Introduction
Tooth movement (TM) is a process in which mechanical
force induces alveolar bone resorption on the pressure side and
alveolar bone deposition on the tension side (1). As it usually takes ≥2 years to
complete orthodontic treatment, it is important that periodontal
accelerate osteogenesis orthodontics (PAOO) can increase
orthodontic TM and reduce the course of orthodontic treatment
(2). The clinical application of
PAOO was first introduced by the Wilcko brothers (a periodontist
and an orthodontist), and has become a useful modality in the field
of surgical orthodontics for the induction of faster TM (3). Traditional orthodontic therapy
focuses on applying forces to the teeth, whereas PAOO utilizes the
dynamics of bone physiology to enhance TM. PAOO involves controlled
surgical damage to cortical bone that accelerates bone metabolism
to aid orthodontic TM, resulting in TM that is 2–3 times faster
compared with traditional orthodontic therapy (4–6).
MicroRNAs (miRNAs/miRs), which are 22–25 nucleotides
in length, post-transcriptionally regulate gene expression in a
sequence-specific manner (7,8). A
previous study has revealed that certain miRNAs are critical
post-transcriptional modulators during osteoblastogenesis and
osteoclastogenesis (9).
Furthermore, previous studies have demonstrated that certain miRNAs
respond to mechanical stimuli in cultured human periodontal
ligament cells and periodontal ligament stem cells (10–12).
However, whether miRNAs regulate PAOO or mediate alveolar bone
remodeling in vivo is not completely understood.
It has been reported that miR-21 mediates
stretch-induced osteogenic differentiation of periodontal ligament
stem cells in vitro, which supports osteoclast
differentiation (10). Although
miR-21 is required for the regulation of gene expression under
mechanical force in several biological processes, such as bone
formation (10,13), its role during PAOO has not been
previously reported. Therefore, the present study aimed to
investigate the effects of miR-21 expression in vivo and
examine its functional roles in the regulation of osteogenesis and
osteoclast differentiation during PAOO.
Materials and methods
Study design
A total of 36 male Sprague-Dawley rats [weight,
286–326 g; age, 8 weeks; provided by Beijing Weitong Lihua
Experimental Animal Technology Co., Ltd.; certificate no. scxk
(Beijing) 2016-0011] were raised under specific pathogen-free
conditions. The experiment was performed under sterile conditions
with humidity 40–60%, 20–26°C and maximum daily temperature
difference of 4°C. The circadian rhythm was changed every 12 h.
Rats were randomly divided into four groups (n=9/group) as
presented in Table I. The present
study was approved by the Experimental Animal Welfare and Ethics
Committee of China Medical University (approval no. 16049R).
 | Table I.Group assignment. |
Table I.
Group assignment.
| Group | Force | Injection |
|---|
| TM | TM | Normal saline |
| PAOO | PAOO + TM | Normal saline |
| agomiR-21 | PAOO + TM | AgomiR-21 |
| antagomiR-21 | PAOO + TM | AntagomiR-21 |
Overexpression and inhibition of
miR-21 in rats
AgomiR-21 and antagomiR-21 were purchased from
Guangzhou RiboBio Co., Ltd., which had the following sequences:
agomiR-21 forward, 5′-UAGCUUAUCAGACUGAUGUUGA-3′ and reverse,
3′-AUCGAAUAGUCUGACUACAACU-5′; and antagomiR-21,
5′-UCAACAUCAGUCUGAUAAGCUA-3′. Rats were anesthetized by the
intraperitoneal injection of sodium pentobarbital (40 mg/kg). Prior
to orthodontic loading, rats received an injection of agomiR-21 (3
nmol/time; 20 µl) per day, antagomiR-21 (10 nmol/time; 20 µl) per
day or normal saline (20 µl) per day into the buccal, palatal and
mesial submucosa of the first maxillary molar continuously for 3
days.
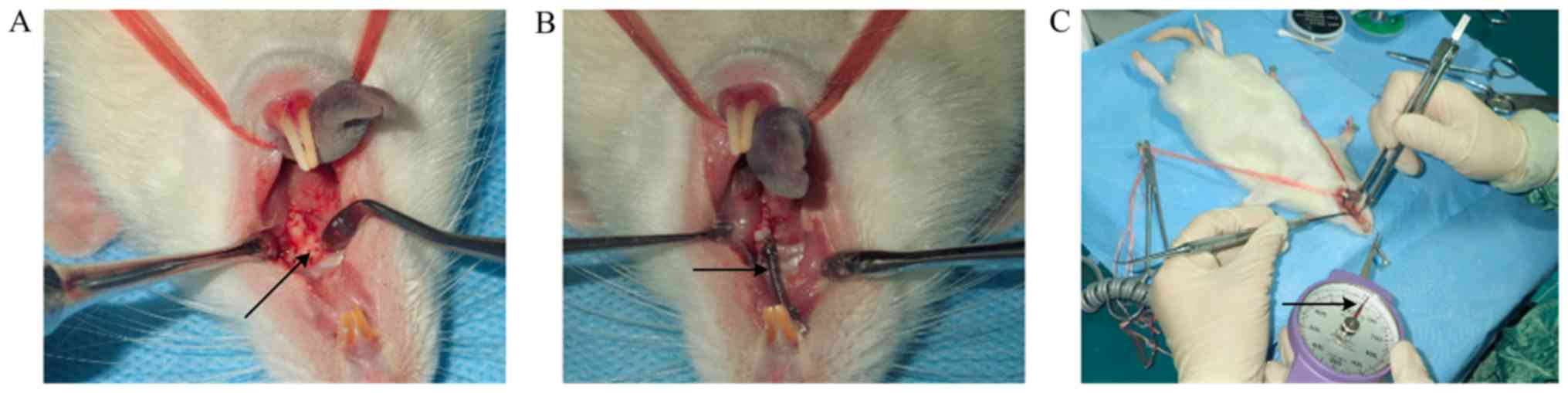
Orthodontic loading
Rats were anesthetized by the intraperitoneal
injection of sodium pentobarbital (40 mg/kg). Subsequently, two
upper central incisors were drilled horizontally with a 0.25-mm
orthodontic ligation wire to penetrate the root hole. The central
incisors were ligated together to enhance anchorage and prevent
continuous eruption.
Groups PAOO, agomiR-21 and antagomiR-21 were
prepared as follows: The labial, palatal and mesial alveolar mucosa
of the left maxillary first molar was incised to expose the
alveolar bone surface; and subsequently, a high-speed hand piece
was used under water irrigation to punch five perforations on the
buccal and palate sides, and two perforations on the mesial side
(diameter, 0.5 mm; depth; 0.5 mm). Group TM did not undergo the
perforation procedure.
To prevent the tension spring from falling off, a
retention groove was grinded into the buccal-palatal surface of the
neck of the left maxillary first molars and a ligation wire was
inserted into the retention groove of the first molar. The tension
spring was fixed between the first maxillary molar and the incisor,
and was stretched to pull the first maxillary molar to the mesial
position. The orthodontic force was maintained at 25 g using a
dynamometer. For the 3 days following surgery, the rats were
provided with soft food. The establishment of the PAOO model is
presented in Fig. 1.
The distance of TM was measured using an electronic
vernier caliper and a stereomicroscope (Motic Instruments). The
distance between the first maxillary molar and the upper incisor
was measured prior to surgery and following sacrifice to calculate
the total distance of TM. The measurements were conducted three
times in each rat.
At day 7 post-surgery, the rats were sacrificed by
carbon dioxide euthanasia, according to the guidelines (14). The flow rate of CO2 did
not displace >30% of the chamber volume/min. Death was verified
by monitoring cessation of breathing, heartbeat and deep pain in
response to the toe pinch method. The flow rate of CO2
was maintained for at least 1 min after cessation of breath, and
absence of heartbeat was assessed for at least 5 min. Subsequently,
the mucosal tissues were removed, and the cortical and cancellous
bones were collected for further analysis.
Tartrate-resistant acid phosphatase
(TRAP) staining
Alveolar bone tissue surrounding the maxillary first
molar were fixed for 48 h at 4°C in precooled 4% paraformaldehyde.
The decalcification was carried out in 10% EDTA decalcification
solution (pH 7.2–3.4). The solution was replaced every 2 days and
shaken. The decalcification time was ~3 months until the alveolar
bone could be penetrated without resistance under 4°C.
Subsequently, the tissue was embedded in paraffin and sectioned
into 5-µm thick occlusal serial sections. The sections were stained
using an Acid Phosphatase, Leukocyte (TRAP) kit (Sigma-Aldrich;
Merck KGaA) according to the manufacturer's protocol. Subsequently,
osteoblasts and TRAP+ multinucleated cells that were
attached to alveolar bone surfaces were counted. The BX53 Olympus
fluorescence microscope (Olympus Corporation) was used under
magnification, ×200.
Reverse transcription-quantitative PCR
(RT-qPCR)
Dentoalveolar bone tissue from the mesial alveolar
bone around the left first maxillary molar, including cortical and
cancellous bone tissue, was collected. Total RNA was extracted
using QIAzol Lysis Reagent (Qiagen, Inc.). Total RNA was reverse
transcribed into cDNA using the TaqMan Reverse Transcription kit
(Guangzhou RiboBio Co., Ltd.) according to the manufacturer's
protocol. Subsequently, qPCR was performed. QuantiFast®
SYBR® Green PCR Master Mix was used
(QuantiFast® SYBR® Green PCR kit; Beijing
Bulader Technology Development Co., Ltd.). The following
thermocycling conditions were used for qPCR: Initial denaturation
at 95°C for 5 min; followed by 40 cycles of 95°C for 10 sec and
60°C for 30 sec. mRNA and miRNA expression levels were normalized
to the internal reference genes GAPDH and U6, respectively. miRNA
expression levels were determined by performing a stem-loop RT-qPCR
assay, as previously described (15). The 2−∆∆Cq method was
used (16). The following primer
sequences were used: Programmed cell death 4 (PDCD4) forward,
5′-CTGTGTTTATGAGACTGTGGTT-3′ and reverse, 5′-CGCGACTTCGTTCGTATC-3′;
activin A receptor type 2B (ACVR2b) forward,
5′-TCTCGTACCTGCATGAGGA-3′ and reverse, 5′-TCGCTCTTCAGCAGAACA-3′;
C-fos forward, 5′-CAGCTCCCACCAGTGTCTA-3′ and reverse,
5′-CGCGTTGAAACCCGAGAA-3′; receptor activator of NF-κΒ ligand
(RANKL) forward, 5′-CGAAGACACAGAAGCACTAC-3′ and reverse,
5′-CACGAACCTTCCATCATAGC-3′; and GAPDH forward,
5′-GCGAGATCCCGCTAACATCA-3′ and reverse,
5′-CTCGTGGTTCACACCCATCA-3′.
Western blotting
Dentoalveolar bone tissues from the mesial alveolar
bone around the left first maxillary molar, including cortical and
cancellous bone tissues, were collected. The tissues were sonicated
in a lysis buffer (Shanghai HuaYi Biology Technology Co., Ltd.)
containing 1 mM PMSF. Total protein was extracted and quantified
using the bicinchoninic acid method. Total protein (50 µg/lane) was
separated via 10% SDS-PAGE and transferred onto nitrocellulose
membranes, which were blocked with 5% non-fat milk for 1 h at 37°C.
Subsequently, the membranes were incubated with primary antibodies
targeted against PDCD4 (1:2,000; cat. no. 12587-1-AP; ProteinTech
Group, Inc.), ACVR2b (1:1,000; cat. no. Ag16744; ProteinTech Group,
Inc.), RANKL (1:1,000; cat. no. 23408-1-AP; ProteinTech Group,
Inc.), C-Fos (1:1,000; cat. no. 15832-1-AP; ProteinTech Group,
Inc.) and GAPDH (1:5,000; cat. no. 10494-1-AP; Santa Cruz
Biotechnology, Inc.) at 4°C overnight. Subsequently, the membranes
were incubated with horseradish peroxidase-conjugated secondary
antibodies, goat anti rabbit IgG (1:5,000; cat. no. CW0156S;
Kangwei Century Biotechnology Co., Ltd.) at 37°C for 45 min.
Protein bands were visualized using Pierce ECL (Pierce; Thermo
Fisher Scientific, Inc. GADPH was used as the loading control. Gel
Pro analyzer software 4.5 (Beijing Zhongsheng Tiancheng Technology
Co., Ltd.) was used to analyze the optical density value of the
target strip.
Immunohistochemical staining
To reduce the error and maintain standards between
groups, the paraffin sections used for immunohistochemical staining
were taken from 1.2 mm away from the root tip. The sections were
fixed for 48 h at 4°C in precooled 4% paraformaldehyde.
Subsequently, 4-µm thick sagittal mouse maxillae were
deparaffinized, treated with 0.25% trypsin for 30 min at 37°C for
antigen retrieval and incubated with 3% hydrogen peroxide for 30
min at 37°C. The sections were blocked with 5% bovine serum albumin
(Wuhan Boster Biological Technology, Ltd.) in PBS for 2 h at room
temperature. Subsequently, the sections were incubated at 4°C
overnight with anti-PDCD4 (1:100; cat. no. 12587-1-AP; ProteinTech
Group, Inc.) and anti-C-Fos (1:100; cat. no. 15832-1-AP;
ProteinTech Group, Inc.) rabbit anti-mouse antibodies. Following
primary incubation, the sections were incubated with a horseradish
peroxidase-conjugated goat anti-rabbit secondary antibody (1:500;
cat. no. CW0103S; Kangwei Century Biotechnology Co., Ltd.) for 2 h
at room temperature. The sections were washed three times by PBS
for 3 min. The slides were stained with DAB for 4 min at 37°C and
counterstained with hematoxylin for 3 min at 37°C. Stained sections
were observed under a fluorescence microscope (N-STORM; Nikon
Corporation) in ≥3 fields of view under magnification ×200. The
number of positive-stained cells over the total periodontal area
was quantified using ImageJ software (version 1.47; National
Institutes of Health). PDCD4 and C-Fos protein levels were
semi-quantitatively detected as the integral optical density.
Cell culture
293T cells (American Type Culture Collection) were
cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.) containing
10% FBS (Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin and 50 mg/l streptomycin at 37°C with 5%
CO2.
Dual-luciferase reporter assay
Renilla luciferase activities was used as
control for normalization. PDCD4 was predicted as a potential
target of miR-21 by TargetScan7.2 (http://www.targetscan.org/vert_72/). The
3′-untranslated region (UTR) of PDCD4 was cloned into the pGL3
plasmid (Suzhou GenePharma Co., Ltd.). Subsequently, 293T cells at
50% density were seeded into 24-well plates and co-transfected with
PDCD4-3′UTR-wild-type (WT) or the mutant (MUT) sequence and miR-21
mimics or mimics control (20 nM; Suzhou GenePharma Co., Ltd.) using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.; 100 nM) according to the manufacturer's protocol.
Following incubation for 48 h at 37°C, luciferase activities were
determined using a Dual-Luciferase Reporter assay system (Promega
Corporation) according to the manufacturer's protocol. miR-21
mimics forward, 5′-UAGCUUAUCAGACUGAUGUUGAC-3′ and reverse,
3′-CAACAUCAGUCUGAUAAGCUAUU-5′; and mimics control forward,
5′-UUCUCCGAACGUGUCACG-3′ and reverse,
3′-ACGUGACACGUUCGGAGAATT-5′.
Statistical analysis
All statistical analyses were performed using SPSS
software (version 13.0; SPSS, Inc.). Data are presented as the mean
± SD. Differences among groups were analyzed using one-way ANOVA
followed by Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference. Each experiment
was repeated three times.
Results
TM in the rat model of PAOO
Following 7 days of treatment, the distance of TM in
group PAOO was significantly longer compared with groups TM and
antagomiR-21, but was significantly shorter compared with group
agomiR-21 (P<0.05; Table
II).
| Table II.Mean difference in tooth movement
between the four groups. |
Table II.
Mean difference in tooth movement
between the four groups.
| Group | Distance (mm) |
|---|
| TM |
0.112±0.013a |
| PAOO | 0.251±0.008 |
| agomiR-21 |
0.856±0.012a |
| antagomiR-21 |
0.072±0.010a |
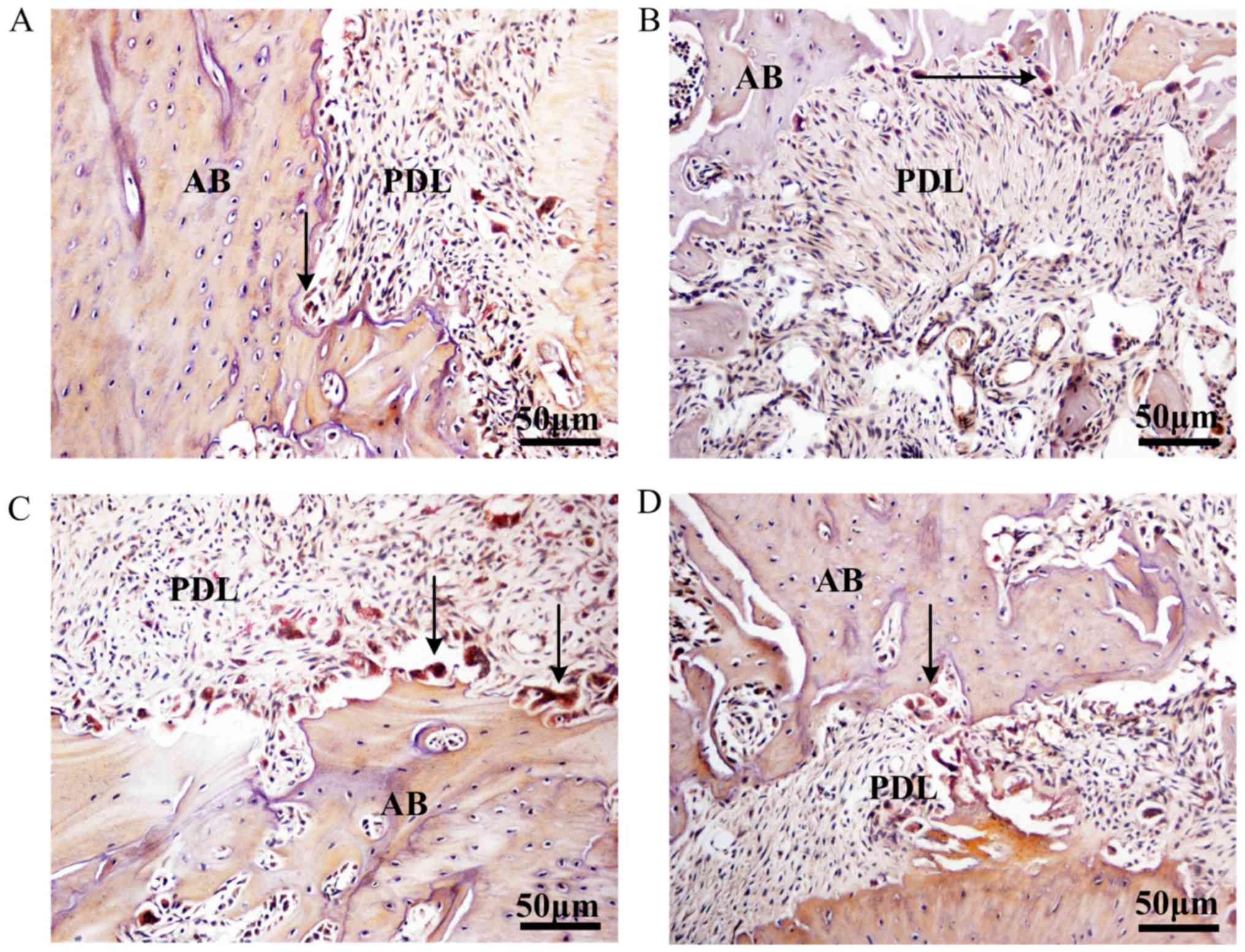
TRAP staining indicated that group PAOO displayed a
higher number of osteoclasts in the alveolar bone surrounding the
first molar compared with group TM (Fig. 2A and B). Following treatment with
agomiR-21, osteoclast activity was notably increased and a reduced
number of osteoclasts were observed in the tissues control compared
with group PAOO (Fig. 2C). The
osteoclast activity was not notably different in group antagomiR-21
control compared with group PAOO (Fig.
2D). The results indicated that the rat model of PAOO had been
successfully established.
miR-21 regulates osteoclastogenesis
during PAOO-facilitated TM
To assess the association between miR-21 and
osteoclastogenesis during PAOO, rats were treated with agomiR-21 or
antagomiR-21. To ensure agomir-21 and antagomir-21 penetrated the
cortical bone and functioned, the mucosal tissues were removed, and
the cortical and cancellous bones were collected. Subsequently, the
efficiency of agomiR-21 and antagomiR-21 in local dentoalveolar
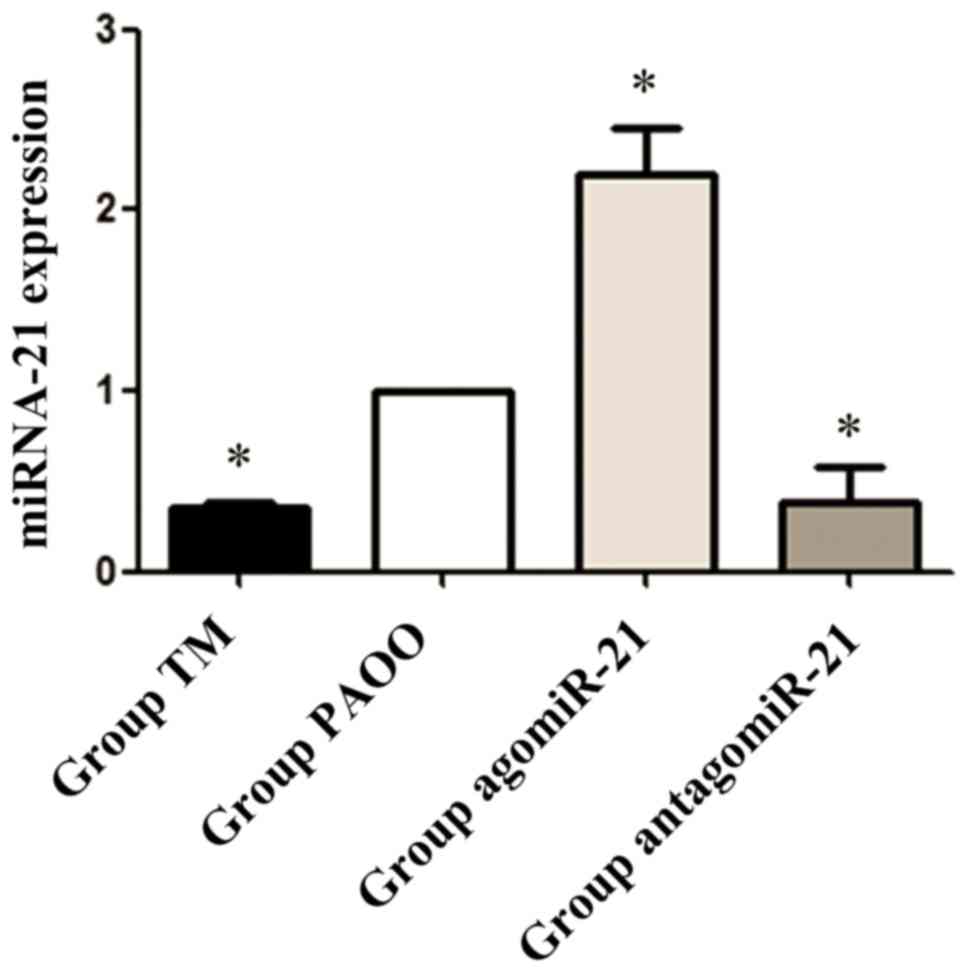
bone tissues was evaluated by RT-qPCR. Compared with group PAOO,
miR-21 expression levels in alveolar bone tissue were significantly
decreased in group TM (P<0.05). Furthermore, compared with group
PAOO, miR-21 expression levels were significantly increased in rats
treated with agomiR-21 (P<0.05), but were significantly
decreased in rats treated with antagomiR-21, which suggested that
agomiR-21 and antagomiR-21 upregulated and downregulated miR-21
expression, respectively, in vivo (P<0.05; Fig. 3).
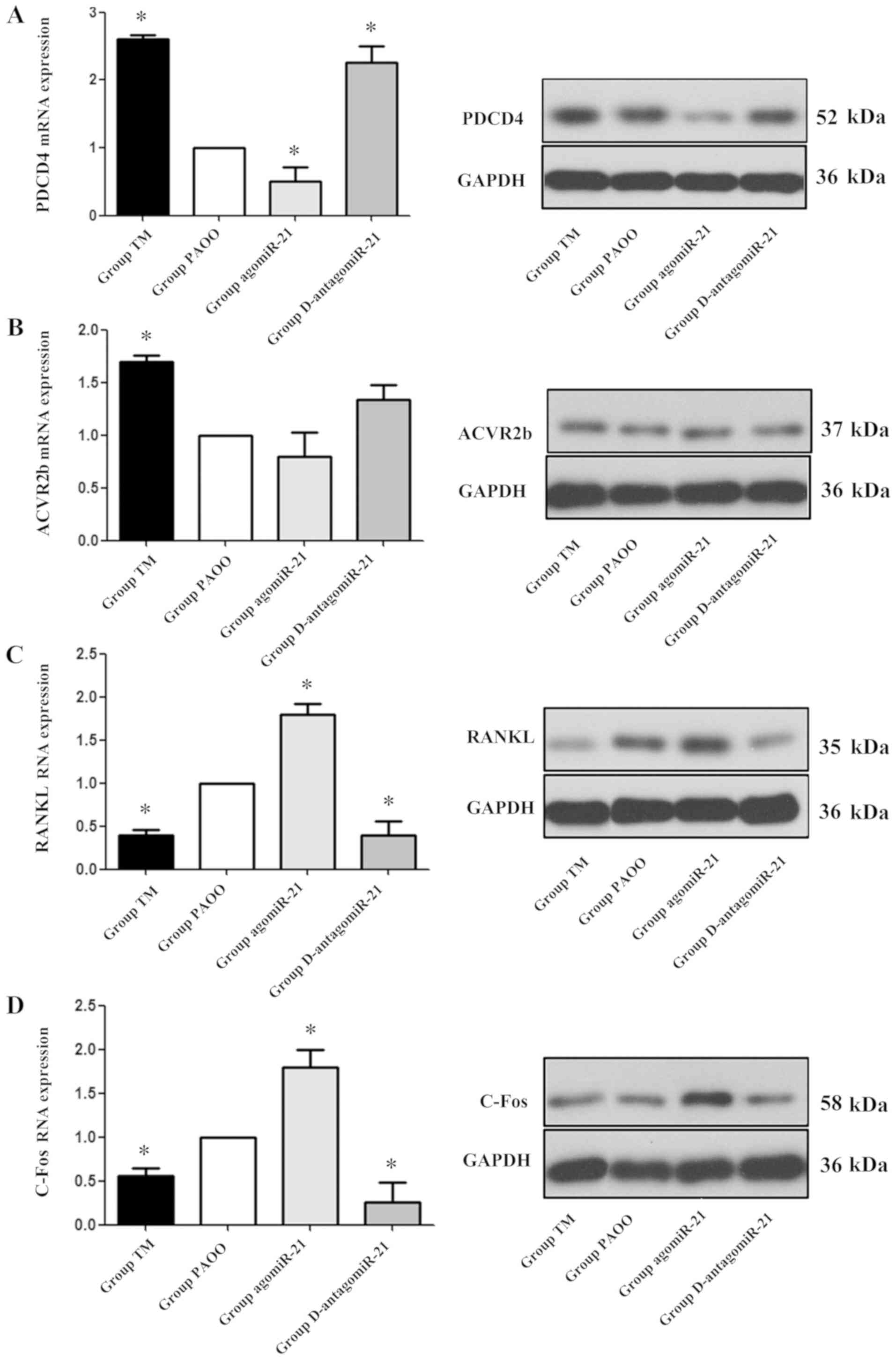
To further determine the role of miR-21 during PAOO,
the expression of two downstream target genes of miR-21, PDCD4 and
ACVR2b, was detected. Compared with group PAOO, rats treated with
agomiR-21 displayed decreased mRNA and protein expression levels of
PDCD4 notably. By contrast, groups TM and antagomiR-21 displayed
significantly increased mRNA and protein expression levels of PDCD4
compared with group PAOO (P<0.05; Fig. 4A). The mRNA and protein expression
levels of ACVR2b were significantly increased in group TM compared
with group PAOO (P<0.05); however, ACVR2b expression levels were
not significantly different between groups PAOO, agomiR-21 and
antagomiR-21(P>0.05; Fig.
4B).
RANKL is an important factor that can be used as an
indicator of osteoclast activity (17). C-Fos is a key regulator of
osteoclast differentiation that is a RANK-activating transcription
factor, which induces the expression of various osteoclast-specific
downstream target genes (18). To
further study the mechanism underlying orthodontic teeth movement,
the expression levels of RANKL and C-Fos were examined. The results
indicated that, compared with group PAOO, the mRNA and protein
expression levels of RANKL and C-Fos were significantly increased
in group agomiR-21, but were significantly decreased in groups TM
and antagomiR-21 (P<0.05; Fig. 4C
and D).
Immunohistochemical staining was used to further
examine the relationship between miR-21 and PDCD4 or C-Fos in rat
alveolar bone tissues. PDCD4 and C-Fos expression was primarily
localized in the cytoplasm and occasionally present in the cell
nuclei. The staining intensity of PDCD4 was significantly decreased
in group agomiR-21, and was significantly increased in groups TM
and antagomiR-21, compared with group PAOO. By contrast, C-Fos
staining displayed the opposite trend (P<0.05; Fig. 5).
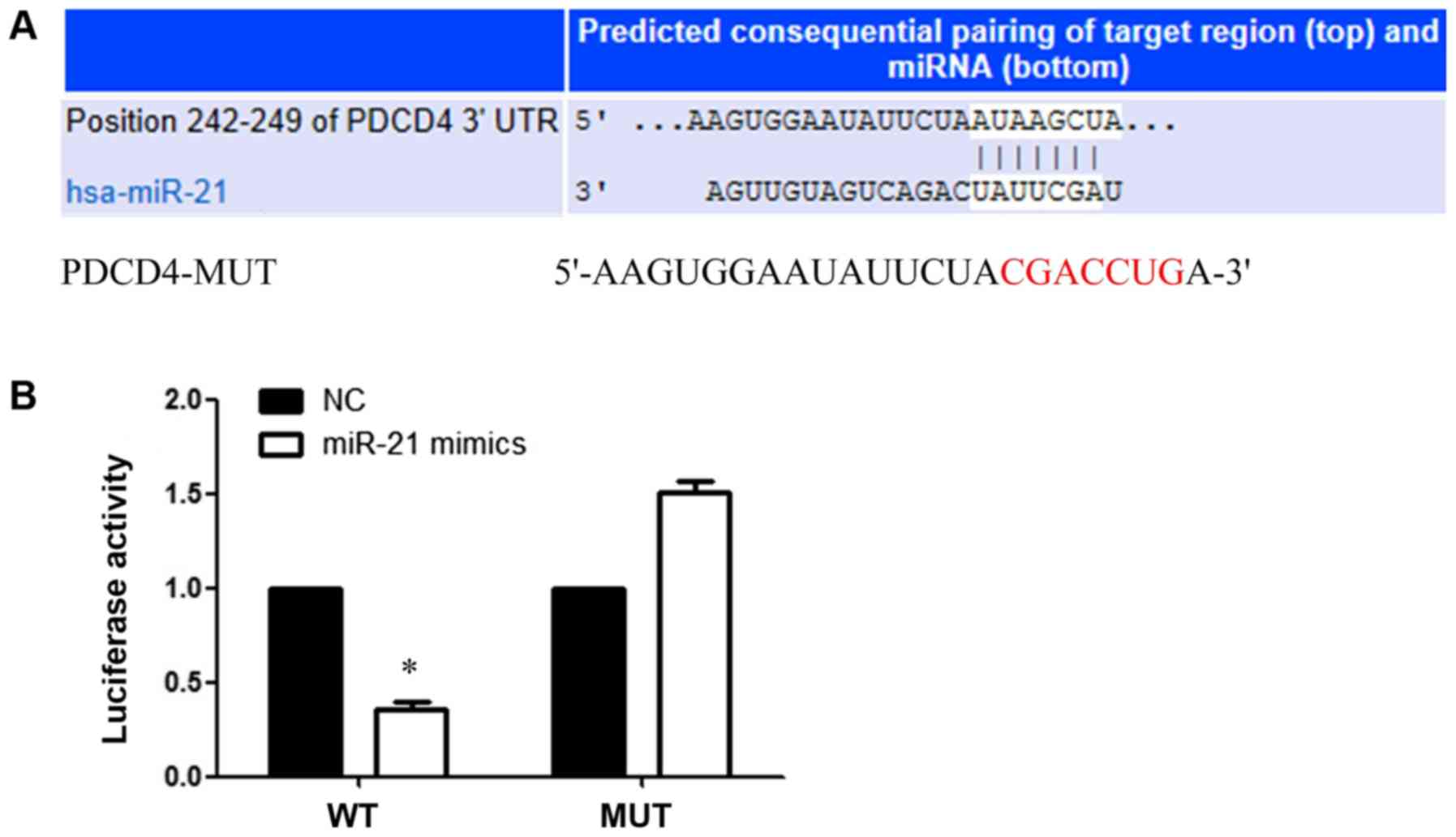
PDCD4 is a direct target of
miR-21
TargetScan was used to predict the binding site
between miR-21 and the 3′UTR of PDCD4 (Fig. 6A). miR-21 overexpression
significantly inhibited the luciferase activities of the
PDCD4-3′UTR-WT group compared with the NC group. However, miR-21
overexpression did not significantly alter the luciferase
activities of the PDCD4-MUT group compared with the NC group, which
indicated that miR-21 specifically targeted the 3′UTR of PDCD4 by
binding to the predicted sequence (P<0.05; Fig. 6B).
Discussion
PAOO is a clinical procedure that combines selective
alveolar corticotomy, particulate bone grafting and the application
of orthodontic forces, and is theoretically based on the
bone-healing pattern that is known as the regional acceleratory
phenomenon (19). PAOO results in
increased alveolar bone width, decreased treatment time, increased
post-treatment stability and reduced apical root resorption
(20,21). To investigate the specific
mechanism underlying the clinical procedure, a rat model of PAOO
was established in the present study. To validate the rat model,
morphological analyses were performed and necessary measurements
were taken. Compared with group TM, group PAOO displayed an
increased rate of TM, a significant reduction in bone volume and
signs of bone resorption, including a higher number of osteoclast
cells at 7 days post-surgery.
miRNAs modulate gene expression by binding to target
mRNAs, thereby inhibiting their translation or promoting their
degradation (22). In a previous
study, gene chip technology indicated that miR-21 expression was
increased in TM (23). A potential
mechanism underlying miR-21 in TM is that the protein that is
post-transcriptionally controlled by miR-21 inhibits
osteoclastogenesis (23). Although
several targets of miR-21 have been identified by bioinformatics
analysis, PDCD4 and ACVR2b were the primary genes that were
investigated in the present study due to their ability to regulate
osteogenic and osteoclastic differentiation (10,24).
High miR-21 expression levels are required for PDCD4 downregulation
during osteoclastogenesis. PDCD4 suppresses cap-dependent
translation of mRNAs with highly structured 5-regions via
interaction with the eukaryotic translation initiation factor 4A
helicase (25). It has also been
reported that PDCD4 can directly alter the activity of the
transcription factor activator protein-1 (AP-1), which triggers the
transcription ofmiR-21 (26).
Several conserved enhancer elements, including the binding site for
AP-1, have been identified in the miR-21 promoter region (27). PDCD4 also regulates C-Fos, which is
a transcription factor associated with osteoclast production
(28,29). As demonstrated by the development
of osteopetrosis in mice lacking C-Fos, C-Fos serves an important
role during osteoclastogenesis. Therefore, although C-Fos is not
required for normal osteoprogenitor development, it is essential
for osteoclastogenesis (30,31).
Moreover, C-Fos is the principal contributor among PDCD4-induced
AP-1 components (32).
In the present study, PDCD4 expression levels were
significantly decreased in the agomiR-21 group compared with group
PAOO, whereas the antagomir-21 group displayed significantly
increased PDCD4 expression levels compared with group PAOO. The
results suggested that miR-21 negatively regulated PDCD4 expression
during PAOO-mediated TM. Furthermore, the results indicated that
C-Fos expression levels increased with agomiR-21 treatment and
decreased with antagomiR-21 treatment, as has been reported in
previous studies (33).
ACVR2b is a transmembrane serine/threonine receptor
kinase that serves a crucial role during the activation of activin,
which forms part of the transforming growth factor-β signaling
pathway, and acts on cell proliferation and differentiation, as
well as several other biological functions (34). Wei et al (10) demonstrated that miR-21 mediated the
osteogenic differentiation effect of stretch by directly targeting
ACVR2b, which is a key regulator of osteogenic differentiation. In
addition, ACVR2b gain- and loss-of-function experiments indicated
an association between miR-21 and ACVR2b during stretch-induced
periodontal ligament stem cells osteogenic differentiation
(10). Moreover, it has been
reported that ACVR2b-mediated effects on osteoblasts are due to its
interaction with bone morphogenetic protein (35). In the present study, the results
indicated that the expression levels of ACVR2b were not
significantly altered by miR-21 overexpression or knockdown, which
suggested that miR-21 did not serve a biological role via ACVR2b
during PAOO-mediated TM.
Several studies have investigated the role of miR-21
during orthodontic TM (10,36);
however, to the best of our knowledge, the present study was the
first to investigate the role of miR-21 during PAOO.
Dual-luciferase reporter assays were performed to verify that
miR-21 downregulated PDCD4 expression levels by targeting the 3′UTR
of PDCD4. Although the present study investigated the role of
miR-21 during PAOO, there were a number of limitations. First, the
present study did not perform RNAscope or RNA-fluorescence in
situ hybridization experiments. Second, the mechanisms
underlying miR-21-mediated regulation of PDCD4, ACVR2b, RANKL and
C-Fos expression in vitro were not investigated.
Furthermore, due to the absence of in vitro experiments, the
dose- and time-dependent relationship between miR-21 and PDCD4,
ACVR2b, RANKL and C-Fos was not investigated; therefore, further
investigation is required.
In conclusion, the present study suggested a
potential mechanism underlying PAOO-mediated acceleration of
orthodontic TM. miR-21 overexpression negatively regulated the
expression levels of the target gene PDCD4, leading to increased
C-Fos expression levels, enhanced RANKL-mediated osteoclast
generation, remodeling of the alveolar bone, development of
temporary osteopenia and local alveolar osteoporosis, and increased
movement of orthodontic teeth.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Department of
Education Science and Technology Program of Liaoning Province
(grant no. LK201639).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YuZ, YT and YaZ conceived and designed the
experiments. XY, ZZ and CF collected and analyzed the imaging and
pathology data. YuZ wrote the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Experimental
Animal Welfare and Ethics Committee of China Medical University
(approval no. 16049R).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Van Schepdael A, Vander Sloten J and Geris
L: A mechanobiological model of orthodontic tooth movement. Biomech
Model Mechanobiol. 12:249–265. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Soltani L, Loomer PM and Chaar EE: A novel
approach in periodontally accelerated osteogenic orthodontics
(PAOO): A case report. Clin Adv Periodontics. 9:110–114. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wilcko WM and Wilcko MT: Accelerating
tooth movement: The case for corticotomy-induced orthodontics. Am J
Orthod Dentofacial Orthop. 114:4–12. 2013. View Article : Google Scholar
|
|
4
|
Adusumilli S, Yalamanchi L and
Yalamanchili PS: Periodontally accelerated osteogenic orthodontics:
An interdisciplinary approach for faster orthodontic therapy. J
Pharm Bioallied Sci. 6 (Suppl 1):S2–S5. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ferguson DJ, Al-Harbi MS, Wilcko WM,
Wilcko MT and Erie P: Lower dental arch decrowding comparing
non-extraction accelerated osteogenesis and distraction techniques.
J Dent Res. 80:181–183. 2001.
|
|
6
|
Yu H, Jiao F, Wang B and Shen SG:
Piezoelectric decortication applied in periodontally accelerated
osteogenic orthodontics. J Craniofac Surg. 24:1750–1752. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bartel D: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lian JB, Stein GS, van Wijnen AJ, Stein
JL, Hassan MQ, Gaur T and Zhang Y: MicroRNA control of bone
formation and homeostasis. Nat Rev Endocrinol. 8:212–227. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wei F, Liu D, Feng C, Zhang F, Yang S, Hu
Y, Ding G and Wang S: microRNA-21 mediates stretch-induced
osteogenic differentiation in human periodontal ligament stem
cells. Stem Cells Dev. 24:312–319. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu Y, Ou Y, Liao C, Liang S and Wang Y:
High-throughput sequencing analysis of the expression profile of
microRNAs and target genes in mechanical force-induced
osteoblastic/cementoblastic differentiation of human periodontal
ligament cells. Am J Transl Res. 11:3398–3411. 2019.PubMed/NCBI
|
|
12
|
Yao S, Zhao W, Ou Q, Liang L, Lin X and
Wang Y: MicroRNA-214 suppresses osteogenic differentiation of human
periodontal ligament stem cells by targeting ATF4. Stem Cells Int.
2017:30286472017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ramanujam D, Sassi Y, Laggerbauer B and
Engelhardt S: Viral vector-based targeting of miR-21 in cardiac
nonmyocyte cells reduces pathologic remodeling of the heart. Mol
Ther. 24:1939–1948. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
NIH ARAC guidelines for euthanasia of
rodents using carbon dioxide. NIH Office of Intramural Research and
Office of Animal Care and Use, . 2017.https://oacu.oir.nih.gov/oacu-staff
|
|
15
|
Feng J, Wang K, Liu X, Chen S and Chen J:
The quantification of tomato microRNAs response to viral infection
by stem-loop real-time RT-PCR. Gene. 437:14–21. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Takegahara N, Kim H, Mizuno H,
Sakaue-Sawano A, Miyawaki A, Tomura M, Kanagawa O, Ishii M and Choi
Y: Involvement of receptor activator of nuclear factor-κB ligand
(RANKL)-induced incomplete cytokinesis in the polyploidization of
osteoclasts. J Biol Chem. 291:3439–3454. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Izawa T, Arakaki R, Mori H, Tsunematsu T,
Kudo Y, Tanaka E and Ishimaru N: The nuclear receptor AhR controls
bone homeostasis by regulating osteoclast differentiation via the
RANK/c-Fos signaling axis. J Immunol. 197:4639–4650. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Amit G, Jps K, Pankaj B, Suchinder S and
Parul B: Periodontally accelerated osteogenic orthodontics (PAOO)-a
review. J Clin Exp Dent. 4:e292–e296. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ma Z, Zheng J, Yang C, Xie Q, Liu X and
Abdelrehem A: A new modified bone grafting technique for
periodontally accelerated osteogenic orthodontics. Medicine
(Baltimore). 97:e120472018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hou HY, Li CH, Chen MC, Lin PY, Liu WC,
Cathy Tsai YW and Huang RY: A novel 3D-printed computer-assisted
piezocision guide for surgically facilitated orthodontics. Am J
Orthod Dentofacial Orthop. 155:584–591. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mRNAs are conserved targets of microRNAs. Genome Res.
19:92–105. 2019. View Article : Google Scholar
|
|
23
|
Pitari MR, Rossi M, Amodio N, Botta C,
Morelli E, Federico C, Gullà A, Caracciolo D, Di Martino MT,
Arbitrio M, et al: Inhibition of miR-21 restores RANKL/OPG ratio in
multiple myeloma-derived bonemarrow stromal cells and impairs the
resorbing activity of mature osteoclasts. Oncotarget.
6:27343–27358. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Frankel B, Christoffersen NR, Jacobsen A,
Lindow M, Krogh A and Lund AH: Programmed cell death 4 (PDCD4) is
an important functional target of the microRNA miR-21 in breast
cancer cells. J Biol Chem. 283:1026–1033. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yasuda M, Nishizawa T, Ohigashi H, Tanaka
T, Hou DX, Colburn NH and Murakami A: Linoleic acid metabolite
suppresses skin inflammation and tumor promotion in mice: Possible
roles of programmed cell death 4 induction. Carcinogenesis.
30:1209–1216. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Loh PG, Yang HS, Walsh MA, Wang Q, Wang X,
Cheng Z, Liu D and Song H: Structural basis for translational
inhibition by the tumour suppressor Pdcd4. EMBO J. 28:274–285.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fujita S, Ito T, Mizutani T, Minoguchi S,
Yamamichi N, Sakurai K and Iba H: miR-21 Gene expression triggered
by AP-1 is sustained through a double-negative feedback mechanism.
J Mol Biol. 378:492–504. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Miao W, Gao H and Hou X: Magnesium
lithospermate B inhibits titanium particles-induced osteoclast
formation by c-fos and inhibiting NFATc1 expression. Connect Tissue
Res. 60:487–494. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Talotta F, Cimmino A, Matarazzo MR,
Casalino L, De Vita G, D'Esposito M, Di Lauro R and Verde P: An
autoregulatory loop mediated by miR-21 and PDCD4 controls the AP-1
activity in RAS transformation. Oncogene. 28:73–84. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Friedman AD: Transcriptional control of
granulocyte and monocyte development. Oncogene. 26:6816–6828. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huhe M, Liu S, Zhang Y, Zhang Z and Chen
Z: Expression levels of transcription factors c-Fos and c-Jun and
transmembrane protein HAb18G/CD147 in urothelial carcinoma of the
bladder. Mol Med Rep. 15:2991–3000. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Haubrock M, Hartmann F and Wingender E:
NF-Y binding site architecture defines a C-Fos targeted promoter
class. PLoS One. 11:e01608032016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sugatani T, Vacher J and Hruska KA: A
microRNA expression signature of osteoclastogenesis. Blood.
117:3648–3657. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shi Y and Massagué J: Mechanisms of
TGF-beta signaling from cell membrane to the nucleus. Cell.
113:685–700. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kokabu S, Gamer L, Cox K, Lowery J, Tsuji
K, Raz R, Economides A, Katagiri T and Rosen V: BMP3 suppresses
osteoblast differentiation of bone marrow stromal cells via
interaction with Acvr2b. Mol Endocrinol. 26:87–94. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen N, Sui BD, Hu CH, Cao J, Zheng CX,
Hou R, Yang ZK, Zhao P, Chen Q, Yang QJ, et al: MicroRNA-21
contributes to orthodontic tooth movement. J Dent Res.
95:1425–1433. 2016. View Article : Google Scholar : PubMed/NCBI
|