Introduction
Lung cancer is a malignant cancer, which
demonstrates the highest rates of morbidity and mortality worldwide
(1); the most common histological
subtype is adenocarcinoma (2).
Although the diagnosis and treatment of lung cancer has progressed
in recent decades, the survival rate has not significantly improved
(3). A total of ~80% of patients
with lung cancer are diagnosed during the advanced stages of the
disease, when treatment strategies are no longer optimal (4). Tumor metastasis is the leading cause
of death in lung adenocarcinoma (LUAD); therefore, further
investigation into the molecular mechanisms underlying LUAD
metastasis is required for the identification of potential
therapeutic targets for the disease.
Cathepsins are the key acid hydrolases in lysosomes
and the main effector enzymes of protein catabolism and autophagy
(5,6). Cathepsin A (CTSA) is a serine
protease member of the cathepsin family that displays
carboxypeptidase, deaminase and esterase activity and regulates the
function of bioactive peptides (7,8).
CTSA protects β-galactosidase and neuraminidase proteins from
lysosomal proteolysis by forming multienzyme complexes (9). CTSA deficiency can cause human
lysosomal storage disease, known as galactosialidosis (10). A recent study demonstrated that
CTSA is a key enzyme involved in the degradation of
lysosome-associated membrane protein type 2a, which is an important
effector of chaperone-mediated autophagy (11). Several studies have confirmed that
cathepsin proteases regulate cancer progression and the therapeutic
response (12–14). High CTSA expression is associated
with various tumors (15–17); however, whether abnormal expression
of CTSA is associated with LUAD development is not completely
understood.
The Cancer Genome Atlas (TCGA) is a public-funded
project, aimed at cataloguing and discovering the major human
genome variations that induce cancerization (18). By analyzing TCGA database,
differential expression data of CTSA in LUAD and normal tissues can
be obtained, which provides a foundation for the investigation of
the roles of CTSA during LUAD molecular pathogenesis.
In the present study, the aim was to investigate
whether CTSA knockdown decreased proliferation, migration and
invasion of A549 cells, and to assess the oncogenic potential of
CTSA in LUAD.
Materials and methods
Data source and bioinformatics
analysis
Human RNA-sequencing data from LUAD projects which
included 515 patients with LUAD and 59 normal tissues were
collected from TCGA database (portal.gdc.cancer.gov). The R software package
(version 3.6.0; RStudio, Inc.) was used to analyze the expression
of CTSA in normal and tumor samples.
Cell culture
The human lung cancer cell line A549 was obtained
from The Cell Bank of Type Culture Collection of the Chinese
Academy of Sciences. Cells were cultured in RPMI-1640 medium
(Biological Industries) supplemented with 10% FBS (Biological
Industries) at 37°C with 5% CO2 and 95% air.
Cell transfection
A total of 5×105 A549 cells/well were
transfected with 50 nM small interfering (si)RNA-CTSA
(5′-GCCUGCCACUCAAGCGGAUTT-3′) or siRNA-negative control (NC;
non-targeting; 5′-UUCUCCGAACGUGUCACGUTT-3′; Shanghai GenePharma
Co., Ltd.) using Lipofectamine® 3000 (Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Following a 48-h incubation at 37°C, cells were harvested for
subsequent experiments.
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from A549 cells using the
RNAiso Plus kit (Takara Biotechnology Co., Ltd.), according to the
manufacturer's protocol. RNA purity was assessed using a
NanoPhotometer® spectrophotometer. Total RNA was reverse
transcribed into cDNA using the PrimeScript RT reagent kit with
gDNA Eraser (Takara Biotechnology Co., Ltd.), according to the
manufacturer's protocol. The following experimental conditions were
used for reverse transcription: 42°C for 2 min, 37°C for 15 min and
85°C for 5 sec. Subsequently, qPCR was performed using an ABI 7500
Fast Real-Time PCR system (Thermo Fisher Scientific, Inc.) and the
SYBR Premix Ex Taq™ II kit (Takara Biotechnology Co., Ltd),
according to the manufacturer's protocol. The following primer
pairs were used for qPCR: CTSA forward, 5′-GTCGCCCAGAGCAATTTTGAG-3′
and reverse, 5′-TCTCCCCGGTCAGGAAAAGTT-3′; and β-actin forward,
5′-CATGTACGTTGCTATCCAGGC-3′ and reverse,
5′-CTCCTTAATGTCACGCACGAT-3′. The following thermocycling conditions
were used for qPCR: Initial denaturation at 95°C for 30 sec;
followed by 40 cycles of 95°C for 5 sec and 60°C for 34 sec. CTSA
mRNA levels were quantified using the 2−ΔΔCq method and
normalized to the internal reference gene β-actin.
Cell proliferation assay
The effect of CTSA knockdown on cell proliferation
was assessed using a Cell Counting Kit-8 (CCK-8) assay (Biosharp
Life Sciences), according to the manufacturer's protocol. A549
cells were harvested 48 h post-transfection, seeded
(5×103 cells/well) into a 96-well microplate and
incubated at 37°C for 24, 48 or 72 h. Subsequently, 10% CCK-8
solution (10 µl CCK-8 reagent; 90 µl RPMI 1640 medium) was added to
each well for 2 h at 37°C. The optical density of each well was
determined at a wavelength of 450 nm using an Elx808 microplate
reader (BioTek Instruments, Inc.).
Cell cycle analysis
Transfected A549 cells (2×106 cells/well)
were seeded into 6-well plates, incubated overnight and cultured in
RPMI-1640 medium without serum for 12 h to synchronize the cell
cycle. The cells were fixed with 70% ethanol for 12 h at 4°C,
washed with 1 ml pre-cooled PBS and stained with propidium iodide
and RNase A solution (25 µg/ml; Beijing 4A Biotech Co., Ltd.) for
30 min at 37°C in the dark. The DNA content of the cells, used as
an indicator of the different phases of the cell cycle, was
measured using a FACSCalibur flow cytometer (BD Biosciences;
Becton, Dickinson and Company) with CellQuest Pro (version 5.2; BD
Biosciences; Becton, Dickinson and Company) and ModFit LT (version
3.0; Verity Software House, Inc.) software.
Western blot analysis
Total protein was extracted from A549 cells using
RIPA lysis buffer (Beyotime Institute of Biotechnology)
supplemented with phenylmethylsulphonyl fluoride (Beyotime
Institute of Biotechnology). Proteins were isolated by
centrifugation at 12,000 × g for 15 min at 4°C. Total protein was
quantified using a bicinchoninic acid assay (Beyotime Institute of
Biotechnology). Subsequently, protein samples were incubated with
Loading Buffer (Beyotime Institute of Biotechnology) at 100°C for
10 min. Equal amounts of protein (30 µg per lane) were separated
via 10% SDS-PAGE for 2 h at a constant voltage (110 V), and
subsequently transferred onto PVDF membranes (EMD Millipore). The
membranes were blocked using TBST (20 mM Tris-Hcl, 150 mM NaCl pH
7.5 and 0.1% Tween-20) containing 5% skim milk at room temperature
for 1–2 h. Subsequently, the membranes were incubated overnight at
4°C with the following primary antibodies: Anti-CTSA (cat. no.
ab184553; 1:10,000; Abcam), anti-E-cadherin (cat. no. WL01482;
1:1,000; Wanleibio Co., Ltd.), anti-p53 (cat. no. WL01919; 1:1,000;
Wanleibio Co., Ltd.), anti-p21 (cat. no. WL0362; 1:1,000; Wanleibio
Co., Ltd.), anti-proliferating cell nuclear antigen (PCNA; cat. no.
WL01482; 1:1,000; Wanleibio Co., Ltd.), anti-N-cadherin (cat. no.
4061S; 1:1,000; Cell Signaling Technology, Inc.), anti-β-catenin
(cat. no. AF6266; 1:1,000; Affinity Biosciences) and anti-β-actin
(cat. no. 60008-1-lg; 1:4,000; ProteinTech, Group, Inc.). Following
primary incubation, the membranes were washed three times with TBST
and incubated with horseradish peroxidase (HRP)-conjugated goat
anti-rabbit immunoglobulin G (IgG; cat. no. ZB-2301; 1:20,000;
OriGene Technologies, Inc.) and HRP-conjugated goat anti-mouse IgG
(cat. no. ZB-2305; 1:20,000; OriGene Technologies, Inc.) secondary
antibodies at 25°C for 2 h. Protein bands were visualized using the
Amersham Imager 600 enhanced chemiluminescence detection system (GE
Healthcare Life Sciences) and quantified using Image J software
(version 1.52; National Institutes of Health), with β-actin as the
loading control.
Wound healing assay
The migratory ability of A549 cells was assessed
using wound healing assays. At 48 h post-transfection, A549 cells
were seeded into a 6-well plate and subsequently grown to 90%
confluence. A 200 µl pipette tip was used to make a single scratch
through the cell monolayer and PBS was used to remove debris. A549
cells were cultured in RPMI-1640 medium, supplemented with 2% FBS
to maintain the adherence of the cells for 48 h. Cell migration was
observed at 0, 24 and 48 h using a light microscope (magnification,
×100; Nikon Corporation) and quantified using ImageJ software
(version 1.37; National Institutes of Health). The migration rate
was defined as the percentage of wound closure.
Transwell assay
Transwell assays were used to assess the invasive
ability of A549 cells. Prior to the assay, the 24-well Transwell
plates (8-µM; Corning Life Sciences) were pre-coated with 40 µl
Matrigel® (1.5 mg/ml; BD Biosciences; Becton, Dickinson
and Company) overnight at 37°C. At 48 h post-transfection, A549
cells (5×104) were suspended in 200 µl RPMI-1640 medium
without serum and seeded into the upper chambers of the Transwell
plates. RPMI-1640 medium supplemented with 20% FBS (500 µl) was
added to the lower chambers of the Transwell plates to act as a
chemoattractant stimulus. Subsequently, the Transwell plates were
incubated for 48 h at 37°C. Cells on the upper surface of the
membrane were wiped off using cotton swabs and invading cells on
the lower surface of the membrane were fixed with 4%
paraformaldehyde for 30 min at room temperature, and stained with
0.1% crystal violet (Beijing Solarbio Science & Technology Co.,
Ltd.) for 15 min at room temperature. The invasive ability of A549
cells was calculated under a light microscope as the mean number of
cells in four randomly selected fields of view (magnification,
×100).
Statistical analysis
Statistical analyses were performed using GraphPad
Prism software (version 5.0; GraphPad Software, Inc.). Data are
presented as the mean ± standard deviation. One-way ANOVA followed
by the least significant distance post hoc test was used make
comparisons. All experiments were repeated at least three times.
P<0.05 was considered to indicate a statistically significant
difference.
Results
CTSA expression in LUAD tissues
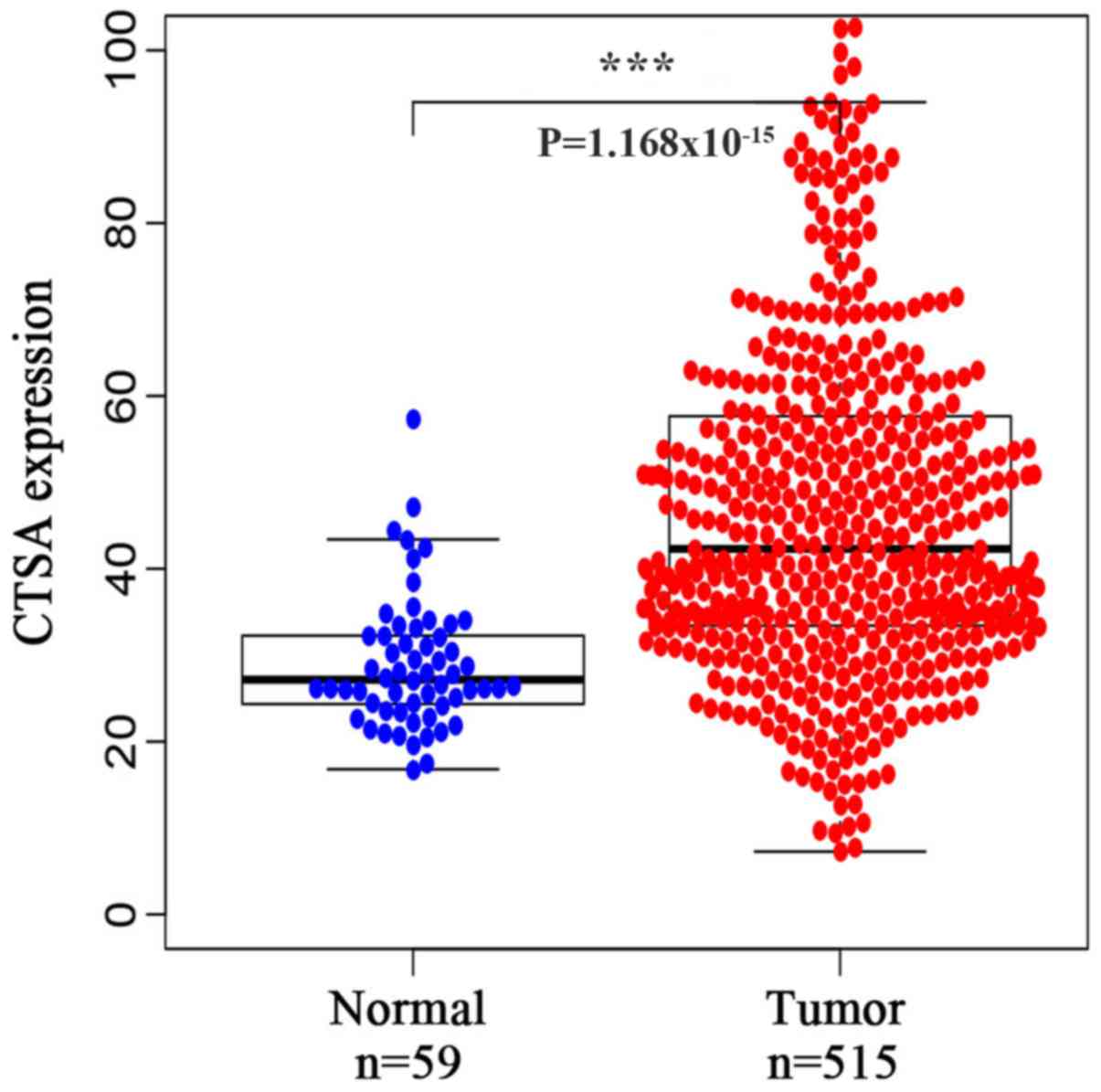
To investigate the expression of CTSA in LUAD, a
bioinformatics analysis of data obtained from TCGA database was
conducted. CTSA mRNA expression was significantly increased in LUAD
tissues compared with normal lung tissues (P<0.001; Fig. 1).
CTSA expression following CTSA
knockdown
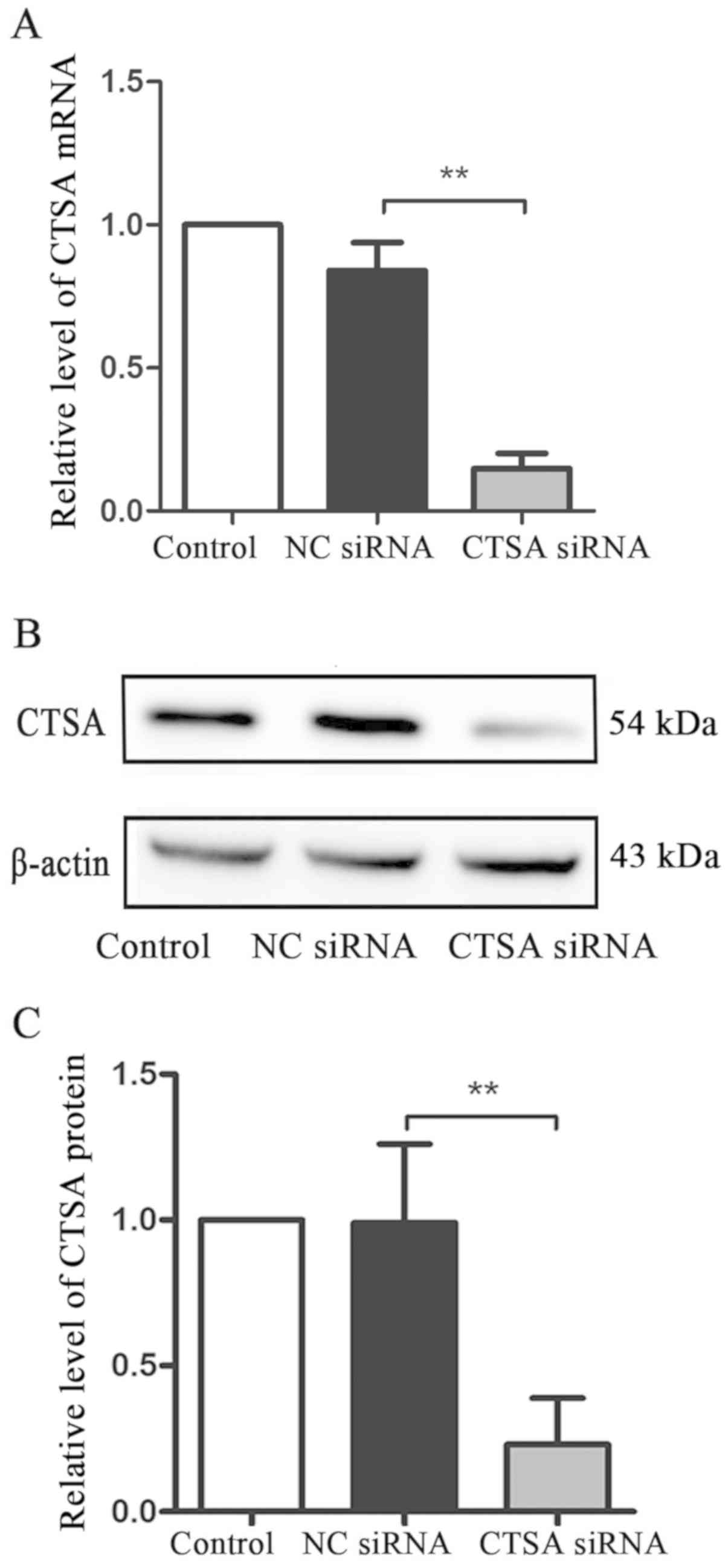
To investigate the effect of CTSA on the biological
characteristics of A549 cells, CTSA siRNA was used to knockdown the
expression of CTSA. Transfection efficiency was demonstrated using
RT-qPCR and western blotting. In A549 cells, CTSA mRNA and protein
expression levels were significantly decreased in CTSA knockdown
cells compared with NC siRNA cells (P<0.01; Fig. 2A-C).
CTSA knockdown decreases the
proliferation of A549 cells
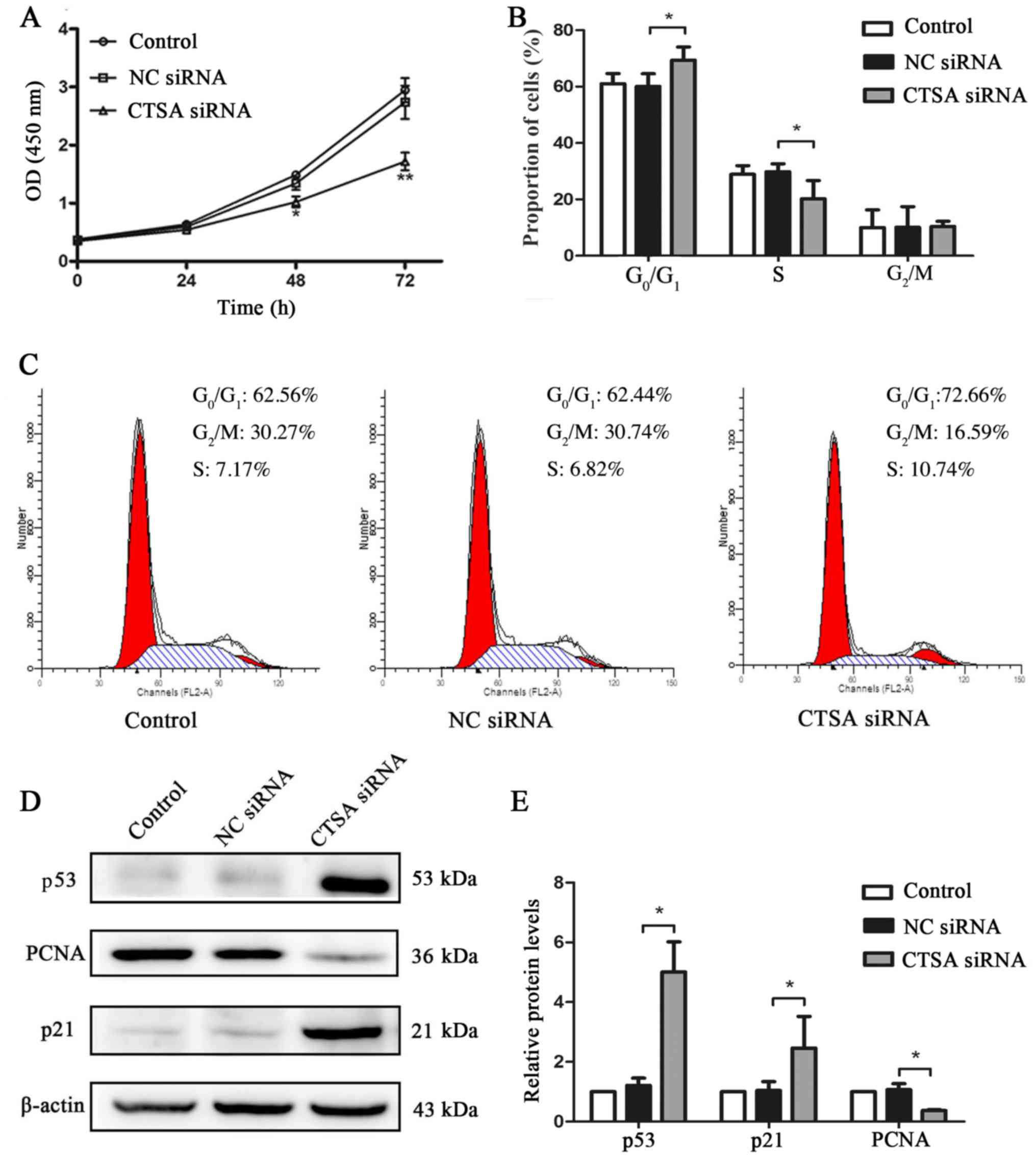
The proliferation of CTSA-knockdown cells was
significantly decreased compared with the negative control cells,
as assessed by the CCK-8 assay (P<0.01; Fig. 3A). CTSA knockdown in A549 cells
resulted in a significant increase in the proportion of cells in
the G0/G1 phase and a decrease in the
proportion of cells in the S phase compared with the negative
control cells (P<0.05; Fig. 3B and
C). In addition, the expression levels of cell proliferation
and cell cycle-associated proteins were detected by western
blotting. The CTSA siRNA group displayed significantly decreased
expression levels of PCNA and significantly increased expression
levels of p53 and p21 compared with the NC siRNA group (P<0.01;
Fig. 3D and E). The results
indicated that CTSA knockdown decreased the proliferation of LUAD
cells.
CTSA knockdown decreases the
migration, invasion and EMT of LUAD cells
The migratory and invasive abilities of
CTSA-knockdown cells were investigated using wound healing and
invasion assays. The siRNA-CTSA group displayed a significant
decrease in migratory (P<0.01; Fig.
4A and B) and invasive (P<0.01; Fig. 4C and D) abilities compared with the
NC siRNA group. The expression of the main proteins involved in EMT
was assessed to investigate the effects of CTSA knockdown on
invasion and migration. The CTSA siRNA group displayed increased
E-cadherin expression (P<0.001), but decreased levels of
N-cadherin (P<0.05) and β-catenin (P<0.01) expression
compared with the NC siRNA group (Fig.
4E and F). The results suggested that CTSA knockdown decreased
the migration and invasion of LUAD cells.
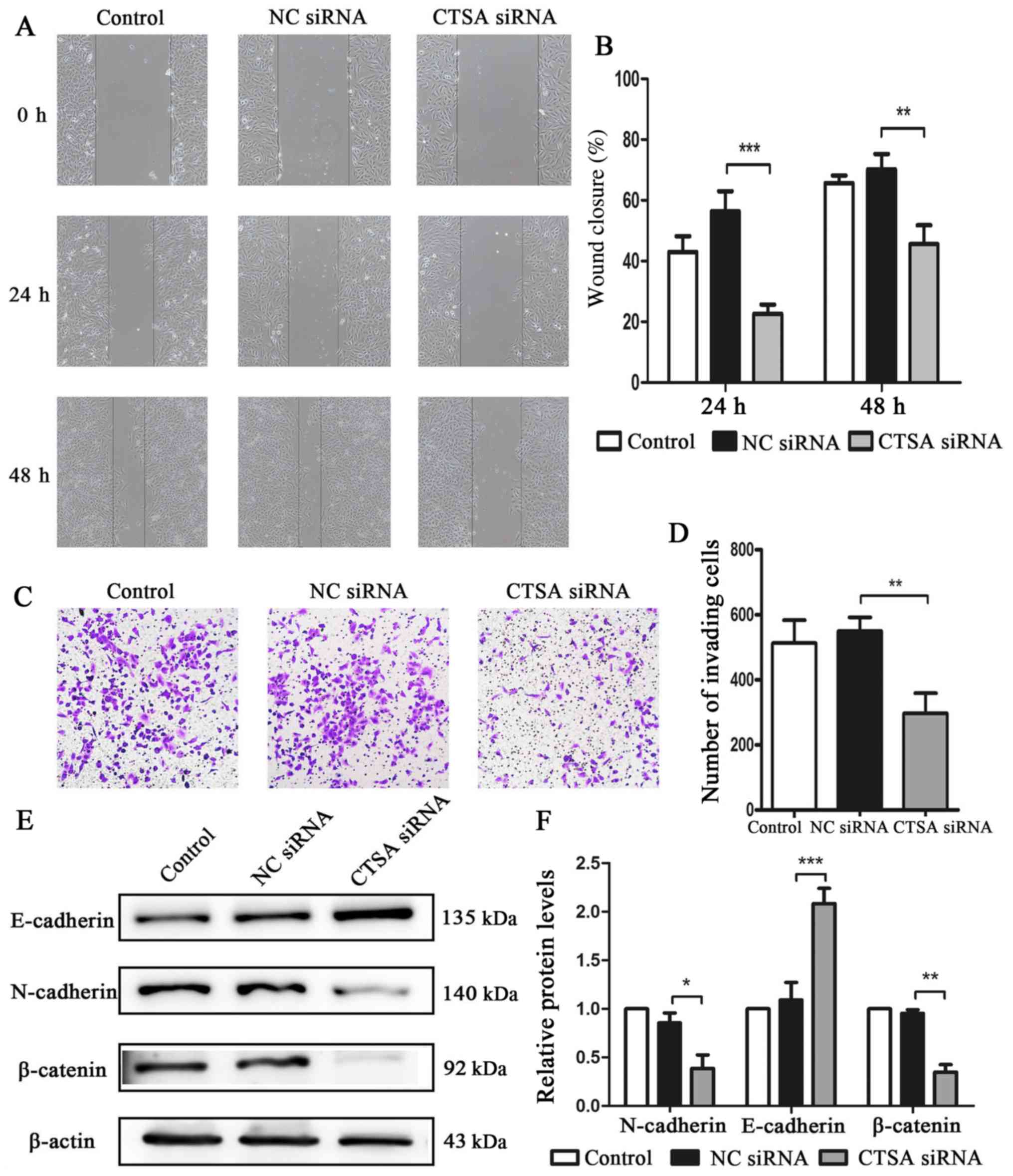 | Figure 4.Effects of CTSA knockdown on the
migration, invasion and EMT of A549 cells. Cell migration was (A)
determined by wound healing assays and (B) quantified
(magnification, ×100). Cell invasion was (C) determined by
Transwell invasion assays and (D) quantified (magnification, ×100).
Protein expression levels of E-cadherin, N-cadherin and β-catenin
were (E) determined by western blotting and (F) quantified.
*P<0.05, **P<0.01 and ***P<0.001, as indicated. CTSA,
cathepsin A; EMT, epithelial-mesenchymal transition; NC, negative
control; siRNA, small interfering RNA. |
Discussion
Lung cancer is one of the most common types of
cancer and is a major cause of cancer-related mortality.
Proliferation and invasion play important roles during tumor
development, and the ability to inhibit tumor growth and metastasis
is key to successful treatment (19). Although various therapeutic targets
for lung cancer have been identified, the survival rate has not
significantly improved (20).
However, there are only a few targets that can inhibit multiple
tumor progression processes; therefore, it is critical to identify
further pleiotropic therapeutic targets for LUAD.
Cathepsins, as lysosomal proteases, participate in
multiple processes associated with cancer, including protein
degradation, autophagy, growth factor receptor recycling and
lysosome-mediated cell death. Abnormal expression of cathepsins has
been reported to be associated with a variety of different types of
cancer (8,21–23).
For example, previous studies have reported that abnormal
expression of cathepsins, including cathepsin K, L and S, is
observed in lung cancer (24–26),
which suggests that variations in cathepsin expression may play a
role in lung cancer. A number of studies have indicated that CTSA
is involved in breast, colorectal and melanocyte cancer, and is
also associated with tumorigenesis, malignant progression and poor
patient prognosis (15–17). However, the role of CTSA in LUAD
has not been reported. In the present study, the bioinformatics
analysis of TCGA database suggested that the expression of CTSA was
upregulated in LUAD tissues compared with normal tissues,
suggesting that high CTSA expression was associated with LUAD.
In various types of cancer, cathepsins are highly
upregulated and their increased expression correlates with more
aggressive tumor types and poor patient prognosis (8,27–33).
Cathepsins contribute to the degradation of the basement membrane
that confines the tumor to promote invasion and tumor metastasis
(34). The expression levels of
CTSA are higher in metastatic melanoma compared with primary
melanoma and are associated with poor prognosis (17). Similarly, higher CTSA expression is
associated with aggressive tumor types and poor outcomes in breast
and colorectal cancer (CRC), and reducing CTSA expression inhibits
the migration and invasion of CRC cells (15,16).
CTSA knockdown led to a reduction in the migration
and invasion of LUAD cells. EMT, which is a cellular process that
involves the loss of polarization and intercellular connections in
epithelial cells, plays an important role in tumor metastasis and
recurrence (35). A key
characteristic of EMT is the loss of E-cadherin expression and the
gain of N-cadherin expression in the cell membrane, which is known
as the E-to-N-cadherin switch (36). E-cadherin has been identified as a
substrate for CTSB, CTSL and CTSS in tumors (37). CTSA is highly expressed in A549
cells; therefore, overexpression of CTSA in A549 cells was not
performed in the present study. However, CTSA knockdown was
performed to investigate the relationship between CTSA and EMT in
LUAD cells. The results suggested that CTSA knockdown increased the
expression of the epithelial marker E-cadherin and decreased the
expression of N-cadherin and β-catenin in A549 cells.
The catabolism of proteins is one of the major
functions of proteases. Cathepsin deficiencies result in
autophagosome accumulation, which indicates impaired catabolism of
the cargo and decreased autophagic flux (6). Increased lysosomal biogenesis and
cathepsin expression in cancer cells supports cell survival
(38). Furthermore, increased
catabolic activity is essential to provide the necessary nutrients
to maintain cell proliferation. Therefore, it was hypothesized that
the loss of cathepsin may inhibit cell proliferation by impairing
the catabolic activity and the lack of essential nutrients. In the
present study, CTSA knockdown decreased the proliferation and
increased cell cycle arrest at the G0/G1
phase in LUAD cells. Further experiments suggested that CTSA
knockdown altered the expression of critical cell cycle-associated
proteins, including p53, p21 and the proliferative marker PCNA. p53
is a transcription factor that initiates cell cycle arrest, cell
senescence or apoptosis to regulate tumor progression. p21 is
involved in p53-mediated cycle arrest, which inhibits cyclin E-CDK2
activity (39).
To the best of our knowledge, the present study
suggested for the first time that CTSA may serve as an oncogene in
LUAD. CTSA knockdown decreased the proliferation, migration and
invasion of A549 cells, and altered the expression of key cell
cycle regulators. However, in order to maintain the cells in a good
adherence state, 2% FBS was used in the wound healing assay, which
is a limitation of the present study in terms of assessing the
migratory ability. CTSA also displayed pleiotropic effects in A549
cells, which suggested that CTSA may serve as a vital effector
during tumor development, indicating that targeting CTSA may allow
multiple tumorigenic processes to be inhibited via a single
therapeutic target. The results of the present study suggested that
CTSA may serve as a potential therapeutic target for LUAD; however,
further investigation into the mechanisms underlying the role of
CTSA in LUAD, including in vitro and in vivo
experiments, is required.
Acknowledgements
Not applicable.
Funding
The present study was supported by The China Medical
University Discipline Enhancement Program Support Project.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request. The datasets generated and/or analyzed during the current
study are available in the TCGA repository, (http://cancergenome.nih.gov).
Authors' contributions
JL, BH and XZ conceived and designed the study. BH
performed the experiments and wrote the manuscript. BH and XZ
analyzed the data and critically revised the manuscript. JL and XZ
supervised the study and revised the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2020. CA Cancer J Clin. 70:7–30. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cancer Genome Atlas Research Network, .
Comprehensive molecular profiling of lung adenocarcinoma. Nature.
511:543–550. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Janssen-Heijnen ML, van Erning FN, De
Ruysscher DK, Coebergh JW and Groen HJ: Variation in causes of
death in patients with non-small cell lung cancer according to
stage and time since diagnosis. Ann Oncol. 26:902–907. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Soda M, Choi YL, Enomoto M, Takada S,
Yamashita Y, Ishikawa S, Fujiwara S, Watanabe H, Kurashina K,
Hatanaka H, et al: Identification of the transforming EML4-ALK
fusion gene in non-small-cell lung cancer. Nature. 448:561–566.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pan L, Li Y, Jia L, Qin Y, Qi G, Cheng J,
Qi Y, Li H and Du J: Cathepsin S deficiency results in abnormal
accumulation of autophagosomes in macrophages and enhances Ang
II-induced cardiac inflammation. PLoS One. 7:e353152012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dennemärker J, Lohmüller T, Müller S,
Aguilar SV, Tobin DJ, Peters C and Reinheckel T: Impaired turnover
of autophagolysosomes in cathepsin L deficiency. Biol Chem.
391:913–922. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Drake MT, Clarke BL, Oursler MJ and Khosla
S: Cathepsin K inhibitors for osteoporosis: Biology, potential
clinical utility, and lessons learned. Endocr Rev. 38:325–350.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Olson OC and Joyce JA: Cysteine cathepsin
proteases: Regulators of cancer progression and therapeutic
response. Nat Rev Cancer. 15:712–729. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Akkari L, Gocheva V, Quick ML, Kester JC,
Spencer AK, Garfall AL, Bowman RL and Joyce JA: Combined deletion
of cathepsin protease family members reveals compensatory
mechanisms in cancer. Genes Dev. 30:220–232. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kolli N and Garman SC: Proteolytic
activation of human cathepsin A. J Biol Chem. 289:11592–11600.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
You Y, Li WZ, Zhang S, Hu B, Li YX, Li HD,
Tang HH, Li QW, Guan YY, Liu LX, et al: SNX10 mediates
alcohol-induced liver injury and steatosis by regulating the
activation of chaperone-mediated autophagy. J Hepatol. 69:129–141.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Verbovšek U, Van Noorden CJ and Lah TT:
Complexity of cancer protease biology: Cathepsin K expression and
function in cancer progression. Semin Cancer Biol. 35:71–84. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sudhan DR and Siemann DW: Cathepsin L
targeting in cancer treatment. Pharmacol Ther. 155:105–116. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yuan L, Sheng L, He W, Zou C, Hu B, Liu J,
Ge W, Liu Y, Wang J and Ma E: Discovery of novel cathepsin
inhibitors with potent anti-metastatic effects in breast cancer
cells. Bioorg Chem. 81:672–680. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ni S, Weng W, Xu M, Wang Q, Tan C, Sun H,
Wang L, Huang D, Du X and Sheng W: miR-106b-5p inhibits the
invasion and metastasis of colorectal cancer by targeting CTSA.
Onco Targets Ther. 11:3835–3845. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Toss MS, Miligy IM, Haj-Ahmad R, Gorringe
KL, AlKawaz A, Mittal K, Ellis IO, Green AR and Rakha EA: The
prognostic significance of lysosomal protective protein (cathepsin
A) in breast ductal carcinoma in situ. Histopathology.
74:1025–1035. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kozlowski L, Wojtukiewicz MZ and Ostrowska
H: Cathepsin A activity in primary and metastatic human melanocytic
tumors. Arch Dermatol Res. 292:68–71. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cancer Genome Atlas Research Network, .
Comprehensive genomic characterization of squamous cell lung
cancers. Nature. 489:519–525. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Molina JR, Yang P, Cassivi SD, Schild SE
and Adjei AA: Non-small cell lung cancer: Epidemiology, risk
factors, treatment, and survivorship. Mayo Clin Proc. 83:584–594.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hung JJ, Yeh YC, Jeng WJ, Wu KJ, Huang BS,
Wu YC, Chou TY and Hsu WH: Predictive value of the international
association for the study of lung cancer/American thoracic
society/European respiratory society classification of lung
adenocarcinoma in tumor recurrence and patient survival. J Clin
Oncol. 32:2357–2364. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Reinheckel T, Hagemann S, Dollwet-Mack S,
Martinez E, Lohmuller T, Zlatkovic G, Tobin DJ, Maas-Szabowski N
and Peters C: The lysosomal cysteine protease cathepsin L regulates
keratinocyte proliferation by control of growth factor recycling. J
Cell Sci. 118:3387–3395. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Platt MO and Shockey WA: Endothelial cells
and cathepsins: Biochemical and biomechanical regulation.
Biochimie. 122:314–323. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Brojatsch J, Lima H Jr, Palliser D,
Jacobson LS, Muehlbauer SM, Furtado R, Goldman DL, Lisanti MP and
Chandran K: Distinct cathepsins control necrotic cell death
mediated by pyroptosis inducers and lysosome-destabilizing agents.
Cell Cycle. 14:964–972. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang L, Zhao Y, Xiong Y, Wang W, Fei Y,
Tan C and Liang Z: K-ras mutation promotes ionizing
radiation-induced invasion and migration of lung cancer in part via
the Cathepsin L/CUX1 pathway. Exp Cell Res. 362:424–435. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Willumsen N, Bager CL, Leeming DJ,
Bay-Jensen AC and Karsdal MA: Nidogen-1 degraded by cathepsin S can
be quantified in serum and is associated with non-small cell lung
cancer. Neoplasia. 19:271–278. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tsai JY, Lee MJ, Dah-Tsyr Chang M and
Huang H: The effect of catalase on migration and invasion of lung
cancer cells by regulating the activities of cathepsin S, L, and K.
Exp Cell Res. 323:28–40. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jechorek D, Votapek J, Meyer F, Kandulski
A, Roessner A and Franke S: Characterization of cathepsin X in
colorectal cancer development and progression. Pathol Res Pract.
210:822–829. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang W, Wang S, Wang Q, Yang Z, Pan Z and
Li L: Overexpression of cysteine cathepsin L is a marker of
invasion and metastasis in ovarian cancer. Oncol Rep. 31:1334–1342.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yamashita K, Iwatake M, Okamoto K, Yamada
SI, Umeda M and Tsukuba T: Cathepsin K modulates invasion,
migration and adhesion of oral squamous cell carcinomas in vitro.
Oral Dis. 23:518–525. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Aggarwal N and Sloane BF: Cathepsin B:
Multiple roles in cancer. Proteomics Clin Appl. 8:427–437. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schweiger A, Staib A, Werle B, Krasovec M,
Lah TT, Ebert W, Turk V and Kos J: Cysteine proteinase cathepsin H
in tumours and sera of lung cancer patients: Relation to prognosis
and cigarette smoking. Br J Cancer. 82:782–788. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ruan J, Zheng H, Fu W, Zhao P, Su N and
Luo R: Increased expression of cathepsin L: A novel independent
prognostic marker of worse outcome in hepatocellular carcinoma
patients. PLoS One. 9:e1121362014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Han ML, Zhao YF, Tan CH, Xiong YJ, Wang
WJ, Wu F, Fei Y, Wang L and Liang ZQ: Cathepsin L
upregulation-induced EMT phenotype is associated with the
acquisition of cisplatin or paclitaxel resistance in A549 cells.
Acta Pharmacol Sin. 37:1606–1622. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Turk V, Turk B, Guncar G, Turk D and Kos
J: Lysosomal cathepsins: Structure, role in antigen processing and
presentation, and cancer. Adv Enzyme Regul. 42:285–303. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pastushenko I and Blanpain C: EMT
Transition states during tumor progression and metastasis. Trends
Cell Biol. 29:212–226. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zeisberg M and Neilson EG: Biomarkers for
epithelial-mesenchymal transitions. J Clin Invest. 119:1429–1437.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gocheva V, Zeng W, Ke D, Klimstra D,
Reinheckel T, Peters C, Hanahan D and Joyce JA: Distinct roles for
cysteine cathepsin genes in multistage tumorigenesis. Genes Dev.
20:543–556. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kallunki T, Olsen OD and Jäättelä M:
Cancer-associated lysosomal changes: Friends or foes? Oncogene.
32:1995–2004. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Harris SL and Levine AJ: The p53 pathway:
Positive and negative feedback loops. Oncogene. 24:2899–2908. 2005.
View Article : Google Scholar : PubMed/NCBI
|