Introduction
Cytoskeletal rearrangement is required for cell
migration and invasion, which are also important steps for the
process of cancer metastasis (1,2).
Dynein transport along the cytoskeletal microtubules towards the
minus end is essential for cell division, migration and
cytoskeletal motors (3–5).
The cytoplasmic dynein complex is implicated in
intracellular vesicular transport (6). Numerous studies have reported that
microtubule-dependent vesicular trafficking serves a central role
in cell function and may impair tumor invasion (7–9). The
expression levels of cytoplasmic dynein, particularly dynein light
chains, have been associated with several types of cancer in
humans; for example, melanoma cell apoptosis was associated with
dynein light chain LC8-type (DYNLL)1 and DYNLL2 expression
(10); and upregulation of dynein
light chain roadblock-type (DYNLRB)1 and downregulation of DYNLRB2
expression levels were detected in hepatocellular carcinoma (HCC)
(11). Moreover, in another study
investigating HCC, aberrant expression levels of dynein cytoplasmic
1 light intermediate chain 1 (DYNC1LI1) were observed (12). DYNC1LI1 expression has also been
detected in the urine of patients with pancreatic ductal
adenocarcinoma (13), and
increased DYNC1LI1 phosphorylation was also found in prostate
cancer (CaP) (14). Altogether,
these findings suggested that DYNC1LI1 may serve a potential
tumorigenic role in numerous types of human cancer.
The epithelia in organ systems, such as the urinary
and gastrointestinal systems, are protected from pathogenic
infections by a mucosal barrier (15,16).
Mucin (MUC) expression is critical for the formation of the mucosal
barrier (17). Two functionally
distinct classes of MUCs, secretory gel-forming MUCs and
transmembrane MUCs, have been identified in different epithelial
cells (18,19). Both MUCs have been discovered to be
intimately involved in the inflammatory response and tumorigenesis
(19). In addition, the
cytoskeletal structures have also been found to be involved in the
regulation of the mucosal barrier integrity and its breakdown
during inflammation (20).
Therefore, molecular detection methods that can improve our
understanding of the mucosal barrier are critical to both cancer
management and treatment. In the present study, the expression
levels of DYNC1LI1 were investigated in colorectal cancer (CRC)
tissues and in the human epithelial colon cell line LS 174T, which
is considered by the American Joint Committee on Cancer (AJCC) to
be a model of stage II CRC and to express different MUCs (21). A series of reverse
transcription-quantitative PCR (RT-qPCR) experiments were performed
to determine the association between the expression levels of
DYNC1LI1 and different MUCs in LS 174T cells.
Materials and methods
Cell lines and reagents
The goblet-cell-like CRC cell line, LS 174T (AJCC
stage II; CL-188) and two other CRC stage II cell lines, SW480
(CCL-228) and HCT 116 (CCL-247), were purchased from the American
Type Culture Collection. The clinical status of the cell lines and
the culture conditions are available on the ATCC website
(www.atcc.org). Briefly, LS 174T cells were
cultured in minimum essential medium (cat. no. 41500-034; Thermo
Fisher Scientific, Inc.), HCT 116 cells were cultured in DMEM (cat.
no. 12800-017; Thermo Fisher Scientific, Inc.) and SW480 cells were
cultured in L-15 medium (cat. no. 41300-039; Thermo Fisher
Scientific, Inc.). All the culture mediums were supplemented with
10% fetal calf serum (cat. no. A6806-35; NQBB International
Biological Corp.) and 1% antibiotic/antimycotic solution (cat. no.
15240-062; Thermo Fisher Scientific, Inc.). LS 174T cells and HCT
116 cells were maintained in a humidified chamber with 95% air and
5% CO2. SW480 cells were maintained in a humidified
incubator with 100% air at 37°C. The Human Reference cDNA (HRC;
cat. no. 636692; Takara Bio, Inc.) was used as an expression
control for DYNC1LI1.
Lentiviral knockdown of DYNC1LI1
The lentiviral construct, pLKO_TRC005-DYNC1LI1
(clone ID: TRCN0000299843), with short hairpin RNA (shRNA) to
target DYNC1LI1 (shDYNC1LI1), was used to knockdown the expression
level of DYNC1LI1. The control pLKO_TRC005-luciferase (LUC; clone
ID: TRCN0000231719) vector targeting LUC was used as the negative
control (shLUC-LS 174T) for the shDYNC1LI1. Both the lentiviral
constructs were obtained from the RNA Technology Platform and Gene
Manipulation Core (Sinica, Taiwan). A total of 1.25×105
LS 174T cells/well were cultured in a 6-well plate for 24 h and the
subsequent lentiviral infections (multiplicity of infection=3) were
performed to knock down the expression of DYNC1LI1 in LS 174T cells
(shDYNC1LI1-LS 174T). Subsequently, medium containing 2 mg/ml
puromycin (Thermo Fisher Scientific, Inc.) was used to select for
and maintain stable clones. Following incubation for 48 h, the
infection efficiency of DYNC1LI1 knockdown in LS 174T cells
compared with the shLUC-LS 174T cells was determined using RT-qPCR
and western blotting.
Western blotting
Total protein was extracted from shLUC-LS 174T and
shDYNC1LI1-LS174T cells using the PRO-PREP™ Protein Extraction
solution (Intron Biotechnology, Inc.) according to the
manufacturer's protocol, supplemented with a protease inhibitor
(cat. no. P8340; Merck KGaA, Inc.). Total protein was quantified
using Bio-Rad Protein Assay reagent (cat. no. 500-0006; Bio-Rad
Laboratories, Inc.) and 15 µg protein/lane was denatured in 1X
NuPAGE™ LDS sample buffer (Thermo Fisher Scientific, Inc.) for 10
min at 95°C, and separated via 12% SDS-PAGE. The separated proteins
were subsequently transferred onto a PolyScreen 2 PVDF Transfer
membrane (0.2 µm; PerkinElmer, Inc.). The membranes were blocked
with 3% bovine serum albumin (cat. no. ALB001.100; BioShop Canada
Inc.) for 1 h at room temperature and incubated with the following
primary antibodies for 1 h at room temperature: Anti-DYNC1LI1
(1:1,000; cat. no. ab154251; Abcam) and anti-β-actin (1:1,000; cat.
no. MAB1501R; Merck KGaA). Following the primary antibody
incubation, the membranes were washed with 1X TBST wash buffer (20
mM Tris, 0.15 M NaCl, 0.1% Tween-20; pH 7.4) three times (15
min/time) at room temperature. Subsequently, the membranes were
incubated with an alkaline phosphatase-conjugated anti-rabbit
antibody (1:5,000; cat. no. A3812; Merck KGaA) or alkaline
phosphatase-conjugated anti-mouse antibody (1:5,000; cat. no.
A9316; Merck KGaA) for 60 min at room temperature. Protein bands
were visualized using a VECTASTAIN® ABC-AmP DuoLuX
chemiluminescent/fluorescent substrate kit (cat. no.AK-6000; Vector
Laboratories, Inc.), according to the manufacturers' protocol, and
a FluorChem FC2 system (Cell Biosciences, Inc.).
CRC cDNA array and RT-PCR
DYNC1LI1 expression levels were quantified using a
cDNA array of colonic tissues covering four CRC stages (n=40) and
normal colonic tissues (n=8; cat. no. HCRT104; OriGene
Technologies, Inc.). Total RNA from the cultured cells was
extracted using the Easy Pure Total RNA Mini kit (Bioman Scientific
Co. Ltd.), according to the manufacturer's protocol. Subsequently,
total RNA (1 g) was reverse transcribed into single-stranded cDNA
using an oligo(dT)12 primer with a High Capacity cDNA Reverse
Transcriptase kit (cat. no. 4368813; Thermo Fisher Scientific,
Inc.), according to the manufacturer's protocol. DYNC1LI1 and MUC2
mRNA expression levels were subsequently quantified by qPCR using
the TaqMan Master mix (Roche Diagnostics GmbH). The following
thermocycling conditions were used for qPCR: Initial denaturation
at 95°C for 10 min; followed by 60 cycles of 95°C for 10 sec and
60°C for 20 sec (22). The primer
pairs and universal TaqMan probes used for the qPCR are presented
in Table I. Expression levels of
MUC1, MUC4, MUC5AC and MUC6 were determined as previously described
by Ohuchida et al (23).
Expression levels were quantified using the 2−∆∆Cq
method and normalized to the internal reference gene GAPDH.
 | Table I.Primers and universal TaqMan probes
used for the reverse transcription-quantitative PCR. |
Table I.
Primers and universal TaqMan probes
used for the reverse transcription-quantitative PCR.
| Gene
numbera | NCBI reference
sequence | Primer sequence
(5′→3′) | Probe |
|---|
| Dynein cytoplasmic
1 light intermediate chain 1 | NM_016141 | F:
CTGGTGTGAGTGGTGGTAGC | 10 |
|
|
| R:
TCTGCATGAACATCTAAGACAGG |
|
| Mucin 2 | NM_002457 | F:
ATGCCAGCATTTGCATCC | 89 |
|
|
| R:
GGCACCCTGGTCTCATTG |
|
| GAPDH | NM_002046 | F:
CTCTGCTCCTCCTGTTCGAC | 60 |
|
|
| R:
ACGACCAAATCCGTTGACTC |
|
Determining the chemosensitivity and
proliferative rate of shLUC-LS 174T and shDYNC1LI1-LS 174T
cells
The chemosensitivity and proliferative rate of
shLUC-LS 174T and shDYNC1LI1-LS 174T cells were determined using a
MTT assay (cat. no. M5655; Sigma-Aldrich; Merck KGaA). Briefly,
3×103 shLUC-LS 174T or shDYNC1LI1-LS 174T cells/well
were cultured in 96-well plates for overnight at 37°C before drug
treatments. The chemosensitivity of cells to the DNA analog
5-fluorouracil (5-FU; Merck KGaA) or the platinum-based drug
oxaliplatin (Merck KGaA) at various concentrations (0.01 µM to 37.5
mM for 5-FU and 2 nM to 1,000 µM for oxaliplatin) was plotted after
72 h of incubation at 37°C and the half-maximal inhibitory
concentration (IC50) for each compound was calculated
using Gen5™ Data Analysis (version 2.04; BioTek Instruments,
Inc.).
To determine the cell proliferative rate,
5×103 shLUC-LS 174T and shDYNC1LI1-LS174T cells/well
were plated into 96-well plates and incubated alone, or with the
IC50 dose of 5-FU or oxaliplatin for the 1–7 days at
37°C.
For both assays, the cells were treated with 10 µl
MTT reagent in the dark for 4 h at room temperature and then 100 µl
DMSO was added/well to dissolve the purple formazan formed by the
live cells. The absorbance at 540 nm of each well was measured
using a Synergy HT Multi-Mode microplate reader (BioTek
Instruments, Inc.). Data were obtained from three independent
experiments.
Statistical analysis
Statistical analysis was performed using SPSS
version 15.0 software (SPSS, Inc.). Statistical differences between
2 groups were determined using a Student's t-test, whereas
statistical differences between >2 groups were determined using
a one-way ANOVA, followed by a Bonferroni's post hoc test for
multiple comparisons. Data was presented as the mean ± SD. P≤0.05
was considered to indicate a statistically significant
difference.
Results
DYNC1LI1 expression levels in CRC
tissues
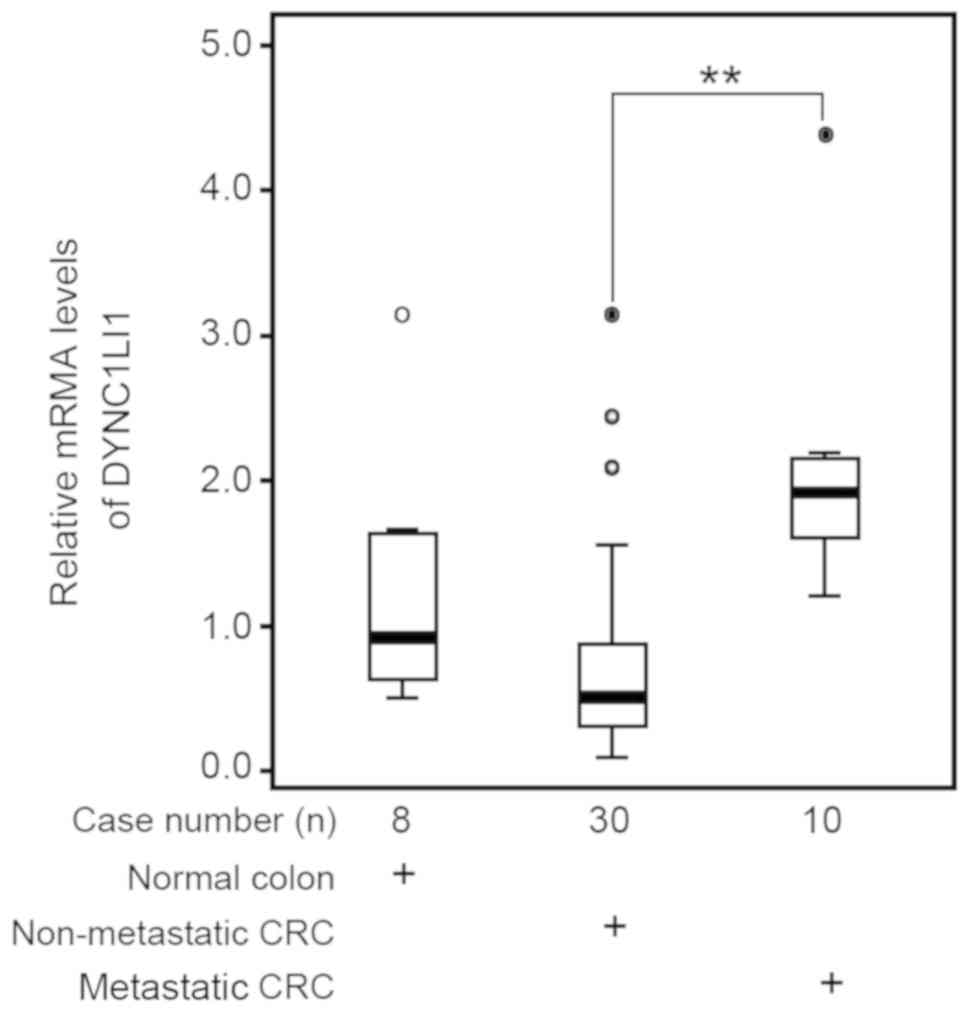
The differential expression levels of DYNC1LI1 in
CRC were analyzed in cDNA arrays of colonic tissues containing
normal colonic tissues and four CRC stages. DYNC1LI1 expression
levels were significantly increased in cDNA samples from patients
with metastasis (n=10) compared with the patients without
metastasis (n=30; P<0.01; Fig.
1).
Differential MUC expression levels in
DYNC1LI1 knockdown LS 174T cells
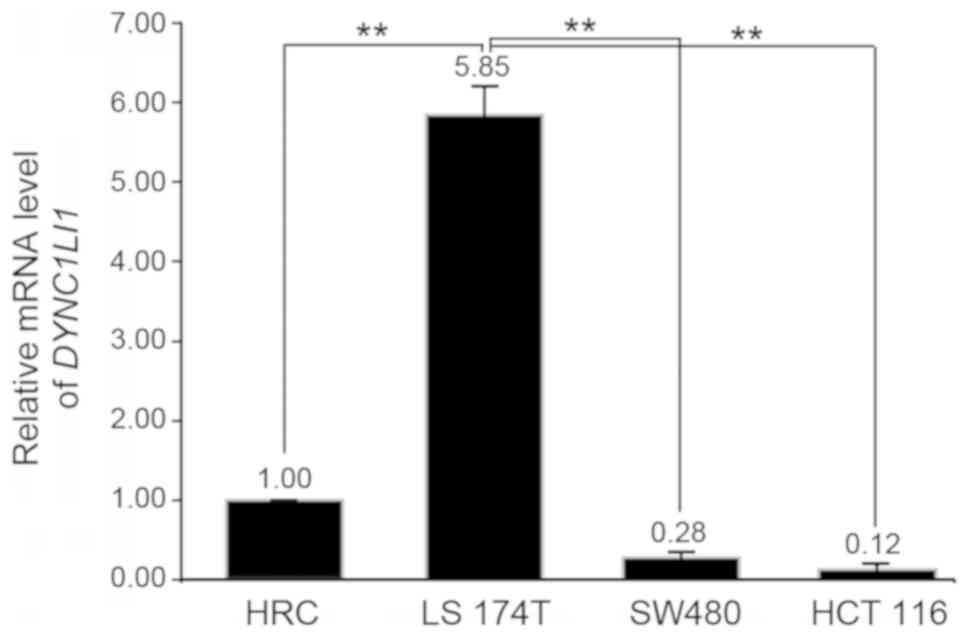
DYNC1LI1 mRNA expression levels were quantified in
three CRC cell lines: HCT 116, SW480 and LS174T. Among these CRC
cell lines (AJCC stage II), LS 174T cells had significantly
increased expression levels of DYNC1LI1 compared with the HRC
control and other CRC cell lines (P<0.01; Fig. 2); therefore, LS 174T cells were
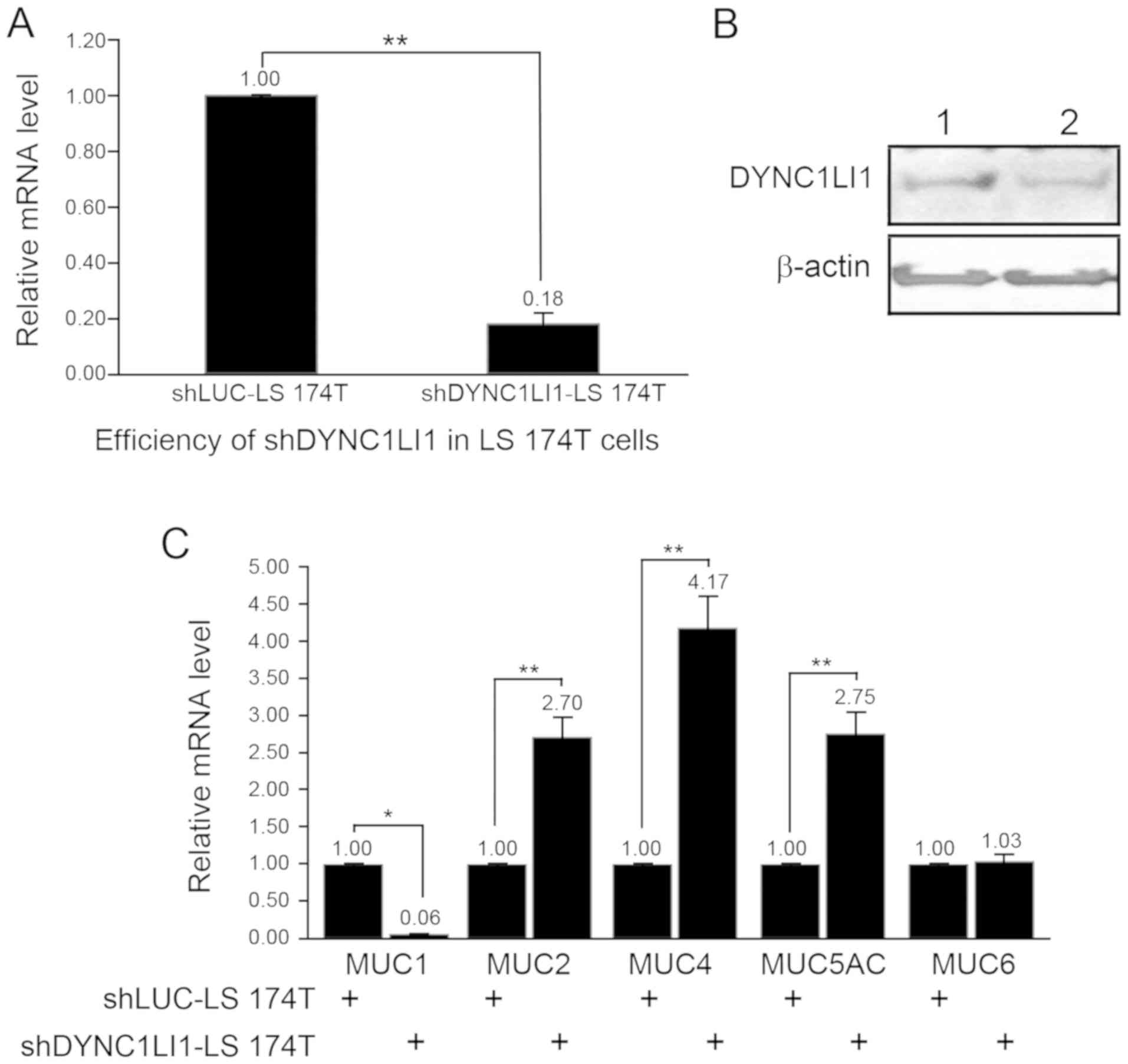
used in subsequent experiments. The knockdown efficiency of
shDYNC1LI1 in LS 174T cells was subsequently determined using
RT-qPCR and western blotting, and it was established to be
successful (Fig. 3A and B).
Notably, MUC mRNA expression levels were discovered to be
differentially altered in the shDYNC1LI1-LS 174T cells (Fig. 3C). Briefly, MUC1 expression levels
were significantly decreased by 0.06-fold, whereas the expression
levels of MUC2, MUC4 and MUC5AC were all significantly increased by
2.70, 4.17 or 2.75-fold, respectively, in shDYNC1LI1-LS 174T cells
compared with the shLUC-LS 174T cells (Fig. 3C). No significant differences were
observed in the expression levels of MUC6 between the shDYNC1LI1-LS
174T and shLUC-LS 174T cells.
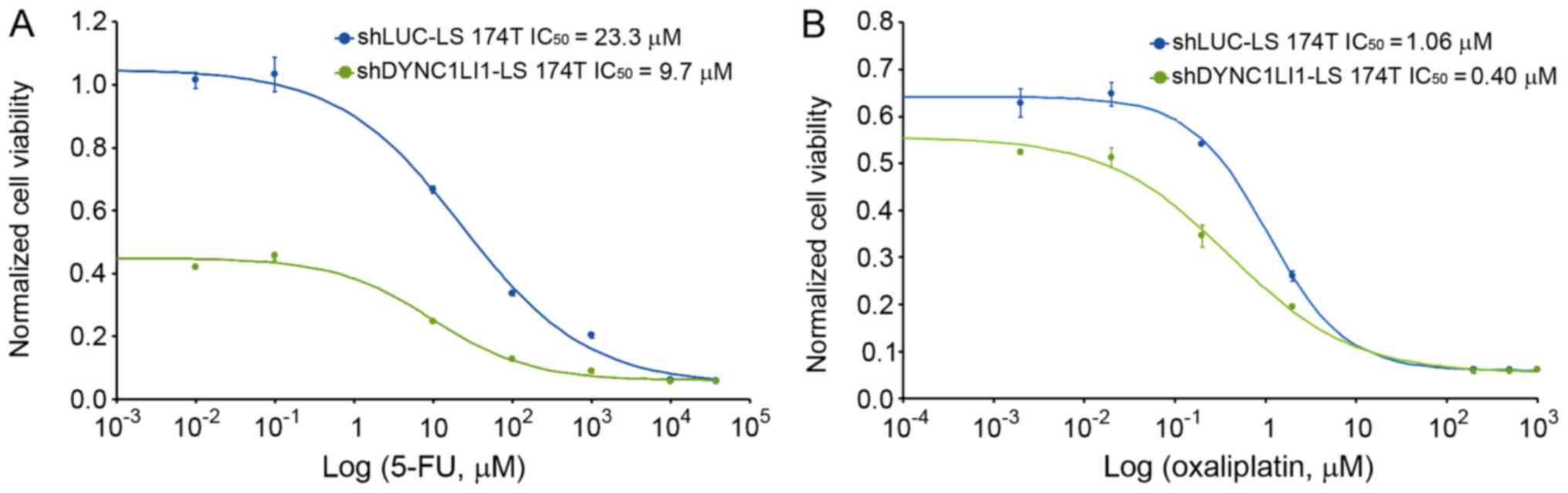
Chemotherapeutic effects in shLUC-LS
174T and shDYNC1LI1-LS 174T cells
5-FU, leucovorin, and oxaliplatin (FOLFOX4) is a
chemotherapeutic reagent used for the treatment of CRC (24,25).
It was discovered that the genetic knockdown of DYNC1LI1 in LS 174T
cells increased the susceptibility of the cells to 5-FU and
oxaliplatin treatment; the IC50 of 5-FU (9.7 µM) and
oxaliplatin (0.4 µM) was significantly decreased in the
shDYNC1LI1-LS 174T cells compared with the shLUC-LS 174T cells
(23.3 µM and 1.06 µM, respectively; Fig. 4A and B). The differences in the
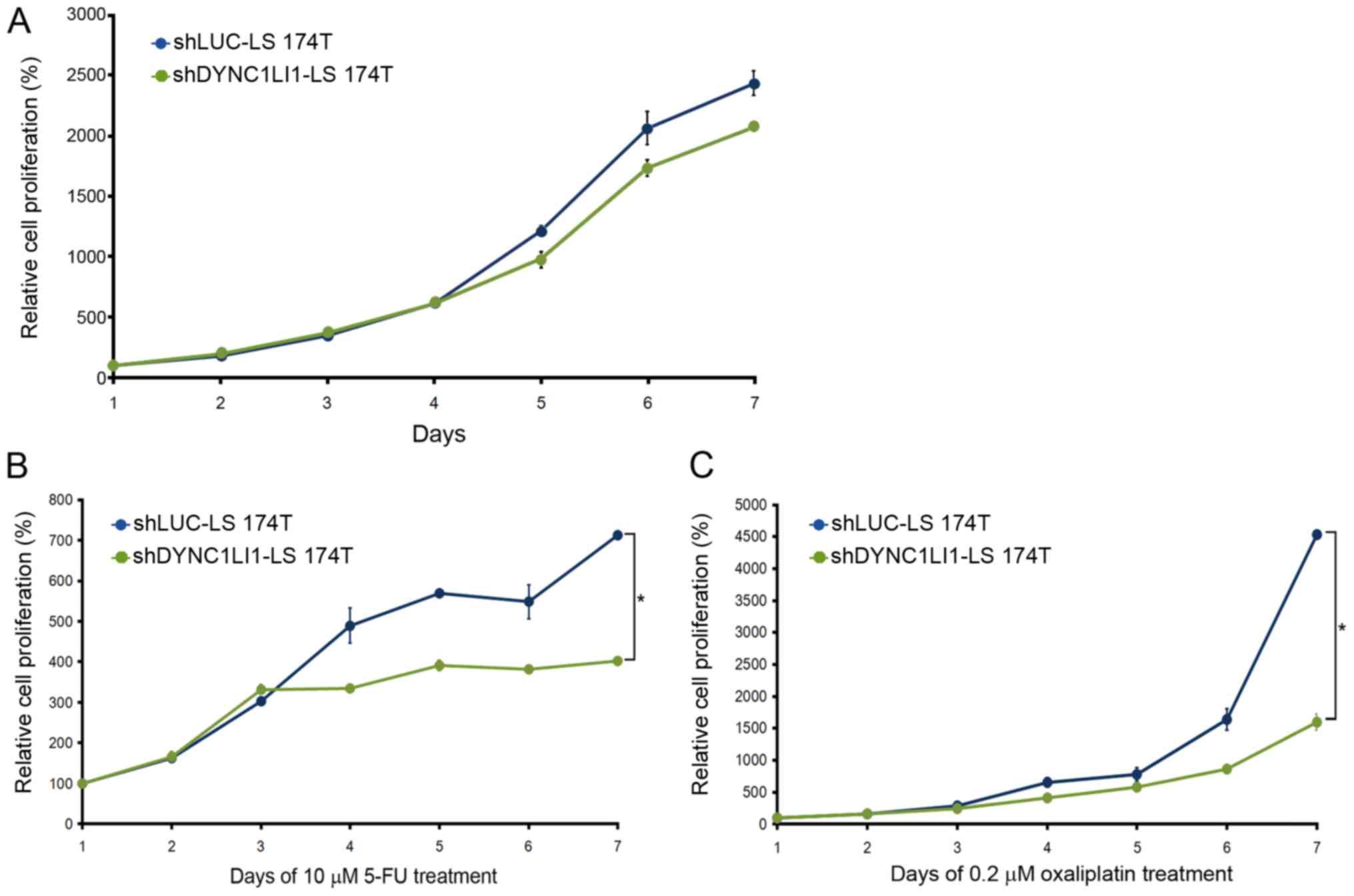
cell proliferative rate of shLUC-LS 174T and shDYNC1LI1-LS 174T
cells began at day 4 without any drug treatment (Fig. 5A), and at day 3 with 5-FU (Fig. 5B) or oxaliplatin chemotherapy
(Fig. 5C). Moreover, the
shDYNC1LI1-LS 174T cells exhibited a significantly reduced
proliferative rate at day 7 compared with the shLUC-LS 174T cells
following both 5-FU or oxaliplatin chemotherapy (P<0.05;
Fig. 5B and C).
Discussion
Tumorigenesis involves cytoskeletal reorganization
(26) and as demonstrated by van
Ree et al (27),
cytoplasmic dyneins can regulate centrosome dynamics. Thus, any
changes in their function have been hypothesized to lead to cancer
formation. For example, it was previously reported that in patients
with advanced CRC, the expression levels of the cytoplasmic dynein
1 heavy chain 1 (DYNC1H1) were decreased (28). This aberrant expression of DYNC1H1
was suggested to impede the assembly of complete cytoplasmic
dynein, resulting in inadequate microtubule dynamics.
In the present study, increased expression levels of
DYNC1LI1 were observed in late-stage, metastatic CRC tissues.
Therefore, the tumorigenic potential of DYNC1LI1 in CRC warrants
further metastatic investigations and research. The
goblet-cell-like LS 174T cells, which exhibit mucinous secretory
granules, have previously provided an excellent in vitro
model for studying the expression levels of MUCs in CRC (21). The present study revealed that
genetic knockdown of DYNC1LI1 induced decreased expression levels
of MUC1 and increased expression levels of MUC2, MUC4 and MUC5AC in
LS 174T cells. The differentially expressed levels of DYNC1LI1 in
LS 174T cells have been suggested to change the MUC profile,
thereby generating differences in the mucosal barrier (29). Thus, it can be further hypothesized
that DYNC1LI1-induced changes in the mucosal barrier may be
positively associated with tumor formation. The integrity of the
gut mucosal barrier, which is composed of a specific profile of
MUCs, has been found to be crucial for protecting epithelial cells
from inflammatory or cancerous responses (30,31).
A previous study provided molecular evidence to
indicate that the changes in the expression levels of
cytoskeleton-related genes may worsen the prognosis of patients
with CRC and lead to the formation of an aberrant mucosal barrier
with a different MUC profile (19). A previous study indicated that
several MUCs, such as increased MUC1 and decreased MUC2, are
associated with CRC (18). Based
on the results of the present study and the aforementioned studies,
it was hypothesized that increased DYNC1LI1 expression levels may
be associated with the advanced stages of CRC. In LS 174T cells,
the expression levels of MUC1, a transmembrane glycosylated
phosphoprotein, and DYNC1LI1 displayed a similar expression trend
in the present study; this result is consistent with that observed
in the CRC cell lines, HCT 116 and SW480. A previous study
indicated that cells with lower DYNC1LI1 expression levels also had
decreased expression levels of MUC1 (32,33).
MUC1, a membrane-bound MUC, was also found to be overexpressed in
numerous types of human cancer, such as endometrioid endometrial
carcinoma and CaP (34,35). Moreover, patients with increased
MUC1 expression levels were suggested to have a poorer prognosis
(36). Betge et al
(17) concluded that MUC1
expression levels had no effect on the clinical outcomes, although
more than half of the studied patients with CRC exhibited MUC1
overexpression. Other previous studies investigating the prognostic
value of MUCs in cancers have also reported contradictory results
(37–39). These discrepancies in the results
of different studies may be attributable to the differences in
cytoskeletal gene expression. Thus, DYNC1LI1 expression levels, and
its effects, must be further understood in order to predict patient
prognosis.
Secreted gel-forming MUCs, another family of MUCs,
include both MUC2 and MUC5AC (18). Betge et al (17) reported that the loss of MUC2 and
MUC5AC expression was associated with favorable CRC outcomes;
however, only reduced expression levels of MUC2, and not of MUC5AC,
were detected in colonic tumors by Byrd and Bresalier (18). Previously, MUC2 was suggested to be
a crucial component of the MUC profile in the mucosal barrier, but
served no role in carcinogenesis (40). In addition, an animal model
indicated that Muc2-knockout mice had a poorly defined mucosal
barrier (16). Collectively, these
results indicated that DYNC1LI1, which upregulates the expression
levels of transmembrane MUCs, such as MUC1, and downregulates the
expression levels of secreted gel-forming MUCs, such as MUC2 and
MUC5AC, may be critical for assessing the composition of the
mucosal barrier in patients with CRC. Compared with normal prostate
or benign tissue, CaP tissue exhibited a similar MUC profile, with
increased MUC1 and decreased MUC2, MUC4, MUC5AC and MUC6 expression
levels (34). In addition, the
difference of immunohistochemical staining patterns of MUC1 and
MUC2 were caused by the carcinomatous transformation of urothelial
neoplastic and preneoplastic lesions (41). Taken together, these findings
suggested that different types of cancer, including those of the
urological and gastrointestinal systems, exhibit diverse MUC
profiles.
However, the possibility that disrupting the
microtubule dynamics in shDYNC1LI1-LS 174T cells may influence cell
proliferation cannot be excluded. For example, Even et al
(42) reported that DYNC1LI1
contributed to the proliferative overgrowth of CRC cells, which is
consistent with the present study results, which found that
shDYNC1LI1-LS 174T cells had a slower proliferative rate compared
with shLUC-LS 174T cells without any drug treatment. Adjuvant
chemotherapy, such as 5-FU or oxaliplatin, which are the
ingredients of FOLFOX4, was discovered to further inhibit the
proliferation of LS 174T cells with low DYNC1LI1 expression levels.
Thus, these findings suggested that DYNC1LI1 may be a marker
molecule for prescribing 5-FU as a first-line chemotherapy
treatment for patients with CRC. In our in vitro cell study,
shDYNC1LI1-LS 174T cells were more sensitive to 5-FU treatment
compared with cells with endogenous DYNC1LI1 expression levels. As
previously discussed, shDYNC1LI1-LS 174T cells had low MUC1, and
high MUC2, MUC4 and MUC5AC expression levels, implying that CRC
cells with low MUC1 expression levels and high expression levels of
other MUCs may benefit from 5-FU-based therapy. Furthermore, this
result was similar to findings in pancreatic cancer cells reported
by Kalra and Campbell (43), where
it was demonstrated that increased MUC1 expression levels
potentially impeded the cytotoxic effects of 5-FU against the
proliferation of pancreatic cancer cells.
In conclusion, based on the findings of the present
results and those of other previous studies, it was suggested that
DYNC1LI1, which is significantly associated with MUC expression
levels, may affect the efficiency of chemotherapy. Additional
functional studies and clinical reports are required for
understanding its molecular significance in tumorigenesis.
Acknowledgements
Not applicable.
Funding
The current study was supported by grants from the
Cathay General Hospital and Taipei Medical University (grant nos.
104CGH-TMU-05 and 105CGH-TMU-04) and the Cathay General Hospital
(grant no. CGH-MR-A10316).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
CCC and KCC performed the experiments and wrote the
manuscript. CJH performed the statistical analysis and helped
revise the manuscript. CSH contributed to the interpretation of the
data. YCW conceived and designed the study. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
DYNC1LI1
|
dynein cytoplasmic 1 light
intermediate chain 1
|
|
CRC
|
colorectal cancer
|
|
MUC
|
mucin
|
|
RT-qPCR
|
reverse transcription-quantitative
PCR
|
|
5-FU
|
5-fluorouracil
|
|
shRNA
|
short hairpin RNA
|
References
|
1
|
Singh R, Kapur N, Mir H, Singh N, Lillard
JW Jr and Singh S: CXCR6-CXCL16 axis promotes prostate cancer by
mediating cytoskeleton rearrangement via Ezrin activation and
alphavbeta3 integrin clustering. Oncotarget. 7:7343–7353. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wu K, Zhang X, Li F, Xiao D, Hou Y, Zhu S,
Liu D, Ye X, Ye M, Yang J, et al: Frequent alterations in
cytoskeleton remodelling genes in primary and metastatic lung
adenocarcinomas. Nat Commun. 6:101312015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kuijpers M, van de Willige D, Freal A,
Chazeau A, Franker MA, Hofenk J, Rodrigues RJ, Kapitein LC,
Akhmanova A, Jaarsma D and Hoogenraad CC: Dynein regulator NDEL1
controls polarized cargo transport at the axon initial segment.
Neuron. 89:461–471. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
McKenney RJ, Huynh W, Tanenbaum ME, Bhabha
G and Vale RD: Activation of cytoplasmic dynein motility by
dynactin-cargo adapter complexes. Science. 345:337–341. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
McLaughlin RT, Diehl MR and Kolomeisky AB:
Collective dynamics of processive cytoskeletal motors. Soft Matter.
12:14–21. 2015. View Article : Google Scholar
|
|
6
|
Johnson JL, Hamm-Alvarez S, Payne G,
Sancar GB, Rajagopalan KV and Sancar A: Identification of the
second chromophore of Escherichia coli and yeast DNA photolyases as
5,10-methenyltetrahydrofolate. Proc Natl Acad Sci USA.
85:2046–2050. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schnaeker EM, Ossig R, Ludwig T, Dreier R,
Oberleithner H, Wilhelmi M and Schneider SW: Microtubule-dependent
matrix metalloproteinase-2/matrix metalloproteinase-9 exocytosis:
Prerequisite in human melanoma cell invasion. Cancer Res.
64:8924–8931. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vidulescu C, Clejan S and O'Connor KC:
Vesicle traffic through intercellular bridges in DU 145 human
prostate cancer cells. J Cell Mol Med. 8:388–396. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yoon SO, Shin S and Mercurio AM: Hypoxia
stimulates carcinoma invasion by stabilizing microtubules and
promoting the Rab11 trafficking of the alpha6beta4 integrin. Cancer
Res. 65:2761–2769. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Izidoro-Toledo TC, Borges AC, Araujo DD,
Mazzi DP, Nascimento Junior FO, Sousa JF, Alves CP, Paiva AP,
Trindade DM, Patussi EV, et al: A myosin-Va tail fragment
sequesters dynein light chains leading to apoptosis in melanoma
cells. Cell Death Dis. 4:e5472013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jiang J, Yu L, Huang X, Chen X, Li D,
Zhang Y, Tang L and Zhao S: Identification of two novel human
dynein light chain genes, DNLC2A and DNLC2B, and their expression
changes in hepatocellular carcinoma tissues from 68 Chinese
patients. Gene. 281:103–113. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dong H, Zhang H, Liang J, Yan H, Chen Y,
Shen Y, Kong Y, Wang S, Zhao G and Jin W: Digital karyotyping
reveals probable target genes at 7q21.3 locus in hepatocellular
carcinoma. BMC Med Genomics. 4:602011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Weeks ME, Hariharan D, Petronijevic L,
Radon TP, Whiteman HJ, Kocher HM, Timms JF, Lemoine NR and
Crnogorac-Jurcevic T: Analysis of the urine proteome in patients
with pancreatic ductal adenocarcinoma. Proteomics Clin Appl.
2:1047–1057. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao H, Pflug BR, Lai X and Wang M:
Pyruvate dehydrogenase alpha 1 as a target of omega-3
polyunsaturated fatty acids in human prostate cancer through a
global phosphoproteomic analysis. Proteomics. 16:2419–2431. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Parsons CL: The role of the urinary
epithelium in the pathogenesis of interstitial
cystitis/prostatitis/urethritis. Urology. 69:9–16. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Petersson J, Schreiber O, Hansson GC,
Gendler SJ, Velcich A, Lundberg JO, Roos S, Holm L and Phillipson
M: Importance and regulation of the colonic mucus barrier in a
mouse model of colitis. Am J Physiol Gastrointest Liver Physiol.
300:G327–G333. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Betge J, Schneider NI, Harbaum L,
Pollheimer MJ, Lindtner RA, Kornprat P, Ebert MP and Langner C:
MUC1, MUC2, MUC5AC, and MUC6 in colorectal cancer: Expression
profiles and clinical significance. Virchows Arch. 469:255–265.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Byrd JC and Bresalier RS: Mucins and mucin
binding proteins in colorectal cancer. Cancer Metastasis Rev.
23:77–99. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kufe DW: Mucins in cancer: function,
prognosis and therapy. Nat Rev Cancer. 9:874–885. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ivanov AI, Parkos CA and Nusrat A:
Cytoskeletal regulation of epithelial barrier function during
inflammation. Am J Pathol. 177:512–524. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bu XD, Li N, Tian XQ and Huang PL: Caco-2
and LS174T cell lines provide different models for studying mucin
expression in colon cancer. Tissue Cell. 43:201–206. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Du J, Zheng X, Cai S, Zhu Z, Tan J, Hu B,
Huang Z and Jiao H: MicroRNA506 participates in pancreatic cancer
pathogenesis by targeting PIM3. Mol Med Rep. 12:5121–5126. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ohuchida K, Mizumoto K, Yamada D, Fujii K,
Ishikawa N, Konomi H, Nagai E, Yamaguchi K, Tsuneyoshi M and Tanaka
M: Quantitative analysis of MUC1 and MUC5AC mRNA in pancreatic
juice for preoperative diagnosis of pancreatic cancer. Int J
Cancer. 118:405–411. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Baratelli C, Zichi C, Di Maio M, Brizzi
MP, Sonetto C, Scagliotti GV and Tampellini M: A systematic review
of the safety profile of the different combinations of
fluoropyrimidines and oxaliplatin in the treatment of colorectal
cancer patients. Crit Rev Oncol Hematol. 122:21–29. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Park SH, Sung JY, Han SH, Baek JH, Oh JH,
Bang SM, Cho EK, Shin DB and Lee JH: Oxaliplatin, folinic acid and
5-fluorouracil (FOLFOX-4) combination chemotherapy as second-line
treatment in advanced colorectal cancer patients with irinotecan
failure: A Korean single-center experience. Jpn J Clin Oncol.
35:531–535. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fearnhead NS, Wilding JL and Bodmer WF:
Genetics of colorectal cancer: Hereditary aspects and overview of
colorectal tumorigenesis. Br Med Bull. 64:27–43. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
van Ree JH, Nam HJ and van Deursen JM:
Mitotic kinase cascades orchestrating timely disjunction and
movement of centrosomes maintain chromosomal stability and prevent
cancer. Chromosome Res. 24:67–76. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chang CC, Chien CC, Yang SH, Chen SH and
Huang CJ: Identification and clinical correlation of decreased
expression of cytoplasmic dynein heavy chain 1 in patients with
colorectal cancer. Clin Mol Medicine. 1:6–10. 2008.
|
|
29
|
Holmberg FEO, Pedersen J, Jorgensen P,
Soendergaard C, Jensen KB and Nielsen OH: Intestinal barrier
integrity and inflammatory bowel disease: Stem cell-based
approaches to regenerate the barrier. J Tissue Eng Regen Med.
12:923–935. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gouyer V, Dubuquoy L, Robbe-Masselot C,
Neut C, Singer E, Plet S, Geboes K, Desreumaux P, Gottrand F and
Desseyn JL: Delivery of a mucin domain enriched in cysteine
residues strengthens the intestinal mucous barrier. Sci Rep.
5:95772015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tadesse S, Corner G, Dhima E, Houston M,
Guha C, Augenlicht L and Velcich A: MUC2 mucin deficiency alters
inflammatory and metabolic pathways in the mouse intestinal mucosa.
Oncotarget. 8:71456–71470. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
McAuley JL, Linden SK, Png CW, King RM,
Pennington HL, Gendler SJ, Florin TH, Hill GR, Korolik V and
McGuckin MA: MUC1 cell surface mucin is a critical element of the
mucosal barrier to infection. J Clin Invest. 117:2313–2324. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ren J, Agata N, Chen D, Li Y, Yu WH, Huang
L, Raina D, Chen W, Kharbanda S and Kufe D: Human MUC1
carcinoma-associated protein confers resistance to genotoxic
anticancer agents. Cancer Cell. 5:163–175. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cozzi PJ, Wang J, Delprado W, Perkins AC,
Allen BJ, Russell PJ and Li Y: MUC1, MUC2, MUC4, MUC5AC and MUC6
expression in the progression of prostate cancer. Clin Exp
Metastasis. 22:565–573. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Morrison C, Merati K, Marsh WL Jr, De Lott
L, Cohn DE, Young G and Frankel WL: The mucin expression profile of
endometrial carcinoma and correlation with clinical-pathologic
parameters. Appl Immunohistochem Mol Morphol. 15:426–431. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Handra-Luca A, Lamas G, Bertrand JC and
Fouret P: MUC1, MUC2, MUC4, and MUC5AC expression in salivary gland
mucoepidermoid carcinoma: Diagnostic and prognostic implications.
Am J Surg Pathol. 29:881–889. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hosseini S, Bananzadeh AM, Salek R,
Zare-Bandamiri M, Kermani AT and Mohammadianpanah M: Prognostic
significance of mucinous histologic subtype on oncologic outcomes
in patients with colorectal cancer. Ann Coloproctol. 33:57–63.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Purdie CA and Piris J: Histopathological
grade, mucinous differentiation and DNA ploidy in relation to
prognosis in colorectal carcinoma. Histopathology. 36:121–126.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xie L, Villeneuve PJ and Shaw A: Survival
of patients diagnosed with either colorectal mucinous or
non-mucinous adenocarcinoma: A population-based study in Canada.
Int J Oncol. 34:1109–1115. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Niv Y and Rokkas T: Mucin expression in
colorectal cancer (CRC): Systematic review and meta-analysis. J
Clin Gastroenterol. 53:434–440. 2018. View Article : Google Scholar
|
|
41
|
Gonul II, Cakir A and Sozen S:
Immunohistochemical expression profiles of MUC1 and MUC2 mucins in
urothelial tumors of bladder. Indian J Pathol Microbiol.
61:350–355. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Even I, Reidenbach S, Schlechter T, Berns
N, Herold R, Roth W, Krunic D, Riechmann V and Hofmann I: DLIC1,
but not DLIC2, is upregulated in colon cancer and this contributes
to proliferative overgrowth and migratory characteristics of cancer
cells. FEBS J. 286:803–820. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kalra AV and Campbell RB: Mucin impedes
cytotoxic effect of 5-FU against growth of human pancreatic cancer
cells: Overcoming cellular barriers for therapeutic gain. Br J
Cancer. 97:910–918. 2007. View Article : Google Scholar : PubMed/NCBI
|