Introduction
Acute lung injury (ALI) is a clinical syndrome
characterized by acute hypoxemic respiratory failure, pulmonary
infiltration and edema in the lung tissues, of which the most
severe manifestation is acute respiratory distress syndrome
(1). The mortality rate of
patients with ALI is ~30%, which has not changed over the past
several years (2,3). The increased permeability of the
alveolocapillary membrane is an important physiological alteration
that is observed in patients with ALI, which is induced by
alveolocapillary barrier dysfunction and epithelial injury.
Increased permeability of the alveolocapillary membrane causes lung
edema due to infiltration of large amounts of intravascular
protein-rich fluid into the pulmonary interstitial and alveolar
space (4). Moreover, overwhelming
inflammation is also a critical pathophysiological process observed
in patients with ALI (5). Previous
studies suggested that the extensive infiltration of neutrophils
and the presence of resident and recruited macrophages could
mediate robust inflammatory responses in patients with ALI and
animal models of ALI (6,7). Immune cells that accumulate in lung
tissues release various proinflammatory mediators, such as
interleukin (IL)-6, tumor necrosis factor (TNF)-α and interferon
(IFN)-γ, which exacerbate inflammation. Moreover, immune cells,
including proinflammatory macrophages, produce nitric oxide (NO)
(8,9).
Interferon regulatory factor 5 (IRF5) is widely
expressed in immune cells, including macrophages, B-cells,
monocytes and dendritic cells (10–12).
Previous studies demonstrated that IRF5 mediated the activation of
inflammatory macrophages in vivo and classical macrophages
in vitro (10–12). Activation of proinflammatory
macrophages leads to excessive secretion of TNF-α, IL-6, IL-12 and
IL-23, and downregulation of IL-10 and transforming growth factor-β
(13). Therefore, the expression
of IRF5 may also promote inflammatory macrophage activation and
inflammatory responses in lung tissue.
It has been reported that p38 mitogen-activated
protein kinase (p38MAPK) serves a crucial role in intracellular
inflammatory signaling pathways, including the TNF and TLR4
signaling pathways (14). However,
the effects of SB203580, an inhibitor of p38MAPK, on lung tissue
injury and the inflammatory response are not completely understood.
In the present study, a mouse model of ALI was established by 24-h
lipopolysaccharide (LPS) stimulation. The aim of the present study
was to investigate the effect of SB203580 on inflammatory response
and lung injury in ALI.
Materials and methods
Animals and treatment
In the present study, a total of 45 healthy male
C57BL/6 mice (age, 8 weeks; weight, 20–25 g) were purchased from
Changzhou Cavans Experimental Animal Co., Ltd. The animals were
kept under standardized conditions with a mean room temperature of
22–24°C, a 12/12h light/dark cycle and a humidity of 50–60%. The
mice were fed a standard animal diet with food and tap water ad
libitum. The mice were randomly divided into three groups
(Ctrl, ALI and SB203580 groups), each containing 15 mice. All
experiments were approved by The Institutional Animal Care and Use
Committee at Nanjing Medical University.
LPS causes ALI after intratracheal administration
and reduces associated non-pulmonary organ dysfunction, whereas
intravenous administration does not lead to tissue-specific lung
injury (15,16). Therefore, in the present study,
mice were treated with 50 µl (5 mg/kg) LPS (Escherichia coli
O111:B4; cat. no. L2630; Sigma-Aldrich; Merck KGaA) by
intratracheal injection under anesthesia with 40 mg/kg
pentobarbital sodium by intraperitoneal injection. In the control
(Ctrl) group, mice were treated with an equal volume of PBS by
intratracheal injection. The absorption rate of intraperitoneal
injection is lower compared with intratracheal injection,
therefore, SB203580 injection was performed prior to LPS treatment.
Thus, in the SB203580 group, mice received 20 mg/kg SB203580 (cat.
no. S1863; Beyotime Institute of Biotechnology) by intraperitoneal
injection 1 h before LPS stimulation. A total of 30 mice were
anesthetized by isoflurane inhalation (5%) and then sacrificed by
cervical dislocation at 24 h post-LPS treatment. For the
histological examination, immunohistochemistry and
immunofluorescence assays, a further 15 mice were anesthetized by
the intraperitoneal injection of pentobarbital sodium (40 mg/kg)
and sacrificed by myocardial perfusion fixation after LPS
stimulation for 24 h. Bronchoalveolar lavage fluid (BALF) and lung
tissues were collected for subsequent analysis at 24 h post-LPS
treatment, since the features of ALI in the model are close to the
diagnostic criteria of patients with ALI at this time point
(15,17).
Histological examination of lung
tissue
At 24 h post-LPS treatment, tissue from the middle
lobe of the right lung was collected, fixed with 10% formaldehyde
solution at room temperature for 24 h, dehydrated using a graded
ethanol series, embedded in paraffin and cut into 4-µm sections.
The sections were stained with hematoxylin for 10 min, followed by
eosin for 3 min, at room temperature. Sections were observed under
a BX53 light microscope (magnification, ×400; Olympus Corporation).
Lung tissue injuries were scored according to a previous study
(18).
Evaluation of pulmonary edema
Native tissue from the right lower lobe of the lung
was weighed to obtain the ‘wet weight’ (W). Subsequently, to obtain
the ‘dry weight’ (D), the tissues was dried at 65°C for 72 h and
then weighed. Pulmonary edema was evaluated by calculating the W/D
ratio.
BALF collection and cell counting
The pulmonary lavage was performed three times with
1 ml PBS. The collected BALF was centrifuged at 845 × g for 10 min
at 4°C. Subsequently, the cell pellet was resuspended with 1 ml
PBS. The total number of BALF cells was counted using a
hematocytometer under a light microscope (magnification, ×200).
Immunohistochemistry
To assess the expression and distribution of IRF5 in
lung tissue, the middle lobe of the right lung was fixed in 10%
formaldehyde at room temperature for 24 h, then dehydrated and
embedded in paraffin. All samples in paraffin were cut into 4-µm
sections. Subsequently, 4-µm sections were dewaxed and rehydrated
through graded alcohol at room temperature and then subjected to
antigen retrieval by high pressure in 10 mmol/l citrate acid (pH
6.0) for 8 min. The sections were blocked with 10% goat serum (cat.
no. AR0009; Boster Biological Technology) at room temperature for 1
h. The sections were incubated with an IRF5 primary antibody (cat.
no. ab181553; 1:600; Abcam) at 4°C overnight. The sections were
washed using PBS and then were incubated with a goat anti-rabbit
biotinylated secondary antibody (cat. no. ab6720; 1:1,000; Abcam)
at room temperature for 1.5 h, and streptavidin-horseradish
peroxidase (HRP; 1:10,000; cat. no. ab7403; Abcam) at room
temperature for 45 min, respectively. Chromagen detection was
performed with the DAB substrate kit (cat. no. ab64238; Abcam),
according to the manufacturer's instructions. Lung tissue sections
were counterstained with hematoxylin for 8 min at room temperature
and rinsed with distilled water, dehydrated through gradient
ethanol and xylene, then mounted with mounting medium (cat. no.
14177; Cell Signaling Technology, Inc.). Images were captured using
a BX53 light microscope (magnification, ×400; Olympus Corporation).
Quantification was performed by calculating the percentage of
IRF5-positive cells. IRF5-positive cells and total cells were
counted in five sections and five different fields/section.
Western blot analysis of the lung
tissues
Native tissue from the left lobe of the lung was
homogenized and protein samples were extracted using RIPA buffer
(cat. no. P0013B; Beyotime Institute of Biotechnology). Protein
content was determined using BCA kit (cat. no. P0012S; Beyotime
Institute of Biotechnology). Proteins (30 µg/line) were separated
via SDS-PAGE on 10% gel and transferred to PVDF membranes. PVDF
membranes were blocked using 5% non-fat milk at room temperature
for 1 h and then incubated with primary antibodies targeted against
IRF5 (cat. no. ab181553; 1:1,000; Abcam), inducible NO synthase
(iNOS; cat. no. ab15323; 1:1,000; Abcam), arginase 1 (Arg1; cat.
no. 93668; 1:1,000; Cell Signaling Technology, Inc.) and β-actin
(cat. no. ab8227; 1:2,000; Abcam) at 4°C overnight. Following
primary incubation, membranes were incubated with HRP-conjugated
goat anti-rabbit IgG (cat. no. A0208; 1:1000; Beyotime Institute of
Biotechnology) secondary antibodies at room temperature for 1.5 h.
In addition, enhanced chemiluminescence reagents (cat. no.
WBKLS0100; EMD Millipore) were used for imaging and quantification
was performed using ImageJ software version 1.47 (National
Institutes of Health) with β-actin as the loading control.
Immunofluorescence
Following treatment with LPS for 24 h, mice were
subjected to myocardial perfusion fixation to preserve tissue.
Tissue from the left lung was fixed with 10% formaldehyde solution
at room temperature for 4 h, dehydrated in 30% sucrose, embedded in
optimal cutting temperature reagent and cut into 7-µm sections
using a freezing microtome. The sections were blocked with 10% goat
serum (cat. no. AR0009; Boster Biological Technology) at room
temperature for 40 min. The frozen lung sections were stained with
F4/80 (1:50; cat. no. ab16911; Abcam) and iNOS (1:50; cat. no.
ab15323; Abcam) primary antibodies at 4°C overnight. Subsequently,
the sections were incubated with anti-rat Alexa Fluor®
594-conjugated (1:500; cat. no. A-11007; Invitrogen; Thermo Fisher
Scientific, Inc.) and anti-rabbit Alexa Fluor®
488-conjugated (1:500; cat. no. A-11034; Invitrogen; Thermo Fisher
Scientific, Inc.) secondary antibodies at room temperature for 2 h.
The localization of protein was examined by confocal laser scanning
microscopy (magnification, ×200; Leica Microsystems GmbH).
Measurements of cytokines
Following LPS treatment for 24 h, lung tissues were
ground into protein homogenate. The levels of the proinflammatory
cytokines TNF-α (cat. no. MTA00B) and IL-1β (cat. no. MLB00C) were
measured using ELISA kits (both R&D Systems, Inc.) according to
the manufacturer's instructions.
Statistical analysis
All experiments were repeated at least three times
and the data are presented as the mean ± SEM. Statistical analyses
were performed using GraphPad Prism software (version 7; GraphPad
Software, Inc.). Multiple comparisons were analyzed using one-way
ANOVA followed by Tukey's post hoc test. P<0.05 was considered
to indicate a statistically significant difference.
Results
SB203580 attenuates LPS-induced lung
histopathological alterations
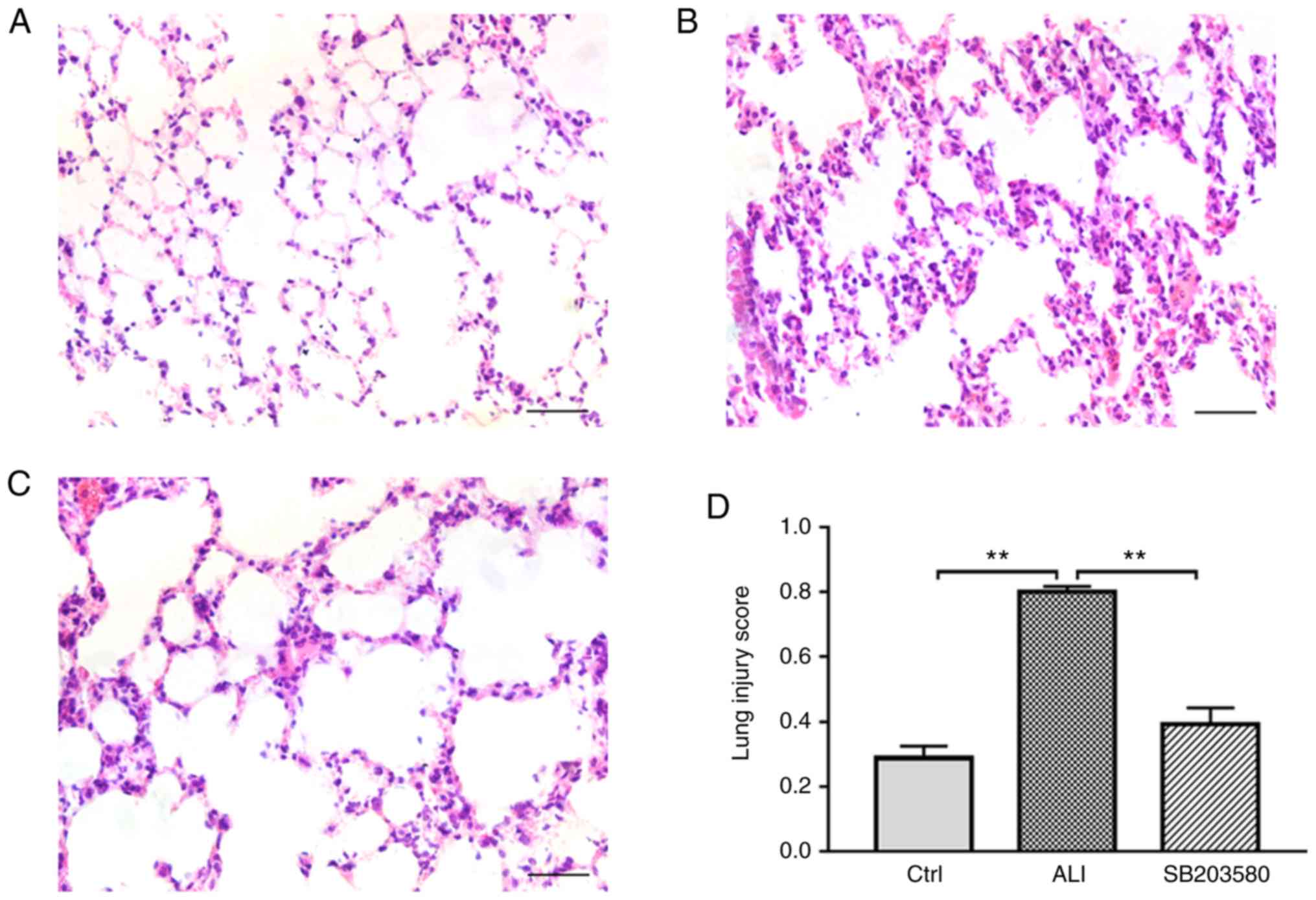
Histopathological alterations in the lung were
detected using hematoxylin and eosin (H&E) staining (Fig. 1). Lung sections of Ctrl group
showed a normal alveolar morphology (Fig. 1A). Histological sections from the
lungs of LPS-induced mice displayed edema, as well as basal
membrane and alveolar wall thickening (Fig. 1B). By contrast, treatment with
SB203580 markedly attenuated LPS-induced lung injury (Fig. 1C). The lung injury score in the ALI
group was significantly increased compared with the Ctrl group.
However, treatment with SB203580 significantly decreased the lung
injury score compared with the ALI group (Fig. 1D). Thus, the intraperitoneal
injection of SB203580 significantly decreased lung injury in ALI
model mice.
SB203580 reduces LPS-induced lung
edema and inflammatory responses in vivo
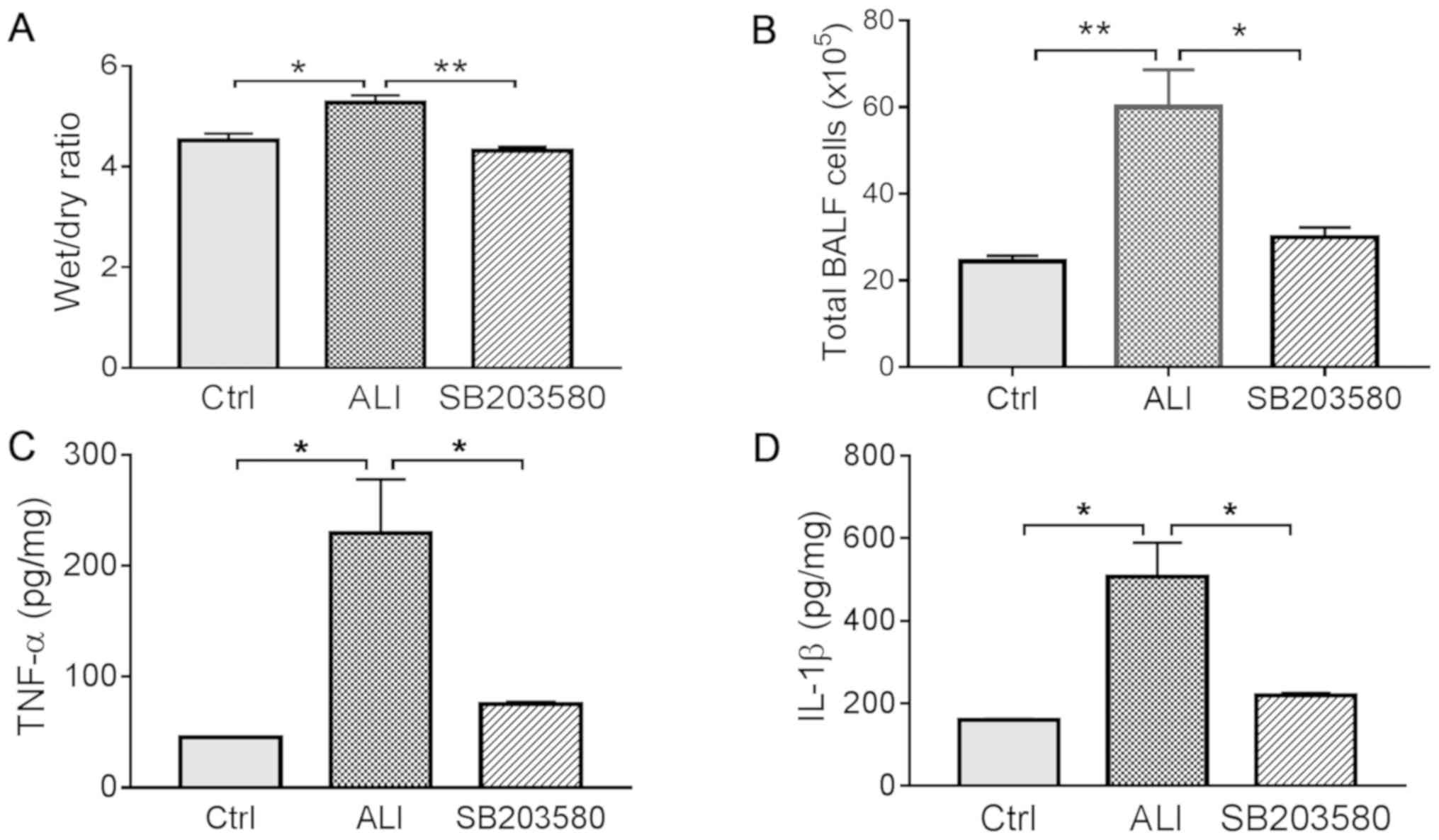
Lung tissue edema was assessed by calculating the
W/D weight ratio. The lung W/D ratio was significantly decreased in
the SB203580 group compared with the ALI group (Fig. 2A). In addition, inflammation was
evaluated by calculating the total number of cells in the BALF and
measuring proinflammatory cytokine levels in lung tissues. Total
cell counts in the BALF were higher in the ALI group compared with
the Ctrl group. However, treatment with SB203580 significantly
decreased cell counts in total BALF compared with the ALI group
(Fig. 2B). Similarly, the levels
of proinflammatory cytokines TNF-α and IL-1β were significantly
elevated in the ALI group compared with the Ctrl group, and
treatment with SB203580 significantly decreased the levels of TNF-α
and IL-1β compared with the ALI group (Fig. 2C and D).
SB203580 inhibits IRF5 expression in
the lung tissues of ALI model mice
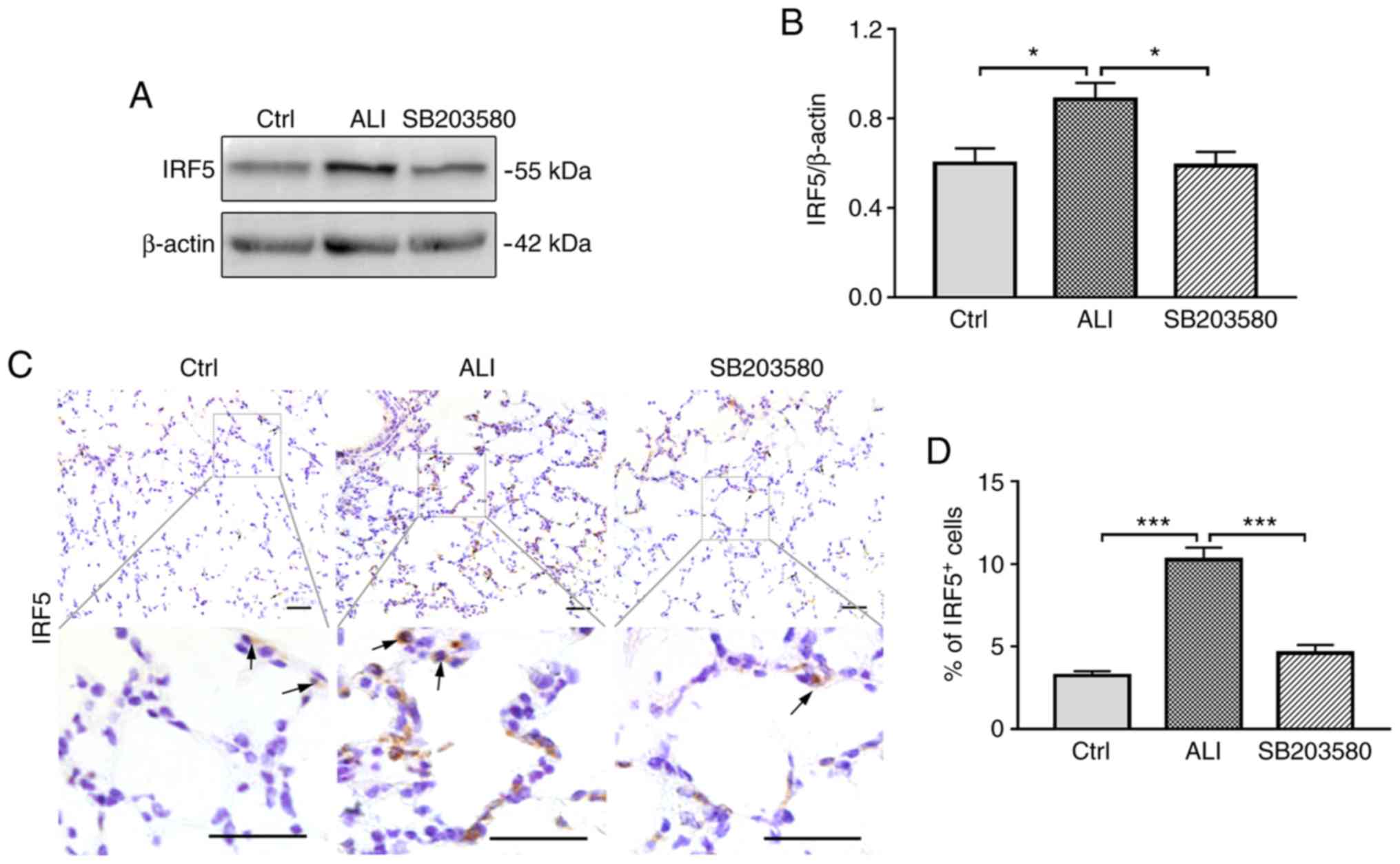
The effect of SB203580 on IRF5 expression was
assessed by western blotting and immunohistochemical staining. The
levels of IRF5 were significantly increased in ALI model mice
compared with Ctrl mice. Treatment with SB203580 resulted in a
significant decrease in IRF5 expression levels compared with the
ALI group (Fig. 3A and B). IRF5
expression was further investigated by immunohistochemical staining
(Fig. 3C). The frequency of
IRF5-postive cells was significantly increased in the lungs in the
ALI group compared with the Ctrl group; however, treatment with
SB203580 significantly inhibited LPS-induced IRF5 expression
(Fig. 3D).
SB203580 decreases the expression of
iNOS and F4/80 in the lung tissues of ALI model mice
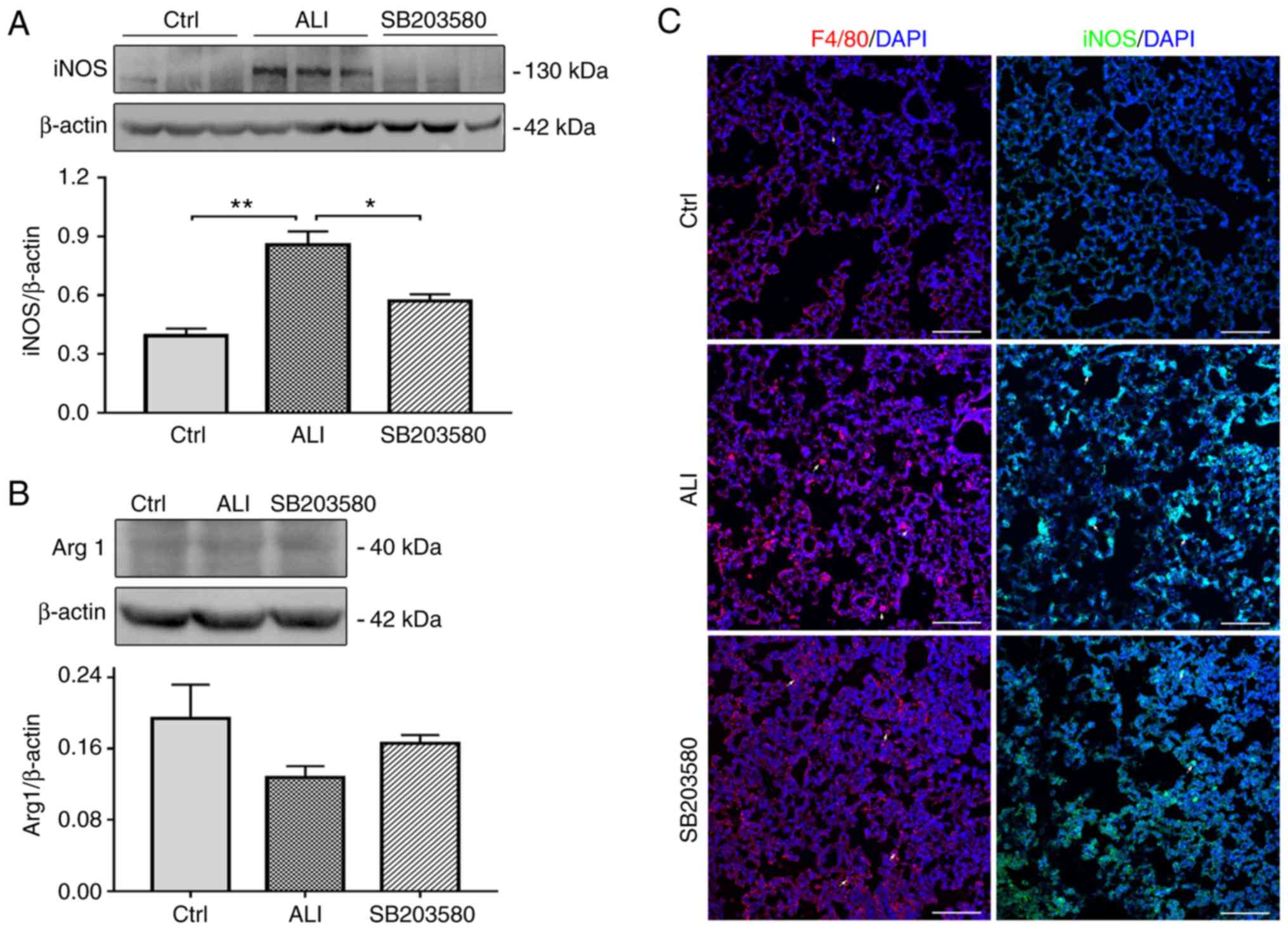
iNOS is an inflammatory gene marker, which serves a
critical role in LPS-induced ALI (19). In the present study, the effect of
SB203580 on iNOS expression was evaluated using western blotting
and immunofluorescence assays. The expression of iNOS was
significantly increased in the lung tissues of ALI model mice
compared with Ctrl mice. However, LPS-induced iNOS expression was
significantly decreased following SB203580 treatment (Fig. 4A and C). The levels of Arg1, were
low in the Ctrl group and remained unchanged in the ALI group
(Fig. 4B).
F4/80 is widely used as a macrophage marker,
although it is also expressed on monocytes and neutrophils in mice
(20,21). The expression of F4/80 was
increased in ALI lung tissues compared with Ctrl lung tissues,
which was inhibited by SB203580 treatment (Fig. 4C).
Discussion
ALI is a common clinical condition characterized by
hypoxia, edema and inflammatory cell infiltration of neutrophils
and macrophages into lung tissues, leading to respiratory failure
(22,23). Alveolar epithelium and capillary
endothelium injuries increase alveolar barrier permeability,
resulting in edema, inflammatory leukocyte infiltration and
bleeding (4,24). Focusing on the treatment of lung
edema and inflammatory injury in patients with ALI is of
significance. Mouse models of LPS-induced lung injury have been
widely used to study the pathogenesis of ALI (4,18).
The mild LPS-induced mouse model of ALI used in the present study
displayed edema and inflammation in lung tissues.
Inhibition of proinflammatory cytokine production
allows the regulation of inflammatory responses (25). The p38MAPK signaling pathway has a
key regulatory role in inflammatory processes taking place in the
lung (26). M39, an inhibitor of
p38MAPK, inhibits the activation of p38MAPK in neutrophils and
macrophages in vitro, which prevents the release of TNF-α
and macrophage inflammatory protein 2, limits the activation of
p38MAPK in vivo and reduces the accumulation of neutrophils
in the airway. Moreover, the novel synthesized flavonoid LFG-500
can alleviate LPS-induced inflammatory responses by inhibiting the
expression of IL-6, TNF-α and IL-1β (27).
In the present study, the effects of SB203580, an
inhibitor of p38MAPK, on ALI and inflammation in LPS-induced mice
were investigated. SB203580 treatment significantly reduced
LPS-induced expression of TNF-α and IL-1β in lung tissues, which
suggested that SB203580 inhibited ALI-stimulated inflammatory
responses by inhibiting the expression of proinflammatory
cytokines. Furthermore, SB203580 reduced the W/D weight ratio
compared with the ALI group, indicating attenuation of LPS-induced
lung edema.
The transcription factor IRF5 is involved in
multiple autoimmune diseases, including systemic lupus
erythematosus, rheumatoid arthritis and inflammatory bowel disease
(28). IRF5 induces the
upregulation of proinflammatory genes, repression of
anti-inflammatory mediators and polarization of macrophages to a
proinflammatory phenotype (28–30).
In the present study, the expression of IRF5 was significantly
upregulated in ALI model mice compared with Ctrl mice, and
significantly downregulated in SB203580-treated ALI model mice. The
results suggested that SB203580 inhibited LPS-stimulated
inflammatory responses by decreasing the expression of IRF5 in lung
tissue.
iNOS is an enzyme expressed in macrophages and
endothelial cells that synthesizes NO via L-arginase oxidation.
iNOS competes with Arg1 for the same substrate and is considered a
marker of inflammatory responses (31,32).
In the present study, the expression of iNOS was increased in the
lungs of ALI mice compared with Ctrl mice, and SB203580 treatment
decreased LPS-induced iNOS expression. By contrast, the expression
of Arg1 in lung tissues remained almost unchanged among the three
groups. Thus, the results suggested that SB203580 attenuated the
inflammatory response by decreasing the expression of iNOS in
LPS-induced lung tissues.
In conclusion, the inhibitor of the p38MAPK
signaling pathway SB203580 attenuated lung injury and inflammatory
responses in ALI model mice. Treatment with SB203580 in LPS-induced
ALI model mice reduced edema and the expression of proinflammatory
cytokines, and alleviated pathological changes in lung tissues.
SB203580 also decreased the expression of IRF5 and iNOS in ALI lung
tissues. Thus, IRF5 may serve as a key regulatory factor of the
inflammatory response in the lungs and may exert its effects by
promoting a proinflammatory macrophage phenotype. Although further
research is required to fully understand the mechanism of SB203580
attenuating lung edema and inflammatory response in ALI, the
present study suggested a protective role of SB203580 in
LPS-induced ALI; therefore, SB203580 may serve as a potential
preventive agent for ALI.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural
Science Foundation of Jiangsu Province (grants nos. BK20190150 and
BK20160196) and the National Natural Science Foundation of China
(grant no. 81500039).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
GL designed the study, analyzed the data and wrote
the manuscript. YD performed the hematoxylin and eosin staining and
immunohistochemistry assay. JT performed immunofluorescence and
ELISA assays. JZo and XN analyzed the data and helped to write the
paper. ZY, JZh and XY performed animal experiments and western blot
assays. JC designed the present study and provided financial
support for this work. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All experiments in the present study were approved
by The Institutional Animal Care and Use Committee at Nanjing
Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ashbaugh DG, Bigelow DB, Petty TL and
Levine BE: Acute respiratory distress in adults. Lancet. 2:319–323.
1967. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Villar J, Blanco J and Kacmarek RM:
Current incidence and outcome of the acute respiratory distress
syndrome. Curr Opin Crit Care. 22:1–6. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Erickson SE, Martin GS, Davis JL, Matthay
MA and Eisner MD; NIH NHLBI ARDS Network, : Recent trends in acute
lung injury mortality: 1996–2005. Crit Care Med. 37:1574–1579.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Patel BV, Wilson MR and Takata M:
Resolution of acute lung injury and inflammation: A translational
mouse model. Eur Respir J. 39:1162–1170. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gill SE, Yamashita CM and Veldhuizen RA:
Lung remodeling associated with recovery from acute lung injury.
Cell Tissue Res. 367:495–509. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Matute-Bello G, Frevert CW and Martin TR:
Animal models of acute lung injury. Am J Physiol Lung Cell Mol
Physiol. 295:L379–L399. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Han S and Mallampalli RK: The acute
respiratory distress syndrome: From mechanism to translation. J
Immunol. 194:855–860. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Robb CT, Regan KH, Dorward DA and Rossi
AG: Key mechanisms governing resolution of lung inflammation. Semin
Immunopathol. 38:425–448. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Freire MO and Van Dyke TE: Natural
resolution of inflammation. Periodontol 2000. 63:149–164. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Krausgruber T, Blazek K, Smallie T,
Alzabin S, Lockstone H, Sahgal N, Hussell T, Feldmann M and Udalova
IA: IRF5 promotes inflammatory macrophage polarization and TH1-TH17
responses. Nat Immunol. 12:231–238. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lien C, Fang CM, Huso D, Livak F, Lu R and
Pitha PM: Critical role of IRF-5 in regulation of B-cell
differentiation. Proc Natl Acad Sci USA. 107:4664–4668. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Krausgruber T, Saliba D, Ryzhakov G,
Lanfrancotti A, Blazek K and Udalova IA: IRF5 is required for
late-phase TNF secretion by human dendritic cells. Blood.
115:4421–4430. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Aggarwal NR, King LS and D'Alessio FR:
Diverse macrophage populations mediate acute lung inflammation and
resolution. Am J Physiol Lung Cell Mol Physiol. 306:L709–L725.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schnyder-Candrian S, Quesniaux VF, Di
Padova F, Maillet I, Noulin N, Couillin I, Moser R, Erard F,
Vargaftig BB, Ryffel B, et al: Dual effects of p38 MAPK on
TNF-dependent bronchoconstriction and TNF-independent neutrophil
recruitment in lipopolysaccharide-induced acute respiratory
distress syndrome. J Immunol. 175:262–269. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Szarka RJ, Wang N, Gordon L, Nation PN and
Smith RH: A murine model of pulmonary damage induced by
lipopolysaccharide via intranasal instillation. J Immunol Methods.
202:49–57. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen H, Bai C and Wang X: The value of the
lipopolysaccharide-induced acute lung injury model in respiratory
medicine. Expert Rev Respir Med. 4:773–783. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
van Helden HP, Kuijpers WC, Steenvoorden
D, Go C, Bruijnzeel PL, van Eijk M and Haagsman HP: Intratracheal
aerosolization of endotoxin (LPS) in the rat: A comprehensive
animal model to study adult (acute) respiratory distress syndrome.
Exp Lung Res. 23:297–316. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Matute-Bello G, Downey G, Moore BB,
Groshong SD, Matthay MA, Slutsky AS and Kuebler WM; Acute Lung
Injury in Animals Study Group, : An official American Thoracic
Society workshop report: Features and measurements of experimental
acute lung injury in animals. Am J Respir Cell Mol Biol.
44:725–738. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mehta S: The effects of nitric oxide in
acute lung injury. Vascul Pharmacol. 43:390–403. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Taylor PR, Martinez-Pomares L, Stacey M,
Lin HH, Brown GD and Gordon S: Macrophage receptors and immune
recognition. Annu Rev Immunol. 23:901–944. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gordon S and Mantovani A: Diversity and
plasticity of mononuclear phagocytes. Eur J Immunol. 41:2470–2472.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hughes KT and Beasley MB: Pulmonary
manifestations of acute lung injury: More than just diffuse
alveolar damage. Arch Pathol Lab Med. 141:916–922. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Butt Y, Kurdowska A and Allen TC: Acute
Lung Injury: A Clinical and Molecular Review. Arch Pathol Lab Med.
140:345–350. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Elicker BM, Jones KT, Naeger DM and Frank
JA: Imaging of Acute Lung Injury. Radiol Clin North Am.
54:1119–1132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pyee Y, Chung HJ, Choi TJ, Park HJ, Hong
JY, Kim JS, Kang SS and Lee SK: Suppression of inflammatory
responses by handelin, a guaianolide dimer from Chrysanthemum
boreale, via downregulation of NF-κB signaling and pro-inflammatory
cytokine production. J Nat Prod. 77:917–924. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nick JA, Young SK, Brown KK, Avdi NJ,
Arndt PG, Suratt BT, Janes MS, Henson PM and Worthen GS: Role of
p38 mitogen-activated protein kinase in a murine model of pulmonary
inflammation. J Immunol. 164:2151–2159. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li C, Yang D, Cao X, Wang F, Jiang H, Guo
H, Du L, Guo Q and Yin X: LFG-500, a newly synthesized flavonoid,
attenuates lipopolysaccharide-induced acute lung injury and
inflammation in mice. Biochem Pharmacol. 113:57–69. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Eames HL, Corbin AL and Udalova IA:
Interferon regulatory factor 5 in human autoimmunity and murine
models of autoimmune disease. Transl Res. 167:167–182. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Khoyratty TE and Udalova IA: Diverse
mechanisms of IRF5 action in inflammatory responses. Int J Biochem
Cell Biol. 99:38–42. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Weiss M, Blazek K, Byrne AJ, Perocheau DP
and Udalova IA: IRF5 is a specific marker of inflammatory
macrophages in vivo. Mediators Inflamm. 2013:2458042013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cook HT, Jansen A, Lewis S, Largen P,
O'Donnell M, Reaveley D and Cattell V: Arginine metabolism in
experimental glomerulonephritis: Interaction between nitric oxide
synthase and arginase. Am J Physiol. 267:F646–F653. 1994.PubMed/NCBI
|
|
32
|
Li Z, Zhao ZJ, Zhu XQ, Ren QS, Nie FF, Gao
JM, Gao XJ, Yang TB, Zhou WL, Shen JL, et al: Differences in iNOS
and arginase expression and activity in the macrophages of rats are
responsible for the resistance against T. gondii infection. PLoS
One. 7:e358342012. View Article : Google Scholar : PubMed/NCBIPubMed/NCBIPubMed/NCBI
|