Introduction
Ischemia, hypoxia, reperfusion injury and other
factors caused by apoplexy may result in brain injury (1). At present, cell therapy is considered
a promising method for the treatment of cerebral injury, and
numerous preclinical studies have made notable progress. For
cytotherapy, stem cells are particularly valued for their strong
proliferation and differentiation characteristics. Several studies
have investigated cytotherapy for the treatment of cerebral injury,
including strategies using bone marrow-derived mesenchymal stem
cells (BMSCs), neural stem cells (NSCs), neural precursor cells
(NPCs), embryonic stem cells and endothelial progenitor cells
(2–7). Stem cell therapy is an emerging
therapeutic modality in the treatment of stroke. The basis stems
from the observation that certain parts of the adult brain are
capable of regeneration (8). A
recent meta-analysis of preclinical studies demonstrated that
mesenchymal stem cells used to treat ischemic stroke were
associated with improvements in neurological function (9); however, important questions remain
unanswered, and translation to the clinical remains a distant
prospect.
The ability of self-renewal and differentiation of
MSCs into neural cells in vitro, as demonstrated by the
expression of neuronal markers such as neuronal nuclear antigen,
and non-ethical and tissue rejection-associated concerns make their
use a promising therapeutic approach for stroke treatment (10). BMSCs can protect neurons from
hypoxia (11). The experimental
rationale for the use of MSCs in stroke therapy involves a number
of divergent mechanisms of action, including differentiation into
cell types relevant to repair, modulation of the immune system,
promotion of angiogenesis, neurogenesis and synapse formation,
paracrine secretion of neuroprotective and neurotrophic factors
(12,13). The interaction between BMSCs and
other cells in the brain-injured area to further promote damage
repair requires further investigation (10).
NSCs/NPCs are present in the subventricular zone and
hippocampal dentate gyrus of mammals (14–16).
NSCs have the ability to self-renew and differentiate into neurons,
astrocytes or microglia with appropriate stimulation (17,18).
Following brain injury, NSCs/NPCs can be activated; they
proliferate and differentiate into neurons under pathological
conditions (19). The migration of
NSCs/NPCs to the focal lesion is key to promoting injury-associated
repair. The effective migration of exogenous or endogenous NSCs can
promote functional reconstruction of the nervous system (15). Investigations have been performed;
however, there is still a lack of clear and effective methods to
promote the directional migration of NSCs/NPCs. The number of NPCs
that migrate to the lesion is very small; thus, it is also
necessary to determine an efficient method to promote NPC migration
and appropriate targeting of NPCs.
MicroRNAs (miRNAs/miRs) are 20–24 nucleotide RNAs
that regulate the translation or expression levels of target mRNA
transcripts (20). Previous
studies suggested that miR-210 specifically is a robust target of
hypoxia-inducible factors, and miR-210 has complex roles in the
cellular responses to hypoxia (21–23).
Previous studies reported that miR-210 is closely associated with
ischemic stroke; miR-210 is not only upregulated in endothelial
cells under hypoxic conditions, but also promotes angiogenesis
(24) and is closely associated
with the physiological and pathological status of the nervous
system (25,26). Hypoxic preconditioning can increase
the expression of miR-210 in BMSCs. Ischemic preconditioning
augments the survival of stem cells via miR-210 expression by
targeting caspase-8-associated protein 2. Liu et al
(18) also confirmed that miR-210
expression in NSCs/NPCs is increased significantly under hypoxic
conditions. Our previous studies revealed that the transplanted
BMSCs localized to the ischemic foci following cerebral ischemia
(27,28). The penumbra surrounding the
ischemic foci was hypoxic following cerebral ischemia (28). Additionally, miR-210 expression was
demonstrated to be significantly increased in ischemic brain tissue
(24,29). Therefore, it was hypothesized that
the expression levels of miR-210 in BMSCs around the ischemic foci
following cerebral ischemia increased in the hypoxic environment,
which may promote the migration of NSCs to the surrounding ischemic
foci. This hypothesis was investigated in the present study by
detecting the expression of miR-210 in BMSCs under hypoxic
conditions and the effect of BMSCs on NPC migration. In addition,
the present study also investigated the effect of miR-210
overexpression and inhibition in BMSCs on the migration of
NPCs.
Materials and methods
Isolation and cultivation of mouse
BMSCs
Initially, ten 4- to 6-week-old C57BL/6 male mice
(weight 12–18 g, Hunan SJA Laboratory Animal Co., Ltd., Changsha,
China; http://www.hnsja.com/) were housed in
specific pathogen free (SPF) conditions (air cleanliness: Level 7,
temperature: 22±2°C) with free access to sterilized food and ultra
pure water. The mice were sacrificed by cervical dislocation. The
femur was obtained under sterile conditions. The ends of the femur
were removed, and PBS was used to flush out the bone marrow.
Following filtering with a 200-µm mesh filter, the bone marrow cell
suspension was centrifuged at 200 × g for 5 min at 4°C. The cell
pellet was resuspended with BMSC culture media [Dulbecco's modified
Eagle's medium (DMEM)/F12+10% fetal bovine serum (FBS; Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA)]. After 24 h in
culture at 37°C in 5% CO2, the media was replaced to
remove the non-adherent cells, and then the media was changed every
3 days. When the cells reached confluence, the cells were passaged.
The cells from passage 4 were evaluated by flow cytometry analysis.
The identification indexes were cluster of differentiation (CD)34,
CD44 and CD71. A total of 1×105−106 cells
were suspended in 100 µl PBS. Then 2 µl CD34 antibody, eFluor660
(cat. no. 50-0341-82; ebioscience; Thermo Fisher Scientific, Inc.),
CD44 antibody, PE (12-0441-82; Thermo Fisher Scientific, Inc.) or
CD71 antibody, PE (12-0711-82; Thermo Fisher Scientific, Inc.) was
added into the BMSC suspension and cultured at 4°C for 45 min, and
then washed with PBS twice. After that the BMSC suspension was
examine by the flow cytometer (Novocyte 2040R,
NovoExpress® 1.2.5 software, ACEA Bioscience Inc., San
Diego, CA, USA). The BMSCs from passages 4 and 5 were used for
further experiments. All procedures and experiments in the present
study were conducted under guidelines and were approved by the
Laboratory Animal Welfare and Ethics Committee of the Third
Military Medical University (Chongqing, China).
Isolation and cultivation of mouse
NPCs
A total of 10 C57BL/6 pregnant mice at day 13 of
gestation (weight 34–35 g; Hunan SJA Laboratory Animal Co., Ltd.)
were housed in SPF conditions (air cleanliness: Level 7,
temperature: 22±2°C) with free access to sterilized food and ultra
pure water. These mice were sacrificed by cervical dislocation.
Under sterile conditions, the abdominal skin, muscle and uterus
were cut so as to obtain mouse embryos. The telencephalon was
isolated, the meninges were removed and the tissue was cut into
pieces under a stereomicroscope. The tissue was digested with 0.05%
trypsin-EDTA at 37°C and agitated every 5 min until a homogenized
solution was obtained. Then, the complete media containing serum
(DMEM/F12+10% FBS) was added to stop the digestion. After a 200 × g
centrifugation for 5 min at 4°C, the cell pellet was suspended with
NPC culture media (DMEM/F12, 1% N-2, 2% B-27, 10 ng/ml
basic-fibroblast growth factor, 20 ng/ml epidermal growth factor)
for cultivation at 37°C in 5% CO2. The NPCs developed
into a bulb shape. The media was changed every 2 days. The NPCs
were identified by the immunofluorescent staining of β-tubulin III,
doublecortin (DCX) and nestin. The NPCs were fixed with 4%
paraformaldehyde at room temperature for 30 min, washed with PBS
twice and treated with 0.2% Triton X-100 for 30 min. Following
washing with PBS twice, the cells were incubated with β-tubulin III
antibody (1:100; cat. no. sc-166729; Santa Cruz Biotechnology,
Inc., Dallas, TX, USA), anti-DCX antibody (1:100; cat. no.
sc-28939; Santa Cruz Biotechnology, Inc.) or anti-nestin antibody
(1:100; cat. no. sc-21248; Santa Cruz Biotechnology, Inc.) at 4°C
over night. After washing with PBS twice, the cells were incubated
with fluorescein isothiocyanate (FITC)-labeled goat-anti-mouse
antibody (1:100; cat. no. ab6785; Abcam, Cambridge, USA),
TRITC-labeled goat-anti-rabbit antibody (1:100; cat. no. ab6718;
Abcam), FITC-labeled donkey-anti-goat antibody (1:100; cat. no.
ab6881; Abcam) at room temperature for 4 h. Then the cells were
washed with PBS and observed under fluorescence microscope. The
NPCs from passage 3 were used for further experiments.
Hypoxic treatment of BMSCs
According to the method of Pulkkinen et al
(22), the BMSCs cultured under
hypoxia were then cultured in a low oxygen incubator chamber with a
94% N2, 1% O2, 5% CO2 low-oxygen
gas mixture for 24 h. The BMSCs that were cultured under normal
conditions (37°C, 5% CO2) were used as a control. Three
sample parallel experiments were performed in each group.
Cell transfection experiments
Previous methods were applied with minor revisions
(30,31). miR-210 mimics and inhibitors were
used to upregulate or downregulate the expression of miR-210,
respectively, in BMSCs. miR-210 mimics and inhibitors were
synthesized by Shanghai GenePharma Co., Ltd. (Shanghai, China). The
BMSCs were seeded into 6-well plates at a density of
5×105 cells/well. Then, the cells were divided into
three groups: Negative control (NC), overexpressing miR-210 and the
suppressing miR-210 groups. On day 2 in culture, the cells were
transfected with NC, miR-210 mimics or inhibitors according to the
manufacturer's protocols for Lipofectamine® 2000 (7.5
µl/well; Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). The doses were 75 pmol/well for the NC miRNA, the miR-210
mimics and miR-210 inhibitors. Following the addition of 100 µl
serum-free medium (DMEM/F12) to three Eppendorf tubes, and the NC
miRNA, miR-210 mimics or miR-210 inhibitors, together with the
Lipofectamine® 2000 reagent, the mixtures were stored at
room temperature for 20 min and then added to the corresponding
wells. The transfected cells were collected at 24 and 48 h
post-transfection. The expression of miR-210 was detected by
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR). The sequence of the double stranded miR-210 mimic was
(5′-3′) CUGUGCGUGUGACAGCGGCUGA and AGCCGCUGUCACACGCACAGUU; the
miR-210 inhibitor sequence was (5′-3′) UCAGCCGCUGUCACACGCACAG; and
the NC sequence was (5′-3′) UUCUCCGAACGUGUCACGUTT and
ACGUGACACGUUCGGAGAATT.
Cell migration experiments
The Transwell cell migration assay was performed to
detect the effect of BMSCs on NPC migration. The BMSCs were seeded
into the lower chamber at a density of 1×104 cells/well
in BMSC culture medium at 37°C. Following culturing for 24 or 48 h,
the NPCs were added to the upper chamber at a density of
5×103 cells/well in NPC culture media. After
co-culturing for 15 or 24 h, the migrated NPCs on the membrane were
stained with 0.1% crystal violet at room temperature for 10 min.
Three parallel samples in each group were analyzed. The cell number
was counted in 20 fields with a light microscope (magnification,
×200).
The effects of miR-210 expression within BMSCs on
NPC migration were investigated. The BMSCs overexpressing miR-210,
suppressing miR-210 expression and the NC-transfected BMSCs were
plated in the lower chamber at a density of 1×104
cells/well in BMSC culture media. Following culturing for 24 h, the
NPCs were added to the upper chamber at a density of
5×103 cells/well in NPC culture media. After 24 h of
co-culturing, the migrated NPCs on the membrane were fixed with 4%
paraformaldehyde at room temperature for 15 min, washed with PBS,
and stained with 0.1% crystal violet at room temperature for 10
min. A total of three parallel samples in each group were analyzed.
The cell number was counted in 20 fields of a light microscope
(magnification, ×200).
RNA extraction and RT-qPCR
Using RT-qPCR, miR-210 expression levels were
detected within BMSCs following hypoxic treatment and transfection
with miR-210 mimics or inhibitors. The mRNA expression levels of
vascular endothelial growth factor-C (VEGF-C), brain derived
neurotrophic factor (BDNF), and chemokine C-C motif ligand 3 (CCL3)
in BMSCs successfully transfected with miR-210 mimics, inhibitors
or NC, were also detected by RT-qPCR after pre-culturing for 24 h.
The BMSCs transfected with NC miRNA (NC BMSCs) served as controls.
Total RNA was extracted with TRIzol® (Thermo Fisher
Scientific, Inc.). The cDNA was obtained via RT reaction by
PrimeScript™ RT reagent kit with gDNA Eraser (Perfect
Real Time; Takara Biotechnology Co., Ltd., Dalian, China) according
to the manufacturer's protocol. Then, qPCR was performed according
to the protocols of SYBR® Premix Ex Taq™ II
(TliRNaseH Plus), Bulk kit (Takara Biotechnology Co., Ltd.). GAPDH
was used as the internal reference. The qPCR conditions were 1
cycle of denaturation at 95°C for 30 sec, then followed 40 cycles
of denaturation at 95°C for 5 sec and annealing at 60°C for 34 sec.
The primer sequences were as follows: miR-210 forward,
5′-GCAGTCTGTGCGTGTGACAGC-3′, reverse, 5′-GTGCAGGGTCCGAGGT-3′;
VEGF-C forward, 5′-ACTTGCTGTGCTTCTTGT-3′, reverse,
5′-CTCATCTACGCTGGACAC-3′; BDNF forward, 5′-CCAGGTGAGAAGAGTGATG-3′,
reverse 5′-AGTGATGTCGTCGTCAGA-3′ and CCL3 forward,
5′-CCTTGCTGTTCTTCTCTGT-3′ and reverse 5′-ATGAATTGGCGTGGAATCT-3′.
GAPDH forward, 5′-TGCACCACCAACTGCTTAGC-3′ and reverse
5′-GGCATGGACTGTGGTCATGAG-3′. The 2−ΔΔCq method was used
for quantification (32).
ELISA
The BMSCs were seeded into 6-well plates at a
density of 5×105 cells/well. The supernatants from
normal and hypoxic BMSCs were collected for the detection of
VEGF-C, BDNF and CCL3 expression. The expression levels of BDNF,
VEGF-C, and CCL3 were analyzed using BDNF, VEGF-C and CCL3 ELISA
kits, respectively (cat. nos. TY0362b, TY0258b and TY3766b;
Shanghai Lichen Trading Company, Shanghai, China) according to the
manufacturer's protocols. The experiments were performed in
triplicate.
Western blotting
The expression levels of VEGF-C, BDNF, and CCL3 in
the BMSCs successfully transfected with miR-210 mimics, inhibitors
or NC, were analyzed by western blotting following pre-culturing
for 24 h. The NC BMSCs served as the control. The cells were lysed
with radioimmunoprecipitation assay lysis buffer (Beyotime
Institute of Biotechnology, Haimen. China). The samples were
centrifuged at 12,092 × g for 10 min at 4°C. The supernatant was
collected, and the protein concentration was determined with a
bicinchoninic protein assay kit (Thermo Fisher Scientific, Inc.).
Then, 30 µg total protein was loaded onto a gel and then separated
by electrophoresis. After transferring to nitrocellulose membranes,
the membrane was blocked with 5% skimmed milk at room temperature
for 1 h and then incubated with primary antibody against BDNF
(1:500; Abcam; cat. no. ab108319), VEGF-C (1:500; Abcam; cat. no.
ab9546), or CCL3 (1:500; Abcam; cat. no. ab179638) overnight at
4°C. β-actin was used as control (1:200; Santa-Cruz Biotechnology,
Inc.; cat. no. SC69879). Following three washes with 1X
Tris-buffered saline with Tween-20, the membranes were incubated
with horseradish peroxidase-conjugated goat anti-rabbit IgG
(1:2,500, diluted in PBS; cat. no. ZB2301; OriGene Technologies,
Inc., Beijing, China) or peroxidase-conjugated goat anti-mouse IgG
(1:2,500, diluted in PBS; cat. no. ZB2305; OriGene Technologies,
Inc.) for 2 h at room temperature. The signal was detected with an
enhanced chemiluminescence substrate Pierce fast western blotting
kit (Thermo Fisher Scientific, Inc.), and the densitometric values
were analyzed with the ImageJ 1.43 program (National Institutes of
Health, Bethesda, MD, USA). All experiments were repeated at least
three times.
Statistical analysis
All values are expressed as the mean ± standard
deviation. The data were analyzed using SPSS 18.0 software (SPSS,
Inc., Chicago, IL, USA). A t test was used to analyze the
difference between two groups. One-way analysis of variance was
used to analyze the difference among three or more groups. When
homogeneity of variance was satisfied, a Tukey's post-hoc test was
used for multiple comparisons. When homogeneity of variance was not
satisfied, a Dunnett's T3 test was used for multiple comparisons.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Isolation and identification of
BMSCs
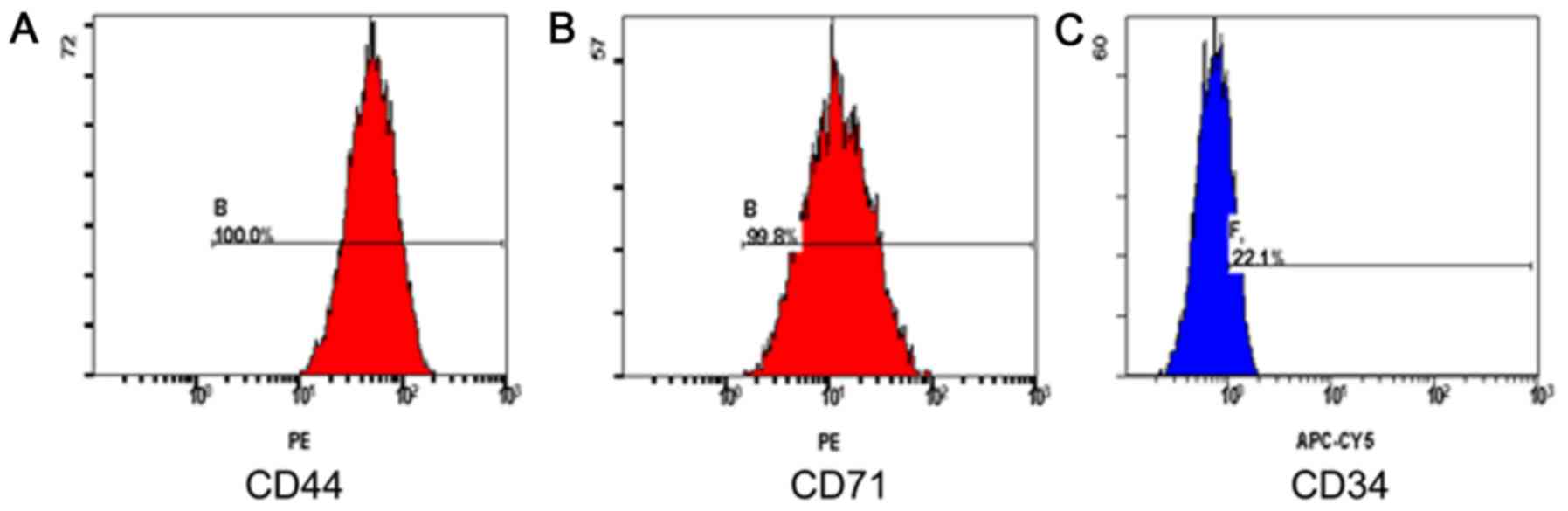
In the present study, the antigenic phenotype of
isolated and cultured BMSCs was detected by flow cytometric
analysis with anti-CD44, anti-CD71 and anti-CD34 antibodies. The
BMSCs from passage 4 were spindle shaped. As presented in Fig. 1, following the analysis and
identification of surface markers by flow cytometry, the positive
rate of CD44 was 100% (Fig. 1A),
the positive rate of CD71 was 99.8% (Fig. 1B), and the positive rate of CD34
was 22.1% (Fig. 1C). Thus, BMSCs
had been successfully isolated.
Identification, isolation and
cultivation of NPCs
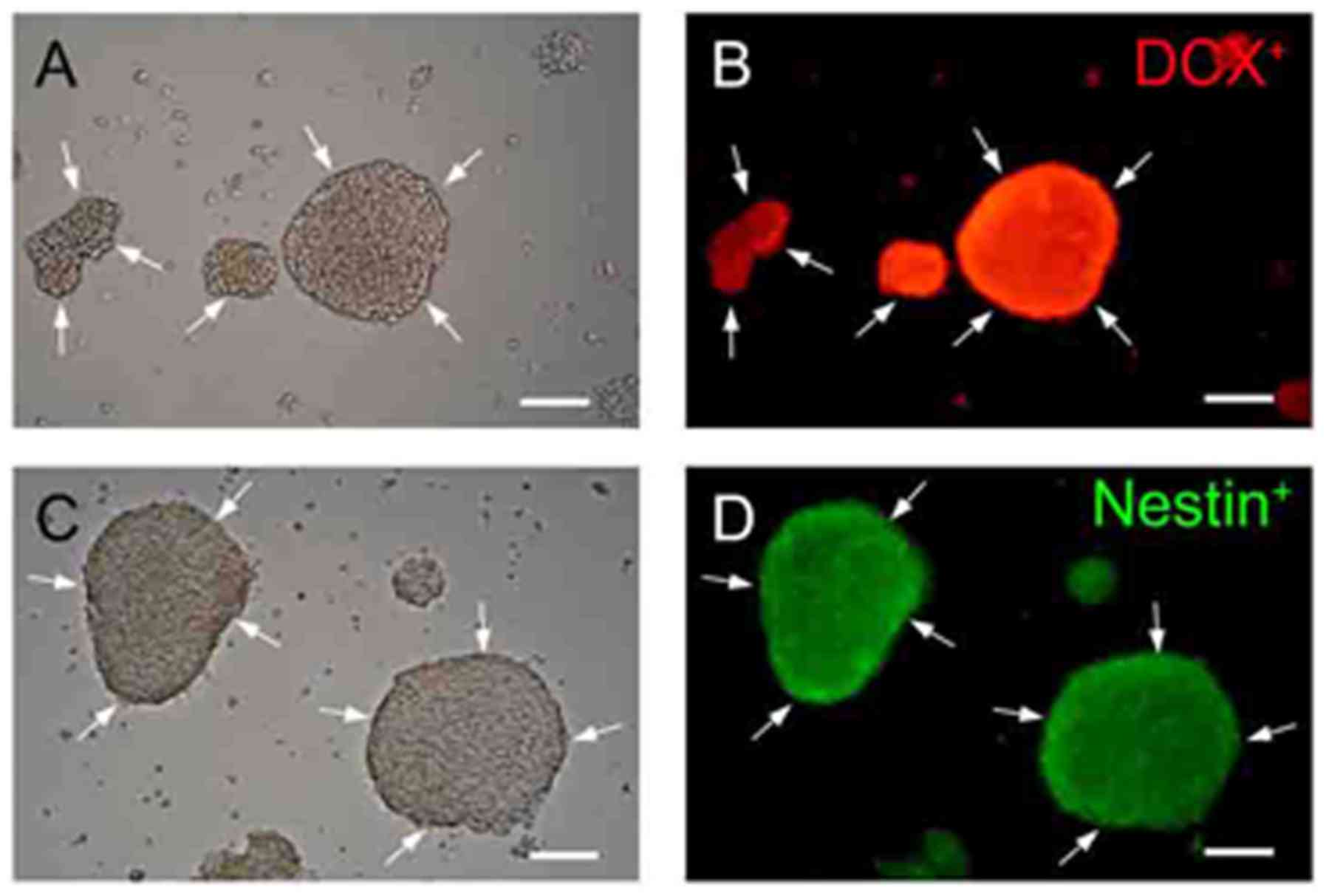
The NPCs exhibited sphere-like morphologies. DCX is
a microtubule-associated protein expressed by NPCs. DCX is
expressed within NPCs during active division, and their neuronal
daughter cells continue to express DCX for 2–3 weeks as the cells
mature into neurons (33). The
protein nestin is widely used as a marker of NSCs and NPCs, and
β-tubulin III is a general marker of neurons. The identification,
isolation and cultivation of NPCs were confirmed by
immunofluorescent staining with anti-nestin, anti-DCX and
anti-β-tubulin III antibodies. As presented in Fig. 2, nestin and DCX staining was
positive; however, β-tubulin III staining was negative (data not
shown). Thus, the isolation and cultivation of NPCs were
successful.
Co-cultured BMSCs and NPCs
significantly promote the migration of NPCs
In the present study, NPCs and BMSCs were
successfully cultured. Subsequently, the effects of BMSCs on NPC
migration were investigated using a Transwell cell migration assay.
The present study examined four groups: Pre-cultured BMSCs for 24 h
and co-cultured BMSCs/NPCs for 15 h; pre-cultured BMSCs for 24 h
and co-cultured BMSCs/NPCs for 24 h; pre-cultured BMSCs for 48 h
and co-cultured BMSCs/NPCs for 15 h, and pre-cultured BMSCs for 48
h and co-cultured BMSCs/NPCs for 24 h. The number of migrated NPCs
was greatest in the group of BMSCs pre-cultured for 24 h and
co-cultured NPCs for 24 h, which was significantly higher than any
other group [pre-cultured BMSCs for 24 h and co-cultured BMSCs/NPCs
for 24 h vs. pre-cultured BMSCs for 24 h and co-cultured BMSCs/NPCs
for 15 h (P=0.045); pre-cultured BMSCs for 48 h and co-cultured
BMSCs/NPCs for 15 h (P=0.041); and pre-cultured BMSCs for 48 h and
co-cultured BMSCs/NPCs for 24 h (P<0.001); Fig. 3]. The number of migrated NPCs in
the group of pre-cultured BMSCs for 48 h and co-cultured BMSCs/NPCs
for 24 h was significantly lower than that in the pre-cultured
BMSCs for 24 h, co-cultured BMSCs/NPCs for 24 h, pre-cultured BMSCs
for 48 h and co-cultured BMSCs/NPCs for 15 h (P<0.001; Fig. 3). Thus, the BMSCs pre-cultured for
24 h and BMSCs/NPCs co-cultured 24 h were the optimal combinations,
which improved the migration ability of NPCs significantly.
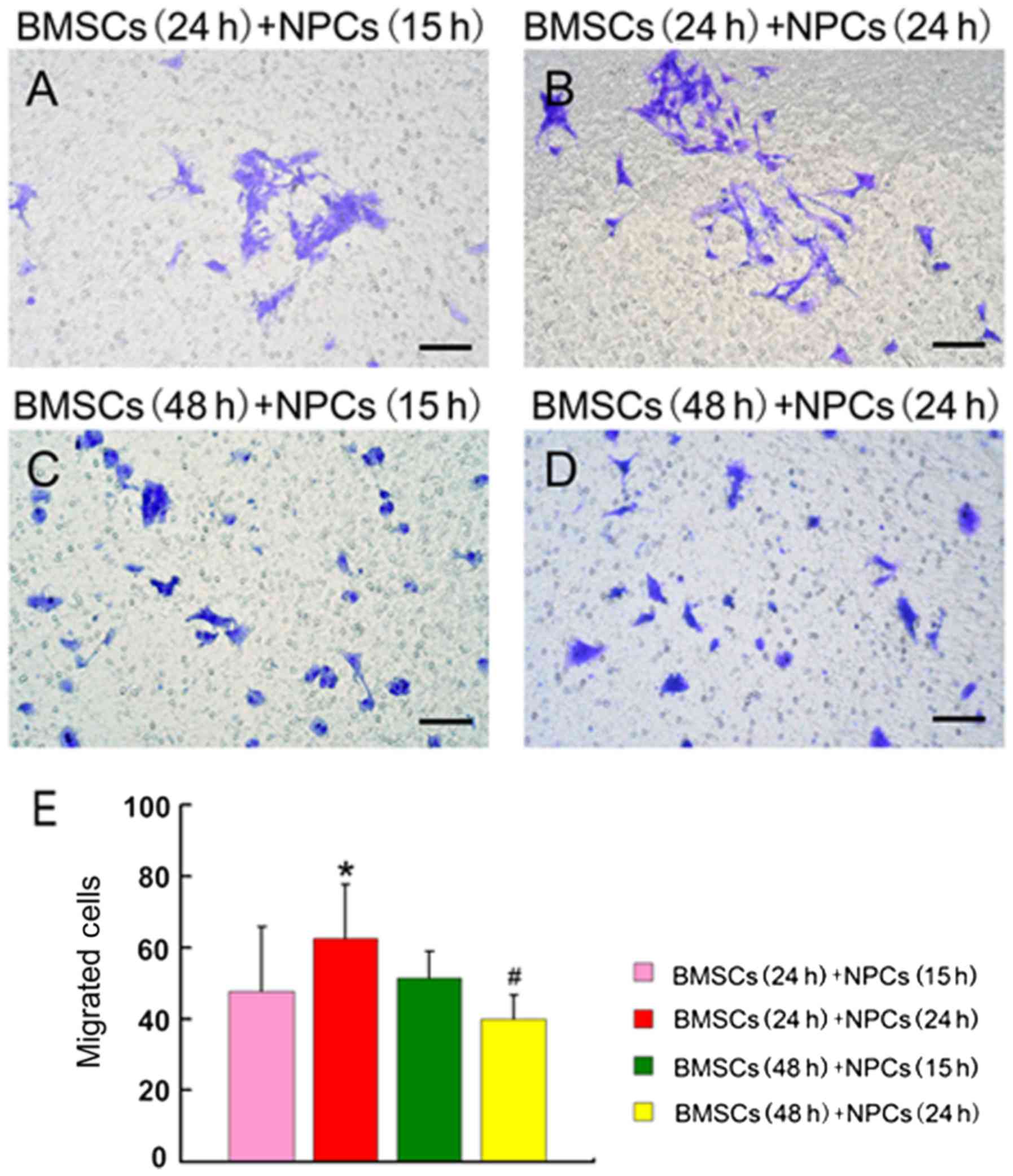 | Figure 3.Effects of BMSCs on the migration of
NPCs. (A) BMSCs pre-cultured for 24 h, BMSCs/NPCs co-cultured for
15 h; (B) BMSCs pre-cultured for 24 h, BMSCs/NPCs co-cultured for
24 h; (C) BMSCs pre-cultured for 48 h, BMSCs/NPCs co-cultured for
15 h; (D) BMSCs pre-cultured for 48 h, BMSCs/NPCs co-cultured for
24 h. (E) Quantification of migrated cells. Number of fields
counted (n=20), *P<0.05 vs. BMSCs (24 h) + NPCs (15 h), BMSCs
(48 h) + NPCs (15 h) and BMSCs (48 h) + NPCs (24 h).
#P<0.05, vs. BMSCs (24 h) + NPCs (24 h) or BMSCs (48
h) + NPCs (15 h). Scale bar, 100 µm. BMSCs, bone marrow-derived
mesenchymal stem cells; NPC, neural precursor cells. |
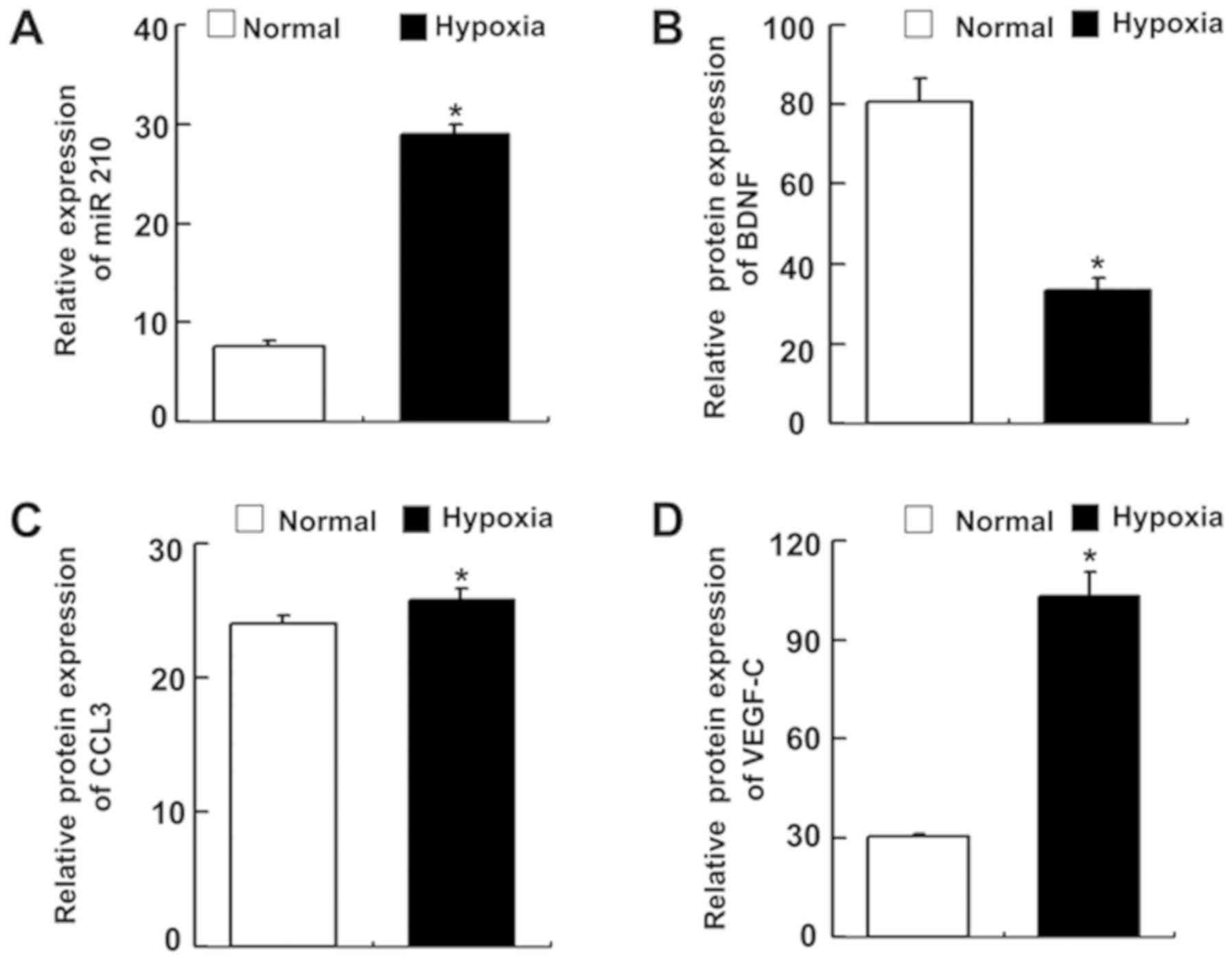
miR-210 expression is significantly
upregulated under hypoxic conditions
As aforementioned, miR-210 is significantly and
consistently upregulated under hypoxic conditions. miR-210
expression levels were significantly upregulated in BMSCs under
hypoxic conditions (Fig. 4A;
P=0.014). The expression levels of BDNF, VEGF-C and CCL3 in the
supernatant of BMSCs were quantified by ELISA. Compared with the
BMSCs cultured under normal conditions, during hypoxic conditions,
BDNF expression levels were significantly decreased (Fig. 4B; P=0.001), CCL3 and VEGF-C
expression levels increased significantly (P=0.049 and P=0.003,
respectively; Fig. 4C and D,
respectively).
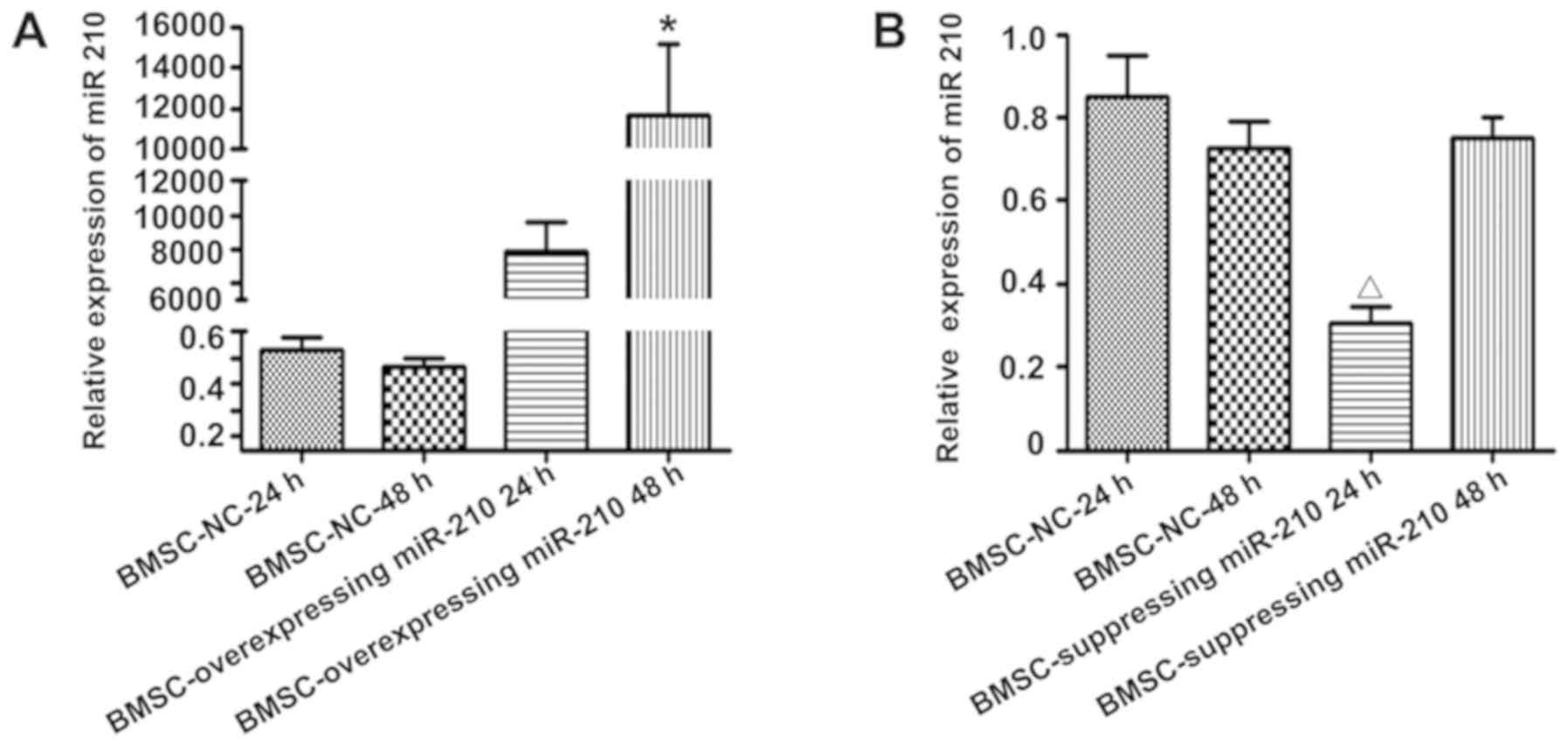
Effects of miR-210 on the migration of
NPCs on the co-culture system
The present study successfully isolated and cultured
BMSCs and NPCs. As miR-210 expression levels were significantly
upregulated in BMSCs under hypoxia, the effects of miR-210
overexpression and inhibition in BMSCs on the migration of NPCs
were investigated in the present study. miR-210 mimics or
inhibitors were transfected into BMSCs to upregulate or
downregulate miR-210 expression, respectively. The miR-210
expression levels in BMSCs were increased following transfection
with miR-210 mimics. The miR-210 expression levels were highest in
BMSCs transfected with miR-210 mimics for 48 h, which was
significantly higher than that in the BMSCs transfected with NC
miRNA for 24 or 48 h, and within BMSCs transfected with miR-210
mimics for 24 h [miR-210 mimics 48 h vs. NC miRNA 24 h (P=0.001),
NC miRNA 48 h (P=0.001) and miR-210 mimics 24 h, (P=0.002);
Fig. 5A].
The miR-210 expression levels in BMSCs decreased
following transfection with miR-210 inhibitors. The miR-210
expression levels in BMSCs was lowest at 24 h post-transfection
with a miR-210 inhibitor, which was significantly lower than BMSCs
transfected with NC miRNA for 24 or 48 h and in BMSCs transfected
with miR-210 inhibitor for 48 h [miR-210 inhibitor 24 h vs. NC
miRNA 24 h (P=0.005), NC miRNA 48 h (P=0.001) and miR-210 inhibitor
48 h (P=0.042); Fig. 5B]. Thus,
the BMSCs transfected with miR-210 mimic for 48 h and BMSCs
transfected with miR-210 inhibitor for 24 h for further experiments
to investigate the effects of miR-210 expression in BMSCs on NPC
migration.
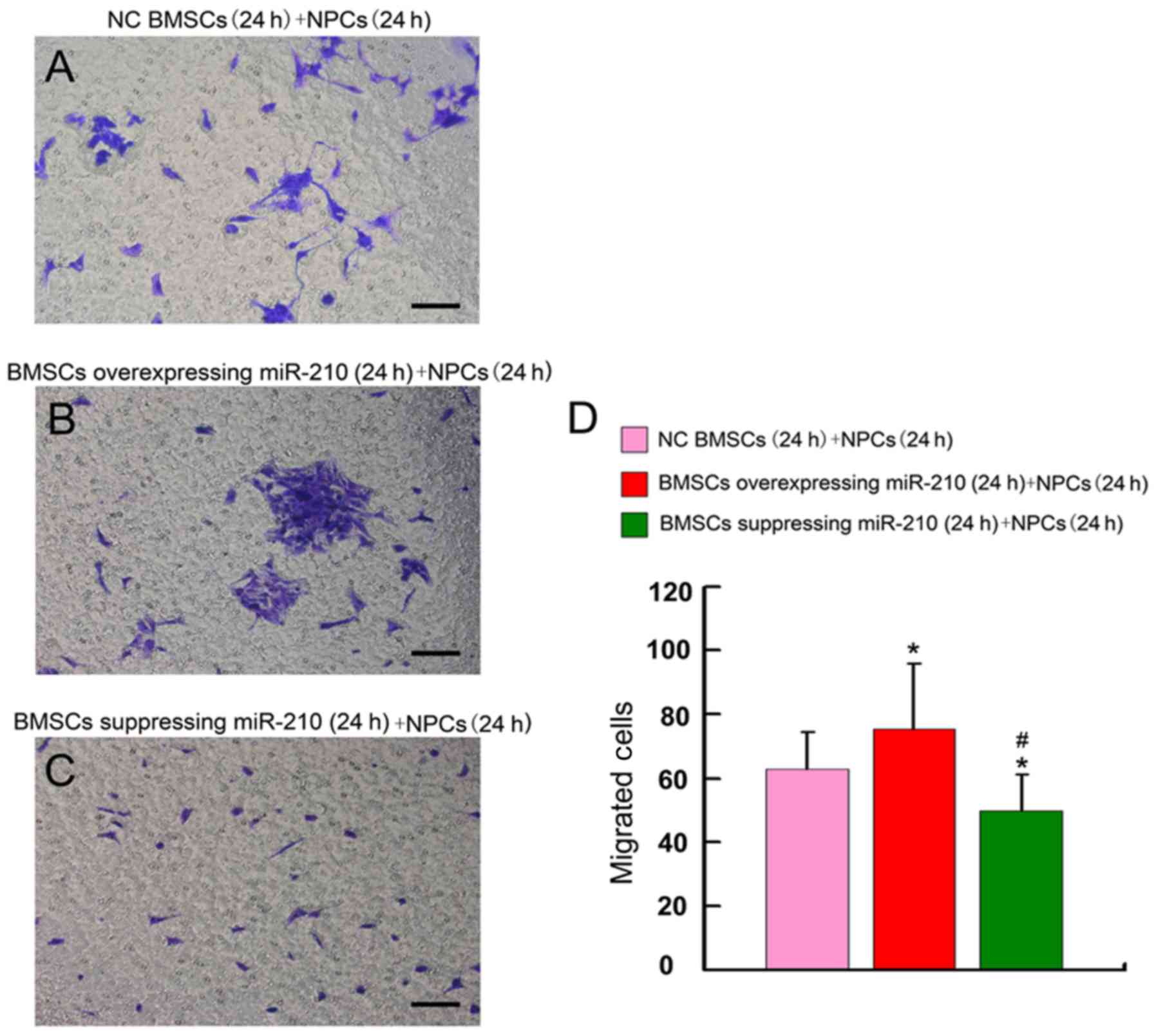
Subsequently, the effect of miR-210 expression in
BMSCs on NPC migration was investigated in the present study. The
number of migrated NPCs was significantly higher in the miR-210
overexpressing BMSC group than in the NC BMSCs group (P=0.028;
Fig. 6). Conversely, the number of
migrated NPCs in the miR-210-suppressed group (BMSCs suppressing
miR-210 expression pre-cultured for 24 h and BMSCs suppressing
miR-210 expression/NPCs co-cultured for 24 h) was significantly
decreased compared with that in the NC BMSCs group (Fig. 6; P=0.023). Compared with in the NC
BMSCs with co-cultured NPCs for 24 h group, the BMSCs
overexpressing miR-210 significantly promoted the migration of
NPCs. Reducing the expression of miR-210 in BMSCs resulted in fewer
migrated NPCs. The results of the present study suggested that
miR-210 had an important role in the migration of NPCs in this
co-culture system.
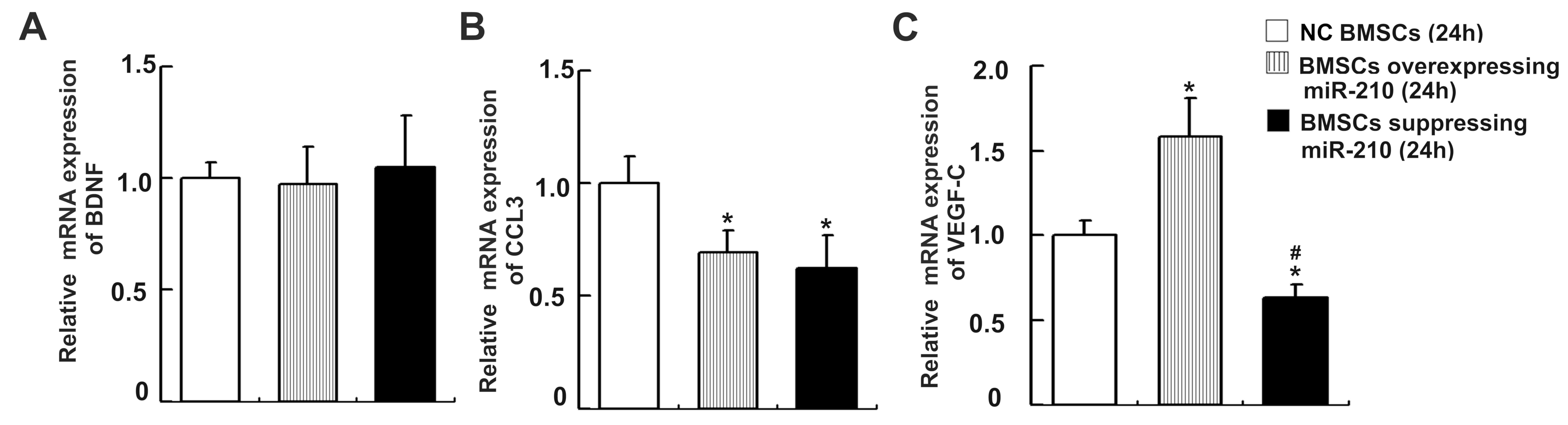
Effect of miR-210 on the expression
levels of BDNF, CCL3 and VEGF-C in BMSC
As aforementioned, under hypoxic conditions, miR-210
was upregulated; BDNF, VEGF-C, and CCL3 expression levels were also
upregulated or downregulated by hypoxia. It was hypothesized that
miR-210 may regulate BDNF, VEGF-C and CCL3 gene expression; the
effects of miR-210 overexpression or inhibition on the expression
of BDNF, CCL3 and VEGF-C in BMSCs were investigated. From RT-qPCR
analyses, BDNF mRNA expression levels in the BMSCs overexpressing
miR-210 or BMSCs suppressing miR-210 expression were not
significantly different from the NC BMSCs group (Fig. 7A). The CCL3 mRNA expression levels
were significantly downregulated in BMSCs when miR-210 was
overexpressed (Fig. 7B; P=0.029)
or suppressed (Fig. 7B; P=0.014).
The VEGF-C mRNA expression levels were significantly increased
(Fig. 7C; P=0.005) when miR-210
was overexpressed in BMSCs and significantly decreased when miR-210
was reduced (Fig. 7C; P=0.037).
The present study determined the effects of miR-210 on the protein
expression levels of BDNF, CCL3 and VEGF-C in BMSCs. Consistent
with the mRNA expression levels, the western blot analyses
indicated that BDNF expression levels were not significantly
different when miR-210 was overexpressed or suppressed in BMSCs
compared with in the NC BMSCs group (Fig. 8A). The expression levels of CCL3
did not significantly change when miR-210 was overexpressed
(P=0.171), while the expression of CCL3 significantly decreased
when miR-210 expression was inhibited (P=0.014) in BMSCs (Fig. 8B). The expression levels of VEGF-C
were significantly increased when miR-210 was overexpressed
(P=0.002) and significantly decreased when miR-210 expression was
inhibited (P=0.039) in BMSCs (Fig.
8C).
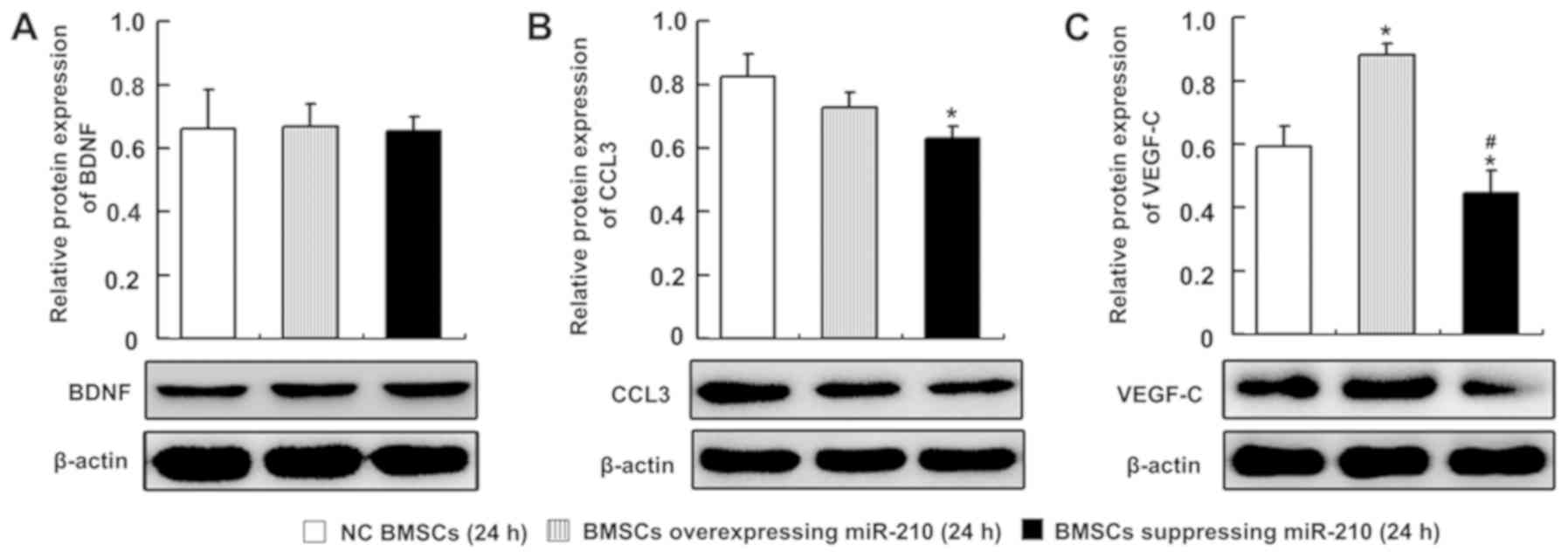 | Figure 8.Western blot analysis of BDNF, CCL3
and VEGF-C expression in the miR-210-regulated BMSCs pre-cultured
for 24 h. Representative bands and densitometry quantification of
(A) BDNF, (B) CCL3 and (C) VEGF-C protein expression in the NC
BMSCs group, BMSCs overexpressing miR-210 group and BMSCs
suppressing miR-210 group. n=3 for each group, *P<0.05 vs. NC
BMSCs group; #P<0.05, vs. BMSCs overexpressing
miR-210 group. BDNF, brain derived neurotrophic factor; NC,
negative control; BMSCs, bone marrow-derived mesenchymal stem
cells; CCL3, chemokine C-C motif ligand 3; VEGF-C, vascular
endothelial growth factor-C. |
Discussion
The migration, proliferation and differentiation of
NPCs located in the subventricular zone and dentate gyrus are key
to promoting self-repair following brain damage; however, the
number of NPCs that migrate to the lesion is limited and
insufficient to meet the requirements of functional reconstruction
(34). Therefore, elucidating the
factors that influence and promote NPC migration is required to
promote self-repair following brain injury.
Cell therapy is considered a prospective treatment
for brain injury with good therapeutic effects. Autologous BMSCs
have the advantages of being obtained easily, proliferative in
vitro, and immune privileged (35). BMSCs are an ideal candidate for
cell therapy. At present, research on BMSCs for the treatment of
brain injury has focused on inducing BMSCs to differentiate into
neurons to replace damaged neurons (36), paracrine-secretion of certain
neurotrophic factors, such as VEGF, BDNF, nerve growth factor,
basic fibroblast growth factor, to support the survival of neural
cells (12,13), promoting neural stem/progenitor
cell proliferation, and regulating the differentiation of neural
stem/progenitor cells to neurons (34,37).
Further investigation into the effects of BMSCs on promoting the
migration of neural stem/progenitor cells via transplantation are
required.
Stem cell transplantation has been proposed as a
means of cell replacement therapy. Once locally or systematically
injected, about 1/3 of the locally injected stem cells migrated
toward the damaged region (38).
Following ischemic stroke, NPCs proliferate within major germinal
niches of the brain (39). In
recent years, numerous studies have reported an interaction between
BMSCs and NSCs during brain injury repair. Haragopal et al
(40) revealed that the direct
effects of BMSCs on NSCs may enhance the stemness of NSCs via the
Notch-1 signaling pathway, which may be useful for establishing
human neural stem cell lines in vitro for basic research,
clinical research and clinical transplantation. Wang et al
(41) reported that BMSCs
regulated the proliferation and differentiation of NSCs via the
Notch signaling pathway. Similarly, NSCs may affect BMSCs;
Alexanian (36) suggested that
NSCs may induce BMSCs to differentiate into neural stem-like cells;
however, it is unknown whether BMSCs regulate the migration of
NSCs/NPCs. In the present study, BMSCs were employed to induce NPC
migration, which may resolve the key problem of NSC/NPC migration
following brain injury. In the present study, the effects of BMSCs
on NPC migration were investigated via a Transwell cell migration
assay. The results of the present study revealed that pre-culturing
BMSCs for 24 h and co-culturing BMSCs and NPCs for 24 h may
significantly improve NPC migration compared with pre-culturing
BMSCs for 48 h. These findings suggested that 24 h may be an
optimal duration for NPC migration; the BMSCs exhibited a stronger
ability to promote NPC migration with in relatively a shorter
duration of pre-culture of 24 h than 48 h. These observations may
be associated with the decreased cell growth and proliferation due
to contact-dependent inhibition when BMSCs are pre-cultured for 48
h (42–44).
BMSCs may be located in a hypoxic environment if
BMSCs are stereotactically injected into the focal ischemic area or
if BMSCs migrate to the infarct border zone via intravenous
injection post-ischemic brain damage. miR-210 is a
hypoxia-responsive miRNA that can combine with hypoxia inducible
factor 1α (HIF-1α). miR-210 expression levels were increased in
cells or tissues significantly under hypoxic conditions (18,45,46).
The results of the present study revealed that miR-210 expression
in hypoxic BMSCs increased significantly, which was consistent with
the results of previous studies (18,45,46).
miR-210 serves an important role in promoting cell
survival under hypoxia. Nie et al (47) confirmed that the overexpression of
miR-210 significantly promoted the survival of BMSCs under hypoxic
conditions. Sun et al (48)
also reported that miR-210 may form a positive feedback loop with
HIF-1α to promote the survival of osteoblast cells post-hypoxia.
miR-210 also exhibited protective effects on NSCs/NPCs; Wang et
al (49) reported that miR-210
inhibited the apoptosis of NPCs by inhibiting BNIP3. In addition to
the protective effects on cell survival during hypoxia, miR-210 may
also exhibit an effect on cell migration. Zhang et al
(50) confirmed that phosphatase
of regenerating liver-3 can promote the migration and invasion of
gastric cancer cells by the nuclear factor-κB/HIF-1α/miR-210 axis.
Similarly, Qu et al (51)
also reported that the overexpression of miR-210 may promote the
migration of colorectal cancer cells. In the present study, the
upregulation of miR-210 in BMSCs may have enhanced the effect of
BMSCs on NPC migration, while the downregulation of miR-210 in
BMSCs may have reduced the effect of BMSCs on NPC migration. Thus,
miR-210 may serve an important role in the process of BMSC-induced
regulation of NPC migration.
In the present study, miR-210 expression levels were
increased in BMSCs under hypoxic conditions. VEGF-C and CCL3
expression levels were significantly increased in the supernatant
of BMSCs cultured under hypoxic conditions compared with that of
BMSCs cultured under normal conditions; while BDNF expression
decreased significantly. To investigate whether the alterations in
BDNF, VEGF-C and CCL3 expression may be regulated by miR-210, the
expression levels of BDNF, VEGF-C and CCL3 in BMSCs overexpressing
miR-210 or suppressing miR-210 expression were analyzed. VEGF-C
mRNA and protein expression levels were increased significantly;
however, the expression levels of CCL3 and BDNF protein did not
change significantly in BMSCs overexpressing miR-210. Additionally,
VEGF-C expression levels were significantly decreased, CCL3
expression decreased significant and BDNF expression did not change
significantly in BMSCs with reduced miR-210 expression levels. The
association between CCL3 or BDNF and miR-210 was unclear; however,
VEGF-C expression increased when miR-210 expression was upregulated
and vice versa. Combined with the results from previous
studies, miR-210 may be associated with the increased level of VEGF
expression (52–54). A high concentration of VEGF in
glioma C6 cells promoted the transmigration of human NSCs (ReNcells
CX cell lines) (55). Therefore,
miR-210 in BMSCs may further promote NPC migration by increasing
the expression of VEGF.
In summary, the present study successfully isolated
BMSCs and NPCs and examined the effects of BMSCs on NPC migration
via a Transwell cell migration assay. BMSCs may promote NPC
migration; pre-culturing BMSCs for 24 h and co-culturing BMSCs/NPCs
for 24 h were the optimal durations of incubation for NPC
migration. The BMSCs overexpressing miR-210 may have promoted NPC
migration further by increasing VEGF-C expression. These data may
provide insight into the mechanism, in which transplanted BMSCs may
promote NPC migration to the ischemic focus following cerebral
ischemia in vivo by increasing miR-210 and VEGF expression;
however, further investigation in vivo in the future is
required. In addition, the mechanism underlying miR-210-mediated
regulation of VEGF remains to be elucidated in future studies. The
results of the present study may assist in developing new
therapeutic regimens for cerebral ischemia. Transplantation of
BMSCs, especially the BMSCs overexpressing miR-210 after cerebral
ischemia may promote NPC migration to the ischemia foci, which is
good for brain damage repair.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81100927
and 81671150) and the Chongqing Science and Technology Commission
(grant no. cstc2011jjA10079).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
FW, acquisition of data, and drafting the
manuscript. ZZ, design of the study, interpretation of the data,
drafting of the manuscript and gave final approval of the version
to be published; JeZ, acquisition and analysis of data. JZ,
interpretation of data, revising the manuscript critically for
important intellectual content; WD, acquisition and analysis of
data.
Ethics approval and consent to
participate
All procedures and experiments in the present study
were conducted under ARRIVE guidelines (http://www.nc3rs.org.uk/arrive-guidelines) and were
approved by the Laboratory Animal Welfare and Ethics Committee of
the Third Military Medical University (Chongqing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Khatri R, McKinney AM, Swenson B and
Janardhan V: Blood-brain barrier, reperfusion injury, and
hemorrhagic transformation in acute ischemic stroke. Neurology. 79
(13 Suppl 1):S52–S57. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Blesch A: Human ESC-derived interneurons
improve major consequences of spinal cord injury. Cell Stem Cell.
19:423–424. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Davoust C, Plas B, Béduer A, Demain B,
Salabert AS, Sol JC, Vieu C, Vaysse L and Loubinoux I: Regenerative
potential of primary adult human neural stem cells on
micropatterned bio-implants boosts motor recovery. Stem Cell Res
Ther. 8:2532017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Feng Y, Ju Y, Cui J and Wang L: Bone
marrow stromal cells promote neuromotor functional recovery, via
upregulation of neurotrophic factors and synapse proteins following
traumatic brain injury in rats. Mol Med Rep. 16:654–660. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hsueh YY, Chang YJ, Huang CW, Handayani F,
Chiang YL, Fan SC, Ho CJ, Kuo YM, Yang SH, Chen YL, et al: Synergy
of endothelial and neural progenitor cells from adipose-derived
stem cells to preserve neurovascular structures in rat
hypoxic-ischemic brain injury. Sci Rep. 5:149852015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Park KJ, Park E, Liu E and Baker AJ: Bone
marrow-derived endothelial progenitor cells protect postischemic
axons after traumatic brain injury. J Cereb Blood Flow Metab.
34:357–366. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gennai S, Monsel A, Hao Q, Liu J, Gudapati
V, Barbier EL and Lee JW: Cell-based therapy for traumatic brain
injury. Br J Anaesth. 115:203–212. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Banerjee S, Williamson DA, Habib N and
Chataway J: The potential benefit of stem cell therapy after
stroke: An update. Vasc Health Risk Manag. 8:569–580. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Steinberg GK, Kondziolka D, Wechsler LR,
Lunsford LD, Coburn ML, Billigen JB, Kim AS, Johnson JN, Bates D,
King B, et al: Clinical outcomes of transplanted modified bone
marrow-derived mesenchymal stem cells in stroke: A phase 1/2a
study. Stroke. 47:1817–1824. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hao L, Zou Z, Tian H, Zhang Y, Zhou H and
Liu L: Stem cell-based therapies for ischemic stroke. Biomed Res
Int. 2014:4687482014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
An SS, Jin HL, Kim KN, Kim DS, Cho J, Liu
ML, Oh JS, Yoon DH, Lee MH and Ha Y: Neuroprotective effect of
combined hypoxia-induced VEGF and bone marrow-derived mesenchymal
stem cell treatment. Childs Nerv Syst. 26:323–331. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Eckert MA, Vu Q, Xie K, Yu J, Liao W,
Cramer SC and Zhao W: Evidence for high translational potential of
mesenchymal stromal cell therapy to improve recovery from ischemic
stroke. J Cereb Blood Flow Metab. 33:1322–1334. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hsuan YC, Lin CH, Chang CP and Lin MT:
Mesenchymal stem cell-based treatments for stroke, neural trauma,
and heat stroke. Brain Behav. 6:e005262016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Doetsch F: A niche for adult neural stem
cells. Curr Opin Genet Dev. 13:543–550. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Koh SH and Lo EH: The role of the PI3K
pathway in the regeneration of the damaged brain by neural stem
cells after cerebral infarction. J Clin Neurol. 11:297–304. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Temple S: Division and differentiation of
isolated CNS blast cells in microculture. Nature. 340:471–473.
1989. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gage FH: Mammalian neural stem cells.
Science. 287:1433–1438. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu ZH, Yang G, Zhao T, Cao GJ, Xiong L,
Xia W, Huang X, Wu LY, Wu K, Fan M, et al: Small ncRNA expression
and regulation under hypoxia in neural progenitor cells. Cell Mol
Neurobiol. 31:1–5. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yao L and Li Y: The role of direct current
electric field-guided stem cell migration in neural regeneration.
Stem Cell Rev. 12:365–375. 2016. View Article : Google Scholar
|
|
20
|
Abe M and Bonini NM: MicroRNAs and
neurodegeneration: Role and impact. Trends Cell Biol. 23:30–36.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ivan M and Huang X: miR-210: Fine-tuning
the hypoxic response. Adv Exp Med Biol. 772:205–227. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pulkkinen K, Malm T, Turunen M, Koistinaho
J and Ylä-Herttuala S: Hypoxia induces microRNA miR-210 in vitro
and in vivo ephrin-A3 and neuronal pentraxin 1 are potentially
regulated by miR-210. FEBS Lett. 582:2397–2401. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Qiu J, Zhou XY, Zhou XG, Cheng R, Liu HY
and Li Y: Neuroprotective effects of microRNA-210 against
oxygen-glucose deprivation through inhibition of apoptosis in PC12
cells. Mol Med Rep. 7:1955–1959. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lou YL, Guo F, Liu F, Gao FL, Zhang PQ,
Niu X, Guo SC, Yin JH, Wang Y and Deng ZF: miR-210 activates notch
signaling pathway in angiogenesis induced by cerebral ischemia. Mol
Cell Biochem. 370:45–51. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Volný O, Kašičková L, Coufalová D,
Cimflová P and Novák J: microRNAs in cerebrovascular disease. Adv
Exp Med Biol. 888:155–195. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rink C and Khanna S: MicroRNA in ischemic
stroke etiology and pathology. Physiol Genomics. 43:521–528. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhu J, Zhou Z, Liu Y and Zheng J:
Fractalkine and CX3CR1 are involved in the migration of
intravenously grafted human bone marrow stromal cells toward
ischemic brain lesion in rats. Brain Res. 1287:173–183. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang Y, Zheng J, Zhou Z, Zhou H, Wang Y,
Gong Z and Zhu J: Fractalkine promotes chemotaxis of bone
marrow-derived mesenchymal stem cells towards ischemic brain
lesions through Jak2 signaling and cytoskeletal reorganization.
FEBS J. 282:891–903. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiang Y, Li L, Tan X, Liu B, Zhang Y and
Li C: miR-210 mediates vagus nerve stimulation-induced antioxidant
stress and anti-apoptosis reactions following cerebral
ischemia/reperfusion injury in rats. J Neurochem. 134:173–181.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang D, Cao X, Li J and Zhao G: MiR-210
inhibits NF-κB signaling pathway by targeting DR6 in
osteoarthritis. Sci Rep. 5:127752015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cao L, Zhang X, Cao F, Wang Y, Shen Y,
Yang C, Uzan G, Peng B and Zhang D: Inhibiting inducible miR-223
further reduces viable cells in human cancer cell lines MCF-7 and
PC3 treated by celastrol. BMC Cancer. 15:8732015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Brown JP, Couillard-Després S, Cooper-Kuhn
CM, Winkler J, Aigner L and Kuhn HG: Transient expression of
doublecortin during adult neurogenesis. J Comp Neurol. 467:1–10.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Croft AP and Przyborski SA: Mesenchymal
stem cells expressing neural antigens instruct a neurogenic cell
fate on neural stem cells. Exp Neurol. 216:329–341. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kuroda S: Current opinion of bone marrow
stromal cell transplantation for ischemic stroke. Neurol Med Chir
(Tokyo). 56:293–301. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Alexanian AR: Neural stem cells induce
bone-marrow-derived mesenchymal stem cells to generate neural
stem-like cells via juxtacrine and paracrine interactions. Exp Cell
Res. 310:383–391. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bai L, Caplan A, Lennon D and Miller RH:
Human mesenchymal stem cells signals regulate neural stem cell
fate. Neurochem Res. 32:353–362. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu X, Ye R, Yan T, Yu SP, Wei L, Xu G,
Fan X, Jiang Y, Stetler RA, Liu G and Chen J: Cell based therapies
for ischemic stroke: From basic science to bedside. Prog Neurobiol.
115:92–115. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lu J, Manaenko A and Hu Q: Targeting adult
neurogenesis for poststroke therapy. Stem Cells Int.
2017:58686322017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Haragopal H, Yu D, Zeng X, Kim SW, Han IB,
Ropper AE, Anderson JE and Teng YD: Stemness enhancement of human
neural stem cells following bone marrow MSC coculture. Cell
Transplant. 24:645–659. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang Y, Tu W, Lou Y, Xie A, Lai X, Guo F
and Deng Z: Mesenchymal stem cells regulate the proliferation and
differentiation of neural stem cells through Notch signaling. Cell
Biol Int. 33:1173–1179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hackstein H, Tschipakow I, Bein G, Nold P,
Brendel C and Baal N: Contact-dependent abrogation of bone
marrow-derived plasmacytoid dendritic cell differentiation by
murine mesenchymal stem cells. Biochem Biophys Res Commun.
476:15–20. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yamada T, Yuasa M, Masaoka T, Taniyama T,
Maehara H, Torigoe I, Yoshii T, Shinomiya K, Okawa A and Sotome S:
After repeated division, bone marrow stromal cells express
inhibitory factors with osteogenic capabilities, and EphA5 is a
primary candidate. Bone. 57:343–354. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang PP, Xie DY, Liang XJ, Peng L, Zhang
GL, Ye YN, Xie C and Gao ZL: HGF and direct mesenchymal stem cells
contact synergize to inhibit hepatic stellate cells activation
through TLR4/NF-kB pathway. PLoS One. 7:e434082012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Huang X, Ding L, Bennewith KL, Tong RT,
Welford SM, Ang KK, Story M, Le QT and Giaccia AJ:
Hypoxia-inducible mir-210 regulates normoxic gene expression
involved in tumor initiation. Mol Cell. 35:856–867. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Xiong L, Wang F, Huang X, Liu ZH, Zhao T,
Wu LY, Wu K, Ding X, Liu S, Wu Y, et al: DNA demethylation
regulates the expression of miR-210 in neural progenitor cells
subjected to hypoxia. FEBS J. 279:4318–4326. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Nie Y, Han BM, Liu XB, Yang JJ, Wang F,
Cong XF and Chen X: Identification of MicroRNAs involved in
hypoxia- and serum deprivation-induced apoptosis in mesenchymal
stem cells. Int J Biol Sci. 7:762–768. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sun G and Peng H: HIF-1α-induced
microRNA-210 reduces hypoxia-induced osteoblast MG-63 cell
apoptosis. Biosci Biotechnol Biochem. 79:1232–1239. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang F, Xiong L, Huang X, Zhao T, Wu LY,
Liu ZH, Ding X, Liu S, Wu Y, Zhao Y, et al: miR-210 suppresses
BNIP3 to protect against the apoptosis of neural progenitor cells.
Stem Cell Res. 11:657–667. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhang C, Tian W, Meng L, Qu L and Shou C:
PRL-3 promotes gastric cancer migration and invasion through a
NF-κB-HIF-1α-miR-210 axis. J Mol Med (Berl). 94:401–415. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Qu A, Du L, Yang Y, Liu H, Li J, Wang L,
Liu Y, Dong Z, Zhang X, Jiang X, et al: Hypoxia-inducible MiR-210
is an independent prognostic factor and contributes to metastasis
in colorectal cancer. PLoS One. 9:e909522014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zeng L, He X, Wang Y, Tang Y, Zheng C, Cai
H, Liu J, Wang Y, Fu Y and Yang GY: MicroRNA-210 overexpression
induces angiogenesis and neurogenesis in the normal adult mouse
brain. Gene Ther. 21:37–43. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Liu F, Lou YL, Wu J, Ruan QF, Xie A, Guo
F, Cui SP, Deng ZF and Wang Y: Upregulation of microRNA-210
regulates renal angiogenesis mediated by activation of VEGF
signaling pathway under ischemia/perfusion injury in vivo and in
vitro. Kidney Blood Press Res. 35:182–191. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kawanishi Y, Nakasa T, Shoji T, Hamanishi
M, Shimizu R, Kamei N, Usman MA and Ochi M: Intra-articular
injection of synthetic microRNA-210 accelerates avascular meniscal
healing in rat medial meniscal injured model. Arthritis Res Ther.
16:4882014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Díaz-Coránguez M, Segovia J, López-Ornelas
A, Puerta-Guardo H, Ludert J, Chávez B, Meraz-Cruz N and
González-Mariscal L: Transmigration of neural stem cells across the
blood brain barrier induced by glioma cells. PLoS One.
8:e606552013. View Article : Google Scholar : PubMed/NCBI
|