was isolated, then the venous catheter was
inserted into the right ventricle from the right external jugular
vein. Significant amplitude of right ventricular wave was observed,
and RVSP was recorded using a miniature pressure transducer (cat.
no. TSD104A; BIOPAC Systems, Inc.) and analyzed using a BIOPAC
MP100 data acquisition system (BIOPAC Systems, Inc.).
Assessment of right heart
hypertrophy
After measuring RVSP, the rats were sacrificed with
pentobarbital sodium (150 mg/kg). Then, heart and lung tissues were
removed. According to a previous study (27), right ventricular hypertrophy can be
given as the ratio of the right ventricle (RV) and the left
ventricle plus septum (LV+S) mass. After the rat was sacrificed,
the heart was separated and the atria were cut off, then the right
ventricle was cut out along the edge of ventricular septum, leaving
ventricular septum and left ventricle. RV and LV+S were weighed and
the right ventricular hypertrophy index (RV/LV+S) was calculated
(28). Finally, the RV/LV+S and
RV/body weight (BW) were used to assess the severity of right
ventricular hypertrophy.
Morphological analysis
After removing superfluous tissues, the remaining
lung tissues were fixed in 4% formalin at room temperature for 48 h
and embedded in paraffin. Subsequently, all paraffin blocks were
sectioned at 4-µm and stained with a Masson assay kit (Beijing
Solarbio Science & Technology, Co., Ltd.; cat. no. G1345) or a
H&E assay kit (Beijing Solarbio Science & Technology, Co.,
Ltd.; cat. no. G1120) according to the manufacturers' protocol. A
total of 5 pulmonary arteries 50–150 µm in diameter were randomly
selected to be viewed using a light microscope (Nikon Corporation;
magnification, ×400. The wall thickness was calculated according to
the following equations: Vascular wall thickness percentage
(WT%)=wall thickness/outer diameter ×100; the percentage of
vascular wall area (WA%)=wall transection area/cross-sectional area
×100.
Immunohistochemical staining was also used for
morphological analysis. After dewaxing in 100% xylene (twice in
total, 20 min each time) and rehydrating in a graded alcohol series
(100% for 5 min, 95% for 5 min and 80% for 5 min), the lung tissues
were blocked with 5% BSA at room temperature for 1 h, and incubated
with anti-TGFβ1 antibody (1:200; cat. no. sc146), anti-MMP2
antibody (1:200; cat. no. ab86607) or anti-MMP9 antibody (1:200;
cat. no. ab38898) at 4°C overnight and subsequently with
anti-rabbit (1:50; cat. no. 7074S) or anti-mouse (1:50; cat. no.
7076S) HRP-conjugated secondary antibody at room temperature for 1
h. Finally, the sections were visualized with 3,3′-DAB at room
temperature for 5 min and counterstained with hematoxylin at room
temperature for 2 min, and observed using a light microscope
(magnification, ×400; Nikon Corporation).
Western blot analysis
Lungs were homogenized with RIPA lysis buffer
containing protease (Beyotime Institute of Biotechnology) and
phosphatase inhibitors (Cell Signaling Technology, Inc.), and
protein concentration was detected using a BCA kit. Total protein
lysate (~50 µg) was separated by 10–12% SDS-PAGE and then was
transferred onto PVDF membranes. All membranes were blocked with 5%
BSA for 2 h at room temperature, then incubated with anti-TGFβ1
antibody (1:1,000; cat. no. sc146), anti-MMP2 antibody (1:1,000;
cat. no. ab86607), anti-MMP9 antibody (1:1,000; cat. no. ab38898),
anti-collagen I antibody (1:1,000; cat. no. ab34710), anti-collagen
III antibody (1:1,000; cat. no. ab7778), anti-FN antibody (1:1,000;
cat. no. ab6328), anti-MCP1 antibody (1:2,000; cat. no. ab25124),
anti-IL-1β antibody (1:1,000; cat. no. ab9722), anti-TNFα antibody
(1:1,000; cat. no. ab6671), anti-p-ERK antibody (1:1,000; cat. no.
9101S), anti-ERK antibody (1:1,000; cat. no. 9102S), anti-p-NF-κB
antibody (1:2,000; cat. no. ab86299), anti-NF-κB antibody (1:1,000;
cat. no. 8242S) or anti-GAPDH (1:1,000; cat. no. 5174S) at 4°C
overnight and anti-rabbit IgG HRP-conjugated antibody (1:1,000;
cat. no. 7074S) or anti-mouse IgG HRP-conjugated antibody (1:1,000;
cat. no. 7076S) at room temperature for 1 h. Finally, the
immunoreactive bands were visualized with ECL reagents (Beyotime
Institute of Biotechnology) in AlphaView software 3.3.0
(ProteinSimple).
Statistical analysis
All experiments were repeated three times. One-way
ANOVA and Student-Newman-Keuls test were performed using GraphPad
Prism 5 software (GraphPad Software, Inc.) to analyze all data. The
data are presented as the mean ± standard error of the mean.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effects of FMN on hemodynamics and
right ventricular hypertrophy
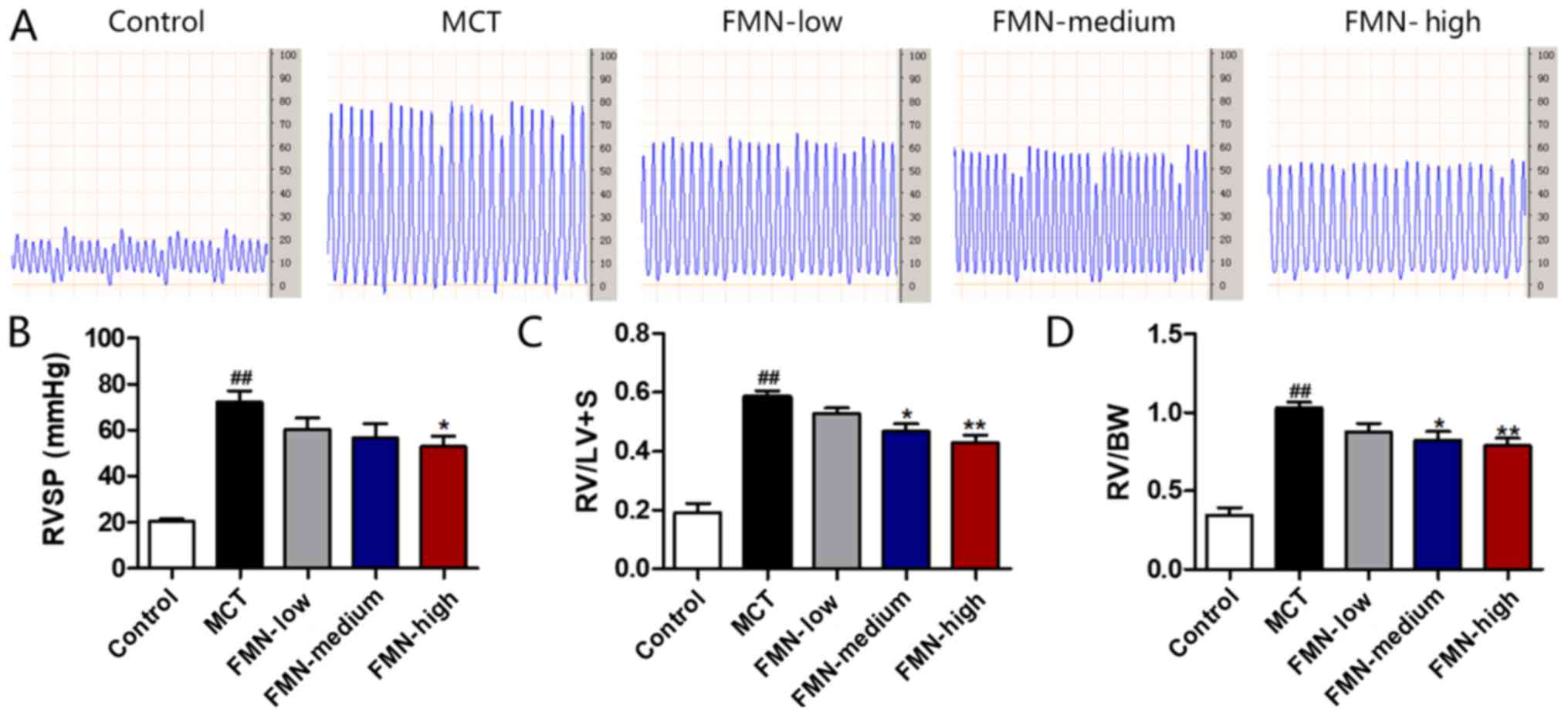
In the present study, PAH, RVSP, RV/LV+S and RV/BW
were measured to assess the inhibitory roles FMN plays in PAH. As
presented in Fig. 1A and B, MCT
administration clearly increased RVSP compared with the control; by
contrast, the increase of RVSP following MCT-treatment was
significantly downregulated by the administration of high-dose FMN
(60 mg/kg). As presented in Fig. 1C
and D, RV/LV+S and RV/BW also significantly increased after MCT
injection compared with the control, while two different doses of
FMN (30 and 60 mg/kg) significantly ameliorated this increase,
thereby decreasing right ventricular hypertrophy.
Effect of FMN on pulmonary vascular
morphology
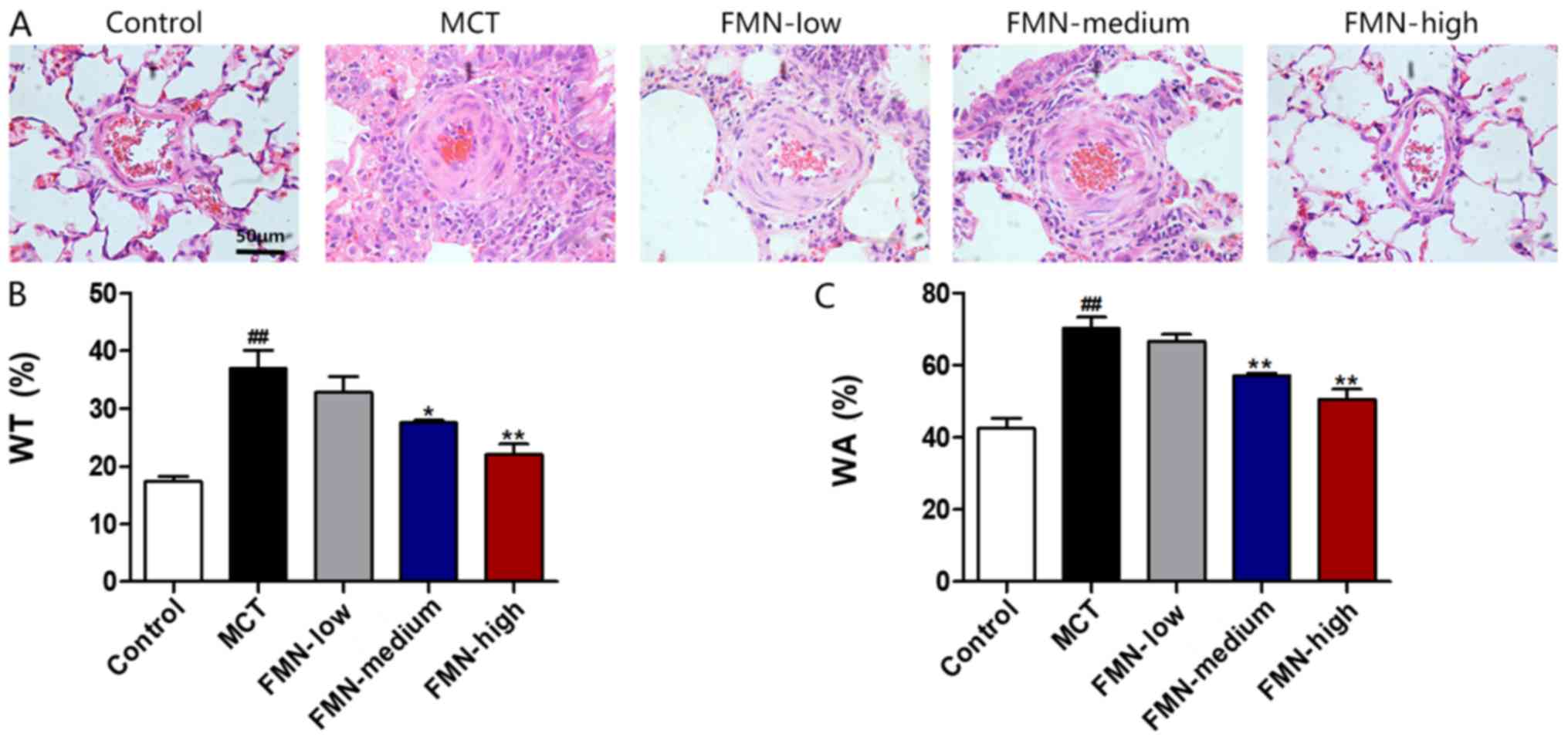
The thickness of pulmonary arterioles with a 50–150
µm diameter was measured. As shown in Fig. 2, MCT administration significantly
enhanced WT and WA% in pulmonary arterioles compared with the
control; however, medium-dose and high-dose FMN treatments
significantly reversed the MCT-induced WT and WA% increases.
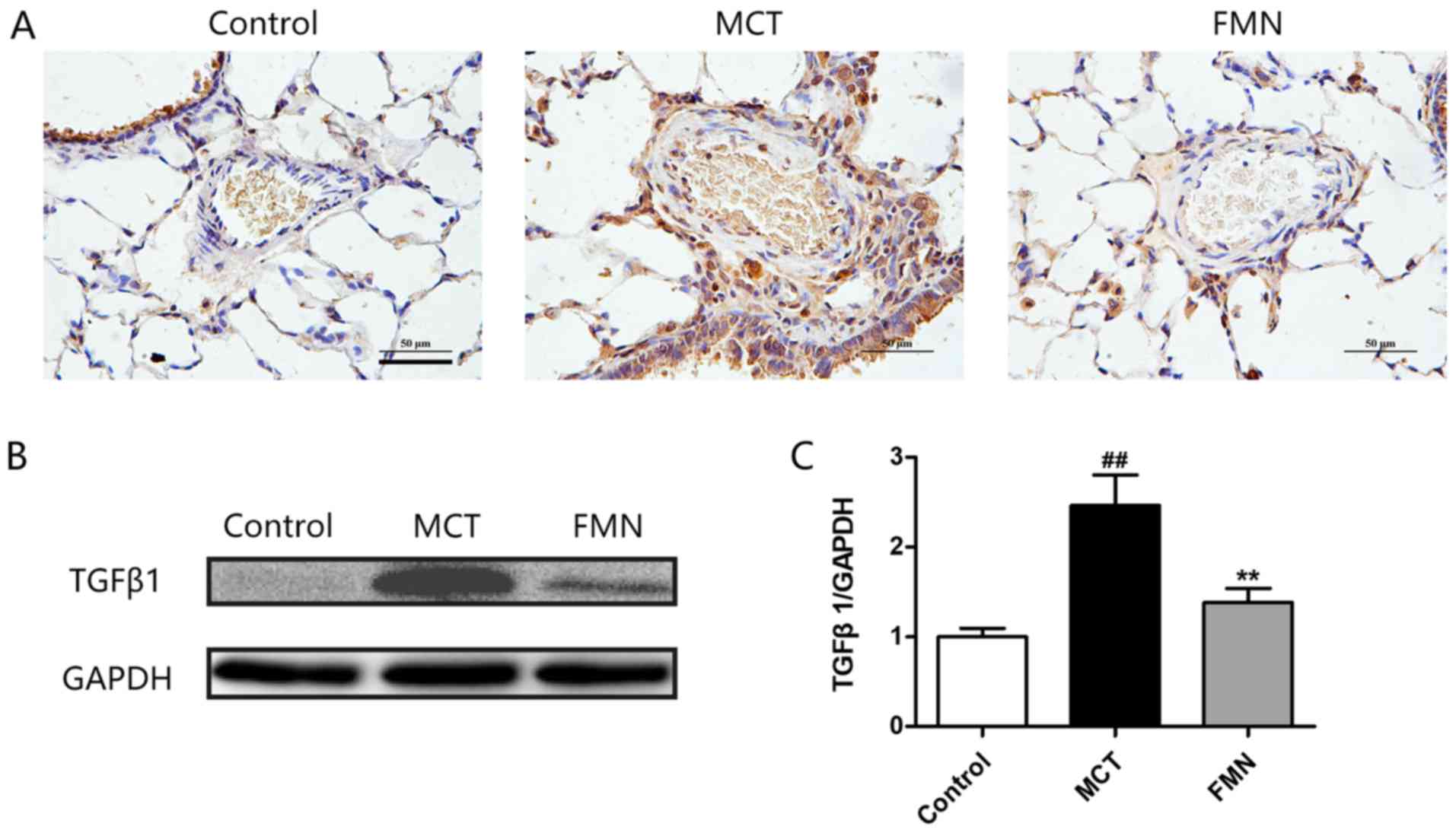
Effect of FMN on TGFβ1 expression
As another key indicator for PAH, TGFβ1 was also
measured by immunohistochemical staining and western blotting. The
results demonstrated that TGFβ1 expression in lungs was
significantly upregulated after MCT injection compared with the
control, but high-dose FMN administration significantly attenuated
this change, and the expression level of TGFβ1 in various groups
detected by western blotting was consistent with the
immunohistochemical results (Fig.
3).
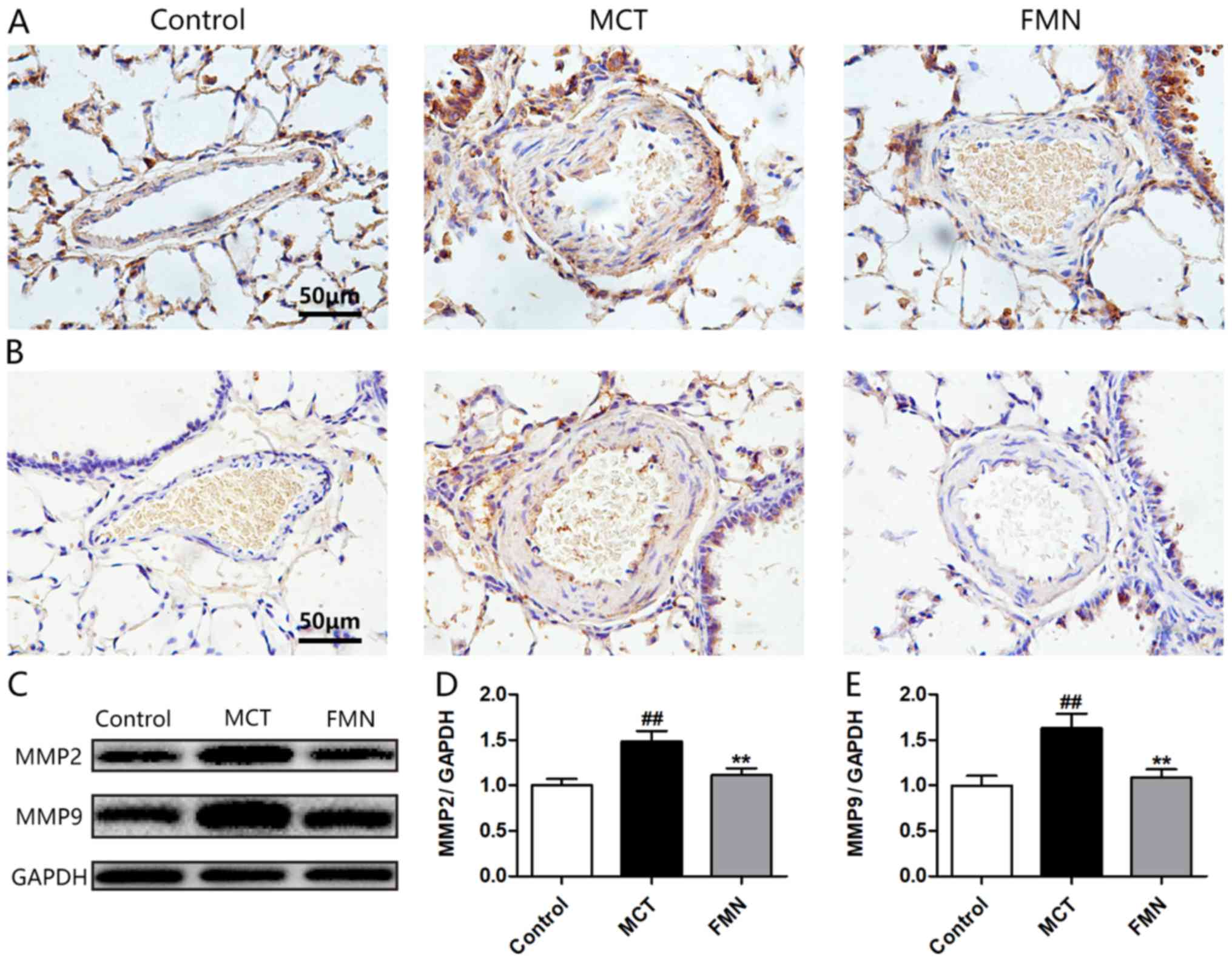
Effects of FMN on the expression of
MMPs
As shown in Fig. 4,
the expression levels of MMP2 and MMP9 in lungs from the MCT group
were higher compared with the control group. However, high-dose FMN
reversed these increases (Fig. 4A and
B). The results from western blotting were consistent with
those from immunohistochemical staining, demonstrating that MCT
significantly upregulated the expression levels of MMPs in lungs,
while the increases of MMPs were significantly alleviated by
high-dose FMN administration (Fig.
4C-E).
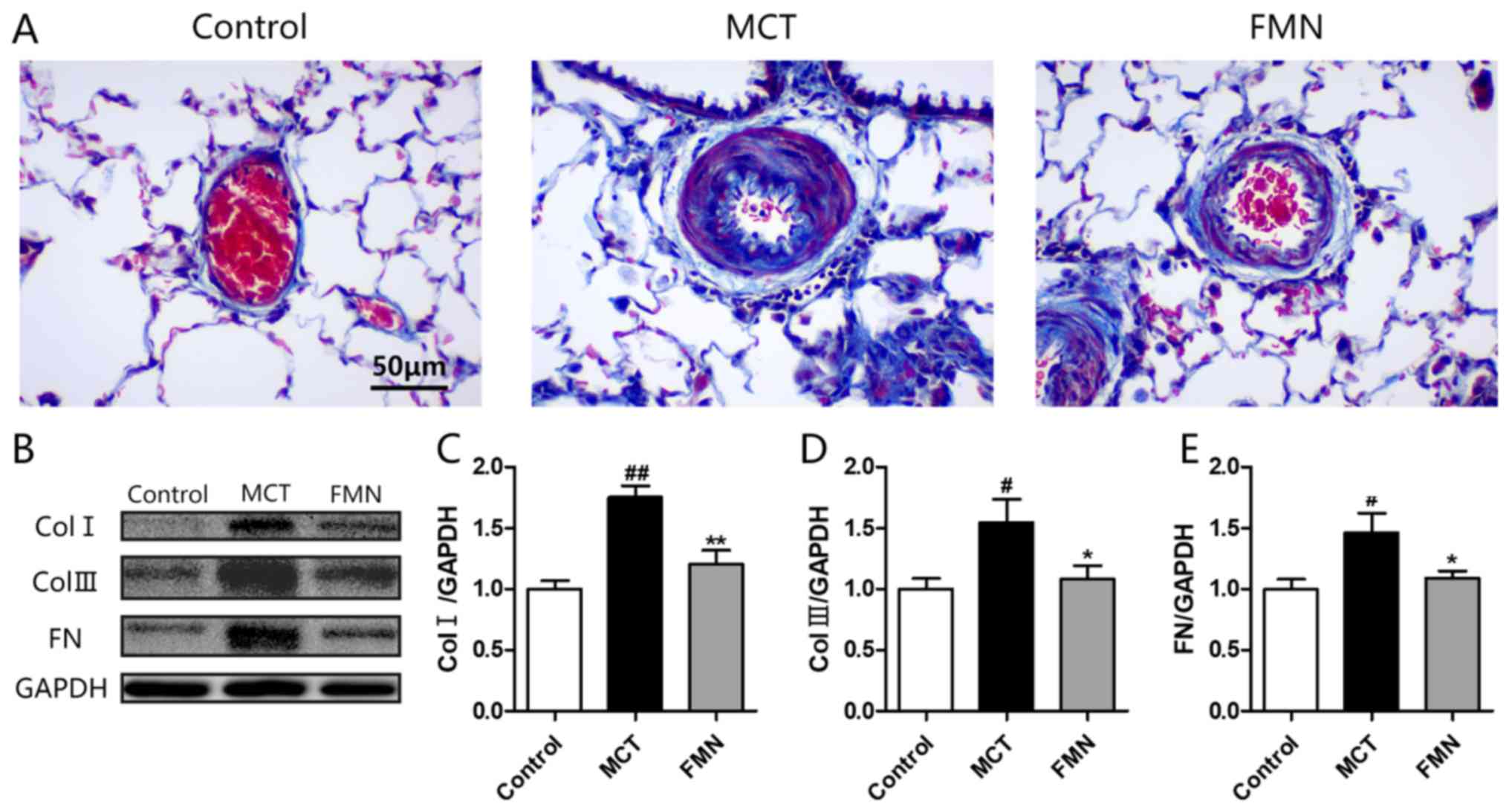
Effect of FMN on ECM deposition
Masson staining demonstrated that high-dose FMN
administration decreased the amount of dense focal collagen
deposition increased by MCT (Fig.
5A). To evaluate the effect of FMN on ECM accumulation, the
levels of biomarkers for ECM in lungs were measured. As western
blotting demonstrated, the expression levels of collagen type I,
collagen type III and fibronectin in the lungs from the MCT group
were significantly higher compared with the control group (Fig. 5B and C). By contrast, high-dose FMN
administration significantly attenuated the increases of these ECM
biomarkers.
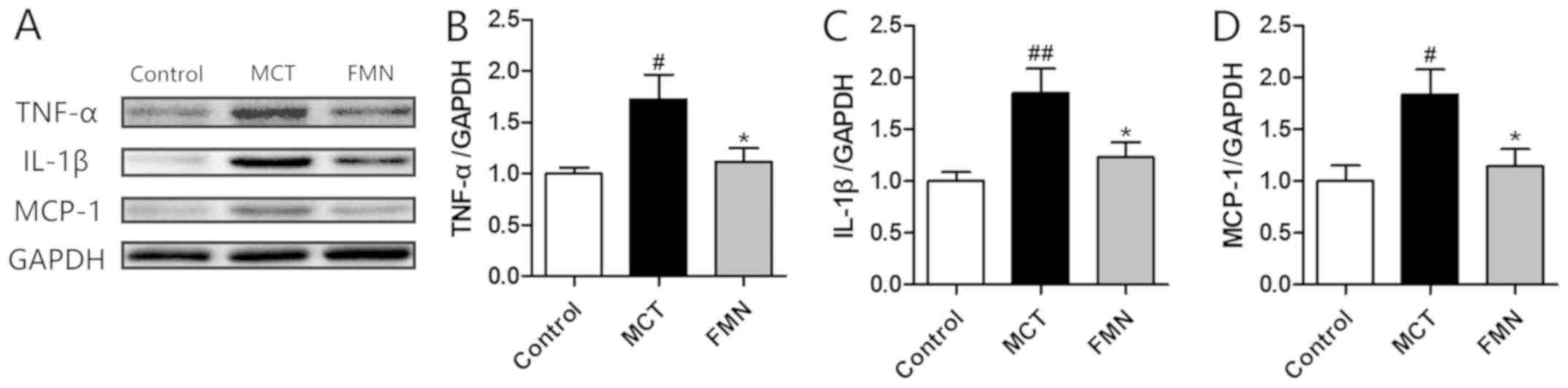
Effect of FMN on perivascular
inflammation
As the H&E staining demonstrated, there were
evident perivascular inflammatory cell infiltrations in the lungs
from MCT group. By contrast, FMN alleviated the MCT-induced
inflammation with the increase of its concentration as demonstrated
in Fig. 2A. The expression levels
of several inflammatory cytokines were also determined. The results
indicated that the expression levels of TNF-α, IL-1β and MCP-1 were
significantly higher in the MCT group compared with the control
group, but high-dose FMN administration significantly attenuated
the increases of these inflammatory cytokines (Fig. 6).
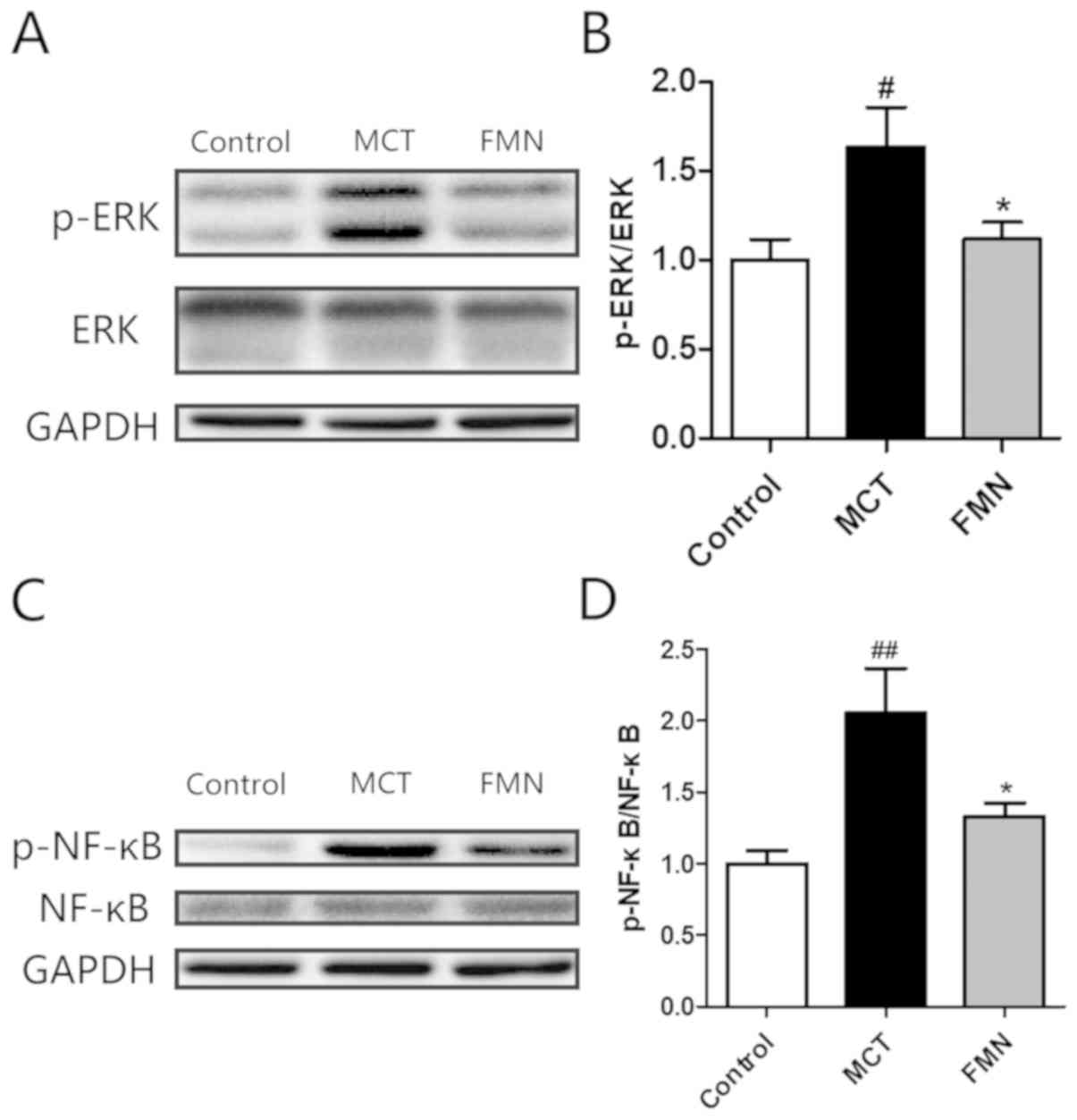
Effects of FMN on ERK and NF-κB
signaling pathways
Western blotting was used to determine and calculate
the ratio of p-ERK and total ERK. The results demonstrated that
this ratio significantly increased in MCT group compared with the
control group, while the increase was significantly attenuated by
high-dose FMN administration (Fig. 7A
and B). The ratio of p-NF-κB and total NF-κB was also
determined. As revealed by western blotting, the ratio was
significantly enhanced in the MCT group compared with the control
group, while high-dose FMN administration significantly attenuated
the increase (Fig. 7C and D).
Discussion
PAH is a fatal syndrome characterized by pulmonary
vascular remodeling, excessive vasoconstriction and subsequent
increased pulmonary artery pressure, and can cause right-sided
heart failure. It is hypothesized that ECM deposition and chronic
inflammation are the main factors causing pulmonary vascular
remodeling (17). FMN, a Chinese
herbal medicine, can be used for cardiovascular diseases (18). The results of the present study
demonstrated that intraperitoneal injection of FMN played an
inhibitory role in MCT-induced PAH in rats. In addition, the
suppression of ERK and NF-κB signaling may be associated with the
mechanism of FMN treatment for MCT-induced PAH.
It has been demonstrated that PAH in humans and
animals is also characterized by adverse changes in pulmonary
artery hemodynamics, including a sharp increase of right
ventricular systolic pressure accompanied with right cardiac
hypertrophy (29,30). The results of the present study
demonstrated that MCT-induced right ventricular hypertrophy was
significantly alleviated with the increase of FMN
concentration.
In addition, the progressive thickening of pulmonary
vascular wall was also a histological characteristic of PAH
(31). As previously described, WT
and WA% can be used to evaluate the degree of vascular thickening
and muscularization of pulmonary arterioles (32). The present study identified that
FMN alleviated the increased WT and WA% with the increase of its
concentration. This finding confirmed that FMN could improve the
MCT-induced PAH. To better illustrate the possible action mechanism
of the inhibitory effect of FMN on pulmonary vascular remodeling,
the high-dose FMN (60 mg/kg) was used for the further
experiments.
TGFβ1 is a crucial cytokine to modulate numerous
cell responses, and increased TGFβ1 expression has been recognized
as an unfavorable factor for PAH (33). Previous studies have indicated that
TGFβ1 was involved in ECM deposition and vascular inflammation by
regulating a variety of signaling pathways (34,35).
Notably, the data from the present study indicated that FMN could
suppress the increased TGFβ1 expression in MCT-induced PAH.
Previous studies have demonstrated that MCT-induced
PAH is involved in ECM deposition of pulmonary arteries and
pulmonary inflammation, in which collagen and fibronectin are
significantly accumulated, thereby increasing the expression levels
of MMPs and inflammatory cytokines (36–38).
The deposition of ECM is a vital change in the pulmonary artery
reconstruction process, and is caused by the interaction between
the synthesis of ECM components and proteolysis (11). Early clinical reports have
indicated that PAH patients exhibit pulmonary artery ruptures and
increase in MMP activity (39,40).
High expression levels of MMP2 and MMP9 are found in lungs from
MCT-induced PAH rats (25). In the
present study, increases of MMP2 and MMP9 expression levels induced
by MCT were inhibited by FMN. Correspondingly, the results from
Masson staining and western blotting suggested that FMN reduced the
MCT-induced ECM accumulation. Certain inflammatory factors are
considered important biomarkers for assessing the severity of PAH
(41). Thus, terminal vascular
remodeling and PAH progression may be improved by suppressing
inflammatory responses. In the present study, the MCT-induced
perivascular inflammatory cell infiltrations in lungs were observed
using H&E staining, and FMN alleviated these inflammatory
changes. As demonstrated by western blotting, FMN suppressed the
MCT-induced inflammatory responses and may serve underlying
therapeutic roles in inflammation-related diseases. Therefore, the
present study indicated that FMN could block pulmonary vascular
remodeling by suppressing the ECM deposition and chronic
inflammation in lung tissues.
The precise action mechanism that leads to PAH
remains to be elucidated, but previous studies have indicated that
ERK and NF-κB signals are closely related to the pathogenesis of
PAH (42,43). ERK is a main factor of the MAPK
family, and its activity in animals with PAH is enhanced (43). In addition, p-ERK is increased in
rat lungs exposed to MCT, and the suppression of p-ERK can prevent
the pulmonary vascular remodeling associated with PAH formation
(44). Notably, ERK signaling has
been demonstrated to coordinate ECM deposition and inflammatory
responses (42,44). The results of the present study
were consistent with these previous studies, demonstrating that
p-ERK was distinctly elevated in the rat lungs exposed to MCT, and
FMN administration markedly decreased the activated ERK induced by
MCT.
The aforementioned results suggest that FMN can
improve MCT-induced PAH by restraining the ERK signaling pathway
mediated by pulmonary vascular remodeling, at least to a certain
extent. Based on previous studies, NF-κB is a multi-functional
transcription factor and can be activated in idiopathic PAH
patients and MCT-induced PAH rats (45,46).
In addition, activated NF-κB can promote inflammation and ECM
deposition, and participates in various pathophysiological and
pathological activities (47,48).
A recent study has indicated that right ventricular hypertrophy
also has certain effects on activated NF-κB in MCT-induced PAH rats
(25). Correspondingly, the
results from the present study indicated that MCT significantly
enhanced the activated NF-κB in rat lungs, while FMN significantly
suppressed this activation, which suggests another underlying
action mechanism of FMN in the suppression of PAH.
Based on results from the present study, injection
of MCT induced the increase in TGFβ1, MMPs, ECM proteins and
inflammatory cytokines, which leads to consideration of their
intrinsic association. According to previous studies, TGFβ1 is
closely related to MMPs and inflammatory factors, and the
relationship between ECM and MMPs is also clear (49,50).
Therefore, it was hypothesized that increased TGFβ1 stimulates the
production of MMPs and inflammatory factors, and MMPs subsequently
affect the synthesis and decomposition of ECM, and this process may
be mediated by the ERK and NF-κB signaling pathways (48,49).
The present study suggested that FMN may play a protective role in
MCT-induced PAH based on the aforementioned hypothesis. In order to
clarify this hypothesis, more cell experiments are required.
In conclusion, the present study demonstrated that
FMN treatment could delay the MCT-induced PAH progression in rats
and alleviate pulmonary vascular remodeling and right ventricular
hypertrophy. These positive effects of FMN may be associated with
suppression of ECM deposition and inflammation, at least in part,
and the suppression of ERK and NF-κB signaling.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YW, CC and CZ designed the study. YW, CC, LY and YX
performed the experiments and analyzed the data, and YW wrote the
manuscript. YX were responsible for data acquisition and provided
technological assistance. HZ provided pathological assistance, and
was involved in the data analysis and interpretation. CZ
participated in critical revisions of the manuscript. All of the
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
The experimental procedure was approved by The
Ethics Review of Animal Use Application of the Fifth Affiliated
Hospital of Wenzhou Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
De Jesus Perez VA: Molecular pathogenesis
and current pathology of pulmonary hypertension. Heart Fail Rev.
21:239–257. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Maciver DH, Adeniran I, Maciver IR, Revell
A and Zhang H: Physiological mechanisms of pulmonary hypertension.
Am Heart J. 180:1–11. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Montani D, Chaumais MC, Guignabert C,
Günther S, Girerd B, Jaïs X, Algalarrondo V, Price LC, Savale L,
Sitbon O, et al: Targeted therapies in pulmonary arterial
hypertension. Pharmacol Ther. 141:172–191. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rabinovitch M, Guignabert C, Humbert M and
Nicolls MR: Inflammation and immunity in the pathogenesis of
pulmonary arterial hypertension. Circ Res. 115:165–175. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huertas A, Perros F, Tu L, Cohen-Kaminsky
S, Montani D, Dorfmuller P, Guignabert C and Humbert M: Immune
dysregulation and endothelial dysfunction in pulmonary arterial
hypertension: A complex interplay. Circulation. 129:1332–1340.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Thenappan T, Chan SY and Weir E.K: Role of
extracellular matrix in the pathogenesis of pulmonary arterial
hypertension. Am J Physiol Heart Circ Physiol. 315:H1322–H1331.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li W, Guo A, Wang L, Kong Q, Wang R, Han L
and Zhao C: Expression of peptide fragments from proADM and
involvement of mitogen-activated protein kinase signaling pathways
in pulmonary remodeling induced by high pulmonary blood flow.
Congenit Anom (Kyoto). 56:28–34. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li L, Wei C, Kim IK, Janssen-Heininger Y
and Gupta S: Inhibition of nuclear factor-κB in the lungs prevents
monocrotaline-induced pulmonary hypertension in mice. Hypertension.
63:1260–1269. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tannenberg P and Tran-Lundmark K: The
extracellular matrix in early and advanced pulmonary arterial
hypertension. Am J Physiol Heart Circ Physiol. 315:H1684–H1686.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Eble JA and Niland S: The extracellular
matrix of blood vessels. Curr Pharm Des. 15:1385–1400. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lammers SR, Kao PH, Qi HJ, Hunter K,
Lanning C, Albietz J, Hofmeister S, Mecham R, Stenmark KR and
Shandas R: Changes in the structure-function relationship of
elastin and its impact on the proximal pulmonary arterial mechanics
of hypertensive calves. Am J Physiol Heart Circ Physiol.
295:H1451–H1459. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ambalavanan N, Nicola T, Li P, Bulger A,
Murphy-Ullrich J, Oparil S and Chen YF: Role of matrix
metalloproteinase-2 in newborn mouse lungs under hypoxic
conditions. Pediatr Res. 63:26–32. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Novotná J and Herget J: Possible role of
matrix metalloproteinases in reconstruction of peripheral pulmonary
arteries induced by hypoxia. Physiol Res. 51:323–334.
2002.PubMed/NCBI
|
|
14
|
Chelladurai P, Seeger W and Pullamsetti
SS: Matrix metalloproteinases and their inhibitors in pulmonary
hypertension. Eur Respir J. 40:766–782. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Perrotta F, Nigro E, Mollica M,
Costigliola A, D'Agnano V, Daniele A, Bianco A and Guerra G:
Pulmonary hypertension and obesity: Focus on adiponectin. Int J Mol
Sci. 20:E9122019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Perros F, Montani D, Dorfmuller P,
Durand-Gasselin I, Tcherakian C, Le Pavec J, Mazmanian M, Fadel E,
Mussot S, Mercier O, et al: Platelet-derived growth factor
expression and function in idiopathic pulmonary arterial
hypertension. Am J Respir Crit Care Med. 178:81–88. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Krenn L and Paper DH: Inhibition of
angiogenesis and inflammation by an extract of red clover
(Trifolium pratense L.). Phytomedicine. 16:1083–1088. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang S, Tang X, Tian J, Li C, Zhang G,
Jiang W and Zhang Z: Cardioprotective effect of sulphonated
formononetin on acute myocardial infarction in rats. Basic Clin
Pharmacol Toxicol. 108:390–395. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sun T, Wang J, Huang LH and Cao YX:
Antihypertensive effect of formononetin through regulating the
expressions of eNOS, 5-HT2A/1B receptors and α1-adrenoceptors in
spontaneously rat arteries. Eur J Pharmacol. 699:241–249. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jin YM, Xu TM, Zhao YH, Wang YC and Cui
MH: In vitro and in vivo anti-cancer activity of formononetin on
human cervical cancer cell line HeLa. Tumour Biol. 35:2279–2284.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang X, Bi L, Ye Y and Chen J:
Formononetin induces apoptosis in PC-3 prostate cancer cells
through enhancing the Bax/Bcl-2 ratios and regulating the p38/Akt
pathway. Nutr Cancer. 66:656–661. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu Y, He J, Chen X, Li J, Shen M, Yu W,
Yang Y and Xiao Z: The proapoptotic effect of formononetin in human
osteosarcoma cells: involvement of inactivation of ERK and Akt
pathways. Cell Physiol Biochem. 34:637–645. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huh JE, Seo DM, Baek YH, Choi DY, Park DS
and Lee JD: Biphasic positive effect of formononetin on metabolic
activity of human normal and osteoarthritic subchondral
osteoblasts. Int Immunopharmacol. 10:500–507. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Alauddin, Chaturvedi S, Malik MY, Azmi L,
Shukla I, Naseem Z, Rao C and Agarwal NK: Formononetin and
biochanin A protects against ritonavir induced hepatotoxicity via
modulation of NfκB/pAkt signaling molecules. Life Sci. 213:174–182.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhu N, Zhao X, Xiang Y, Ye S, Huang J, Hu
W, Lv L and Zeng C: Thymoquinone attenuates monocrotaline-induced
pulmonary artery hypertension via inhibiting pulmonary arterial
remodeling in rats. Int J Cardiol. 221:587–596. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ichimura K, Matoba T, Koga JI, Nakano K,
Funamoto D, Tsutsui H and Egashira K: Nanoparticle-mediated
targeting of pitavastatin to small pulmonary arteries and
leukocytes by intravenous administration attenuates the progression
of monocrotaline-induced established pulmonary arterial
hypertension in rats. Int Heart J. 59:1432–1444. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Seimetz M, Parajuli N, Pichl A, Veit F,
Kwapiszewska G, Weisel FC, Milger K, Egemnazarov B, Turowska A,
Fuchs B, et al: Inducible NOS inhibition reverses
tobacco-smoke-induced emphysema and pulmonary hypertension in mice.
Cell. 147:293–305. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wu F, Yao W, Yang J, Zhang M, Xu Y, Hao Y,
Yan L, Niu Y, Sun T, Yu J and Zhou R: Protective effects of
aloperin on monocroline-induced pulmonary hypertension via
regulation of Rho A/Rho kinsase pathway in rats. Biomed
Pharmacother. 95:1161–1168. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li Q, Wang J, Zhu X, Zeng Z, Wu X, Xu Y,
Xie J and Yu J: Dihydromyricetin prevents monocrotaline-induced
pulmonary arterial hypertension in rats. Biomed Pharmacother.
96:825–833. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schäfer M, Ivy DD, Abman SH, Stenmark K,
Browne LP, Barker AJ, Mitchell MB, Morgan GJ, Wilson N, Shah A, et
al: Differences in pulmonary arterial flow hemodynamics between
children and adults with pulmonary arterial hypertension as
assessed by 4D-flow CMR studies. Am J Physiol Heart Circ Physiol.
316:H1091–H1104. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Han X, Zhang Y, Zhou Z, Zhang X and Long
Y: Hydroxysafflor yellow A improves established
monocrotaline-induced pulmonary arterial hypertension in rats. J
Int Med Res. 44:569–584. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bai Y, Wang HM, Liu M, Wang Y, Lian GC,
Zhang XH, Kang J and Wang HL: 4-Chloro-DL-phenylalanine protects
against monocrotaline-induced pulmonary vascular remodeling and
lung inflammation. Int J Mol Med. 33:372–382. 2014. View Article : Google Scholar
|
|
33
|
Long L, Crosby A, Yang X, Southwood M,
Upton PD, Kim DK and Morrell NW: Altered bone morphogenetic protein
and transforming growth factor-beta signaling in rat models of
pulmonary hypertension: Potential for activin receptor-like
kinase-5 inhibition in prevention and progression of disease.
Circulation. 119:566–576. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hodges MM, Zgheib C, Xu J, Hu J, Dewberry
LC, Hilton SA, Allukian MW, Gorman JH III, Gorman RC and Liechty
KW: Differential expression of transforming growth factor-β1 is
associated with fetal regeneration after myocardial infarction. Ann
Thorac Surg. 108:59–66. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tang PM, Nikolic-Paterson DJ and Lan HY:
Macrophages: Versatile players in renal inflammation and fibrosis.
Nat Rev Nephrol. 15:144–158. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Crosswhite P and Sun Z: Nitric oxide,
oxidative stress and inflammation in pulmonary arterial
hypertension. J Hypertens. 28:201–212. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gao Y, Lu J, Zhang Y, Chen Y, Gu Z and
Jiang X: Baicalein attenuates bleomycin-induced pulmonary fibrosis
in rats through inhibition of miR-21. Pulm Pharmacol Ther.
26:649–654. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mathew R: Inflammation and pulmonary
hypertension. Cardiol Rev. 18:67–72. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lepetit H, Eddahibi S, Fadel E, Frisdal E,
Munaut C, Noel A, Humbert M, Adnot S, D'Ortho MP and Lafuma C:
Smooth muscle cell matrix metalloproteinases in idiopathic
pulmonary arterial hypertension. Eur Respir J. 25:834–842. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Keller KE, Aga M, Bradley JM, Kelley MJ
and Acott TS: Extracellular matrix turnover and outflow resistance.
Exp Eye Res. 88:676–682. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Dorfmüller P, Perros F, Balabanian K and
Humbert M: Inflammation in pulmonary arterial hypertension. Eur
Respir J. 22:358–363. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bai Y, Li ZX, Wang HL, Lian GC and Wang Y:
The protective effects of PCPA against monocrotaline-induced
pulmonary arterial hypertension are mediated through the
downregulation of NFAT-1 and NF-κB. Int J Mol Med. 40:155–163.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kiss T and Kovacs K, Komocsi A, Tornyos A,
Zalan P, Sumegi B, Gallyas F Jr and Kovacs K: Novel mechanisms of
sildenafil in pulmonary hypertension involving
cytokines/chemokines, MAP kinases and Akt. PLoS One. 9:e1048902014.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kovacs L, Cao Y, Han W, Meadows L,
Kovacs-Kasa A, Kondrikov D, Verin AD, Barman SA, Dong Z, Huo Y and
Su Y: PFKFB3 in smooth muscle promotes vascular remodeling in
pulmonary arterial hypertension. Am J Respir Crit Care Med.
200:617–627. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sawada H, Mitani Y, Maruyama J, Jiang BH,
Ikeyama Y, Dida FA, Yamamoto H, Imanaka-Yoshida K, Shimpo H,
Mizoguchi A, et al: A nuclear factor-kappaB inhibitor pyrrolidine
dithiocarbamate ameliorates pulmonary hypertension in rats. Chest.
132:1265–1274. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Price LC, Caramori G, Perros F, Meng C,
Gambaryan N, Dorfmuller P, Montani D, Casolari P, Zhu J, Dimopoulos
K, et al: Nuclear factor κ-B is activated in the pulmonary vessels
of patients with end-stage idiopathic pulmonary arterial
hypertension. PLoS One. 8:e754152013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang AW, Song L, Miao J, Wang HX, Tian C,
Jiang X, Han QY, Yu L, Liu Y, Du J, et al: Baicalein attenuates
angiotensin II-induced cardiac remodeling via inhibition of
AKT/mTOR, ERK1/2, NF-κB, and calcineurin signaling pathways in
mice. Am J Hypertens. 28:518–526. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Morin C, Hiram R, Rousseau E, Blier PU and
Fortin S: Docosapentaenoic acid monoacylglyceride reduces
inflammation and vascular remodeling in experimental pulmonary
hypertension. Am J Physiol Heart Circ Physiol. 307:H574–H586. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Peng X, Li HX, Shao HJ, Li GW, Sun J, Xi
YH, Li HZ, Wang XY, Wang LN, Bai SZ, et al: Involvement of
calcium-sensing receptors in hypoxia-induced vascular remodeling
and pulmonary hypertension by promoting phenotypic modulation of
small pulmonary arteries. Mol Cell Biochem. 396:87–98. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhou XM, Wang GL, Wang XB, Liu L, Zhang Q,
Yin Y, Wang QY, Kang J and Hou G: GHK peptide inhibits
bleomycin-induced pulmonary fibrosis in mice by suppressing
TGFβ1/Smad-mediated epithelial-to-mesenchymal transition. Front
Pharmacol. 8:9042017. View Article : Google Scholar : PubMed/NCBI
|