Introduction
Intervertebral disc degeneration (IDD) is a major
cause of low back pain (1).
Intervertebral discs consist of cartilage endplate and nucleus
pulposus (NP) and annulus fibrosus (AF) cells. Cartilage endplates
are required for stress distribution and nutrient transport to the
intervertebral discs (2,3). It has been reported that degeneration
of the endplates may result in IDD (4,5).
Since the endplate capillaries are responsible for supplying
nutrients to the intervertebral discs, degeneration of the
endplates is thought to be a risk factor for IDD, as it can hinder
the transport of nutrients and eventually lead to IDD (6–8).
Therefore, CEPs may be particularly susceptible to the vasculopathy
associated with DM and increasing evidence has revealed that DM
plays an important role in CEP degeneration (2,9–12).
Fields et al revealed that endplate sclerosis, increased
endplate thickness and decreased endplate porosity were observed in
rats with type 2 diabetes (9).
Furthermore, high-glucose was revealed to induce the apoptosis of
rat cartilage endplate cells in a dose- and time-dependent manner
(12).
Long non-coding RNAs (lncRNAs) are a class of
non-coding RNA molecules >200 nucleotides in length that play a
key role in a variety of cellular processes. The dysregulation of
lncRNAs is closely related to the occurrence and development of
human diseases, including cancer, degenerative neurological
diseases, and diabetic microangiopathy (13–16).
The long non-coding RNA metastasis associated lung adenocarcinoma
transcript 1 (MALAT1) is highly phylogenetic and conserved in
mammals. MALAT1 is highly expressed in a variety of tumours and
promotes the proliferation, metastasis and invasion of tumour cells
(17,18). In recent years, studies have
revealed that MALAT1 also plays an important role in
diabetes-related complications. It has been reported that MALAT1
contributes to certain diabetes-related complications, including
cataracts, retinopathy, cardiomyopathy, gastropathy and kidney
disease (19–24).
Mitogen-activated protein kinase (MAPK), a
serine/threonine kinase, includes three major classes: p38,
extracellular signal-regulated kinases (ERKs) and c-jun N-terminal
kinase. The p38/MAPK signalling pathway is an important signalling
transduction pathway (25). It has
been reported that activation of the p38/MAPK signalling pathway is
closely related to apoptosis (26,27).
Studies have demonstrated that aberrant MALAT1 expression causes a
series of cellular effects via the p38/MAPK signalling pathway
(24,28).
The aforementioned studies indicated that MALAT1
plays an essential role in several diabetes-related complications.
However, whether MALAT1 affects CEP degeneration or how MALAT1
contributes to CEP degeneration have not been determined.
Therefore, the present study investigated the effect and mechanism
of lncRNA-MALAT1 underlying high glucose-induced apoptosis in rat
CEP cells.
Materials and methods
Rat CEP cell culture and
treatment
All animal experiments were approved by the Ethics
Committee on Animal Experiments of Fudan University (Shanghai,
China). A total of three 12-week-old male Sprague-Dawley rats were
anesthetized with a single intraperitoneal injection of 1% sodium
pentobarbital (40 mg/kg; Sigma-Aldrich; Merck KGaA) and then
euthanized by cervical dislocation. Death was confirmed by checking
breathing and heartbeat. Verification of death was supplemented by
percutaneous cardiac puncture before intervertebral discs (L1-L6)
were harvested from rats. CEPs were carefully dissected under a
microscope and homogenized. The tissue was digested with 0.25%
trypsin (Sigma-Aldrich; Merck KGaA) for 2 h followed by treatment
with 0.02% collagenase type II (Sigma-Aldrich; Merck KGaA) at 37°C
for 12 h. Cells were cultured in Dulbecco's Modified Eagle's Medium
(DMEM; Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10%
foetal bovine serum (FBS; Thermo Fisher Scientific, Inc.) at 37°C
and 5% CO2. The chondrocytic phenotype of the primary CEP cells was
confirmed by toluidine blue staining (Shanghai Haoran Biotechnology
Co., Ltd.). The cells were then trypsinized and sub-cultured on
6-well plates (5×105 cells/well) for subsequent experimentation.
After reaching ~80% confluence, CEP cells were cultured in DMEM
supplemented with 10% FBS (5 mM glucose; control) or high-glucose
DMEM supplemented with 10% FBS (25 mM glucose) for 24 and 72 h. The
glucose concentrations used in the present study were based on the
literature (24).
RNA interference and cell
transfection
The stealth RNAi specifically targeting MALAT1 was
designed and synthesized by Shanghai Genechem Co., Ltd. The
lncRNA-MALAT1-RNAi sequence was as follows:
5′-GAGGUGUAAAGGGAUUUAUTT-3′. lncRNA-MALAT1-RNAi was transfected
into rat CEP cells using Lipofectamine 2000 (Thermo Fisher
Scientific, Inc.). Following 48 h in culture, the cells were
collected for future experiments.
Reverse transcription quantitative
polymerase chain reaction (RT qPCR)
MALAT1 mRNA expression levels in the different
groups were measured by RT-qPCR. Total RNA was reversed-transcribed
into cDNA using the advantage rT-for-PCR kit (Takara Biotechnology
Co., Ltd). qPCR was performed using a 20-µl reaction system
consisting of 10 µl SYBR Green mix (Thermo Fisher Scientific,
Inc.), 1 µl RT-primer, 1 µl template DNA and 8 µl DEPC-treated
water in a TP800 Thermal Cycler Dice (Takara Biotechnology Co.,
Ltd). The primer sequences were as follows: Rat MALAT1 forward,
5′-GTGATGCGAGTTGTTCTCCG-3′ and reverse, 5′-CTGGCTGCCTCAATGCCTAC-3′;
rat GAPDH forward, 5′-ACAGTCAGCCGCATCTTCTT-3′ and reverse,
5′-GACAAGCTTCCCGTTCTCAG-3′. The following thermocycling conditions
were used: Melting at 95°C for 10 sec, annealing at 95°C for 5 sec,
and extension at 60°C for 20 sec for 45 cycles. MALAT1 expression
was quantified using the 2−ΔΔCq method and normalized to
GAPDH (29).
Determination of apoptosis in CEP
cells
Apoptosis was detected using an Annexin
V-fluorescein isothiocyanate (FITC)/ propidium iodide (PI)
Apoptosis Detection kit (Nanjing KeyGen Biotech Co., Ltd.). Rat CEP
cells were collected and washed with ice-cold phosphate-buffered
saline (PBS; Beyotime Institute of Biotechnology) and resuspended
in binding buffer (100 µl). The cells were incubated with Annexin
V-FITC (5 µl) and PI (5 µl) for 15 min at room temperature in the
dark. Apoptosis was subsequently detected using a FACScan flow
cytometer (BD Biosciences).
Western blot assay
Western blotting was used to assess the expression
levels of p38 and phosphorylated (p)-p38. After 24 or 72 h of
treatment, total protein was extracted from CEP cells using
radioimmunoprecipitation assay buffer (Sigma-Aldrich; Merck KGaA).
The samples were sonicated for 10 sec. Following centrifugation at
2,000 × g for 15 min at 4°C, protein concentrations were measured
using a bicinchoninic acid protein assay kit (Thermo Fisher
Scientific, Inc.). Total protein (20 µg per sample) was
subsequently separated by SDS-PAGE on a 12% gel and then
transferred onto nitrocellulose membranes (EMD Millipore). The
membranes were incubated overnight at 4°C with the following
antibodies: anti-p53 (1:1,000; cat. no. 32523), anti-p53 (phosopho
ser15) (1:1,000; cat. no. 9284), and anti-β-actin (1:1,000; cat.
no. 4970; all from Cell Signalling Technology Inc.). After washing
with TBST for 30 min, the membranes were incubated with a
corresponding secondary antibody (1:5,000; cat. no. 7074; Cell
Signaling Technology Inc.) for 2 h at room temperature.
Electrochemiluminescence Plus (Tanon Science and Technology Co.,
Ltd.) was used to visualize the protein bands. Densitometry
analysis was performed using ImageJ software (version 1.8.0;
National Institutes of Health) with β-actin as the internal
control.
Statistical analysis
Each experiment was performed in triplicate and data
were expressed as the mean ± standard deviation. Statistical
analyses were performed using SPSS software (version 20.0; IBM
Corp.). Statistical comparisons among different groups were
performed using the one-way analysis of variance followed by
Bonferroni's multiple comparison test. Moreover, qPCR data were
analysed using analysis of covariance. P<0.05 was considered to
indicate a statistically significant difference.
Results
High-glucose promotes apoptosis of rat
CEP cells and upregulates MALAT1 expression
Toluidine blue staining was performed to confirm the
CEP cell phenotype. Light blue staining in the cytoplasm was
observed, which is considered to be a feature of chondrocyte
proteoglycans. Therefore, the results demonstrated that cells
cultured from the CEP maintained a chondrocytic phenotype (Fig. 1).
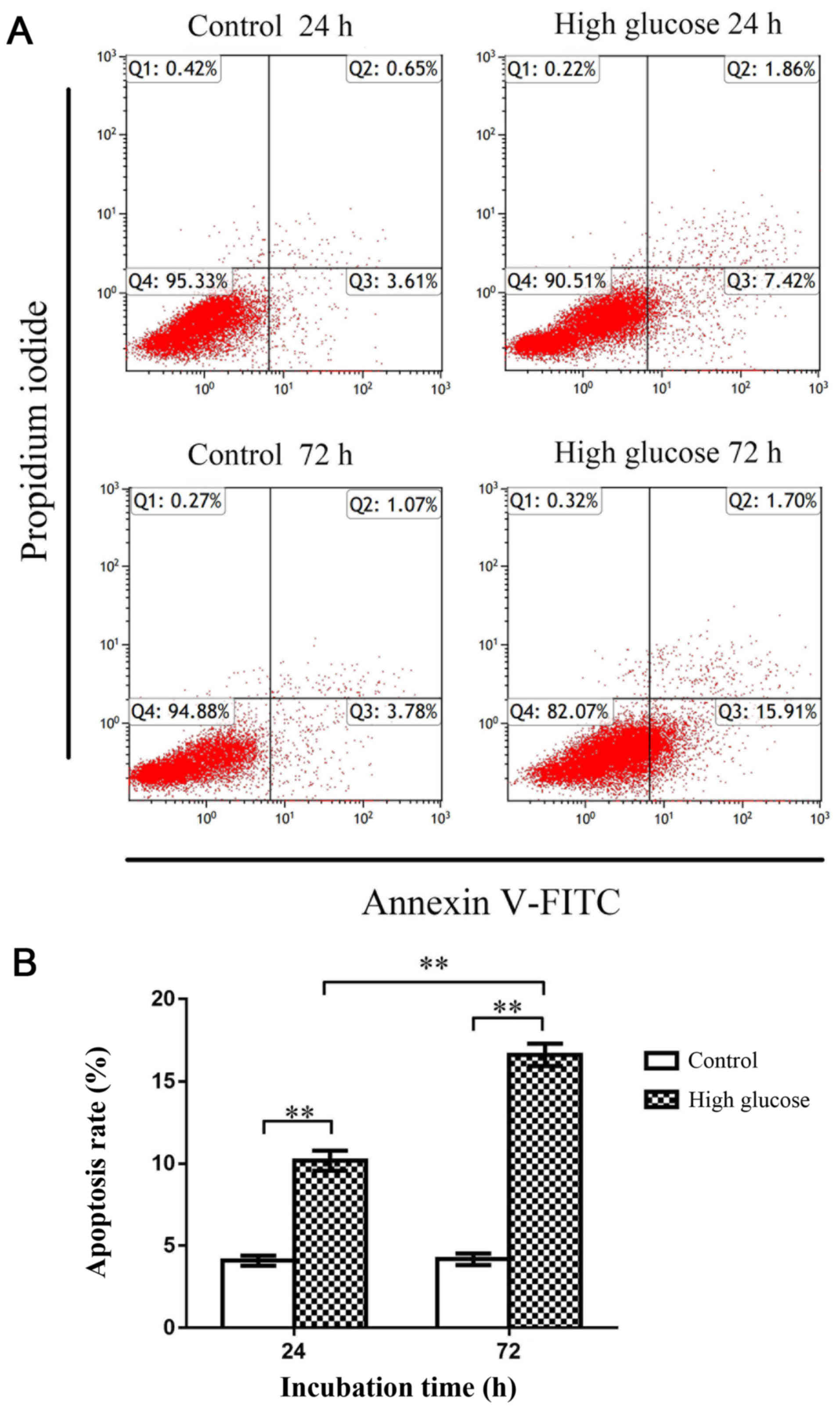
Flow cytometry was subsequently used to investigate
apoptosis of rat CEP cells. The apoptosis rate in the high-glucose
group (25 mM glucose) was significantly increased compared with the
control group (5 mM glucose) at both 24 and 72 h (P<0.01),
indicating that high-glucose conditions promoted rat CEP cell
apoptosis (Fig. 2). Moreover,
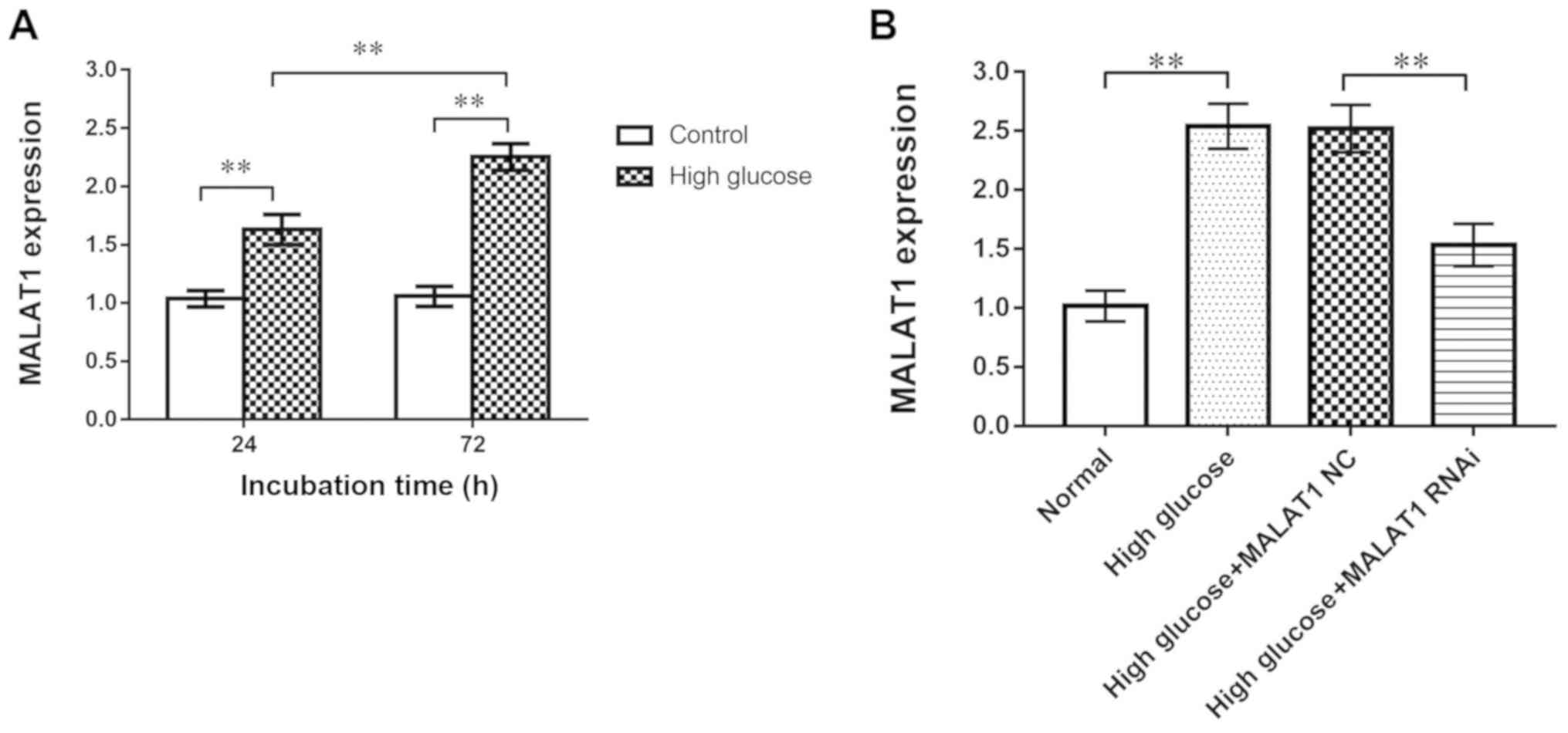
RT-qPCR revealed that the expression of MALAT1 in the high-glucose
group was significantly increased compared with the control group
at 24 and 72 h (P<0.01), indicating that the expression of
MALAT1 was upregulated in rat CEP cells treated with high glucose
(Fig. 3A).
MALAT1 knockout decreases the
expression levels of MALAT1
CEP cells in the four groups [control, high glucose,
high glucose + MALAT1 negative control (NC), high glucose + MALAT1
RNAi] were cultured for 72 h. RT-qPCR was used to analyse the siRNA
transfection efficiency in rat CEP cells. MALAT1 RNAi significantly
decreased the expression of MALAT1 in CEP cells (P<0.01). No
difference in expression was observed between the high glucose and
the high glucose + MALAT1 NC groups (Fig. 3B).
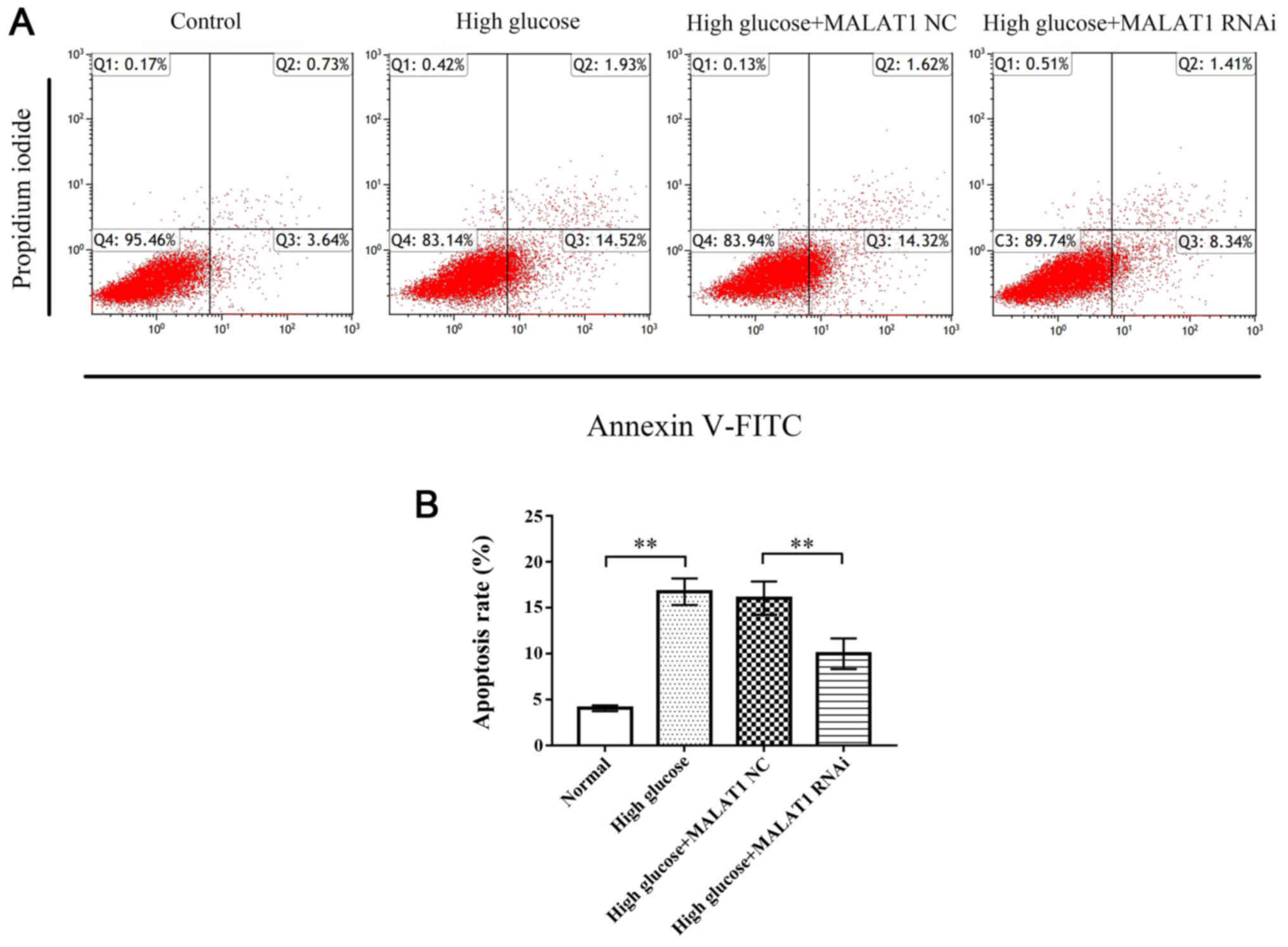
MALAT1 knockout decreases the
apoptosis of rat CEP cells
Flow cytometric analysis revealed that the apoptosis
rate of CEP cells in the high glucose and high glucose + MALAT1 NC
groups was significantly increased compared with the control group
(P<0.01), while the apoptosis rate of CEP cells in the high
glucose + MALAT1 RNAi group was significantly decreased compared
with the high glucose and high glucose + MALAT1 NC groups
(P<0.01; Fig. 4).
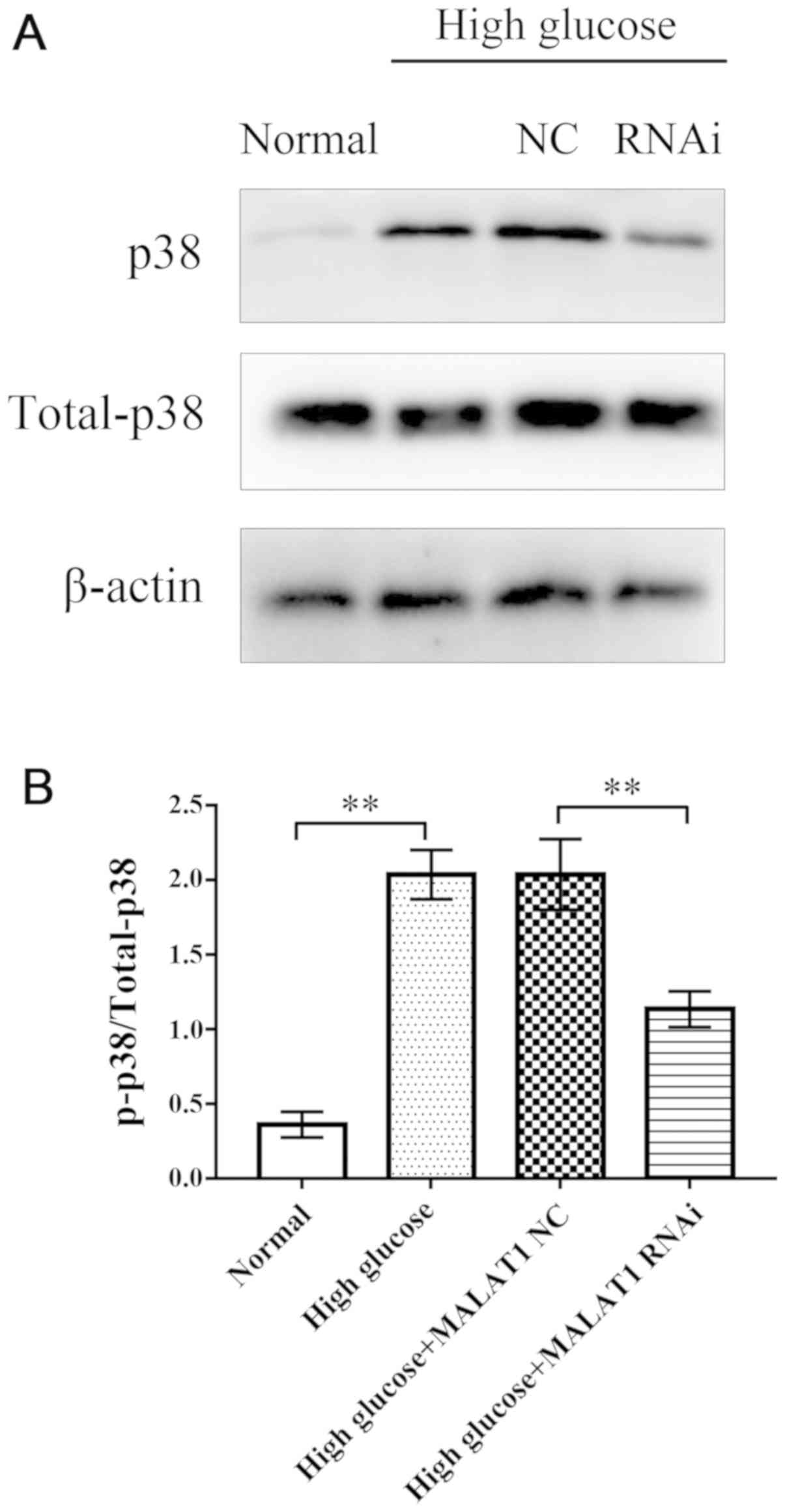
MALAT1 knockout decreases the
expression levels of total p38 and p-p38
After 72 h of treatment, western blotting was used
to assess the expression levels of total p38 and p-p38 in rat CEP
cells in the four groups. Western blot analysis revealed that in
the high glucose and high glucose + MALAT1 NC groups, the
expression levels of total p38 and p-p38 were significantly
increased compared with the control group (P<0.01). However, the
expression levels of total p38 and p-p38 in CEP cells treated with
high glucose + MALAT1 RNAi were significantly decreased compared
with CEP cells treated with high glucose or high glucose + MALAT1
NC (P<0.01). No significant difference was observed between the
high glucose and the high glucose + MALAT1 NC groups (Fig. 5).
Discussion
Previous study has revealed that apoptosis of CEP
cells induced by high glucose contributes to IDD (12). MALAT1 plays an essential role in a
number of diabetes-related complications (20–24).
However, the role of MALAT1 in diabetes-associated IDD has not been
determined. The present study revealed that 25 mM glucose promoted
rat CEP cell apoptosis. Furthermore, high-glucose concentrations
upregulated MALAT1 expression in rat CEP cells. MALAT1 knockout
significantly inhibited high glucose-induced apoptosis of CEP
cells. High-glucose treatment increased the expression of MALAT1,
p38 and p-p38. MALAT1 knockdown significantly decreased MALAT1, p38
and p-p38 expression, indicating that MALAT1 promoted high
glucose-induced apoptosis of rat CEP cells by activating the
p38/MAPK signalling pathway.
While DM has not been characterized as a direct
cause of IDD, a growing body of evidence suggests that diabetes
affects the initiation and development of IDD to a certain extent
(9–12,30).
Clinical studies have revealed that the incidence of DM in patients
undergoing surgery for lumbar disc degenerative disease was
significantly higher than that of patients undergoing surgery for
other reasons (11,31). Patients with diabetes experience
decreased physiological function of intervertebral discs compared
with patients without DM. Recent studies have also revealed that
high glucose concentrations lead to apoptosis of intervertebral
disc cells, including NP and AF cells (31,32).
A previous study demonstrated that high glucose concentrations can
enhance oxidative stress, impair mitochondrial functions and
ultimately lead to rat CEP cell apoptosis in a concentration- and
time-dependent (12).
Recent studies have revealed that MALAT1 contributes
to certain diabetes-related complications, including cerebral
ischaemic reperfusion injury, atherosclerosis, cataracts,
retinopathy, cardiomyopathy, gastropathy and kidney disease. It has
been reported that the expression of MALAT1 is upregulated in
multiple cells and tissues in patients and animals with diabetes
(20–24). Increased MALAT1 expression is an
important reason for the initiation and development of
diabetes-related diseases. Therefore, MALAT1 is considered as both
a therapeutic target and a potential biomarker for DM and
diabetes-related diseases. However, the association between MALAT1
and diabetes-related IDD remains unclear. Therefore, the present
study investigated the roles of MALAT1 in diabetes-related IDD.
The present study demonstrated the roles of MALT1 in
rat CEP cell apoptosis induced by high glucose. Previous studies on
diabetes-associated IDD have investigated the use of 50, 100 and
200 mM glucose concentrations, while 25 mM glucose served as a
control. To clarify the role of MALAT1 in diabetes-associated IDD
in the present study, 25 mM glucose was selected and 5 mM glucose
was used as a control. The results revealed that 25 mM glucose
induced rat CEP cell apoptosis, which is consistent with results
obtained in a previous study (12). Compared with previous studies, the
present study investigated the role of high glucose in promoting
rat CEP cell apoptosis by using more physiologically relevant
glucose concentrations (5 and 25 mM). MALAT1 expression was
upregulated in CEP cells cultured with high glucose concentrations,
which is similar to what occurs in other tissues and cells in DM
(21–24). MALAT1 expression was downregulated
following MALAT1 knockout. Flow cytometric analysis revealed that
rat CEP cell apoptosis in the MALAT RNAi group was significantly
decreased compared with the high glucose and MALAT1 NC groups,
indicating that inhibition of MALAT1 expression effectively
prevented rat CEP cell apoptosis induced by high glucose
concentrations. This finding also indicated that MALAT1 serves a
crucial role in high glucose-induced rat CEP cell apoptosis.
Western blotting revealed that the expression levels of p38 and
p-p38 significantly increased in rat CEP cells incubated with 25 mM
glucose at 72 h but decreased when MALAT1 expression was inhibited
by MALAT1 RNAi, suggesting that MALAT1 affects rat CEP cell
apoptosis by activating the p38/MAPK signalling pathway.
In conclusion, the present study revealed that
incubation with a high glucose concentration induced rat CEP cell
apoptosis and increased MALAT1 expression. However, considering the
multifaceted effects of high glucose on CEP cells, further research
is required to explore influences such as aging and necrosis.
Diabetic patients are in different degrees of hypertonic state,
which may be harmful and cannot be ignored (33–35).
Considering that hypertonicity is one of the effects of high
glucose, we have not eliminated the effects of hypertonicity in the
present study. To better understand the role of high glucose in
IDD, some measures will be taken to normalize the hyperosmolality
of hyperglycemia in future research. MALAT1 promoted high
glucose-induced rat CEP cell apoptosis by activating the p38/MAPK
signalling pathway. However, the p38 MAPK signalling pathway is not
the only mechanism by which MALAT1 promotes rat CEP cell apoptosis.
Therefore, further experiments should be performed to elucidate the
mechanisms of MALAT1 in diabetes-related IDD. The results obtained
in the present study indicated that MALAT1 may serve as a
therapeutic target and biomarker in diabetes-related IDD.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Shanghai
Municipality Health Bureau (grant no. 201640101) and Jinshan
Municipality Health Bureau (grant no. JSKJ-KTMS-2018-01).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
QZ and JW designed the study. ZJ, DL and LD
conducted the majority of the experiments. ZJ, MB and WL assisted
with the analysis and interpretation of data. ZJ and WL prepared
the manuscript. All authors were involved in drafting the article
or revising it critically for important scientific content. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Ethics
Committee on Animal Experiments of Fudan University (Shanghai,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
DM
|
diabetes mellitus
|
|
IDD
|
intervertebral disc degeneration
|
|
CEP
|
cartilage endplate
|
|
lncRNAs
|
long non-coding RNAs
|
|
MALAT1
|
metastasis associated lung
adenocarcinoma transcript 1
|
|
MAPK
|
mitogen-activated protein kinase
|
|
ERK
|
extracellular signal-regulated
kinase
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
References
|
1
|
Dowdell J, Erwin M, Choma T, Vaccaro A,
Iatridis J and Cho SK: Intervertebral Disk Degeneration and Repair.
Neurosurgery. 80 (Suppl 3):S46–S54. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gu W, Zhu Q, Gao X and Brown MD:
Simulation of the Progression of Intervertebral Disc Degeneration
due to Decreased Nutrition Supply. Spine. 39:E1411–E1417. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhu Q, Gao X, Levene HB, Brown MD and Gu
W: Influences of Nutrition Supply and Pathways on the Degenerative
Patterns in Human Intervertebral Disc. Spine. 41:568–576. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Määttä JH, Kraatari M, Wolber L, Niinimäki
J, Wadge S, Karppinen J and Williams FM: Vertebral endplate change
as a feature of intervertebral disc degeneration: A heritability
study. Eur Spine J. 23:1856–1862. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang Y, Videman T and Battié MC: ISSLS
prize winner: Lumbar vertebral endplate lesions: associations with
disc degeneration and back pain history. Spine. 37:1490–1496. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kang R, Li H, Ringgaard S, Rickers K, Sun
H, Chen M, Xie L and Bünger C: Interference in the endplate
nutritional pathway causes intervertebral disc degeneration in an
immature porcine model. Int Orthop. 38:1011–1017. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yin S, Du H, Zhao W, Ma S, Zhang M, Guan M
and Liu M: Inhibition of both endplate nutritional pathways results
in intervertebral disc degeneration in a goat model. J Orthop Surg
Res. 14:1382019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hutton WC, Murakami H, Li J, Elmer WA,
Yoon ST, Minamide A, Akamaru T and Tomita K: The effect of blocking
a nutritional pathway to the intervertebral disc in the dog model.
J Spinal Disord Tech. 17:53–63. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fields AJ, Berg-Johansen B, Metz LN,
Miller S, La B, Liebenberg EC, Coughlin DG, Graham JL, Stanhope KL,
Havel PJ, et al: Alterations in intervertebral disc composition,
matrix homeostasis and biomechanical behavior in the UCD-T2DM rat
model of type 2 diabetes. J Orthop Res. 33:738–746. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Agius R, Galea R and Fava S: Bone mineral
density and intervertebral disc height in type 2 diabetes. J
Diabetes Complications. 30:644–650. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sakellaridis N: The influence of diabetes
mellitus on lumbar intervertebral disk herniation. Surg Neurol.
66:152–154. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jiang Z, Lu W, Zeng Q, Li D, Ding L and Wu
J: High glucose-induced excessive reactive oxygen species promote
apoptosis through mitochondrial damage in rat cartilage endplate
cells. J Orthop Res. 36:2476–2483. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jathar S, Kumar V, Srivastava J and
Tripathi V: Technological Developments in lncRNA Biology. Adv Exp
Med Biol. 1008:283–323. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kung JT, Colognori D and Lee JT: Long
noncoding RNAs: Past, present, and future. Genetics. 193:651–669.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jarroux J, Morillon A and Pinskaya M:
History, Discovery, and Classification of lncRNAs. Adv Exp Med
Biol. 1008:1–46. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen WK, Yu XH, Yang W, Wang C, He WS, Yan
YG, Zhang J and Wang WJ: lncRNAs: Novel players in intervertebral
disc degeneration and osteoarthritis. Cell Prolif. 50:e123132017.
View Article : Google Scholar
|
|
17
|
Zhang X, Hamblin MH and Yin KJ: The long
noncoding RNA Malat1: Its physiological and pathophysiological
functions. RNA Biol. 14:1705–1714. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li ZX, Zhu QN, Zhang HB, Hu Y, Wang G and
Zhu YS: MALAT1: A potential biomarker in cancer. Cancer Manag Res.
10:6757–6768. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Abdulle LE, Hao JL, Pant OP, Liu XF, Zhou
DD, Gao Y, Suwal A and Lu CW: MALAT1 as a Diagnostic and
Therapeutic Target in Diabetes-Related Complications: A Promising
Long-Noncoding RNA. Int J Med Sci. 16:548–555. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gong Y, Zhu Y, Zhu B, Si X, Heng D, Tang
Y, Sun X and Lin L: LncRNA MALAT1 is up-regulated in diabetic
gastroparesis and involved in high-glucose-induced cellular
processes in human gastric smooth muscle cells. Biochem Biophys Res
Commun. 496:401–406. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yan B, Tao ZF, Li XM, Zhang H, Yao J and
Jiang Q: Aberrant expression of long noncoding RNAs in early
diabetic retinopathy. Invest Ophthalmol Vis Sci. 55:941–951. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang M, Gu H, Xu W and Zhou X:
Down-regulation of lncRNA MALAT1 reduces cardiomyocyte apoptosis
and improves left ventricular function in diabetic rats. Int J
Cardiol. 203:214–216. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hu M, Wang R, Li X, Fan M, Lin J, Zhen J,
Chen L and Lv Z: LncRNA MALAT1 is dysregulated in diabetic
nephropathy and involved in high glucose-induced podocyte injury
via its interplay with β-catenin. J Cell Mol Med. 21:2732–2747.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gong W, Zhu G, Li J and Yang X: LncRNA
MALAT1 promotes the apoptosis and oxidative stress of human lens
epithelial cells via p38MAPK pathway in diabetic cataract. Diabetes
Res Clin Pract. 144:314–321. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim EK and Choi EJ: Pathological roles of
MAPK signaling pathways in human diseases. Biochim Biophys Acta.
1802:396–405. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sui X, Kong N, Ye L, Han W, Zhou J, Zhang
Q, He C and Pan H: p38 and JNK MAPK pathways control the balance of
apoptosis and autophagy in response to chemotherapeutic agents.
Cancer Lett. 344:174–179. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cuadrado A and Nebreda AR: Mechanisms and
functions of p38 MAPK signalling. Biochem J. 429:403–417. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu JY, Yao J, Li XM, Song YC, Wang XQ, Li
YJ, Yan B and Jiang Q: Pathogenic role of lncRNA-MALAT1 in
endothelial cell dysfunction in diabetes mellitus. Cell Death Dis.
5:e15062014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Teraguchi M, Yoshimura N, Hashizume H,
Yamada H, Oka H, Minamide A, Nagata K, Ishimoto Y, Kagotani R,
Kawaguchi H, et al: Progression, incidence, and risk factors for
intervertebral disc degeneration in a longitudinal population-based
cohort: The Wakayama Spine Study. Osteoarthritis Cartilage.
25:1122–1131. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jiang L, Zhang X, Zheng X, Ru A, Ni X, Wu
Y, Tian N, Huang Y, Xue E, Wang X, et al: Apoptosis, senescence,
and autophagy in rat nucleus pulposus cells: Implications for
diabetic intervertebral disc degeneration. J Orthop Res.
31:692–702. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shan L, Yang D, Zhu D, Feng F and Li X:
High glucose promotes annulus fibrosus cell apoptosis through
activating the JNK and p38 MAPK pathways. Biosci Rep.
39:BSR201908532019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wachtel TJ: The diabetic hyperosmolar
state. Clin Geriatr Med. 6:797–806. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kitabchi AE and Nyenwe EA: Hyperglycemic
crises in diabetes mellitus: Diabetic ketoacidosis and
hyperglycemic hyperosmolar state. Endocrinol Metab Clin North Am.
35725–751. (viii)2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Umpierrez G and Korytkowski M: Diabetic
emergencies - ketoacidosis, hyperglycaemic hyperosmolar state and
hypoglycaemia. Nat Rev Endocrinol. 12:222–232. 2016. View Article : Google Scholar : PubMed/NCBI
|