Introduction
Myocardial ischemia/reperfusion injury (MI/RI)
refers to rapid tissue damage caused by the recovery of blood flow
from myocardial cells following ischemia (1). MI/RI is a common complication during
the perioperative period of cardiac and macrovascular surgeries
that can lead to the aggravation of myocardial damage and seriously
affect the recovery of postoperative cardiac function (2). Cardiomyocyte hypoxia/reoxygenation
(H/R) injury can be used to mimic myocardial ischemia/reperfusion
in vitro (3,4). Morphine is a widely used analgesic in
cardiac and macrovascular anesthesia (5,6).
Compared with ischemic pre-conditioning, morphine could be
conveniently and easily administered and cause less trauma to
patients (7). Previous studies
demonstrated that morphine pre-conditioning (MPC) significantly
reduced myocardial ischemic injury and inhibited cardiomyocyte
apoptosis, although the mechanism remains unclear (8,9).
MicroRNAs (miRNAs/miRs) are a class of endogenous,
single-stranded, non-coding RNAs that regulate multiple biological
processes such as cell proliferation, differentiation and apoptosis
(10). Cardiomyocyte apoptosis is
an important characteristic of MI/RI (11). Accumulating evidence indicates that
miRNAs can regulate apoptosis of cardiomyocytes through their
target genes and downstream signaling pathways (12). For example, Dong et al
(13) suggested that miR-21 could
inhibit ischemia-induced apoptosis by studying acute myocardial
infarction in rats. Cheng et al (14) demonstrated that miR-21 also
protected cardiomyocytes from hydrogen peroxide-induced damage by
regulating programmed cell death 4. Moreover, miR-320 was also
implicated in the regulation of myocardial ischemia/reperfusion
injury, and miR-320 upregulation could promote cardiomyocyte
apoptosis (15). Additionally,
miR-320 has multiple functions in different environments (16), and its expression level is closely
related to tumor migration and invasion. Indeed, overexpression of
miR-320 is associated with high risk of metastasis and poor
prognosis (17).
Previous studies demonstrated that 5′-adenosine
monophosphate-activated protein kinase (AMPK) could ameliorate
myocardial ischemia through the regulation of oxidative stress
(18,19), autophagy (20,21)
and apoptosis (22) in
cardiomyocytes. Moreover, AMPK exerts its protective effects with
other molecules that may have crosstalk with each other against
myocardial ischemia, for example, mesenchymal stem cell
(MSC)-derived exosomes could reduce MI/RI by inducing cardiomyocyte
autophagy via AMPK/mTOR and Akt/mTOR pathways (21,23,24).
Sun et al (25)
demonstrated that dexmedetomidine protected mice against MI/RI by
activating the AMPK/phosphatidylinositol-4,5-bisphosphate 3-kinase
(PI3K)/Akt/endothelial nitric oxide synthase pathway. PI3K/Akt
signaling is an important signaling pathway associated with many
diseases, including cancer and neurological diseases (26). Akt kinase consists of three
subtypes, Aktl, Akt2 and Akt3, which are the key factors downstream
of the PI3K signaling pathway that regulate cell proliferation,
apoptosis and metastasis (27).
Previous studies indicated that activation of the PI3K-Akt
signaling pathway protected the heart from reperfusion injury
(28). However, whether morphine
pre-conditioning participates in myocardial H/R injury through the
regulation of miR-320-3p and the PI3K/Akt signaling pathway is not
fully understood. Therefore, the current study examined the
myocardial protection mechanism of MPC following at the miRNA
level, and aimed to elucidate the relationship between MPC,
miR-320-3p and the PI3K/Akt signaling pathway in the regulation of
H/R injury of cardiomyocytes.
Materials and methods
Cell culture
H9c2 cells (cat. no. CRL-1446) were obtained from
the American Type Culture Collection and divided into four groups:
i) Control, ii) MPC, iii) H/R, and iv) MPC+H/R. Cells from the
control group were cultured in DMEM/F-12 containing 10% FBS, and 1%
penicillin/streptomycin at 37°C with 5% CO2 (all from
Thermo Fisher Scientific, Inc.). After one day of culture, the
original medium was discarded, and the cells were washed once or
twice with PBS, then resuspended in 5 ml fresh DMEM/F-12 medium and
returned to the CO2 incubator. The cells were passaged
when grown to 80 to 90% confluence. In the MPC group, cells were
cultured normally following a 10-min treatment with 1 µM morphine
(cat. no. 121206-2; Northeast Pharmaceutical Group Co., Ltd.). In
the H/R group, DMEM/F12 medium containing 10% serum was replaced by
anoxic solution (i.e., glucose-free and serum-free medium
containing 2.3 mM CaCl2, 5.6 mM KCl, 154 mM NaCl, 5 mM
Hepes, and 3.6 mM NaHCO3; pH 7.4). The culture plate was
maintained in anoxic chamber for 5 h at 37°C, and then
reoxygenation for 1 h in DMEM/F12 medium containing 10% serum under
a humidified atmosphere (95% air and 5% CO2) at 37°C.
For the MPC + H/R group, the cells were treated with 1 µM morphine
in serum-free DMEM/F12 for 10 min. Following the treatment,
serum-free DMEM/F12 was replaced by DMEM/F12 medium containing 10%
serum and the MPC pre-treatment was completed after 30 min of
normal culture. Cells then received the same treatment as the H/R
group.
Cell transfection
Cells were cultured for 12–16 h to 60–70%
confluence, and then transfected with Lipofectamine®
3000 (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. miR-320-3p mimic (miR10000903-1-5;
5′-AAAAGCUGGGUUGAGAGGGCGA-3′), mimic negative control
(miRB160401025525-2-1; 5′-UUUGUACUACACAAAAGUACUG-3′), and
miR-320-3p inhibitor (miR20000903-1-5;
5′-UCGCCCUCUCAACCCGCUUUU-3′), and inhibitor negative control
(miR2N0000001-1-5; 5′-CAGUACUUUUGUGUAGUACAAA-3′) were purchased
from Guangzhou RiboBio Co., Ltd. Briefly, when cells were at 60 to
70% confluence, the transfection was performed using
Lipofectamine® 3000 (Thermo Fisher Scientific, Inc.).
miR-320-3p mimic, inhibitor, corresponding control and
Lipofectamine® RNAiMAX (Thermo Fisher Scientific, Inc.)
were diluted in Opti-MEM medium (Thermo Fisher Scientific, Inc.).
Then, the diluted RNAs were added into the diluted
Lipofectamine® reagent and incubated for 10 min at room
temperature. The mixture containing the RNA was added to the cells.
Following an 8-h incubation in the presence of transfection
reagent, the medium was replaced, and the cells incubated for 2–4
days at 37°C prior to subsequent experimentation.
Bioinformatics prediction
TargetScan7.2. (http://www.targetscan.org) was used for the biological
prediction of miR-320-3p target genes.
Dual luciferase activity assay
The sequence of Akt3-3′-UTR was cloned into the
pmirGLO luciferase vector (cat. no. E1330; Promega Corp.). H9c2
cells were transfected with miR-320-3p mimic to detect the binding
of Akt3 and miR-320-3p. Cells transfected with miR-320-3p mimic
negative control were used as the blank group. The cell
transfection was performed by transfected with
Lipofectamine® 3000 (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. Luciferase activity was
determined using the Double-Luciferase Reporter Assay kit (cat. no.
FR201-01; TransGen Biotech) according to the manufacturer's
protocol. Briefly, the cell culture medium was removed, and the
cells were carefully washed twice in PBS, then added to 20 µl 1X
Cell Lysis Buffer from the kit. The cells were lysed for 10 min at
room temperature and centrifuged at 12,000 × g for 10 min at 4°C to
collect supernatant for later use. After centrifugation, 100 µl
Luciferase Reaction Reagent was added to the tube at room
temperature, and 20 µl cell lysate was carefully pipetted into the
tube and mixed gently to measure the activity of the firefly
luciferase reporter gene in a luminometer (SpectraMaxL, Molecular
Devices, LLC). Finally, 100 µl Luciferase Reaction Reagent II was
added to the above reaction tube at room temperature, vortexed, and
the activity of the Renilla luciferase reporter gene was
detected using a SpectraMaxL luminometer (Molecular Devices,
LLC).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA extraction from lysed H9c2 cells was
carried out at 4°C using TRIzol® (Invitrogen; Thermo
Fisher Scientific, Inc.), and all the consumables and reagents used
in the extraction process were subjected to DEPC treatment. The RT
of miRNA and mRNA was conducted using TaqMan MicroRNA Reverse
Transcription kit (cat. no. 4366597; Thermo Fisher Scientific,
Inc.) and TaqMan Reverse Transcription Reagents (cat. no. N8080234;
Thermo Fisher Scientific, Inc.). GAPDH served as internal control
for mRNA, while U6 served as internal control for miRNA
quantification. The primer sequences were as follows:
miR-320-3p-forward (F), 5′-TAAGTGCTTCCATGTTTTGGTGA-3′;
miR-320-3p-reverse (R), 5′-GAACATGTCTGCGTATCTCAGACTTC-3′; Akt3-F,
5′-TCCCCCGAACACTCTCTTCA-3′; Akt3-R, 5′-CCCTCCACCAAGGCGTTTAT-3′;
GAPDH-F, 5′-ATGACTCTACCCACGGCAAG-3′; GAPDH-R,
5′-GGAAGATGGTGATGGGTTTC-3′; U6-F, 5′-CTCGCTTCGGCAGCACA-3′; U6-R,
5′-AACGCTTCACGAATTTGCGT-3′. RT-qPCR was performed using the ABI7500
Real-Time PCR System (Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions. The thermocycling conditions were
set as follows: Initial denaturation at 95°C for 10 min, followed
by 45 cycles at 95°C for 30 sec, 59°C for 1 min, and finally at
60°C for 1 min, and preservation at 4°C. The relative expression
levels were calculated using the 2−ΔΔCq method (29).
MTT assay
Cell viability was detected using
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (cat.
no. M6494; Thermo Fisher Scientific, Inc.). H9c2 cells were seeded
in 96-well plates at a density of 5×104/ml and incubated
for 24, 48 and 72 h at 37°C. A volume of 10 µl MTT reagent was
added into each well and cells were then cultured for an additional
4 h at 37°C. After removing the MTT, 100 µl DMSO was added.
Finally, the optical density was measured at 490 nm using a
microplate reader (cat. no. 24072800; Thermo Fisher Scientific,
Inc.). Cells were seeded in triplicate wells and the experiment was
repeated three times.
Lactate dehydrogenase (LDH) content
determination
LDH content was measured using the Cytotoxicity LDH
Assay Kit-WST (cat. no. CK12; Dojindo Molecular Technologies,
Inc.), according to the manufacturer's protocol. Briefly, the cell
suspension was diluted in 50 µl medium, added to the 96-well plate
with 50 µl medium and incubated in a CO2 incubator at
37°C for 24 h. A volume of 10 µl Lysis Buffer was added into the
control well and the reaction was incubated at 37°C with 5%
CO2 for 30 min. After adding 100 µl Working Solution
into each well, the cells were incubated in the dark at room
temperature for 30 min. Next, 50 µl Stop Solution was added to
terminate the reaction, and the absorbance at 490 nm was determined
immediately using a microplate reader (cat. no. 24072800; Thermo
Fisher Scientific, Inc.).
Flow cytometry
A total of 1×106 H9c2 cells were
harvested then centrifuged at 1,200 × g for 1 min at 4°C, then
resuspended in 300 µl pre-chilled PBS containing 10% calf serum.
Cells were then fixed in 700 µl absolute ethanol at −20°C for
>24 h, and centrifuged at 140 × g for 30 sec at 4°C. Fixed cells
were resuspended in 500 µl cold PBS, and centrifuged at 1,200 × g
for 1 min at 4°C. After discarding the supernatant, the pelleted
cells were suspended in 480 µl cold PBS and 10 µl of 0.1 mg/ml
RNase A, and allowed to stand at 37°C for 30 min. Annexin V-FITC
and propidium iodide (PI) staining solution (5 µl each; cat. no.
APOAF; Sigma-Aldrich; Merck KGaA) were added to the cell suspension
for 15 min at room temperature in the dark. After the incubation,
cells were filtered once through a 400-mesh sieve, and apoptosis
was detected using a BD FACS Canto™ flow cytometer. Analysis was
carried out using FlowJo version 10.0 (FlowJo, LLC). The lower left
quadrant (FITC-H−/PI−) represents healthy
living cells, while the lower right quadrant
(FITC-H+/PI−) represents early apoptotic
cells. Late apoptotic cells are gated in the upper right quadrant
(FITC-H+/PI+) and necrotic cells are found in
the upper left quadrant (FITC-H−/PI+).
Western blot analysis
Total protein was extracted from H9c2 cells using
RIPA buffer (cat. no. 89901; Thermo Fisher Scientific, Inc.)
containing protease inhibitor and phosphatase inhibitor. The
protein concentrations were measured using Pierce Rapid Gold BCA
Protein Assay kit (cat. no. A53227; Thermo Fisher Scientific,
Inc.). 30 µg protein was separated by SDS-PAGE on 10% gels, then
transferred to PVDF membranes (cat. no. HVLP04700; EMD Millipore).
The films were fully rinsed in TBS, and then transferred to a dish
containing 5% skim milk solution and blocked at 37°C for 1 h, then
shaken for 2 h at room temperature on a decolorizing shaker. The
membranes were then incubated with rabbit primary antibody against
Akt3 (72 kDa; 1:2,000; cat. no. ab152157; Abcam) or mouse
anti-GAPDH (36 kDa; 1:20,000; cat. no. ab8245; Abcam) overnight at
4°C. The membrane was then incubated with an HRP-conjugated
secondary antibody (1:5,000; cat. nos. ab205718 and ab205719;
Abcam) for 2 h at room temperature. Finally, SignalFire
electrochemiluminescence reagent (cat. no. 6883; Cell Signaling
Technology, Inc.) was used for visualization. The ImageJ version
5.0 software (Bio-Rad Laboratories, Inc.) for quantification. GAPDH
was used as the reference protein.
Statistical analysis
Data are presented as the mean ± SD. Multi-group
comparisons were conducted using one-way ANOVA, followed by Tukey's
post hoc test. Statistical analysis was carried out using GraphPad
Prism 7.0 (GraphPad Software, Inc.). P<0.05 was considered to
indicate a statistically significant difference. All experiments
were performed in triplicate.
Results
MPC protects rat cardiomyocytes from
H/R damage
To investigate the effects of MPC on reperfusion
injury in rat cardiomyocytes, H9c2 cells were divided into Control,
MPC, H/R, MPC+H/R groups. An MTT assay demonstrated that the
viability of cells in the H/R group was reduced, compared with the
control. However, cell viability in the MPC+H/R group was
significantly higher than that of the H/R group (P<0.05;
Fig. 1A).
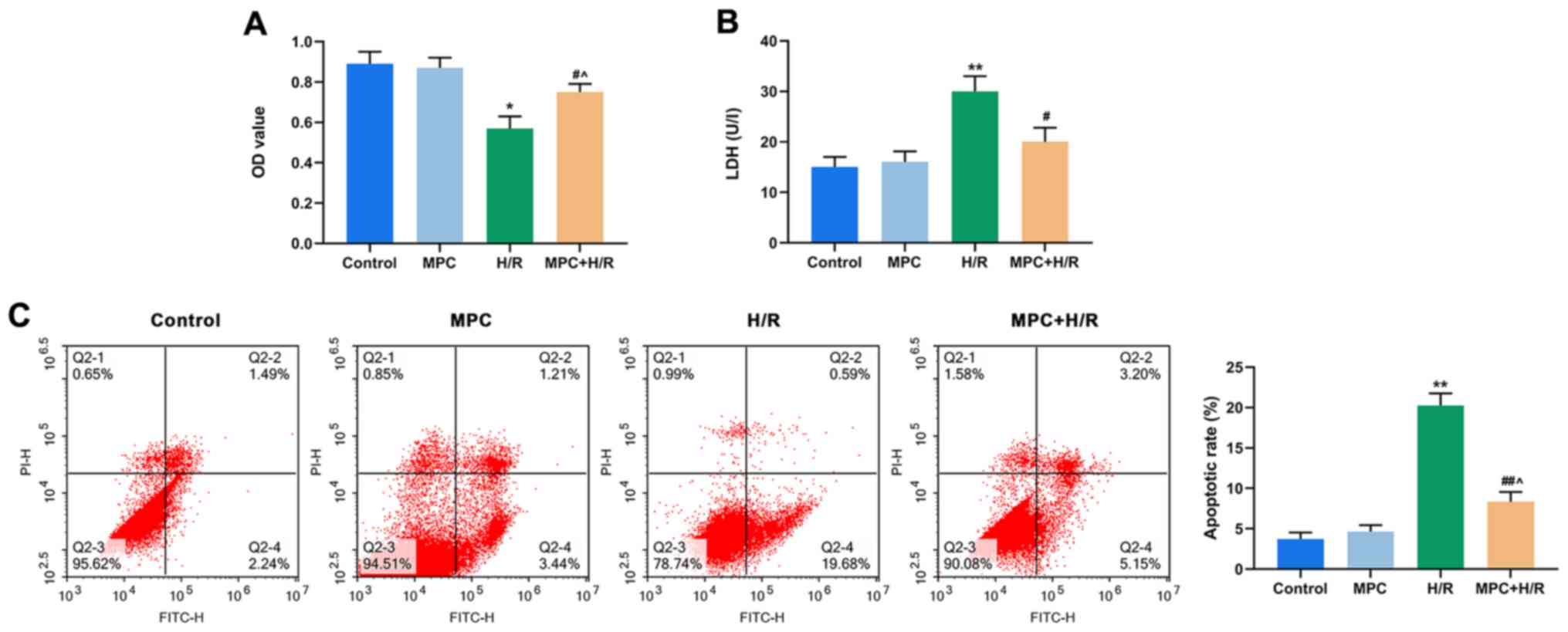 | Figure 1.MPC protects rat cardiomyocytes from
H/R damage. (A) MTT assay was used to assess the viability of H9c2
cells in the control, MPC, H/R and MPC+H/R groups. (B) Colorimetric
assay was used to measure LDH levels in the control, MPC, H/R and
MPC+H/R groups. (C) Flow cytometry was used to detect apoptosis in
the control, MPC, H/R and MPC+H/R groups. *P<0.05 and
**P<0.001 vs. control; #P<0.05 and
##P<0.001 vs. H/R; ^P<0.05 vs. MPC
group. MPC, morphine pre-conditioning; H/R, hypoxia/reoxygenation;
LDH, lactate dehydrogenase; OD, optical density; PI, propidium
iodide; FITC, fluorescein isothiocyanate. |
LDH levels indicate the number of dead and damaged
cells. In the H/R group, the LDH content was increased, compared
with the control. However, the LDH content in the MPC+H/R group was
significantly lower than that in the H/R group (P<0.05; Fig. 1B). In addition, the levels of
apoptosis were also examined in each of the four groups of cells.
The frequency of apoptotic cells in the H/R group was increased,
compared with control, but significantly decreased in the MPC+H/R
group, compared with the H/R group (P<0.05; Fig. 1C).
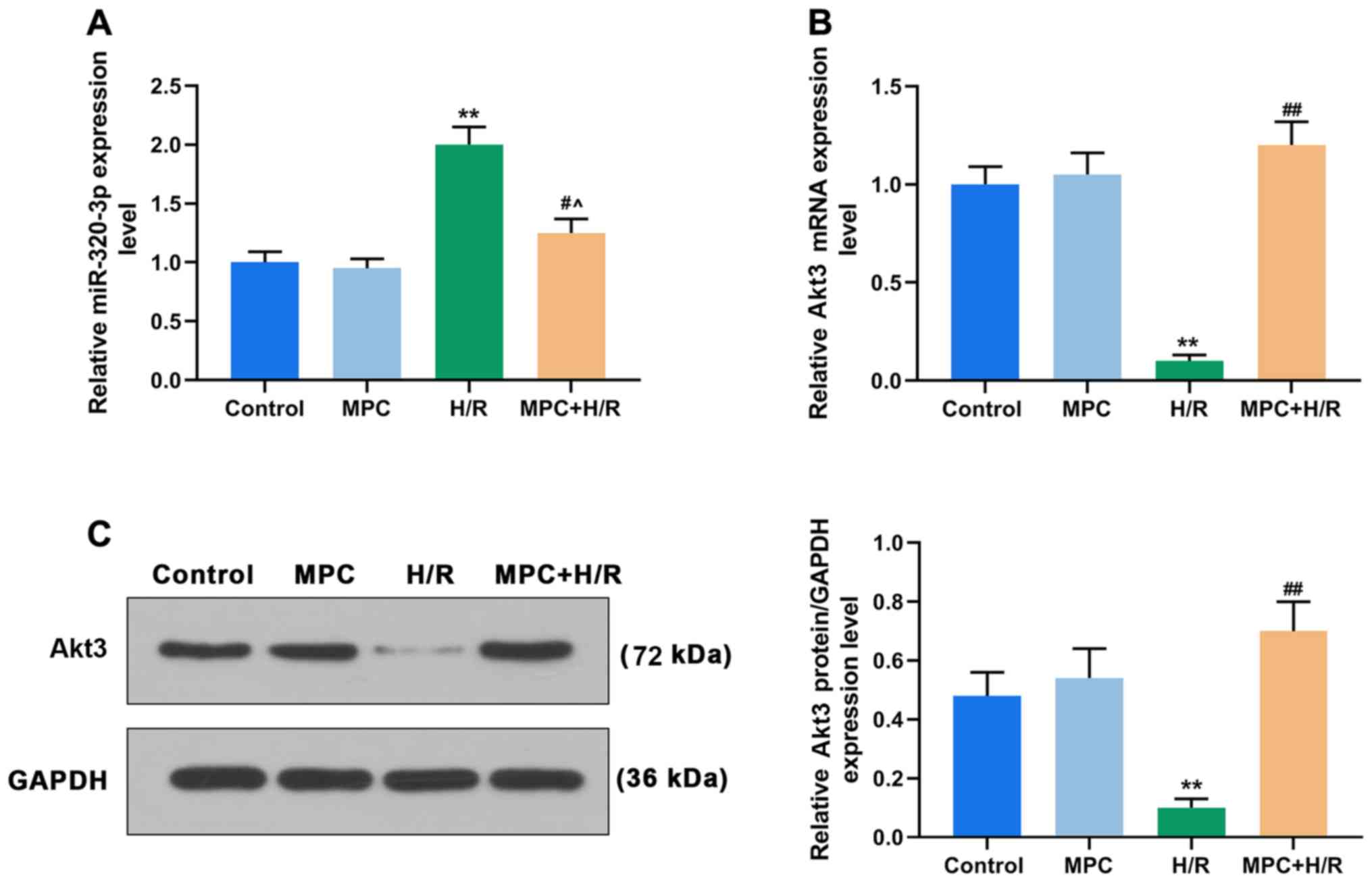
MPC regulates the expression of
miR-320-3p and Akt3
To further investigate the molecular mechanism of
MPC in rat cardiomyocytes, RT-qPCR was performed to detect the
expression of miR-320-3p. MPC treatment alone did not affect the
expression of miR-320-3p. The expression of miR-320-3p in the
MPC+H/R group was significantly lower than that in the H/R group
(P<0.05; Fig. 2A). Moreover,
the mRNA and protein expression levels of Akt3 were also examined.
MPC treatment alone did not affect the expression of Akt3. However,
the expression of Akt3 in the H/R group was reduced, compared with
the control. Additionally, expression of Akt3 in the MPC+H/R group
was significantly higher than that in the H/R group (P<0.05;
Fig. 2B and C).
Downregulation of miR-320-3p reduces
H/R damage to H9c2 cells
To further understand the biological role of
miR-320-3p in reperfusion injury of rat cardiomyocytes, H9c2 cells
were transfected with an miR-320-3p inhibitor (Fig. 3A). In the H/R group, an MTT assay
demonstrated that the viability of cells transfected with the
inhibitor (Inhibitor+H/R) was significantly higher than
untransfected cells and was similar to the MPC+H/R group
(P<0.05; Fig. 3B). Furthermore,
the LDH content in the Inhibitor+H/R group was lower than that in
the unstranfected H/R group and was similar to the MPC+H/R group
(P<0.05; Fig. 3C). As indicated
by flow cytometry, the rates of apoptosis of in the Inhibitor+H/R
group was similar to that of the MPC+H/R group. However, this was
significantly lower than that of the H/R group (P<0.001;
Fig. 3D).
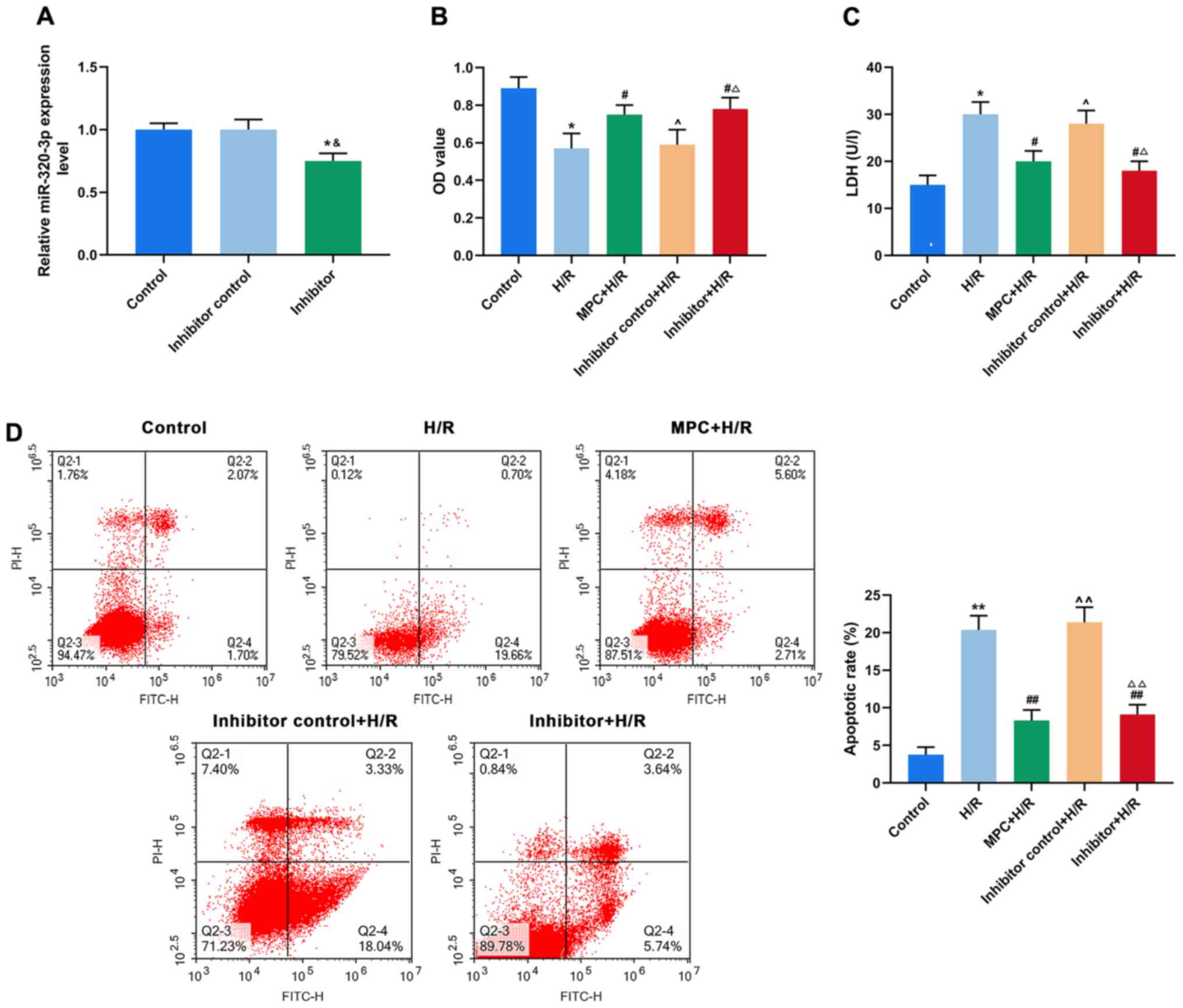 | Figure 3.Downregulation of miR-320-3p
attenuates H/R damage to cells. (A) Reverse
transcription-quantitative PCR was used to determine the effect of
miR-320-3p inhibitor transfection. (B) H9c2 cell viability in the
control, H/R, MPC+H/R, inhibitor control+H/R and inhibitor+H/R
groups using an MTT assay. (C) LDH content was measured in H9c2
cells in the control, H/R, MPC+H/R, inhibitor control+H/R and
inhibitor+H/R groups using a colorimetric assay. (D) Apoptosis of
H9c2 cells in the control, H/R, MPC+H/R, inhibitor control+H/R and
inhibitor+H/R groups was detected using flow cytometry.
&P<0.05 vs. Inhibitor control; *P<0.05 and
**P<0.001 vs. control; #P<0.05 and
##P<0.001 vs. H/R; ^P<0.05 and
^^P<0.001 vs. MPC+H/R; ΔP<0.05 and
ΔΔP<0.001 vs. Inhibitor control+H/R. MPC, morphine
pre-conditioning; H/R, hypoxia/reoxygenation; miR, microRNA; LDH,
lactate dehydrogenase; OD, optical density; PI, propidium iodide;
FITC, fluorescein isothiocyanate. |
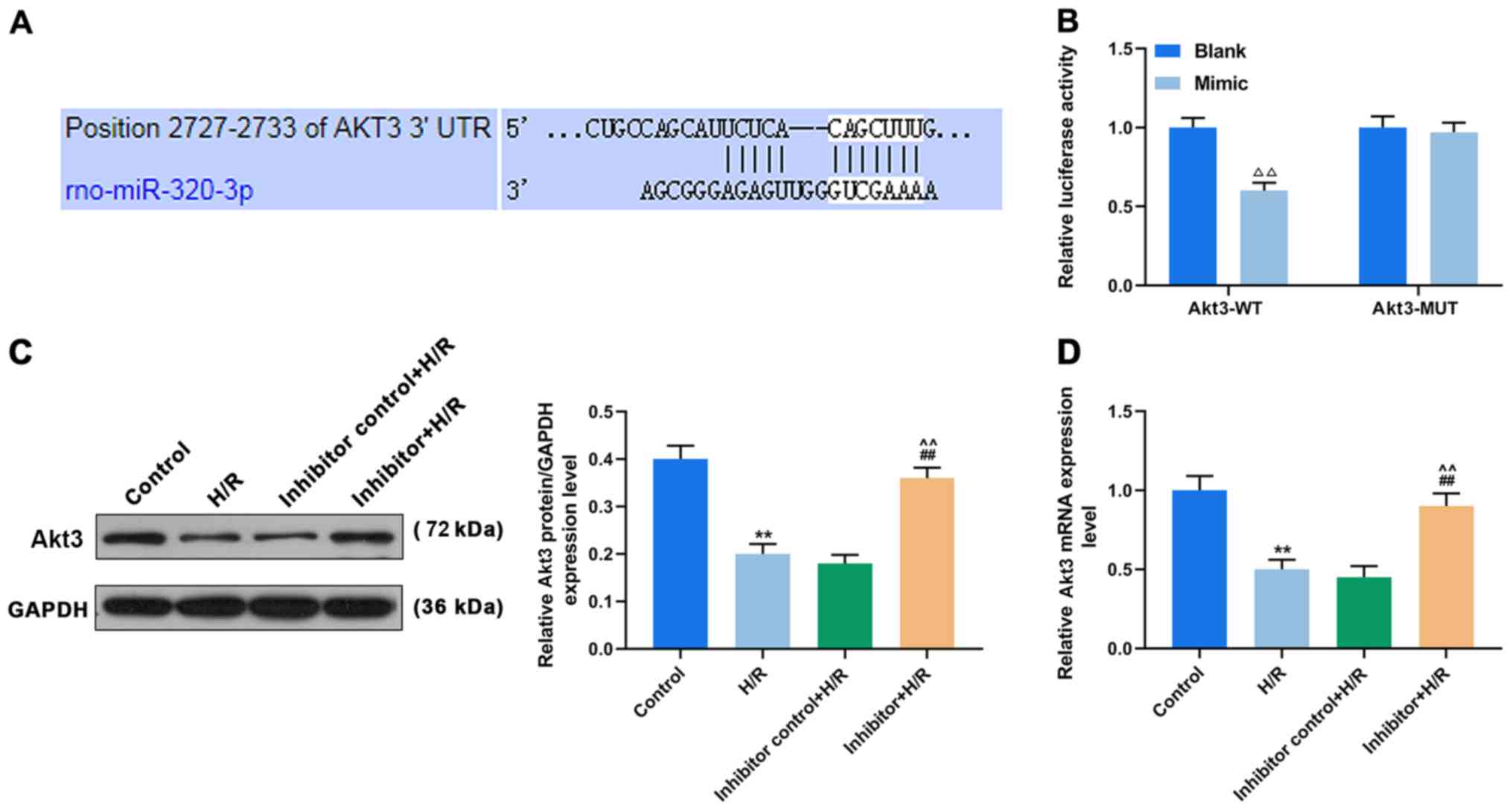
Upregulation of miR-320-3p targeting
Akt3 inhibits the protective effect of MPC on H9c2 cells
Using TargetScan v7.2 bioinformatics analysis and
luciferase assay, Akt3 was identified as the direct target of
miR-320-3p (Fig. 4A and B). Both
at the mRNA and protein levels, the expression of Akt3 was
significantly reduced in the untransfected cells of the H/R group,
compared with control. However, Akt3 expression in the H/R-treated
cells transfected with the inhibitor was significantly higher,
compared to the untransfected cells (P<0.001; Fig. 4C and D).
To better understand the effects of MPC, miR-320-3p
and Akt3 on reperfusion injury of rat cardiomyocytes, H9c2 cells
were transfected with an miR-320-3p mimic (Fig. 5A). The expression of Akt3 in
transfected cells of the MPC+H/R group (Mimic+MPC+H/R group) was
significantly lower than that in the untransfected MPC+H/R group
(P<0.05; Fig. 5B and C).
Moreover, an MTT assay suggested that cell viability in the
Mimic+MPC+H/R group was significantly lower than that in the
MPC+H/R group (P<0.05; Fig.
5D). To further assess the biological function of miR-320-3p in
H9c2 cells, LDH content was measured by colorimetry in each group.
The LDH content of the Mimic+MPC+H/R group was significantly higher
than that of the MPC+H/R group (P<0.05; Fig. 5E). In addition, the rates of
apoptosis in the Mimic+MPC+H/R group was significantly increased,
compared with the MPC+H/R group (P<0.05; Fig. 5F).
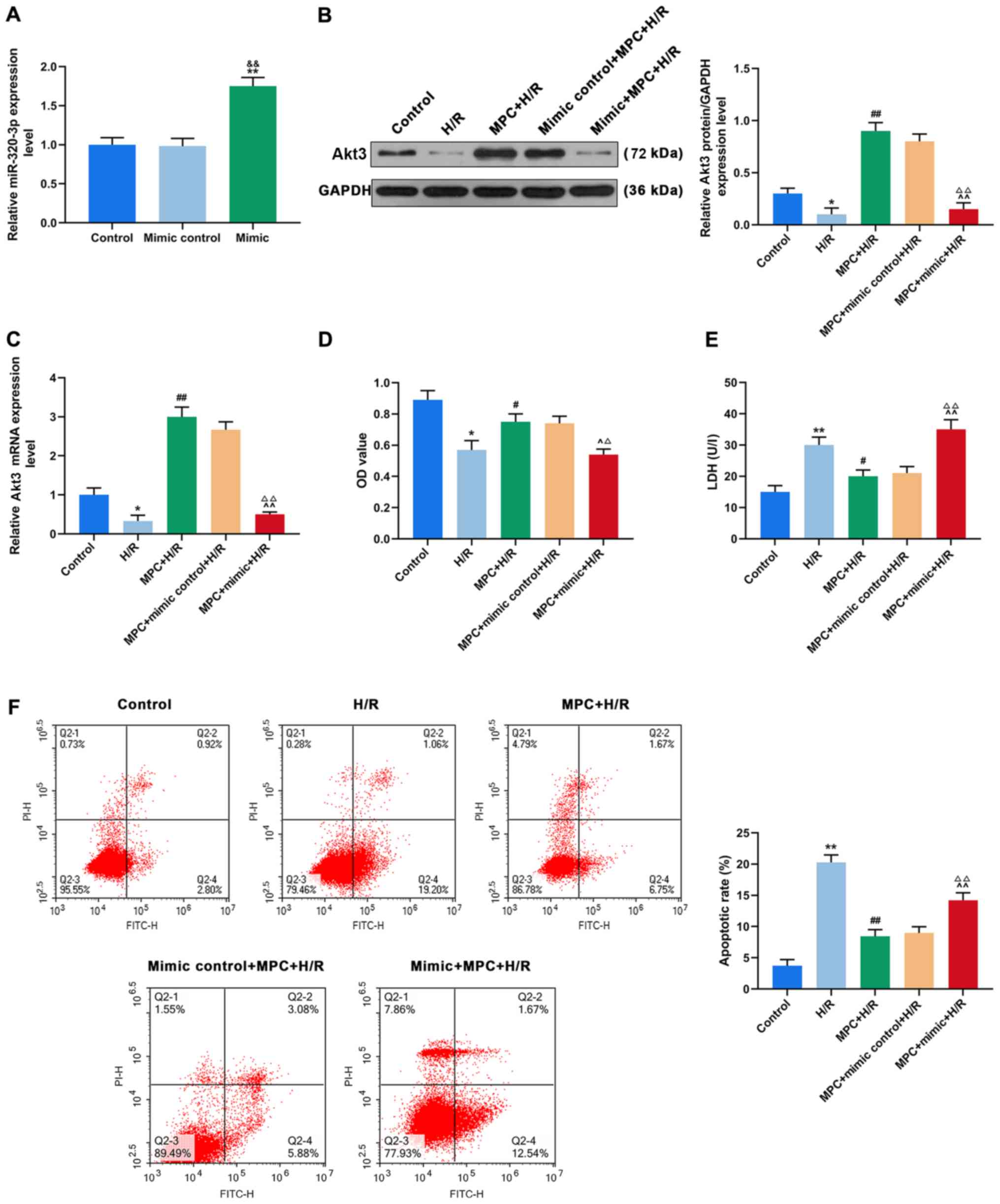 | Figure 5.Upregulation of miR-320-3p targeting
Akt3 inhibits the protective effect of MPC on H9c2 cells. (A)
RT-qPCR was used to determine the effect of miR-320-3p mimic
transfection. (B) RT-qPCR and (C) western blot analysis were used
to detect Akt3 expression in the control, H/R, MPC+H/R, MPC+Mimic
control+H/R and MPC+Mimic+H/R groups. (D) MTT assay was used to
assess H9c2 cell viability in the control, H/R, MPC+H/R, MPC+Mimic
control+H/R and MPC+Mimic+H/R groups. (E) LDH content was measured
in the control, H/R, MPC+H/R, MPC+Mimic control+H/R and
MPC+Mimic+H/R groups using a colorimetric assay. (F) Flow cytometry
was used to detect the apoptosis of H9c2 cells in the control, H/R,
MPC+H/R, MPC+Mimic control+H/R and MPC+Mimic+H/R groups.
&&P<0.001 vs. mimic control, *P<0.05 and
**P<0.001 vs. control, #P<0.05 and
##P<0.001 vs. H/R; ^P<0.05 and
^^P<0.001 vs. MPC+H/R, ΔP<0.05 and
ΔΔP<0.001 vs mimic control+MPC+H/R. MPC, morphine
pre-conditioning; H/R, hypoxia/reoxygenation; miR, microRNA;
RT-qPCR, reverse transcription-quantitative PCR; LDH, lactate
dehydrogenase; OD, optical density; PI, propidium iodide; FITC,
fluorescein isothiocyanate. |
Discussion
Myocardial ischemia/reperfusion injury (MI/RI) is a
pathophysiological process that occurs after revascularization in
patients with ischemic heart disease, such as myocardial infarction
(30,31). Morphine is a non-selective opioid
receptor agonist widely used as an anesthetic and in the treatment
and myocardial infarction (32).
Previous research has demonstrated that morphine has an early
pre-conditioning effect on cardiomyocytes that is triggered by
activation of opioid receptors (33). In the present study, the H9c2
cardiomyocyte hypoxia/reoxygenation injury model was established as
previously described (34) but
modified to incorporate 1 µM morphine as pre-treatment (35). In the present study, MPC
significantly enhanced the viability of cardiomyocytes, reduced the
levels of LDH resulting from H/R injury, and inhibited the
apoptosis of cardiomyocytes, indicating that MPC alleviates H/R
injury and plays a role in myocardial protection.
miRNAs are abundantly expressed in cardiomyocytes
and participate in pathophysiological processes, such as
cardiomyocyte apoptosis, myocardial remodeling and heart failure
(10). Several previous studies
suggest that MI/RI could be alleviated through the upregulation or
downregulation of the expression of target miRNAs, pointing to a
new therapeutic target for clinical myocardial protection (36,37).
Yang et al (38)
demonstrated that miRNA-22 can reduce apoptosis in MI/RI in a rat
model by inhibiting CREB-binding protein expression and its
downstream signaling pathway. Li et al (39) suggested that miRNA-145 expression
was downregulated in an MI/RI model, whereas upregulation of
miRNA-145 inhibited hydrogen peroxide-induced apoptosis, reactive
oxygen species production, and activation of key signaling proteins
in the mitochondrial apoptotic pathway. In addition, it has been
reported that morphine can upregulate the expression of miR-133b-5p
and protect cardiomyocytes from H/R injury, indicating that the
biological function of morphine may be achieved by regulating miRNA
expression (9).
Previous studies indicate that the regulation of
multiple signaling pathways could improve myocardial injury. For
example, melatonin protects the diabetic heart against
ischemia-reperfusion injury through membrane receptor-dependent
activation of cyclic GMP-dependent guanosine monophosphate protein
kinase (40). The inhibition of
miR-101a-3p alleviates H/R injury in H9c2 cells by regulating the
JAK2/STAT3 pathway (41).
Downregulation of miR-134 protects against myocardial H/R injury
through targeting nitric oxide synthase 3 and regulating the
PI3K/Akt pathway (42). However,
whether myocardial protection by MPC involves the regulation of
miR-320-3p and PI3K/Akt signaling pathways is still unclear.
The present findings demonstrated that miR-320-3p
expression was upregulated and Akt3 expression was downregulated in
cardiomyocytes after H/R injury, while MPC partially reversed the
changes in miR-320-3p and Akt3 expression induced by the injury.
This indicated that MPC plays an important role in the protection
of H9c2 cardiomyocytes by regulating miR-320-3p expression and the
PI3K/Akt signaling pathway. Previous studies also demonstrated that
miR-320 serves critical functions in many diseases such as glioma,
ovarian cancer and osteosarcoma (16,43–46).
Another study reported that miR-320 targeted A-kinase interacting
protein 1 to induce mitochondrial apoptosis, and contributed to
MI/RI (47). In the present study,
the role of miR-320-3p in myocardial ischemia/reperfusion was
further investigated using transfection of an miR-320-3p inhibitor,
which indicated that inhibition of miR-320-3p expression enhanced
cell viability, reduced LDH content and inhibited apoptosis.
Moreover, these results were consistent with a previous study that
suggested that miR-320-3p is involved in MI/RI (47).
In line with previous studies, it was hypothesized
that downregulation of miR-320-3p may be involved in mediating MPC
to protect myocardial cells from H/R injury through the regulation
of apoptosis. Through bioinformatics analysis and experimental
validation, it was determined that miR-320-3p could directly
interact with Akt3. The Akt3 gene encodes protein kinase B and
plays an important role in the regulation of several physiological
processes (48,49). Transgenic mice display significant
cardiac hypertrophy, suggesting that one of the functions of Akt3
is to promote cell growth (28).
The PI3K/Akt signaling pathway is also involved in cardiac
hypertrophy, myocardial cell remodeling and the prevention of
inflammation, thereby alleviating myocardial ischemia-reperfusion
injury (50–52). The present study suggested that
overexpression of miR-320-3p inhibited the expression Akt3
following MPC, and reduced the protective effect of MPC on H/R
injury in cardiomyocytes. This indicated that MPC may protect
cardiomyocytes from H/R injury through inhibiting miR-320-3p
expression and the PI3K/Akt signaling pathway.
In conclusion, the present study demonstrated that
MPC could significantly reduce H/R injury of H9c2 cardiomyocytes,
and its mechanism of action may be related to miR-320-3p and the
PI3K/Akt3 signaling pathway. The present findings provide a basis
for further research on miRNA regulation mechanism of morphine
pre-treatment.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed data sets used and/or analyzed during
the current study are available from the corresponding author on
reasonable request.
Authors' contributions
LC and SC made substantial contributions to
conception and design. LC and SC acquired, analyzed and interpreted
data. LC and SC drafted the article and critically revised the
draft for important intellectual content. LC and SC agree to be
accountable for all aspects of the work in ensuring that questions
related to the accuracy or integrity of the work are appropriately
investigated and resolved. All authors approved the final version
of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Frank A, Bonney M, Bonney S, Weitzel L,
Koeppen M and Eckle T: Myocardial ischemia reperfusion injury: From
basic science to clinical bedside. Semin Cardiothorac Vasc Anesth.
16:123–132. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Konstantinidis K, Whelan RS and Kitsis RN:
Mechanisms of cell death in heart disease. Arterioscler Thromb Vasc
Biol. 32:1552–1562. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen C, Jia KY, Zhang HL and Fu J: MiR-195
enhances cardiomyocyte apoptosis induced by hypoxia/reoxygenation
injury via downregulating c-myb. Eur Rev Med Pharmacol Sci.
20:3410–3416. 2016.PubMed/NCBI
|
|
4
|
Cao H, Xu H, Zhu G and Liu S: Isoquercetin
ameliorated hypoxia/reoxygenation-induced H9C2 cardiomyocyte
apoptosis via a mitochondrial-dependent pathway. Biomed
Pharmacother. 95:938–943. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
McGilliard KL and Takemori AE: Alterations
in the antagonism by naloxone of morphine-induced respiratory
depression and analgesia after morphine pretreatment. J Pharmacol
Exp Ther. 207:884–891. 1978.PubMed/NCBI
|
|
6
|
Swartjes M, Mooren RA, Waxman AR, Arout C,
van de Wetering K, den Hartigh J, Beijnen JH, Kest B and Dahan A:
Morphine induces hyperalgesia without involvement of µ-opioid
receptor or morphine-3-glucuronide. Mol Med. 18:1320–1326. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Okubo S, Tanabe Y, Takeda K, Kitayama M,
Kanemitsu S, Kukreja RC and Takekoshi N: Ischemic preconditioning
and morphine attenuate myocardial apoptosis and infarction after
ischemia-reperfusion in rabbits: Role of delta-opioid receptor. Am
J Physiol Heart Circ Physiol. 287:H1786–H1791. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang Y, Wang L, Li JH, Zhao HW and Zhang
FZ: Morphine alleviates myocardial ischemia/reperfusion injury in
rats by inhibiting TLR4/NF-κB signaling pathway. Eur Rev Med
Pharmacol Sci. 23:8616–8624. 2019.PubMed/NCBI
|
|
9
|
He SF, Zhu HJ, Han ZY, Wu H, Jin SY, Irwin
MG and Zhang Y: MicroRNA-133b-5p is involved in cardioprotection of
morphine preconditioning in rat cardiomyocytes by targeting fas.
Can J Cardiol. 32:996–1007. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Whelan RS, Kaplinskiy V and Kitsis RN:
Cell death in the pathogenesis of heart disease: Mechanisms and
significance. Annu Rev Physiol. 72:19–44. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hwang CK, Wagley Y, Law PY, Wei LN and Loh
HH: MicroRNAs in opioid pharmacology. J Neuroimmune Pharmacol.
7:808–819. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dong S, Cheng Y, Yang J, Li J, Liu X, Wang
X, Wang D, Krall TJ, Delphin ES and Zhang C: MicroRNA expression
signature and the role of microRNA-21 in the early phase of acute
myocardial infarction. J Biol Chem. 284:29514–29525. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cheng Y, Liu X, Zhang S, Lin Y, Yang J and
Zhang C: MicroRNA-21 protects against the H(2)O(2)-induced injury
on cardiac myocytes via its target gene PDCD4. J Mol Cell Cardiol.
47:5–14. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ren XP, Wu J, Wang X, Sartor MA, Jones K,
Qian J, Nicolaou P, Pritchard TJ and Fan GC: MicroRNA-320 is
involved in the regulation of cardiac ischemia/reperfusion injury
by targeting heat-shock protein 20. Circulation. 119:2357–2366.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu H, Li W, Zhang M, Zhu S, Zhang D and
Wang X: Inhibitory roles of miR-320 in osteosarcoma via regulating
E2F1. J Cancer Res Ther. 12:S68–S71. 2016. View Article : Google Scholar
|
|
17
|
Wang W, Yang J, Xiang YY, Pi J and Bian J:
Overexpression of hsa-miR-320 is associated with invasion and
metastasis of ovarian cancer. J Cell Biochem. 118:3654–3661. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen Q and Lesnefsky EJ: A new strategy to
decrease cardiac injury in aged heart following
ischaemia-reperfusion: Enhancement of the interaction between AMPK
and SIRT1. Cardiovasc Res. 114:771–772. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tian L, Cao W, Yue R, Yuan Y, Guo X, Qin
D, Xing J and Wang X: Pretreatment with Tilianin improves
mitochondrial energy metabolism and oxidative stress in rats with
myocardial ischemia/reperfusion injury via AMPK/SIRT1/PGC-1 alpha
signaling pathway. J Pharmacol Sci. 139:352–360. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen WR, Liu HB, Chen YD, Sha Y, Ma Q, Zhu
PJ and Mu Y: Melatonin attenuates myocardial ischemia/reperfusion
injury by inhibiting autophagy via an AMPK/mTOR signaling pathway.
Cell Physiol Biochem. 47:2067–2076. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu L, Jin X, Hu CF, Li R, Zhou Z and Shen
CX: Exosomes derived from mesenchymal stem cells rescue myocardial
ischaemia/reperfusion injury by inducing cardiomyocyte autophagy
Via AMPK and Akt pathways. Cell Physiol Biochem. 43:52–68. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kosuru R, Cai Y, Kandula V, Yan D, Wang C,
Zheng H, Li Y, Irwin MG, Singh S and Xia Z: AMPK contributes to
cardioprotective effects of pterostilbene against myocardial
ischemia-reperfusion injury in diabetic rats by suppressing cardiac
oxidative stress and apoptosis. Cell Physiol Biochem. 46:1381–1397.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gu C, Li T, Jiang S, Yang Z, Lv J, Yi W,
Yang Y and Fang M: AMP-activated protein kinase sparks the fire of
cardioprotection against myocardial ischemia and cardiac ageing.
Ageing Res Rev. 47:168–175. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen K, Li G, Geng F, Zhang Z, Li J, Yang
M, Dong L and Gao F: Berberine reduces ischemia/reperfusion-induced
myocardial apoptosis via activating AMPK and PI3K-Akt signaling in
diabetic rats. Apoptosis. 19:946–957. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun Y, Jiang C, Jiang J and Qiu L:
Dexmedetomidine protects mice against myocardium
ischaemic/reperfusion injury by activating an AMPK/PI3K/Akt/eNOS
pathway. Clin Exp Pharmacol Physiol. 44:946–953. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu J, Huang W, Ren C, Wen Q, Liu W, Yang
X, Wang L, Zhu B, Zeng L, Feng X, et al: Flotillin-2 promotes
metastasis of nasopharyngeal carcinoma by activating NF-κB and
PI3K/Akt3 signaling pathways. Sci Rep. 5:116142015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhou K, Fan YD, Wu PF, Duysenbi S, Feng
ZH, Du GJ and Zhang TR: MicroRNA-145 inhibits the activation of the
mTOR signaling pathway to suppress the proliferation and invasion
of invasive pituitary adenoma cells by targeting AKT3 in vivo and
in vitro. Onco Targets Ther. 10:1625–1635. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Khedr RM, Ahmed AAE, Kamel R and Raafat
EM: Sitagliptin attenuates intestinal ischemia/reperfusion injury
via cAMP/PKA, PI3K/Akt pathway in a glucagon-like peptide 1
receptor-dependent manner. Life Sci. 211:31–39. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tang H, Song X, Ling Y, Wang X, Yang P,
Luo T and Chen A: Puerarin attenuates myocardial
hypoxia/reoxygenation injury by inhibiting autophagy via the Akt
signaling pathway. Mol Med Rep. 15:3747–3754. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xiao R, Xiang AL, Pang HB and Liu KQ:
Hyperoside protects against hypoxia/reoxygenation induced injury in
cardiomyocytes by suppressing the Bnip3 expression. Gene.
629:86–91. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Coles JA Jr, Sigg DC and Iaizzo PA: Role
of kappa-opioid receptor activation in pharmacological
preconditioning of swine. Am J Physiol Heart Circ Physiol.
284:H2091–H2099. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Peart JN, Patel HH and Gross GJ:
Delta-opioid receptor activation mimics ischemic preconditioning in
the canine heart. J Cardiovasc Pharmacol. 42:78–81. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sun L, Zhao M, Yu XJ, Wang H, He X, Liu JK
and Zang WJ: Cardioprotection by acetylcholine: A novel mechanism
via mitochondrial biogenesis and function involving the PGC-1α
pathway. J Cell Physiol. 228:1238–1248. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liang BT and Gross GJ: Direct
preconditioning of cardiac myocytes via opioid receptors and KATP
channels. Circ Res. 84:1396–1400. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pan Z, Sun X, Ren J, Li X, Gao X, Lu C,
Zhang Y, Sun H, Wang Y, Wang H, et al: miR-1 exacerbates cardiac
ischemia-reperfusion injury in mouse models. PLoS One.
7:e505152012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yang Y, Del Re DP, Nakano N, Sciarretta S,
Zhai P, Park J, Sayed D, Shirakabe A, Matsushima S, Park Y, et al:
miR-206 mediates YAP-induced cardiac hypertrophy and survival. Circ
Res. 117:891–904. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang J, Chen L, Yang J, Ding J, Li S, Wu
H, Zhang J, Fan Z, Dong W and Li X: MicroRNA-22 targeting CBP
protects against myocardial ischemia-reperfusion injury through
anti-apoptosis in rats. Mol Biol Rep. 41:555–561. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li R, Yan G, Li Q, Sun H, Hu Y, Sun J and
Xu B: MicroRNA-145 protects cardiomyocytes against hydrogen
peroxide (H2O2)-induced apoptosis through
targeting the mitochondria apoptotic pathway. PLoS One.
7:e449072012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yu LM, Di WC, Dong X, Li Z, Zhang Y, Xue
XD, Xu YL, Zhang J, Xiao X, Han JS, et al: Melatonin protects
diabetic heart against ischemia-reperfusion injury, role of
membrane receptor-dependent cGMP-PKG activation. Biochim Biophys
Acta Mol Basis Dis. 1864:563–578. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu J, Wang J, Ning Y and Chen F: The
inhibition of miR-101a-3p alleviates H/R injury in H9C2 cells by
regulating the JAK2/STAT3 pathway. Mol Med Rep. 21:89–96.
2020.PubMed/NCBI
|
|
42
|
Xiao JM, Wang JJ and Sun LL: Effect of
miR-134 against myocardial hypoxia/reoxygenation injury by directly
targeting NOS3 and regulating PI3K/Akt pathway. Acta Cir Bras.
34:e2019008022019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Luo L, Yang R, Zhao S, Chen Y, Hong S,
Wang K, Wang T, Cheng J, Zhang T and Chen D: Decreased miR-320
expression is associated with breast cancer progression, cell
migration, and invasiveness via targeting Aquaporin 1. Acta Biochim
Biophys Sin (Shanghai). 50:473–480. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Pan C, Gao H, Zheng N, Gao Q, Si Y and
Zhao Y: miR-320 inhibits the growth of glioma cells through
downregulating PBX3. Biol Res. 50:312017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang T, Zou P, Wang T, Xiang J, Cheng J,
Chen D and Zhou J: Down-regulation of miR-320 associated with
cancer progression and cell apoptosis via targeting Mcl-1 in
cervical cancer. Tumour Biol. 37:8931–8940. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhu H, Jiang X, Zhou X, Dong X, Xie K,
Yang C, Jiang H, Sun X and Lu J: Neuropilin-1 regulated by miR-320
contributes to the growth and metastasis of cholangiocarcinoma
cells. Liver Int. 38:125–135. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Tian ZQ, Jiang H and Lu ZB: miR-320
regulates cardiomyocyte apoptosis induced by ischemia-reperfusion
injury by targeting AKIP1. Cell Mol Biol Lett. 23:412018.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Alcantara D, Timms AE, Gripp K, Baker L,
Park K, Collins S, Cheng C, Stewart F, Mehta SG, Saggar A, et al:
Mutations of AKT3 are associated with a wide spectrum of
developmental disorders including extreme megalencephaly. Brain.
140:2610–2622. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Chin YR, Yoshida T, Marusyk A, Beck AH,
Polyak K and Toker A: Targeting Akt3 signaling in triple-negative
breast cancer. Cancer Res. 74:964–973. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Carloni S, Girelli S, Buonocore G, Longini
M and Balduini W: Simvastatin acutely reduces ischemic brain damage
in the immature rat via Akt and CREB activation. Exp Neurol.
220:82–89. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yao H and Han X and Han X: The
cardioprotection of the insulin-mediated PI3K/Akt/mTOR signaling
pathway. Am J Cardiovasc Drugs. 14:433–442. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Song HP, Chu ZG, Zhang DX, Dang YM and
Zhang Q: PI3K-AKT pathway protects cardiomyocytes against
hypoxia-induced apoptosis by MitoKATP-mediated mitochondrial
translocation of pAKT. Cell Physiol Biochem. 49:717–727. 2018.
View Article : Google Scholar : PubMed/NCBI
|