Introduction
Dental bone implants are frequently used for
replacing missing teeth. The clinical success of bone implants is
dependent on the successful integration of the biomaterial into
regenerated bone tissue, which occurs via the adhesion,
proliferation and differentiation of osteoblasts(1–4). The adhesion
process is followed by the production of mineralized matrix
directly on the surface of the biomaterial (1–4).
However, it is estimated that 5–11% dental implants fail within
10–15 years, which then must be removed (5,6). The
loss of a dental implant is generally attributed to peri-implant
bone loss. Therefore, it is important to elucidate the mechanism of
peri-implant bone loss and facilitate the understanding of
osseointegration of implants in order to improve the success rate
of clinical implantation.
Titanium (Ti) and its alloys are choice materials
for dental implants due to the high biocompatibility and corrosion
resistance properties (7). However,
Ti implants can dissolve due to stress corrosion, galvanic
corrosion and chemical corrosion in the oral environment, resulting
in the precipitation of Ti ions (8). In vivo studies have
demonstrated that the release of Ti ions into surrounding tissue,
regional lymph nodes and internal organs can occur during or after
implant placement (9–11). In a rabbit model based on
synchrotron radiation X-ray fluorescence, Ti ions were detected in
bony tissues 12 weeks and 1 year after implant placement (12).
Ti ions released from dental implants play an
important role in peri-implant complications, including bone loss
via several mechanisms (13–15).
Bitar and Parvizi (16) identified
cytokine-mediated osteolytic changes caused by reactive oxygen
species (ROS) intermediates and activation of the complement
cascade as factors affecting bone metabolismin macrophages from
mice in vitro. Mitochondria, as the power plant of cells,
are the primary location of ROS production. However, to the best of
our knowledge, the effect of Ti ions on mitochondria derived ROS
(mROS) has not been previously studied in depth.
Therefore, the present study investigated the
mechanism of Ti ion damage to osteoblasts, and the possible
mechanism of increased mROS production.
Materials and methods
Cell culture
Osteoblastic cells were purchased from Lonza
(Clonetics Normal Human Osteoblast Cell System). Cells were
cultured in RPMI-1640 medium (Gibco; Thermo Fisher Scientific,
Inc.) containing 10% FBS (Gibco; Thermo Fisher Scientific, Inc.)
and 1% (v/v) penicillin/streptomycin (cat. no. P4333;
Sigma-Aldrich; Merck KGaA) and incubated at a humidified atmosphere
with 5% CO2 at 37°C. The culture medium was changed
every 2 days. At 70–80% confluency, adhered cells were detached
using 0.25% trypsin/EDTA solution. Osteoblastic cells of passages
4–6 were used for the subsequent experiments. All experiments were
performed three times.
Cell experimental protocol
The effect of Ti ions on autophagy in osteoblastic
cells was evaluated. Cells were treated with TiCl4 (cat.
no. 7550-45-0; Shanghai Aladdin Bio-Chem Technology Co., Ltd.) at
different concentrations (0, 10, 25 and 50 µM) at 37°C for 24 h.
The cells were also treated with various concentrations (0, 10, 25
and 50 µM) of TiCl4 for 24 h with or without lysosomal
inhibitor bafilomycin A1 (Baf-A1; 20 nM; Cell Signaling Technology,
Inc.) at 37°C for 24 h. The vehicle control used for each in
vitro assay was 0.1% DMSO. The role of the sirtuin3
(SIRT3)/superoxide dismutase 2 (SOD2) pathway in osteoblasts was
investigated.
Cell viability
Cell viability was analyzed using the Cell Counting
Kit-8 (CCK-8) according to the manufacturer's instructions (cat.
no. CK04; Dojindo Molecular Technologies, Inc.). Briefly,
1×104 cells were inoculated into 96-well plates. After
treatment, 90 ml RPMI 1640 medium and 10 ml CCK-8 solution were
added to each well. The cells were then incubated at 37°C for 2 h,
and the absorption was measured at 450 nm using an Infinite M200
microplate reader (Tecan Group, Ltd.). Data are presented as
percentages relative to the control.
Determination of mROS
After treatment, osteoblastic cells were incubated
with culture medium containing 10 mM MitoSOX (cat. no. M36008;
Invitrogen; Thermo Fisher Scientific, Inc.) for 20 min at 37°C.
Osteoblastic cells were pre-incubated at 37°C with 10 mM Mito-TEMPO
(Sigma-Aldrich; Merck KGaA) for 2 h before the 24 h
TiCl4 exposure.The fluorescence intensity was analyzed
with an Infinite M200 Microplate Reader (Tecan Group, Ltd.) at an
excitation wavelength of 492 nm and an emission wavelength of 595
nm. Cellular fluorescence intensity was expressed as fold-change
relative to control.
Measurement of SOD2 enzyme
activity
SOD2 enzymatic activity was assessed using a SOD1
and SOD2 assay kit (cat. no. S0103; Beyotime Institute of
Biotechnology) according to the manufacturer's instructions. For
quantification, one unit of SOD was defined as the amount of enzyme
that inhibited the rate of nitro-blue tetrazoliumreduction observed
in a blank sample by 50% (17). The
SOD isoforms were identified by adding the SOD1 inhibitor A and B
according to the manufacturer's instructions to inhibit SOD1
activity (i.e., to detect SOD2). The absorption at 450 nm was
measured using an Infinite M200 microplate reader.
Plasmids and transfection
LV5-SIRT3 overexpression lentivirus were purchased
from Yuxi Biotechnology Corporation (Fig. S1A). The transfection assay was
performed according to the manufacturer's protocols. Osteoblastic
cells were grown in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
with 20% FBS (Gibco; Thermo Fisher Scientific, Inc.) on culture
dishesat a humidified atmosphere of 5% CO2 at 37°C, and
were infected with the lentivirus for 72–96 h (Fig. S1B). The lentivirus contained green
fluorescent protein, which was visible in fluorescence microscopy.
Cells were then washedwith DMEM (Gibco; Thermo Fisher Scientific,
Inc.) and processed for subsequent experiments. Western blotting
was used to show that transfection was successful (Fig. S1C).
Reverse transcription-quantitative PCR
(RT-qPCR)
TRIzol® (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to extract RNA from osteoblastic cells,
and its concentration and purity were detected with a UV
spectrophotometer. cDNA was synthesized using a Sensiscript RT kit
(Thermo Fisher Scientific, Inc.). All reagents used for RT-qPCR to
detect SIRT3 mRNA were obtained from Thermo Fisher Scientific, Inc.
The following primer sequences were used: SIRT3 forward, 5′-
GACATCCCGTACCCTGAAG-3; and reverse, 5′-AGCTGTTACAAAGGTCCCGT-3′; and
GAPDH forward, 5′-CGAGAATGGGAAGCTTGTCA-3′ and reverse,
5′-GACATCCCGTACCCTGAAG-3′. The reaction system consisted of 4 µl 5X
buffer + 1.5 µl MgCl2 + 0.5 µl dNTP + 0.8 µl sense and
antisense primers + 0.3 µl Taq enzyme + 2 µl cDNA template, + 0.4
µl passive dye (SYBR Green; Thermo Fisher Scientific, Inc.) and
11.5 µl water to 20 µl. The reaction conditions were: Initial
denaturation at 95°C for 3 min, followed by 35 cycles at 95°C for
30 sec, 55°C for 20 sec and 72°C for 20 sec; and a final extension
step at 72°C for 10 min. The dissociation curve was obtained and
the 2−ΔΔCq method (18)
was used to calculate the relative expression of mRNA.
SIRT3 activity
A fluorometric kit (cat. no. BML-AK557-0001; Enzo
Life Sciences, Inc.) was used to examine SIRT3 enzymatic activity.
The osteoblastic cells were treated and lysed with cell lysis
buffer (Beyotime Institute of Biotechnology). Lysates were
centrifuged at 12,000 × g for 15 min at 4°C and the resulting
supernatant was transferred to a new tube. The protein
concentrations were determined using a Bradford protein assay kit
(Beyotime Institute of Biotechnology). In total, 40 mg protein was
incubated at 37°C for 45 min with specific substrates, following
the manufacturer's instructions. Then, 25 ml developer was added
and samples were incubated for 45 min at 37°C. SIRT3 activity was
measured using an Infinite M200 microplate reader at the
wavelengths 350 and 450 nm.
Immunoprecipitation
Osteoblastic cells were treated with
TiCl4 at different concentrations (0, 10, 25 and 50 µM)
at 37°C for 24 h and lysed with cell lysis buffer (Beyotime
Institute of Biotechnology; cat. no. P0013). Lysates were clarified
by centrifugation at 12,000 × g for 15 min at 4°C and were used for
immunoprecipitation. The protein concentrations were determined
using a Bradford protein assay kit (Beyotime Institute of
Biotechnology). A total of 2 µg anti-SOD2 (1:200; Abcam; cat. no.
Ab137037) was incubated with 500-1,000 µg protein overnight at 4°C.
This was followed by incubation with 20 µl protein A beads
(Beyotime Institute of Biotechnology; cat. no. P2006) overnight at
4°C. After incubation, the beads were washed three times with PBS
and subsequently boiled in 40 µl 1X sample buffer for 5 min at
100°C, and analyzed by western blotting.
Western blotting
Osteoblastic cells were treated at 37°C for 24 h and
lysed with cell lysis buffer (Beyotime Institute of Biotechnology;
cat. no. P0013). Lysates were clarified by centrifugation at 12,000
× g for 15 min at 4°C and the resulting supernatant was transferred
to a new tube. The protein concentrations were determined using a
Bradford protein assay kit (Beyotime Institute of Biotechnology).
The protein samples (60 µg/lane) were separated by SDS-PAGE (12%
gel). Following protein transfer onto PVDF membranes, the membranes
were blocked with 20 ml sealing liquid (1 g skimmed milk powder, 20
ml 2% PBS-Tween 20) on a horizontal shaker and left to agitate at
25°C for 2 h. Then, membranes were incubated overnight at 4°C with
antibodies against microtubule associated protein 1 light chain 3 α
(LC3;1:1,000; cat. no. L7543; Sigma-Aldrich; Merck KGaA),
SQSTM1/p62 (1:1,000; Abcam; cat. no. ab56416), SIRT3 (1:500; cat.
no. sc-99143; Santa Cruz Biotechnology, Inc.), SOD2 (1:1,000; cat.
no.sc-33254; Santa Cruz Biotechnology, Inc.), Ac-lysine (1:500;
cat. no. Ab137037; Abcam) and GAPDH (1:5,000; cat. no. A5441;
Sigma-Aldrich; Merck KGaA). The membranes were incubated for 1 h at
25°C with the secondary antibodies, horseradish
peroxidase-conjugated affinity-purified goat anti-rabbit IgG
(1:2,000; cat. no. AP132; Sigma-Aldrich; Merck KGaA). Signals were
detected using Super Signal West Femto Trial kit (Thermo Fisher
Scientific, Inc.). The membranes were exposed to Hyper Performance
Chemiluminescence film (GE Healthcare Life Sciences). The relative
amount of proteins was quantified using gel analysis software
UNSCAN-IT gel (version 5.3; Silk Scientific, Inc.) and normalized
to GAPDH internal loading control.
Immunofluorescence
Cells were collected and fixed with4%
paraformaldehyde for 10 min at room temperature., and subsequently
incubated with 0.5% Triton X-100 at 25°C for 20 min. Then, cells
were incubated with anti-LC 3(1:200; cat. no. L7543; Sigma-Aldrich;
Merck KGaA) overnight at 4°C after blocking with 5% BSA (Gibco;
Thermo Fisher Scientific, Inc.) in PBS. After washingwith PBS, the
cells were incubated with FITC labeled goat anti-rabbit
Immunoglobulin G (1:2,000; cat. no. AP132; Sigma-Aldrich; Merck
KGaA) at 25°C for 1 hand PI (50 µg/ml; cat. no. C1002; Beyotime
Institute of Biotechnology) for 15 min. Then, images
(magnification, ×1000) were captured using a fluorescent microscope
(BX51; Olympus Corporation).
Statistical analysis
Data are presented as the mean ± standard deviation
from ≥3 independent experiments. The data were analyzed by one-way
ANOVA test using GraphPad Prism-5 software (GraphPad Software,
Inc.), followed by Tukey's test for multiple comparisons using SPSS
(v.10.01; IBM Corp.). P<0.05 was considered to indicate a
statistically significant difference.
Results
Ti ions can lead to osteoblastic
cellular damage via autophagy
To determine whether Ti ions can damage osteoblastic
cells, the present study established a cellular model. It was found
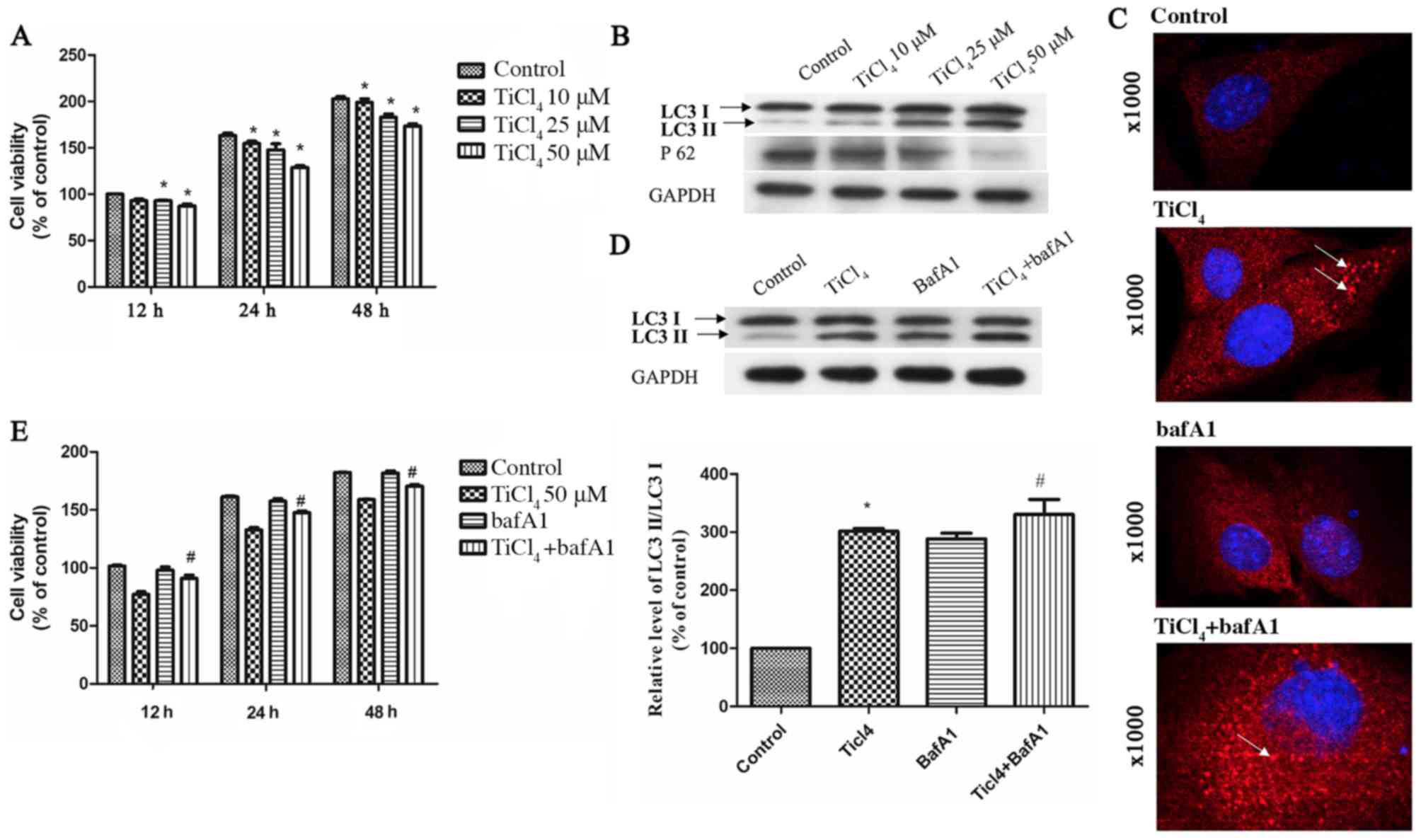
that Ti ion exposure decreased osteoblastic cell viability in a
dose-dependent manner (Fig. 1A). To
investigate whether autophagy is involved in osteoblast injury
caused by Ti ions, the expression level of LC3II was assessed as a
marker of autophagy in osteoblastic cells exposed to Ti ions. The
present results suggested that the protein expression level of
LC3II increased, while the expression level of sequestosome 1(P62)
decreased with increasing Ti ion dosage (Fig. 1B). The present study also examined
osteoblastic cells by fluorescence microscopy, and found a
significant increase in autophagy in TiCl4 group
(Fig. 1C).
The basal level of autophagy has a
protective effect on cells, but excessive autophagy can induce
autophagic cell death
To investigate the effects of autophagy on
osteoblastic cells caused by Ti ions, the apoptotic rate of
osteoblastic cells treated with Ti ions was examined. No
significant rate of apoptosis of osteoblastic cells was identified
(data not shown). In addition, osteoblasts were treated with
Bafilomycin A1 (BafA1), an autophagic inhibitor of the proton pump
of autophagic lysosomes that can inhibit the activity of
autophagosomes and reduce their degradation (19). After treatment with BafA1, the
expression level of LC3II increased in the osteoblasts (Fig. 1C and D), suggesting that enhanced
LC3II after Ti ion exposure increased the autophagic flow. In
addition, it was found that cell viability improved after treatment
with BafA1 compared with TiCl4 group (Fig. 1E).
Ti ions increase autophagy via
mROS
Autophagy has been shown to be related to mROS
(20). The initial oxygen reduction
product in mitochondria is O2·, which is then rapidly
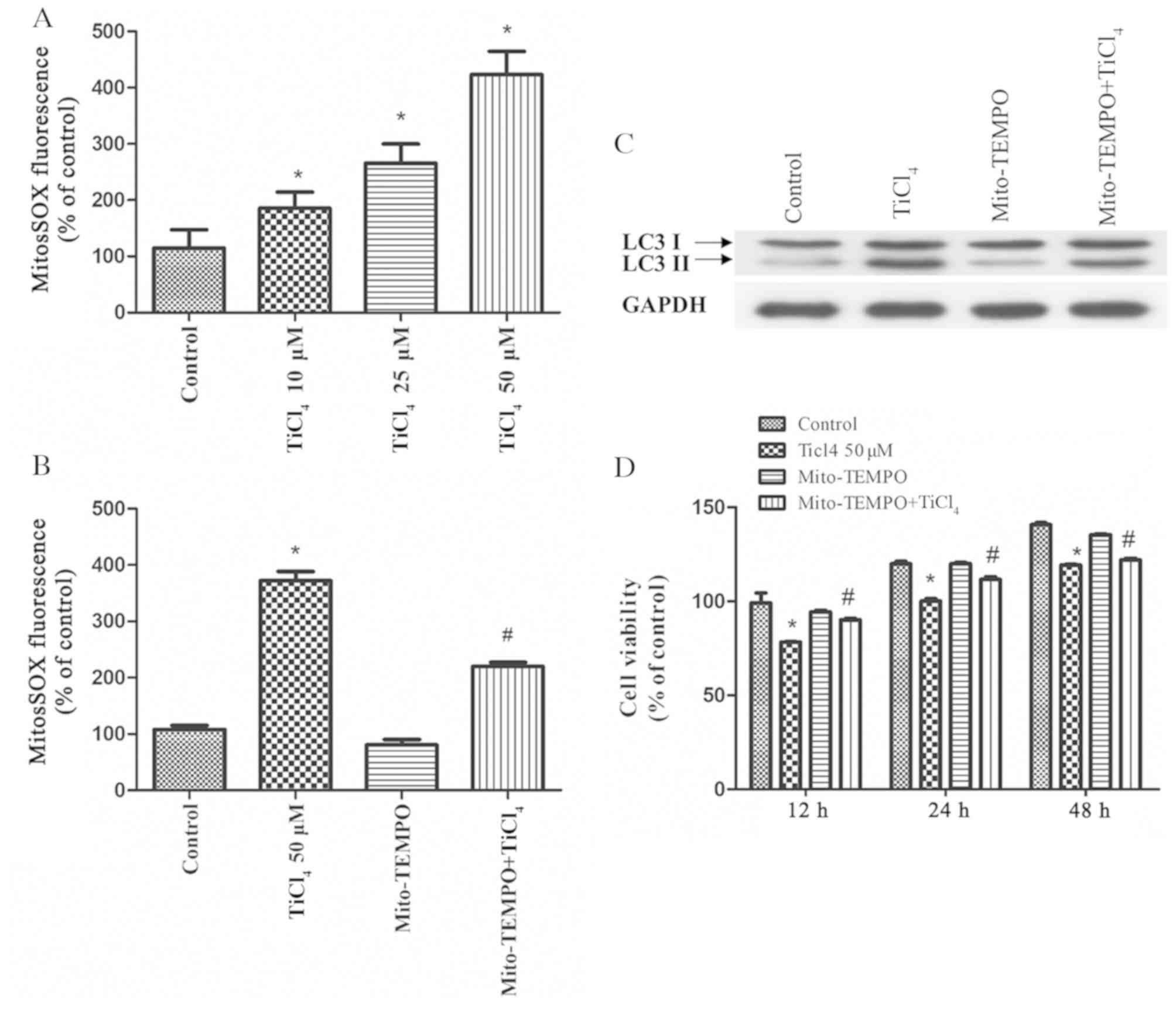
converted to H2O2 (21). Therefore, the present study
investigate the levels of mROS in osteoblasts treated with Ti ions,
and identified significantly elevated of mROS levels in these cells
(Fig. 2A). Furthermore,
osteoblastic cells were pretreated with the mitochondrial
antioxidant, Mito-TEMPO, which targets SOD2 in mitochondria, for 2
h before co-culture with Ti ions. It was demonstrated that
Mito-TEMPO inhibited the production of mROS (Fig. 2B), and it inhibited the expression
level of LC3II caused by Ti ions (Fig.
2C). Moreover, the viability of osteoblastic cells was
significantly increased by pretreatment with Mito-TEMPO compared
with TiCl4 group (Fig.
2D). Collectively, these results suggested that oxidative
stress induced by Ti ions triggers autophagy, while antioxidants,
such as Mito-TEMPO, can mitigate mitochondrial oxidative stress and
reduce this excess autophagy.
Ti ions can increase SOD2 acetylation
by inhibiting SIRT3 expression and activity
SOD2 is the main scavenger of oxygen free radicals
in mitochondria (22). SOD2
catalyzes the conversion of O2· to
H2O2 and plays an important role in
regulating mROS in mitochondria (22). Therefore, the present study examined
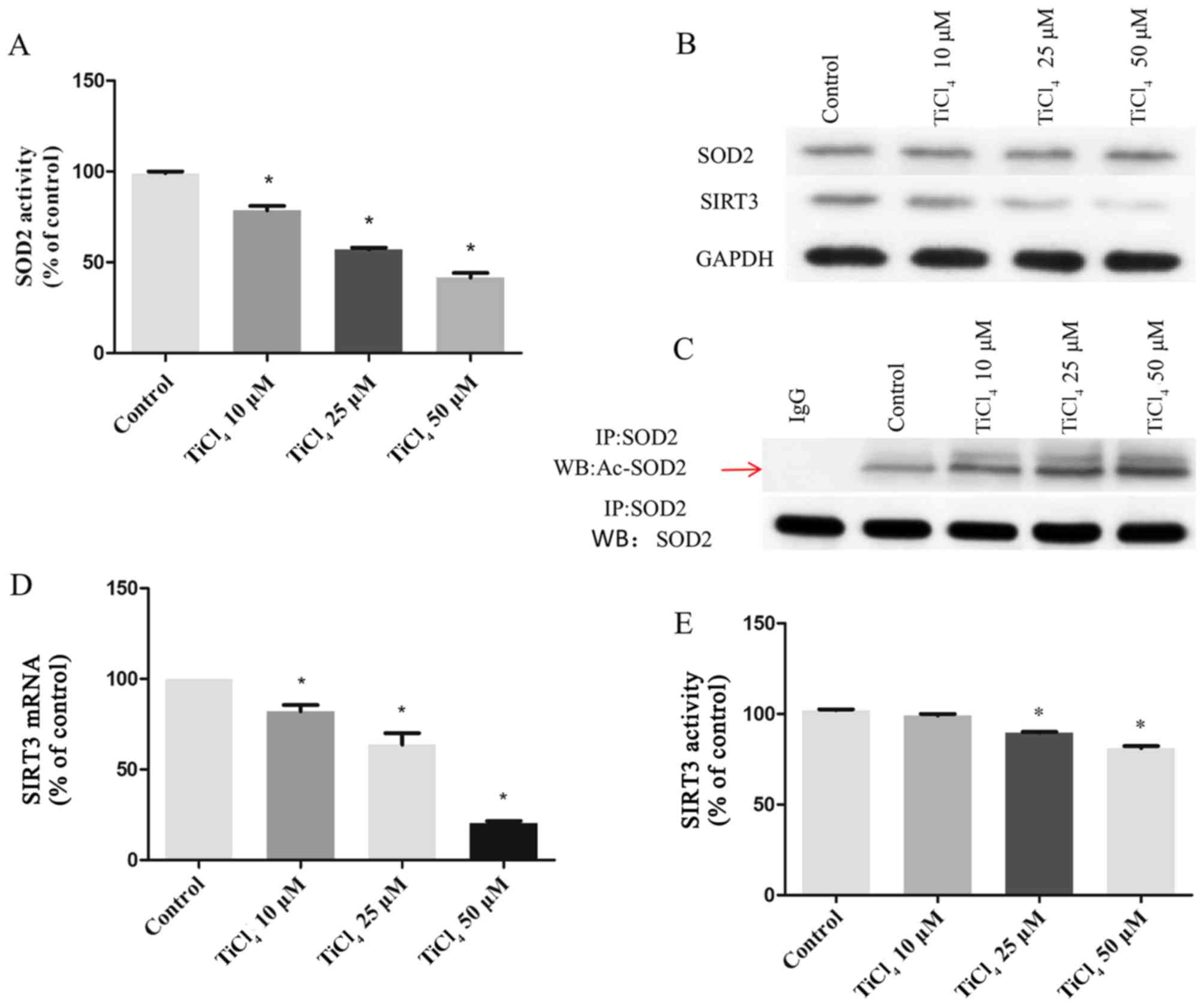
the effects of Ti ions on SOD2 expression levels. It was
demonstrated that the activity of SOD2 decreased with increasing Ti
ion dose (Fig. 3A), but no effect
was observed on the protein expression levels of SOD2 (Fig. 3B). SOD2 activity is regulated by the
acetylation of lysine residues (22–24).
Therefore, the present study detected the acetylation level of SOD2
by coimmunoprecipitation and western blotting. It was found that Ti
ion exposure resulted in a progressive increase in SOD2 acetylation
level (Fig. 3C). Deacetylation of
SOD2 is mainly regulated by the mitochondrial deacetylase SIRT3
(25–27). Therefore, the present study
investigated SIRT3 activity and its concentration in osteoblastic
cells treated with Ti ions. The present results suggested that Ti
ions caused a significant decrease in the mRNA (Fig. 3D) and protein (Fig. 3B) expression levels of SIRT3 in a
dose-dependent manner. The present results also indicated that the
levels of SIRT3 activity in the Ti ion group were significantly
lower compared with the control group (Fig. 3E).
SIRT3 and SOD2 modulate Ti ion-induced
mROS accumulation and autophagy
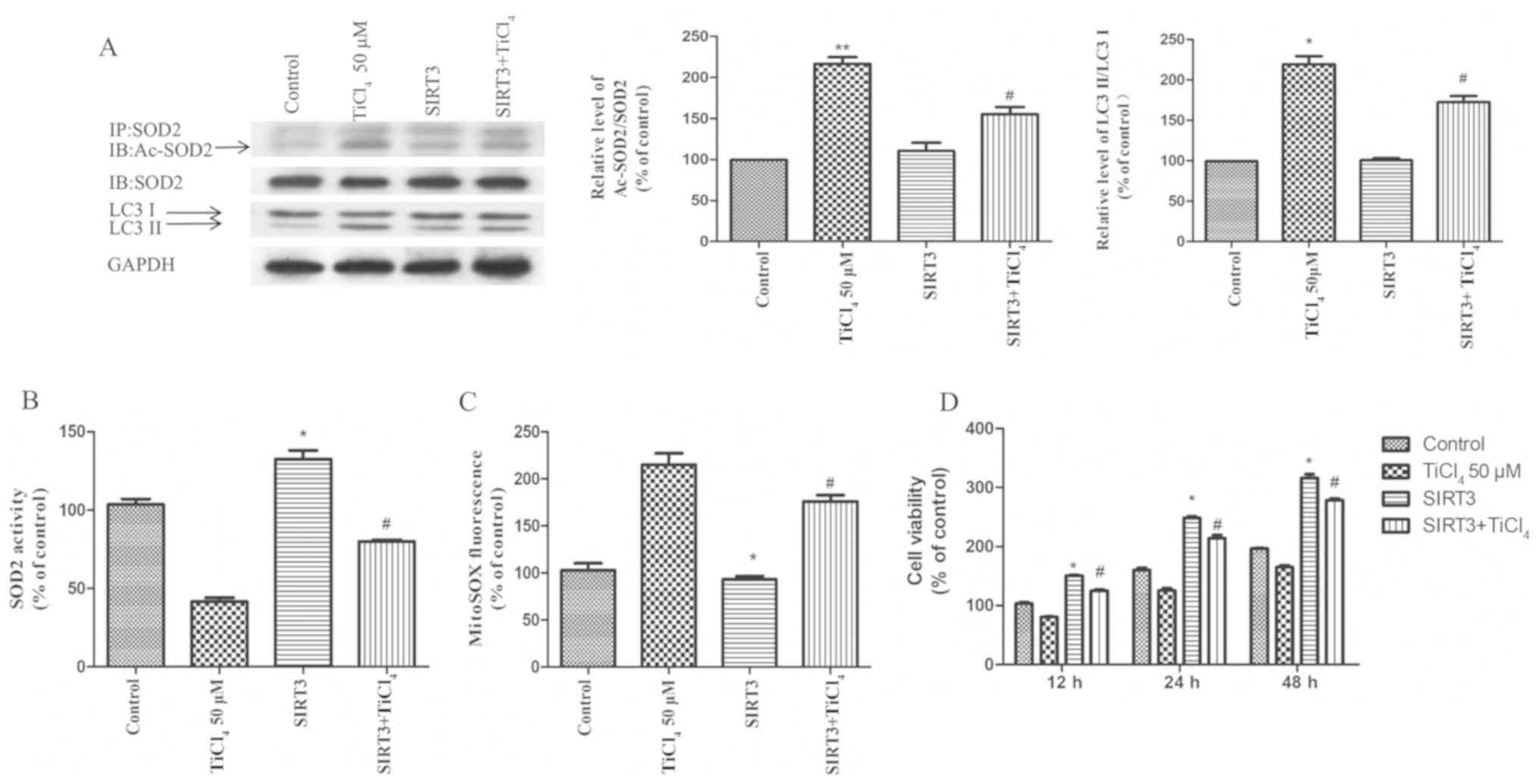
To examine whether the recovery of SIRT3 protein
expression level or activity could prevent autophagy induced by Ti
ion exposure, the present study transfected SIRT3 into osteoblastic
cells to overexpress SIRT3. It was demonstrated that overexpression
of SIRT3 ameliorated the inhibition of SIRT3 activity induced by Ti
ions, reduced the acetylation of SOD2 (Fig. 4A) and increased the activity of SOD2
(Fig. 4B). Furthermore, SIRT3
overexpression significantly reduced the level of mROS produced by
Ti ions (Fig. 4C). The present
results also suggested that the overexpression of SIRT3 increased
cell viability and significantly reduced the expression levels of
LC3II in Ti ion treated cells (Fig. 4A
and D).
Discussion
Ti is used extensively in dental implants. However,
clinical studies have found that Ti particles released from Ti
implants can lead to aseptic loosening of orthopedic implants
(28,29). The corrosive properties of Ti are
increased in low dissolved-oxygen conditions such as in the oral
cavity, and especially in the presence of small amounts of fluoride
(8,30,31).
During implant placement, increasing osteoblast adhesion and
proliferation on a Ti surface is essential for successful
osseointegration (32). The
osseointegration process can be divided into hemostasis,
inflammatory, proliferation and a remodeling phase (33). In the inflammatory phase,
infiltrated inflammatory cells synthesize and secrete ROS (34). Furthermore, Ti ions released from
metal graft material can stimulate ROS synthesis (35). This overproduction of ROS can result
in failure of implant placement by reducing osseointegration via
dysfunction and apoptosis of the osteoblasts (36). N-acetyl-cysteine, a potent ROS
inhibitor, enhances osseointegration by scavenging ROS in
osteoblasts on the Ti alloy surface (37). Moreover, insulin improves the
impaired osteogenesis of osteoblasts on the Ti surface by
inhibiting ROS overproduction (38). In contrast to previous studies
(35,37,38),
the present results suggested that Ti ions could lead to the
production of ROS via the mitochondrial pathway. Instead of
apoptotic cell death (36), the
present results suggested that ROS could lead to the autophagic
death of osteoblasts.
By removing cells damaged by intracellular pathogens
and toxic metabolites, autophagy aids in cell survival (39). However, autophagy may cause cell
death via the degradation of essential cellular constituents and
excessive self-digestion (40). The
present results indicated that Ti ion induced increases in LC3
expression may be due to an elevation of autophagic flux. In
addition, it was demonstrated that Ti ion exposure decreased the
viability of osteoblasts. Collectively, the present results
suggested that autophagy may contribute to Ti ion-mediated
osteoblast damage in the periodontium.
Increased generation of ROS is an important stimulus
of autophagy in several diseases (41–45).
It is reported that autophagic removal and degradation of damaged
proteins caused by mitochondrial oxidative stress is beneficial for
cell survival (46). However,
excessive amounts of ROS and severe oxidative stress may activate
signaling pathways, and cause autophagic cell death (20,47).
The present results suggested that Ti ion exposure promoted mROS
2-fold compared with the control group. Moreover, Mito-TEMPO
treatment, the mitochondrial-targeted SOD mimetic, reduced
oxidative stress and decreased autophagic cell death, suggesting
that mROS accumulation is an important mechanism underlying the
sensitization of cells to autophagy.
Although mitochondrial oxidative stress is
associated with autophagy (48),
the molecular mechanism of oxygen free radical aggregation from
mitochondria is not fully understood. Cells can regulate oxygen
free radicals derived from mitochondria via a specific mechanism
(47). Furthermore, mitochondria
have a special function of removing oxygen free radicals from
mitochondria to maintain homeostasis (49). Mitochondria consume 90% of
intracellular oxygen, so SOD2 activity is important in maintaining
the balance of oxygen free radicals derived from mitochondria
(23,50). SOD2 is an enzyme that scavenges free
radicals, and is only found in mitochondria (51,52).
However, in the present study, it was found that the protein
expression of SOD2 was not affected by Ti ion treatment. The amount
of SOD2 protein is regulated by transcription, and its activity is
regulated by acetylation level (23,24,50).
Consistent with previous studies (53,54),
Ti ion exposure in the present study resulted in an elevated level
of acetylation of SOD2 in a dose-dependent manner. Therefore, the
present results suggested that Ti ions increased mitochondrial
oxygen free radicals by increasing SOD2 acetylation levels, rather
than by decreasing SOD2 protein expression levels.
Lysine acetylation is an important
post-translational modification for the regulation of mitochondrial
proteins and autophagy (55–57).
In total, three deacetylases SIRT3, SIRT4 and SIRT5, are found in
mitochondria. SIRT3 is the strongest deacetylase in mitochondria,
and is directly involved in mitochondrial energy synthesis and the
control of oxygen free radical levels (47,58).
Moreover, SIRT3 controls SOD2 activity by regulating the
acetylation level of SOD2, and its target lysine has been
identified (59). SIRT3
deacetylates SOD2 in response to ionizing radiation, indicating
that SOD2 is a major downstream signal of SIRT3-mediated
mitochondrial derived O2 reduction (24,47).
The present results indicated that Ti ions did not decrease SOD2
expression levels, but increased acetylation levels. Furthermore,
it was demonstrated that SIRT3 expression level and activity were
decreased. It was also found that SIRT3 overexpression could repair
the increase inSOD2 acetylation caused by Ti ions, reduce the
acetylation of SOD2 and increase SOD2 activity, thus maintaining
the stability of mitochondrial oxygen free radicals and reversing
autophagy induced by Ti ions. Therefore, SIRT3/SOD2 mediated
autophagy may be an important mechanism of osteoblastic cells
damage induced by Ti ions.
In conclusion, the present results suggested that
there is a dose-response relationship between Ti ions and
osteoblastic cells damage. Furthermore, increased autophagy-induced
by mROS may be one of the important factors of Ti ion-induced
osteoblastic cell damage. Moreover, Ti ions may lead to the
dysfunction of SIRT3 and SOD2 in mitochondria. Therefore,
inhibition of SIRT3 activity is an important mechanism of
osteoblastic cellular damage induced by Ti ion exposure.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
JM and XX contributed to the concept and the design
of the study. SW, JY and TL performed research. SH acquired the
data and performed statistical analysis. SW and JY drafted the
manuscript. XX revised the manuscript. All authors read and
approved the final manuscript
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Anselme K: Osteoblast adhesion on
biomaterials. Biomaterials. 21:667–681. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Anselme K: Biomaterials and interface with
bone. Osteoporos Int. 22:2037–2042. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Davies JE: Mechanisms of endosseous
integration. Int J Prosthodont. 11:391–401. 1998.PubMed/NCBI
|
|
4
|
Davies JE: Understanding peri-implant
endosseous healing. J Dent Educ. 67:932–949. 2003.PubMed/NCBI
|
|
5
|
Hermann JS, Buser D, Schenk RK,
Schoolfield JD and Cochran DL: Biologic Width around one- and
two-piece titanium implants. Clin Oral Implants Res. 12:559–571.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lindhe J and Meyle J; Group D of European
Workshop on Periodontology, : Peri-implant diseases: Consensus
Report of the Sixth European Workshop on Periodontology. J Clin
Periodontol. 35 (Suppl):282–285. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brånemark PI, Adell R, Albrektsson T,
Lekholm U, Lundkvist S and Rockler B: Osseointegrated titanium
fixtures in the treatment of edentulousness. Biomaterials. 4:25–28.
1983. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wachi T, Shuto T, Shinohara Y, Matono Y
and Makihira S: Release of titanium ions from an implant surface
and their effect on cytokine production related to alveolar bone
resorption. Toxicology. 327:1–9. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schwarz F, Sahm N, Mihatovic I, Golubovic
V and Becker J: Surgical therapy of advanced ligature-induced
peri-implantitis defects: Cone-beam computed tomographic and
histological analysis. J Clin Periodontol. 38:939–949. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kumazawa R, Watari F, Takashi N, Tanimura
Y, Uo M and Totsuka Y: Effects of Ti ions and particles on
neutrophil function and morphology. Biomaterials. 23:3757–3764.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wennerberg A, Ide-Ektessabi A, Hatkamata
S, Sawase T, Johansson C, Albrektsson T, Martinelli A, Södervall U
and Odelius H: Titanium release from implants prepared with
different surface roughness. Clin Oral Implants Res. 15:505–512.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ellingsen JE, Johansson CB, Wennerberg A
and Holmén A: Improved retention and bone-tolmplant contact with
fluoride-modified titanium implants. Int J Oral Maxillofac
Implants. 19:659–666. 2004.PubMed/NCBI
|
|
13
|
Pajarinen J, Kouri VP, Jämsen E, Li TF,
Mandelin J and Konttinen YT: The response of macrophages to
titanium particles is determined by macrophage polarization. Acta
Biomater. 9:9229–9240. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Obando-Pereda GA, Fischer L and
Stach-Machado DR: Titanium and zirconia particle-induced
pro-inflammatory gene expression in cultured macrophages and
osteolysis, inflammatory hyperalgesia and edema in vivo. Life Sci.
97:96–106. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Brettin BT, Grosland NM, Qian F, Southard
KA, Stuntz TD, Morgan TA, Marshall SD and Southard TE: Bicortical
vs monocortical orthodontic skeletal anchorage. Am J Orthod
Dentofacial Orthop. 134:625–635. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bitar D and Parvizi J: Biological response
to prosthetic debris. World J Orthop. 6:172–189. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Grunow M and Schöpp W: [Determination of
the activity of superoxide dismutase in terms of rea substrate
conversion]. Biomed Biochim Acta. 48:185–199. 1989.(In German).
PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Klionsky DJ, Elazar Z, Seglen PO and
Rubinsztein DC: Does bafilomycin A1 block the fusion of
autophagosomes with lysosomes? Autophagy. 4:849–850. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sena LA and Chandel NS: Physiological
roles of mitochondrial reactive oxygen species. Mol Cell.
48:158–167. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lambert AJ and Brand MD: Reactive oxygen
species production by mitochondria. Methods Mol Biol. 554:165–181.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tao R, Vassilopoulos A, Parisiadou L, Yan
Y and Gius D: Regulation of MnSOD enzymatic activity by Sirt3
connects the mitochondrial acetylome signaling networks to aging
and carcinogenesis. Antioxid Redox Signal. 20:1646–1654. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen Y, Zhang J, Lin Y, Lei Q, Guan KL,
Zhao S and Xiong Y: Tumour suppressor SIRT3 deacetylates and
activates manganese superoxide dismutase to scavenge ROS. EMBO Rep.
12:534–541. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tao R, Coleman MC, Pennington JD, Ozden O,
Park SH, Jiang H, Kim HS, Flynn CR, Hill S, Hayes McDonald W, et
al: Sirt3-mediated deacetylation of evolutionarily conserved lysine
122 regulates MnSOD activity in response to stress. Mol Cell.
40:893–904. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Di Tucci AA, Murru R, Alberti D, Rabault
B, Deplano S and Angelucci E: Correction of anemia in a
transfusion-dependent patient with primary myelofibrosis receiving
iron chelation therapy with deferasirox (Exjade, ICL670). Eur J
Haematol. 78:540–542. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Messa E, Cilloni D, Messa F, Arruga F,
Roetto A and Saglio G: Deferasirox treatment improved the
hemoglobin level and decreased transfusion requirements in four
patients with the myelodysplastic syndrome and primary
myelofibrosis. Acta Haematol. 120:70–74. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Angelucci E, Santini V, Di Tucci AA,
Quaresmini G, Finelli C, Volpe A, Quarta G, Rivellini F, Sanpaolo
G, Cilloni D, et al: Deferasirox for transfusion-dependent patients
with myelodysplastic syndromes: Safety, efficacy, and beyond
(GIMEMA MDS0306 Trial). Eur J Haematol. 92:527–536. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jacobs JJ, Gilbert JL and Urban RM:
Corrosion of metal orthopaedic implants. J Bone Joint Surg Am.
80:268–282. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gallo J, Kamínek P, Tichá V, Riháková P
and Ditmar R: Particle disease. A comprehensive theory of
periprosthetic osteolysis: A review. Biomed Pap Med Fac Univ
Palacky Olomouc Czech Repub. 146:21–28. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nakagawa M, Matsuya S and Udoh K: Effects
of fluoride and dissolved oxygen concentrations on the corrosion
behavior of pure titanium and titanium alloys. Dent Mater J.
21:83–92. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Matono Y, Nakagawa M, Matsuya S, Ishikawa
K and Terada Y: Corrosion behavior of pure titanium and titanium
alloys in various concentrations of Acidulated Phosphate Fluoride
(APF) solutions. Dent Mater J. 25:104–112. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Roehlecke C, Witt M, Kasper M, Schulze E,
Wolf C, Hofer A and Funk RW: Synergistic effect of titanium alloy
and collagen type I on cell adhesion, proliferation and
differentiation of osteoblast-like cells. Cells Tissues Organs.
168:178–187. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Terheyden H, Lang NP, Bierbaum S and
Stadlinger B: Osseointegration--communication of cells. Clin Oral
Implants Res. 23:1127–1135. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Rajendrakumar SK, Revuri V, Samidurai M,
Mohapatra A, Lee JH, Ganesan P, Jo J, Lee YK and Park IK:
Peroxidase-mimicking nanoassembly mitigates
lipopolysaccharide-induced endotoxemia and cognitive damage in the
brain by impeding inflammatory signaling in macrophages. Nano Lett.
18:6417–6426. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Landgraeber S, Jäger M, Jacobs JJ and
Hallab NJ: The pathology of orthopedic implant failure is mediated
by innate immune system cytokines. Mediators Inflamm.
2014:1851502014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li X, Ma XY, Feng YF, Ma ZS, Wang J, Ma
TC, Qi W, Lei W and Wang L: Osseointegration of chitosan coated
porous titanium alloy implant by reactive oxygen species-mediated
activation of the PI3K/AKT pathway under diabetic conditions.
Biomaterials. 36:44–54. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Feng YF, Wang L, Zhang Y, Li X, Ma ZS, Zou
JW, Lei W and Zhang ZY: Effect of reactive oxygen species
overproduction on osteogenesis of porous titanium implant in the
present of diabetes mellitus. Biomaterials. 34:2234–2243. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang L, Zhao X, Wei BY, Liu Y, Ma XY, Wang
J, Cao PC, Zhang Y, Yan YB, Lei W, et al: Insulin improves
osteogenesis of titanium implants under diabetic conditions by
inhibiting reactive oxygen species overproduction via the PI3K-Akt
pathway. Biochimie. 108:85–93. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Klionsky DJ, Abdelmohsen K, Abe A, Abedin
MDJ, Abeliovich H, Arozena AA, Adachi H, Adams CM, Adams PD, Adeli
K, et al: Guidelines for the use and interpretation of assays for
monitoring autophagy (3rd edition). Autophagy. 12:1–222. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu Y and Levine B: Autosis and autophagic
cell death: The dark side of autophagy. Cell Death Differ.
22:367–376. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Essick EE and Sam F: Oxidative stress and
autophagy in cardiac disease, neurological disorders, aging and
cancer. Oxid Med Cell Longev. 3:168–177. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang T, Li Y, Park KA, Byun HS, Won M,
Jeon J, Lee Y, Seok JH, Choi SW, Lee SH, et al: Cucurbitacin
induces autophagy through mitochondrial ROS production which
counteracts to limit caspase-dependent apoptosis. Autophagy.
8:559–576. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shen W, Tian C, Chen H, Yang Y, Zhu D, Gao
P and Liu J: Oxidative stress mediates chemerin-induced autophagy
in endothelial cells. Free Radic Biol Med. 55:73–82. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Dai DF and Rabinovitch P: Mitochondrial
oxidative stress mediates induction of autophagy and hypertrophy in
angiotensin-II treated mouse hearts. Autophagy. 7:917–918. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kim Y, Kim YS, Kim DE, Lee JS, Song JH,
Kim HG, Cho DH, Jeong SY, Jin DH, Jang SJ, et al: BIX-01294 induces
autophagy-associated cell death via EHMT2/G9a dysfunction and
intracellular reactive oxygen species production. Autophagy.
9:2126–2139. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liang Q, Benavides GA, Vassilopoulos A,
Gius D, Darley-Usmar V and Zhang J: Bioenergetic and autophagic
control by Sirt3 in response to nutrient deprivation in mouse
embryonic fibroblasts. Biochem J. 454:249–257. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Pi H, Xu S, Reiter RJ, Guo P, Zhang L, Li
Y, Li M, Cao Z, Tian L, Xie J, et al: SIRT3-SOD2-mROS-dependent
autophagy in cadmium-induced hepatotoxicity and salvage by
melatonin. Autophagy. 11:1037–1051. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lee J, Giordano S and Zhang J: Autophagy,
mitochondria and oxidative stress: Cross-talk and redox signalling.
Biochem J. 441:523–540. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zorov DB, Juhaszova M and Sollott SJ:
Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS
release. Physiol Rev. 94:909–950. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhu Y, Park SH, Ozden O, Kim HS, Jiang H,
Vassilopoulos A, Spitz DR and Gius D: Exploring the electrostatic
repulsion model in the role of Sirt3 in directing MnSOD acetylation
status and enzymatic activity. Free Radic Biol Med. 53:828–833.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zeng L, Yang Y, Hu Y, Sun Y, Du Z, Xie Z,
Zhou T and Kong W: Age-related decrease in the mitochondrial
sirtuin deacetylase Sirt3 expression associated with ROS
accumulation in the auditory cortex of the mimetic aging rat model.
PLoS One. 9:e880192014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Li M, Chiu JF, Mossman BT and Fukagawa NK:
Down-regulation of manganese-superoxide dismutase through
phosphorylation of FOXO3a by Akt in explanted vascular smooth
muscle cells from old rats. J Biol Chem. 281:40429–40439. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Niska K, Pyszka K, Tukaj C, Wozniak M,
Radomski MW and Inkielewicz-Stepniak I: Titanium dioxide
nanoparticles enhance production of superoxide anion and alter the
antioxidant system in human osteoblast cells. Int J Nanomedicine.
10:1095–1107. 2015.PubMed/NCBI
|
|
54
|
Li Y, Wang W, Wu Q, Li Y, Tang M, Ye B and
Wang D: Molecular control of TiO2-NPs toxicity formation
at predicted environmental relevant concentrations by Mn-SODs
proteins. PLoS One. 7:e446882012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Bánréti A, Sass M and Graba Y: The
emerging role of acetylation in the regulation of autophagy.
Autophagy. 9:819–829. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Papanicolaou KN, O'Rourke B and Foster DB:
Metabolism leaves its mark on the powerhouse: Recent progress in
post-translational modifications of lysine in mitochondria. Front
Physiol. 5:3012014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Hebert AS, Dittenhafer-Reed KE, Yu W,
Bailey DJ, Selen ES, Boersma MD, Carson JJ, Tonelli M, Balloon AJ,
Higbee AJ, et al: Calorie restriction and SIRT3 trigger global
reprogramming of the mitochondrial protein acetylome. Mol Cell.
49:186–199. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Giralt A and Villarroya F: SIRT3, a
pivotal actor in mitochondrial functions: Metabolism, cell death
and aging. Biochem J. 444:1–10. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Kim H, Lee YD, Kim HJ, Lee ZH and Kim HH:
SOD2 and Sirt3 Control Osteoclastogenesis by Regulating
Mitochondrial ROS. J Bone Miner Res. 32:397–406. 2017. View Article : Google Scholar : PubMed/NCBI
|