Introduction
Neurodegenerative diseases, including Parkinson's
disease, Alzheimer's disease and amyotrophic lateral sclerosis, are
a major cause of disability and death in elderly individuals, which
seriously impact the physical and mental health of the affected
individual (1–4). With an increasing aged population and
the pressure of life, neurological diseases pose a large burden on
an individual's life and health; therefore, identifying drugs that
protect against neurodegeneration is important.
Ginsenosides are the active ingredients of ginseng,
which has been used as a tonic drug in East Asia for >2,000
years (5). In recent years, a
number of studies have reported that ginsenosides have certain
roles in the prevention and treatment of neurological diseases. For
example, Ginsenoside Rb1 promoted the proliferation and
differentiation of neural stem cells (NSCs) in a rat model of
Alzheimer's disease (6). Gerbil
models of global ischemia have also indicated the significance of
the ginsenosides in neurodegenerative diseases (7). However, research is still in its
infancy, and effective physiological dosages and the mechanisms
underlying the action of ginsenosides have not yet been fully
elucidated. Ginsenosides can be classified into two categories,
which display distinct pharmacological roles, based on the
functional groups attached at the C6 position:
20(S)-protopanaxadiol (PPD) type and 20(S)-protopanaxatriol type
(PPT) (8). In the present study,
the effect of the ginsenoside de-glycosylated metabolite PPD on
neurogenesis in rat NSC cultures was investigated. The results
suggested that the treatment of NSCs with PPD significantly
promoted the differentiation of NSCs by inducing autophagy and cell
cycle arrest.
Materials and methods
NSC cultures
NSCs were prepared as previously described (9). A total of 160 pups were used in the
present study. Postnatal day 1 Sprague-Dawley rats (specific
pathogen-free grade) were purchased from the Slack Jinda Laboratory
Animal Company (Hunan, China). The animals were maintained at
19–22°C with 40–50% humidity, 12-h light/dark cycles. Primary NSCs
derived from postnatal day 1 rats were isolated and cultured as
follows. Following sacrifice by decapitation, rat brains were
dissected in ice-cold DMEM/F12 medium (1:1; Thermo Fisher
Scientific, Inc.) supplemented with 2% penicillin and 50 µg/ml
gentamycin. The subventricular zone was treated with 1 ml Stem Pro
Accutase (Thermo Fisher Scientific, Inc.) for 10 min at 37°C.
Following digestion, the samples were filtered using a 200-mesh
sieve and centrifuged at 240 × g for 5 min at room temperature. The
obtained NSCs (5-7×106) were cultured in fresh DMEM/F12
supplemented with basic fibroblast growth factor (20 ng/ml;
PeproTech, Inc.), epidermal growth factor (20 ng/ml; PeproTech,
Inc.), 2% B27 (Thermo Fisher Scientific, Inc.), glutamine (2 mM;
Thermo Fisher Scientific, Inc.), penicillin (50 U/ml) and
gentamycin (50 µg/ml; Thermo Fisher Scientific, Inc.) at 37°C with
5% CO2. The culture medium was replaced every 2 days and
cells were subcultured every 5 days. Neurospheres were observed on
days 1, 3 and 6 using an inverted microscope (magnification, ×400;
Olympus Corporation). All procedures involving animals were
approved by the Institutional Animal Care and Use Committee of
Central South University (Changsha, China). All efforts were made
to minimize animal suffering and reduce the number of animals
used.
Cell viability and proliferation
assays
Cell viability was assessed using the Cell Counting
Kit-8 (CCK-8; Vazyme Biotech Co., Ltd.), according to the
manufacturer's protocol. Ginsenoside PPD (purity, >95%) was
obtained from the Department of Chemistry, Jilin University, and
dissolved in DMSO to make a 100-mg/l stock solution. Neurospheres
(passage 2) were digested to form single-cell suspensions. Cell
suspensions (1×105/well) were seeded into 96-well plates
in DMEM/F12 medium containing different concentrations of PPD (5,
10, 20 or 40 µM). The control group was incubated with DMEM/F12
medium containing 0.1% DMSO. Following incubation for 72 h at 37°C,
20 µl CCK-8 reagent was added to each well and incubated for a
further 4 h at 37°C. Absorbance was measured at 450 nm using a
microplate reader (BioTek Instruments, Inc.). The IC50
was calculated according to the optical density values using SPSS
software (version 18.0; IBM Corp.).
Cell proliferation was assessed by BrdU
incorporation. The single-cell suspensions (1×105
cells/well) were plated on a 20-mm coverslip (Fisherbrand; Thermo
Fisher Scientific, Inc.) in a 6-well plates in DMEM/F12 medium
containing different concentrations of PPD (5,10, 20 or 40 µM).
Cells co-cultured with DMEM/F12 medium containing 0.1% DMSO were
used as the control group. All cells were incubated with BrdU (15
µm/l; Sigma-Aldrich; Merck KGaA) for 24 h at 37°C with 5%
CO2 subsequently assessed using an immunofluorescence
assay.
Cell cycle assay
The cells were analyzed by flow cytometry (FC 500;
Beckman Coulter, Inc.) and analyzing software (CXP 2.1; Beckman
Coulter, Inc.). Cells were seeded (1×106 cells/well)
into 6-well plates and incubated without or with PPD (10, 20 and 40
µM) for 24 h at 37°C with 5% CO2. Cells were harvested
and permeabilized overnight at 4°C with pre-cooled 75% ethanol.
Subsequently, cells were treated with 1 mg/ml RNase A for 30 min at
37°C and stained with 50 µg/ml propidium iodide in the dark for 15
min at room temperature. The cell cycle was analyzed by flow
cytometry.
Cell differentiation assay
Single cell suspensions were seeded
(3.5×105 cells/well) onto 20-mm round coverslip in
6-well plates. Cells were cultured in differentiation medium
(DMEM/F12 medium without growth factors, but supplemented with 5%
FBS (Gibco; Thermo Fisher Scientific, Inc.) and incubated without
or with PPD (10, 20, 40 and 60 µM) for 72 h at 37°C with 5%
CO2. Differentiation was measured by immunostaining and
western blotting for neuronal markers tubulin-β3.
Detection of autophagy
Single cell suspensions (1×106) were
transferred into a culture bottle and cultured in differentiation
medium alone or with 20 µM PPD for 24, 48 or 72 h at 37°C with 5%
CO2. Changes in the expression of autophagy markers,
including LC3 and p62 were assessed by western blot analysis.
Following cells cultured for 72 h, LC3 punctae formation and
autophagic vacuoles were also monitored by immunofluorescence and
Transmission electron microscopy (TEM).
To further assess the role of autophagy during NSCs
differentiation, NSCs were treated with the autophagic inhibitor
wortmannin (WM, 30 µM, Abcam; cat. no. ab120148) alone or incubated
with 20 µM PPD in the differentiation medium for 72 h at 37°C with
5% CO2. Then, neuronal marker Microtubule-Associated
Protein 2 (MAP2) expression was assessed by immunofluorescence.
Immunofluorescence
Following fixation with 4% formaldehyde, pre-coated
chamber slides containing the growing cells were washed once with
PBS, permeabilized with 0.1% Triton-100 for 5 min and blocked with
0.5% BSA for 30 min. Subsequently, cells were incubated with the
following primary antibodies overnight at 4°C: Mouse monoclonal
anti-BrdU (1:100; Sigma-Aldrich; Merck KGaA; cat. no. NA61), Rabbit
monoclonal anti-tubulin-β3 (1:100; Abcam; cat. no. ab18207) and
Rabbit polyclonal anti-LC3 (1:100; Bioworld Technology, Inc.; cat.
no. AP0762). Cells were washed three times with PBS and incubated
for 1 h at 25°C with the goat anti-rabbit IgG or goat anti-mouse
IgG (1:300; Abcam; cat. nos. ab97050 and ab97022) secondary
antibodies. Cells were washed with PBS and incubated with DAPI
(1:200) for 10 min. Stained cells and LC3 punctae were observed in
five randomly selected fields of view using an Eclipse confocal
fluorescence microscope (Nikon Corporation) and analyzed using
Image J 1.46r software (National Institutes of Health).
Western blot analysis
Cells were washed twice with PBS and lysed in RIPA
buffer (50 mM Tris-HCl, 150 mM NaCl, 1.0 mM Na3VO4, 1 mM EDTA, 1%
NP-40, 0.5% sodium deoxycholate, 0.1% SDS, 100 µg/ml
phenylmethylsulfonyl fluoride, 30 µl/ml aprotinin and 4 µg/ml
leupeptin, pH 7.5). Lysates were centrifuged for 10 min at 10,000 ×
g for 10 min at 4°. The protein concentrations of the collected
supernatants were then determined by bicinchoninic acid Protein
Assay kit (Abcam; cat. no. ab102536). Proteins (50 µg per lane)
were separated via 10–15% SDS-PAGE for 1.5 h at 120 V and
transferred to PVDF membranes for 1.5 h at 40 V. The membranes
subsequently blocked with 5% skimmed milk in PBS plus 0.1% Tween-20
(PBST) for 60 min at room temperature. The membranes were incubated
overnight at 4°C with primary antibodies (1:1,000) against Rabbit
polyclonal anti-LC3 (Bioworld Technolgies, Inc.; cat. no. AP0762)),
Rabbit p62 polyclonal antibody (Bioworld Technolgies, Inc.; cat.
no. AP6006), Rabbit monoclonal anti-tubulin-β3 (Abcam; cat. no.
ab18207) and Rabbit polyclonal β-actin (Abcam; cat. no. AP0714).
LC3-I is ~14 kDa and lipidated LC3 (LC3-II) is ~16 kDa. Primary
antibodies were diluted in 1X PBST containing 5% BSA. Following
incubation, the membranes were washed with 1X PBST and incubated
for 2 h at room temperature with a goat anti-rabbit horseradish
peroxidase-conjugated IgG secondary antibody (1:2,500; Abcam; cat.
no. ab97051). Bands were developed with ECL reagent (Thermo Fisher
Scientific, Inc.; cat. no. 32209). Protein expression was
quantified using Image Lab software 4.1 (Bio-Rad Laboratories,
Inc.) with β-actin as the loading control.
TEM
At the end of incubation, cell monolayers were
washed with PBS and scraped gently with a plastic cell scraper. The
harvested cells were pelleted by centrifugation at 10,000 × g for
10 min at 37°C. Subsequently, cells were fixed with 4%
glutaraldehyde for 3 h at 4°C, post-fixed with 1% perosmic acid for
2 h at room temperature, dehydrated with acetone and embedded in an
epoxy resin. Serial ultrathin sections (70 nm) were placed on
400-mesh grids, and double-stained with 2% uranyl acetate and 0.4%
lead citrate for 15 min each at room temperature. Sections were
observed using a JEM1230 TEM (JEOL, Ltd.) at a magnification of
×8,000 in eight randomly selected fields of view. The percentage of
autophagic vacuoles area in each per field was calculated.
Statistical analysis
Statistical analyses were performed using SPSS
software (version 18.0; IBM Corp.). Data are presented as the mean
± standard deviation; all analyses were based on biological
replicates (n=3) from the same independent experiment. Comparisons
between groups were performed using one-way or two-way ANOVA
followed by Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
PPD inhibits NSC viability and
proliferation
To investigate the effect of PPD on NSC
proliferation in vitro, NSCs were treated with different
concentrations of PPD for 72 h. Cell viability was evaluated using
the CCK-8 assay. Cell viability displayed a dose-dependent
relationship with increasing concentrations of PPD (Fig. 1A); the IC50 of PPD was
calculated as 180.0±10.2 µM. The effect of PPD on cell
proliferation was further evaluated using a BrdU incorporation
assay. Following treatment with different concentrations of PPD for
24 h, the percentage of BrdU-positive cells in five randomly
selected fields of view was calculated for each group. The
percentage of BrdU-positive cells decreased significantly with
increasing concentrations of PPD compared with the percentage in
the control group (Fig. 1B and C).
The results indicated that PPD inhibited NSC proliferation in
vitro.
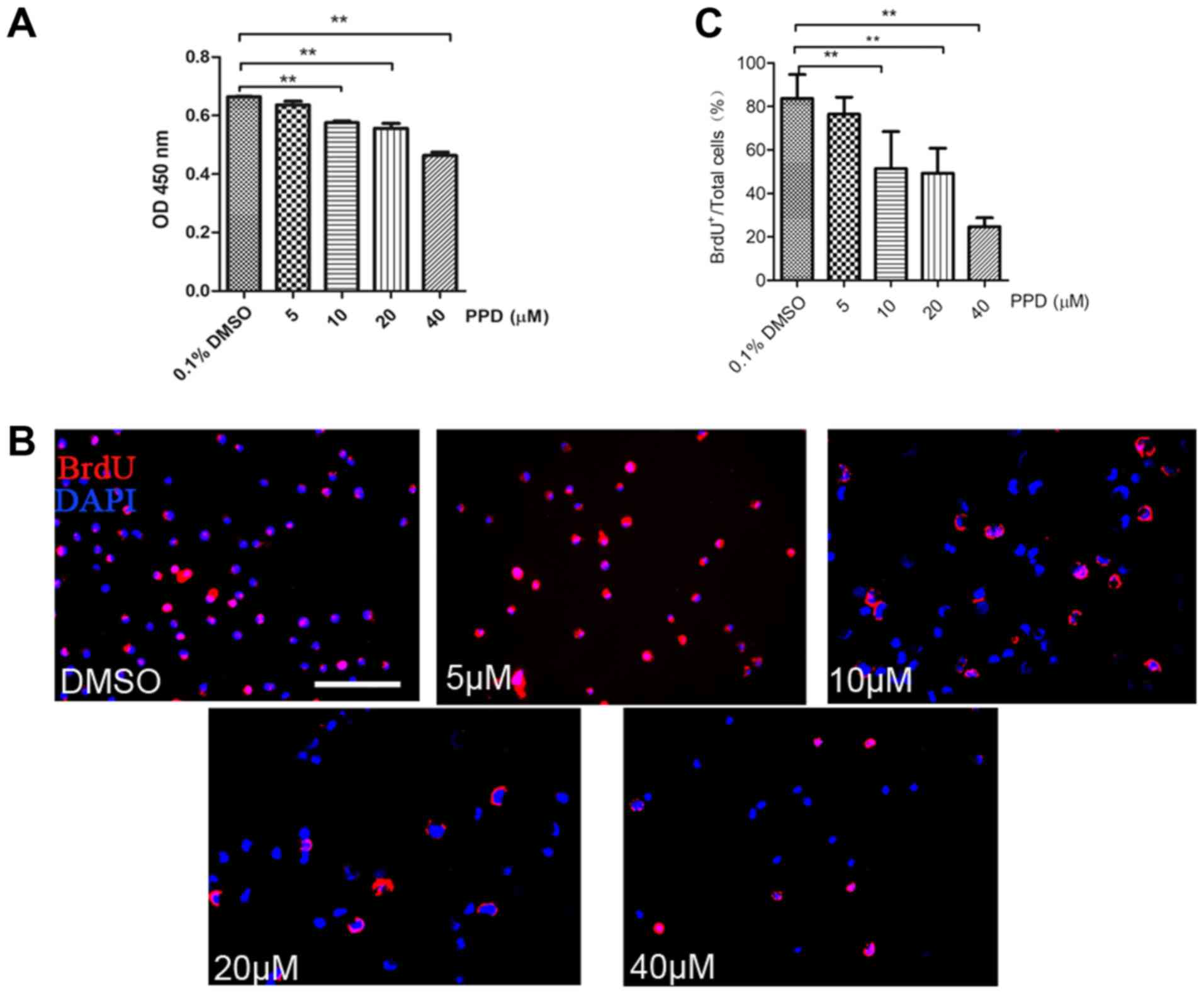 | Figure 1.PPD inhibits the viability and
proliferation of NSCs in vitro. (A) NSCs were treated with
5, 10, 20 or 40 µM PPD for 72 h, and viability was assessed using a
Cell Counting Kit-8 assay, cells incubated with DMEM/F12 medium
containing 0.1% DMSO as control group. (B) NSCs were treated with
5, 10, 20 or 40 µM PPD for 24 h, and proliferation was analysed
using a BrdU incorporation assay, cells incubated with DMEM/F12
medium containing 0.1% DMSO as control group; scale bar, 50 µm. (C)
Proportion of BrdU-positive cells following PPD treatment.
**P<0.01 vs. DMSO group. NSCs, neural stem cells; OD, optical
density; PPD, 20(S)-protopanaxadiol. |
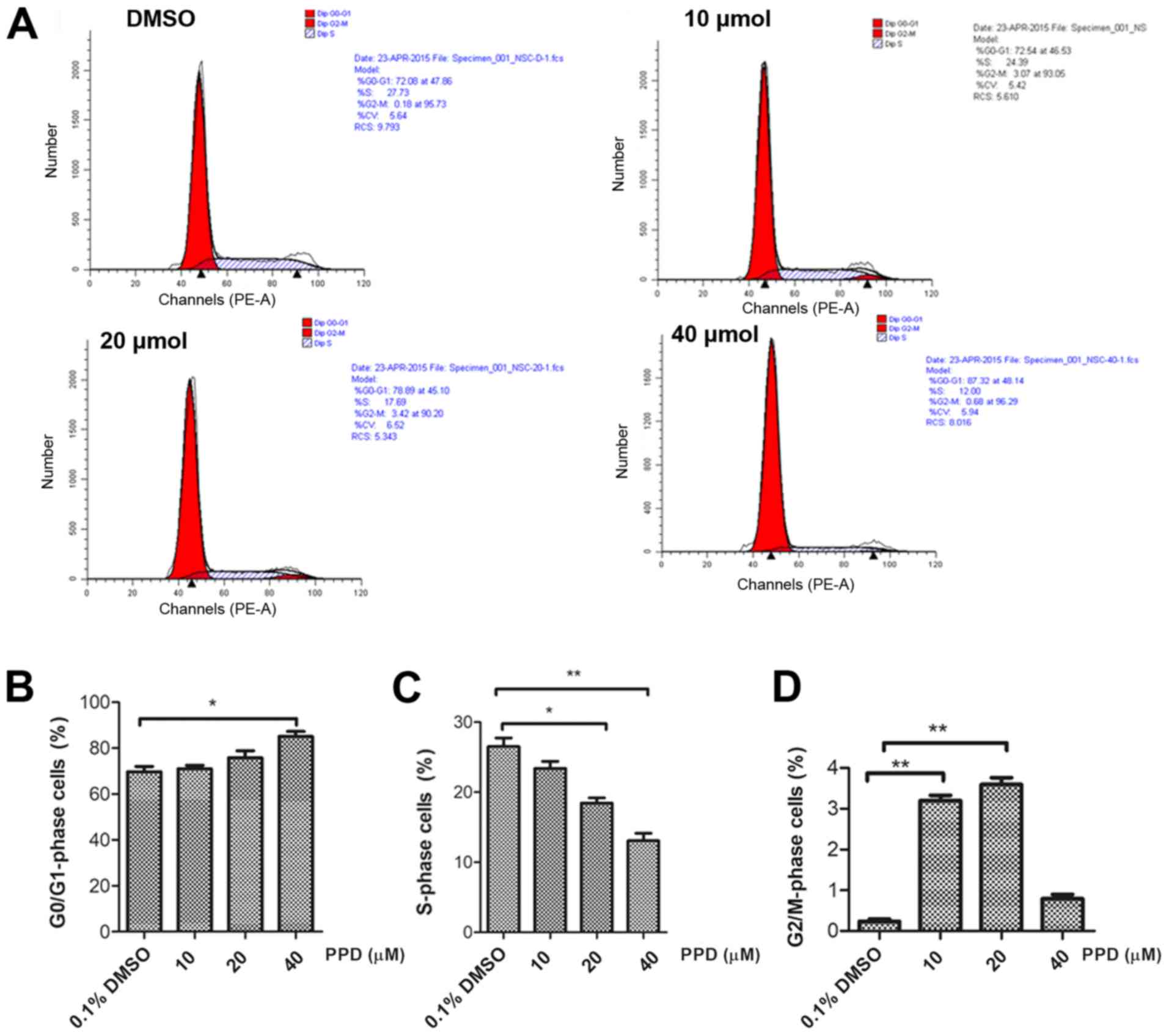
PPD induces cell cycle arrest
To further investigate the effect of PPD on NSC
proliferation, PPD-induced alterations to the cell cycle were
investigated (Fig. 2A). Flow
cytometric analysis indicated that PPD treatment increased the
proportion of cells in the G0/G1 phase in a
dose-dependent manner from 69.65% in the control group to 85.10% in
the 40 µM PPD group (Fig. 2B).
Furthermore, PPD treatment decreased the proportion of cells in the
S phase from 26.53% in the control group to 13.07% in the 40 µM PPD
group (Fig. 2C). The increased
proportion of cells in the G2/M phase in the 10 and 20
µM PPD groups was significantly higher compared with the control
group (Fig. 2D). The results
suggested that PPD inhibited NSC proliferation, which may be
associated with cell cycle arrest induction.
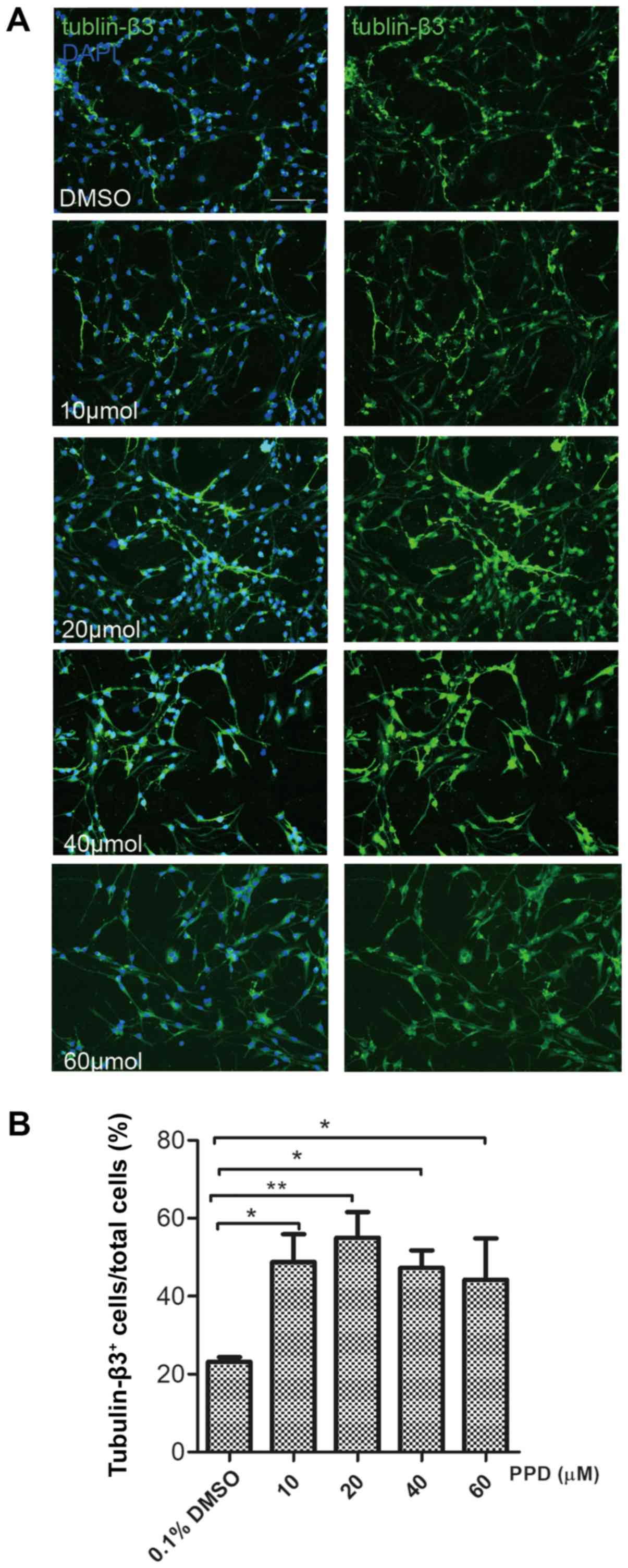
PPD promotes NSC differentiation by
inducing autophagy
To investigate the effect of PPD treatment on cell
differentiation, immunocytochemistry and western blot assays were
performed. After a 72-h culture in differentiation medium,
immunofluorescence expression of the neuronal marker tubulin-β3 was
evaluated. The 20 µM PPD group displayed the most significant
increase in tubulin-β3-positive cells compared with the control
group (P<0.05; Fig. 3A and B).
Western blotting also indicated that the expression level of
tubulin-β3 was significantly increased in NSCs following treatment
with 20 µM PPD, compared with the control group at each respective
time point (P<0.05; Fig. 4A and
B). These results indicated that PPD can effectively promote
the differentiation of neural stem cells, with the highest effect
at 20 µm. Therefore, in the subsequent experiments, 20 µm was
selected to study the mechanism of PPD promoting the
differentiation of neural stem cells.
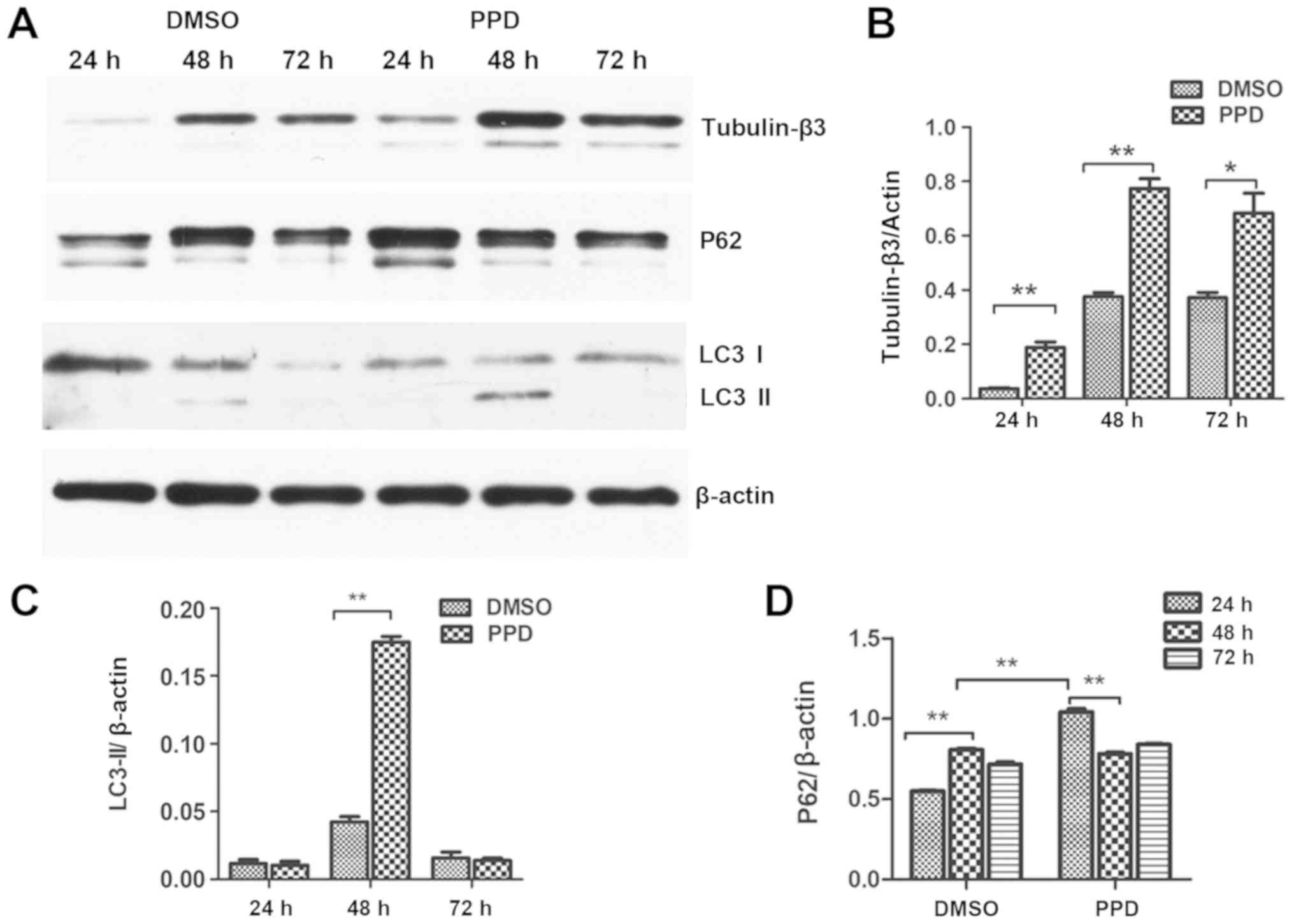
To further investigate the effect of PPD on
differentiation, NSC autophagy was assessed. LC3 is an essential
component of autophagosomes that has been widely used as a marker
of autophagy (10). Alterations in
the protein expression levels of LC3II were determined by western
blotting. It was demonstrated that 20 µM PPD treatment for 48 h
significantly increased the expression of LC3II compared with the
respective control group (P<0.01; Fig. 4A and C). p62 expression, which
often serves as another index of autophagy, increased quickly and
significantly at 24 h after PPD treatment, which was significantly
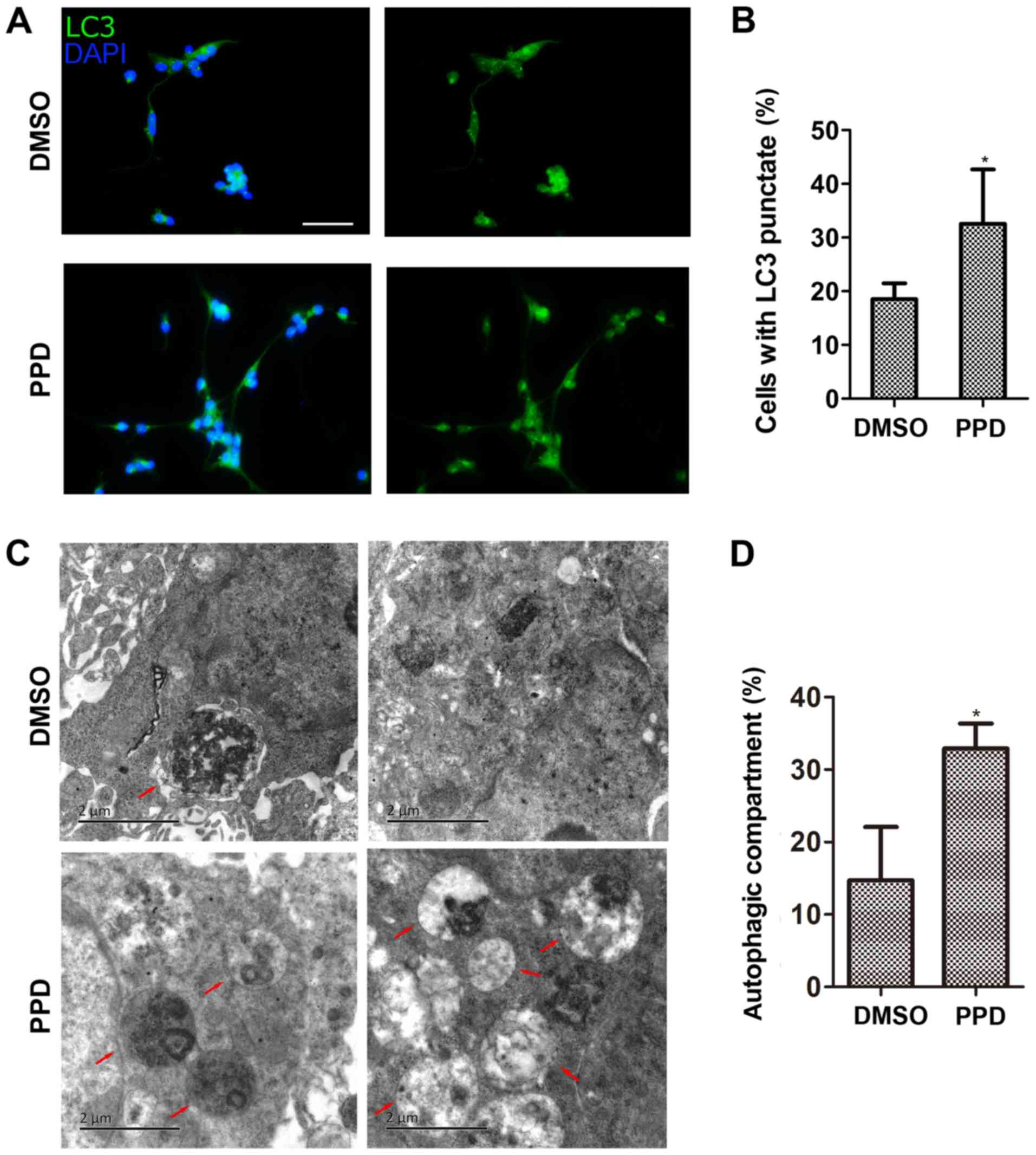
earlier than 48 h in the control group (Fig. 4A and D). LC3 punctate structures
were also detected using an immunofluorescence assay. Treatment
with 20 µM PPD significantly increased the percentage of LC3
punctae formation compared with the control group (P<0.05;
Fig. 5A and B). The TEM results
also indicated that the proportion of NSC autophagosomes was higher
by PPD treatment compared with the control group (Fig. 5C and D).
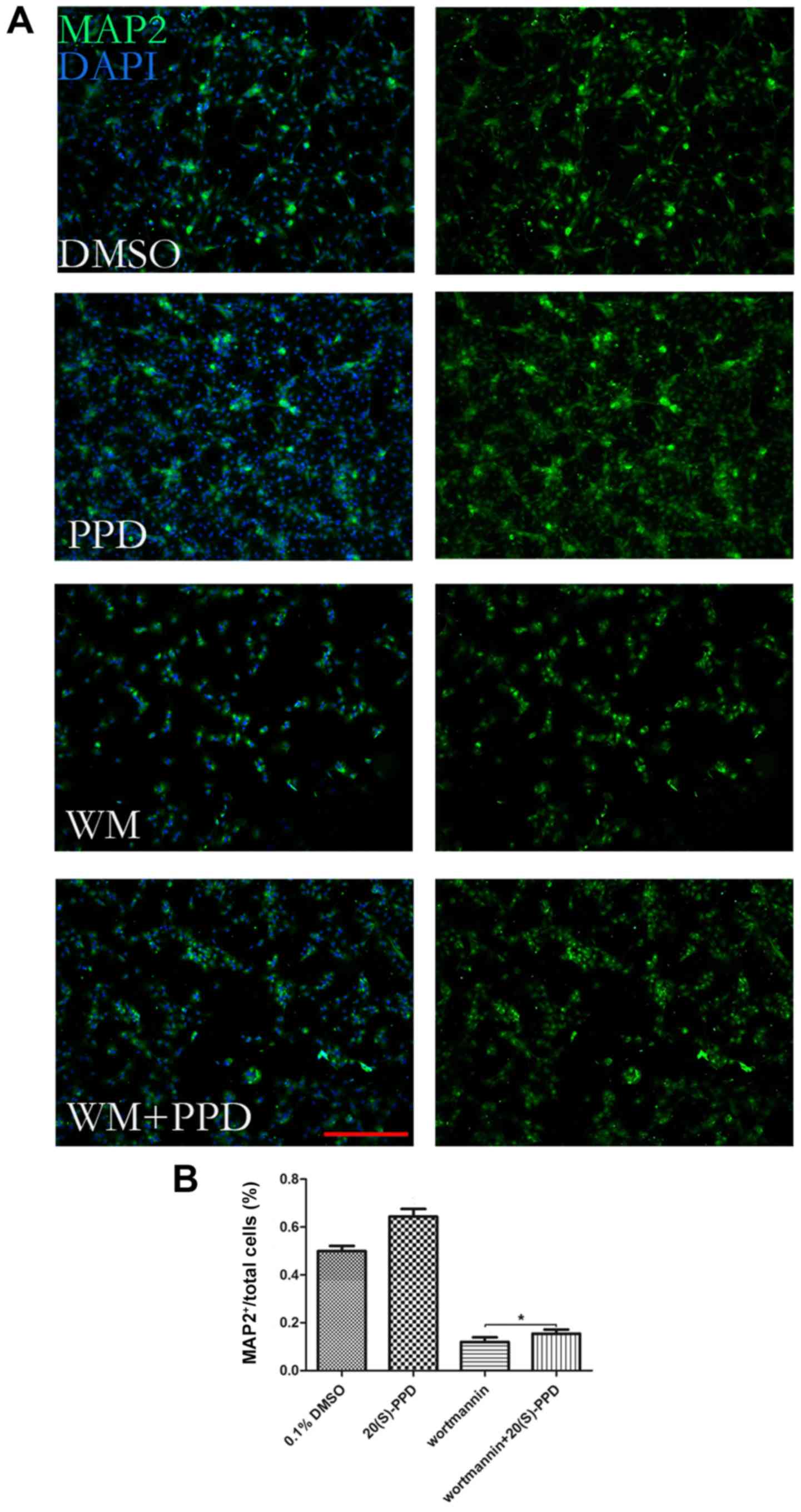
To further assess the role of autophagy during NSC
differentiation, NSCs were treated with the autophagic inhibitor
wortmannin (WM) alone or incubated with 20 µM PPD for 72 h in the
differentiation medium. The results suggested that PPD partially
rescued NSCs from WM-induced inhibitory effects on MAP2 expression,
indicating that the effect of PPD on NSC differentiation was
associated with autophagy (Fig. 6A and
B).
Discussion
Generation of new neurons involves proliferation of
progenitors, withdrawal from the cell cycle and subsequent
differentiation. Precursor cells continue to divide before
acquiring a fully differentiated state, whereas terminal
differentiation usually coincides with proliferation arrest and
permanent exit from the cell cycle. The proportion of neural
progenitors that remain in and exit from the cell cycle determines
the degree of neurogenesis (11,12).
The results from the present study suggested that treatment with
PPD reduced the proliferation of NSCs, arrested the cell cycle at
the G0/G1 and G2/M phases, and
promoted NSC differentiation into neurons in vitro.
Therefore, it was hypothesized that the therapeutic effect of PPD
on nervous system injuries and degenerative disease may occur by
promoting NSC cell cycle arrest and triggering neuronal
differentiation. The results of the present study were consistent
with a previous study, which reported that oral administration of
ginsenoside Rb1 significantly increased cell survival but not
proliferation in the hippocampus (13). Further investigation into the
molecular mechanisms linking NSC cell cycle arrest to cell
differentiation is required.
There is evidence for an active role for autophagy
during NSC differentiation. Vázquez et al (14) reported an increase in the
expression levels of the autophagy genes Autophagy Related 7,
Beclin1, activating molecule in beclin1-regulated autophagy
(Ambra1) and LC3 in the mouse embryonic olfactory bulb during the
initial period of neuronal differentiation, along with a parallel
increase in neuronal markers. Furthermore, Fimia et al
(15) revealed that Ambra1
knockout in mouse embryos leads to severe neural tube defects
associated with autophagy impairment, the accumulation of
ubiquitinated proteins, unbalanced cell proliferation and excessive
cell death. Chemical inhibitors, including 3-methyladenine and
LY294002, can reverse retinoic acid-induced neuronal
differentiation of neuroblastoma N2a cells, and RNA interference of
Beclin 1 significantly delays this process (16).
Results from the present study indicated that LC3II
expression was significantly increased following treatment with PPD
for 48 h compared with the control group. The p62 expression, which
often serves as another index of autophagy, increased quickly and
significantly at 24 h after PPD treatment, which was significantly
earlier than 48 h in the control group. Previous studies have
reported that p62 protein, via LC3, might be involved in
facilitating the clearance of polyubiquitinated protein aggregates
by linking the aggregates to the autophagic machinery (17,18).
Deterioration of the p62 promoter results in a blockade of p62
expression and can also impair the autophagic elimination of Tau
aggregates (18). Based on the
results of the present study, it was hypothesized that PPD may
accelerate the process of linking polyubiquitinated protein
aggregates to the autophagic machinery, which may also be the
mechanism of PPD inhibiting NSCs proliferation and promoting cell
differentiation. Future studies investigating the mechanisms
underlying the effects of PPD on NSC differentiation and survival
are required to verify the results of the present study.
In conclusion, the results indicated that PPD
inhibited NSC proliferation and promoted NSC differentiation,
potentially through a mechanism associated with autophagy and cell
cycle arrest. However, the present study was only preliminary and
included a number of limitations, such as the lack of in
vivo experiments and a failure to present data regarding
alterations to the expression levels of LC3II and tubulin-β3 in the
presence of the autophagy inhibitor WM. The present study may
provide a theoretical basis for the development of novel
regenerative therapeutic strategies using ginsenoside, an approved
and safe drug.
Acknowledgements
Not applicable.
Funding
The present study was supported by The National
Natural Science Foundation of China (grant nos. 81673544, 81973710
and 81903107), The Hunan Provincial Natural Science Foundation of
China (grant nos. 2016JJ4113 and 2018SK2110), The Hunan Innovation
Projects for University Students in 2016, Xiangya Hospital Central
South University Natural Science Foundation for the Youth (grant
no. 2014Q06), and The Changzhi Medical College Research Startup
Fund (grant no. QDZ201523).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
ZL, QW and JL conceived and designed the study. SC,
JH, XQ, TL, SL and AP performed the experiments and data analyses.
ZL, QW, SC and AP drafted the manuscript and figures.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Animal Care and Use Committee of Central South University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
LC3
|
light chain 3
|
|
NSCs
|
neural stem cells
|
|
PPD
|
20(S)-protopanaxadiol
|
|
PI
|
propidium iodide
|
|
TEM
|
transmission electron microscopy
|
References
|
1
|
Blin P, Dureau-Pournin C, Foubert-Samier
A, Grolleau A, Corbillon E, Jové J, Lassalle R, Robinson P,
Poutignat N, Droz-Perroteau C and Moore N: Parkinson's disease
incidence and prevalence assessment in France using the national
healthcare insurance database. Eur J Neurol. 22:464–471. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bhullar KS and Rupasinghe HP: Polyphenols:
Multipotent therapeutic agents in neurodegenerative diseases. Oxid
Med Cell Longev. 2013:8917482013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rocca WA, Petersen RC, Knopman DS, Hebert
LE, Evans DA, Hall KS, Gao S, Unverzagt FW, Langa KM, Larson EB and
White LR: Trends in the incidence and prevalence of Alzheimer's
disease, dementia, and cognitive impairment in the United States.
Alzheimers Dement. 7:80–93. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Davinelli S, Maes M, Corbi G, Zarrelli A,
Willcox DC and Scapagnini G: Dietary phytochemicals and
neuro-inflammaging: From mechanistic insights to translational
challenges. Immun Ageing. 13:162016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shin BK, Kwon SW and Park JH: Chemical
diversity of ginseng saponins from Panax ginseng. J Ginseng Res.
39:287–298. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhao J, Lu S, Yu H, Duan S and Zhao J:
Baicalin and ginsenoside Rb1 promote the proliferation and
differentiation of neural stem cells in Alzheimer's disease model
rats. Brain Res. 1678:187–194. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cheng Y, Shen LH and Zhang JT:
Anti-amnestic and anti-aging effects of ginsenoside Rg1 and Rb1 and
its mechanism of action. Acta Pharmacol Sin. 26:143–149. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wong A, Che C and Leung K: Recent advances
in ginseng as cancer therapeutics: A functional and mechanistic
overview. Nat Prod Rep. 32:256–272. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Marshall GP II, Ross HH, Suslov O, Zheng
T, Steindler DA and Laywell ED: Production of neurospheres from CNS
tissue. Methods Mol Biol. 438:135–150. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Klionsky DJ, Abdelmohsen K, Abe A, Abedin
MJ, Abeliovich H, Acevedo Arozena A, Adachi H, Adams CM, Adams PD,
Adeli K, et al: Guidelines for the use and interpretation of assays
for monitoring autophagy (3rd edition). Autophagy.
12:1–222. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ruijtenberg S and van den Heuvel S:
Coordinating cell proliferation and differentiation: Antagonism
between cell cycle regulators and cell type-specific gene
expression. Cell Cycle. 15:196–212. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang RL, Zhang ZG, Roberts C, LeTourneau
Y, Lu M, Zhang L, Wang Y and Chopp M: Lengthening the G(1) phase of
neural progenitor cells is concurrent with an increase of symmetric
neuron generating division after stroke. J Cereb Blood Flow Metab.
28:602–611. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu L, Hoang-Gia T, Wu H, Lee MR, Gu L,
Wang C, Yun BS, Wang Q, Ye S and Sung CK: Ginsenoside Rb1 improves
spatial learning and memory by regulation of cell genesis in the
hippocampal subregions of rats. Brain Res. 1382:147–154. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vázquez P, Arroba AI, Cecconi F, de la
Rosa EJ, Boya P and de Pablo F: Atg5 and Ambra1 differentially
modulate neurogenesis in neural stem cells. Autophagy. 8:187–199.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fimia GM, Stoykova A, Romagnoli A, Giunta
L, Di Bartolomeo S, Nardacci R, Corazzari M, Fuoco C, Ucar A,
Schwartz P, et al: Ambra1 regulates autophagy and development of
the nervous system. Nature. 447:1121–1125. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zeng M and Zhou JN: Roles of autophagy and
mTOR signaling in neuronal differentiation of mouse neuroblastoma
cells. Cell Signal. 20:659–665. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shvets E, Fass E, Scherz-Shouval R and
Elazar Z: The N-terminus and Phe52 residue of LC3 recruit
p62/SQSTM1 into autophagosomes. J Cell Sci. 121:2685–2695. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pankiv S, Clausen TH, Lamark T, Brech A,
Bruun JA, Outzen H, Øvervatn A, Bjørkøy G and Johansen T:
p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of
ubiquitinated protein aggregates by autophagy. J Biol Chem.
282:24131–24145. 2007. View Article : Google Scholar : PubMed/NCBI
|