Introduction
Coronary heart disease is caused by structural or
functional coronary lesions in the coronary artery, affecting the
blood supply and leading to demand imbalances as a result of
myocardial damage (1). Acute
myocardial infarction (AMI) is a serious disease associated with
high mortality rates (2).
According to the guidelines of the American College of Cardiology
published in 2014, re-establishing the blood supply to ischemic
myocardial tissues is the most viable therapeutic approach for the
treatment of AMI (3). However,
coronary revascularization can induce myocyte damage such as the
injury of myocardial structure, cardiac function, myocardial cell
metabolism and electrophysiological activity (4). In addition, pathological and
physiological changes in endothelial cells, microvascular
dysfunction, myocardial necrosis, arrhythmia and myocardial
stunning are also associated with coronary revascularization
(4). These effects are known as
myocardial ischemic reperfusion injury (MIRI) (5). The mechanisms of MIRI have not been
fully characterized.
Toll-like receptor 4 (TLR4) is a member of the
mammalian TLR family. TLR4 can activate myeloid differentiation
factor 88 (MyD88)-dependent pathways (6). TLR4 can also regulate the
transcriptional activity of NF-κB and interferon regulating factor
3 (IRF3), which promotes the production of inflammatory factors
(7–9). As a member of TLR family, TLR2 can
participate in the inflammatory response by modulating the MyD88
signaling pathway, and the activation of NF-κB and mitogen
activated protein kinase (MAPK) (10). The aim of the present study was to
investigate the mechanism of the TLR4/TLR2 signaling pathway in
MIRI in order to facilitate the treatment development for MIRI.
Radioprotective 105 kDa protein (RP105) is a
single-pass type I membrane protein belonging to the TLR family
(11). RP105 plays an important
role in regulating the proliferation and radio-activation of murine
B cells (12). The expression
level of RP105 is closely associated with the inflammatory response
in MIRI (13). RP105 acts as a key
inhibitor of the TLR4 pathway by forming the RP105/myeloid
differentiation protein-1 complex (MD-1) and decreases the binding
of TLR4 to ligands (14,15). MD-2 is a glycoprotein that was
discovered by sequence homology with MD-1. MD-2 determines the
surface expression and signal transmission of TLR4 (11,16).
Transfection of RP105 into cardiocytes significantly decreased the
production of pro-inflammatory cytokines to alleviate the heart
from MIRI by inhibiting both the TLR4/MyD88 and
TLR4/TIR-domain-containing adapter-inducing interferon-β (TRIF)
signaling pathways (17,18). The TLR2-dependent p38/MAPK and
transcription factor activator protein-1 (AP-1) signaling pathways
play critical roles in the pathogenesis of reperfusion injury of
the myocardium, retina, liver and intestine (19–21).
In the present study, the role of RP105 and the mechanism via which
RP105 regulated TLR2/4 in MIRI were investigated by knockdown of
RP105 in the cardiomyocytes of rats. The present results indicated
the role of the RP105/TLR2/4 axis in MIRI.
Materials and methods
Animals
In total, 75 adult Sprague-Dawley rats (males;
weight, 220–250 g; age, 7–8 weeks) were provided by the Animal
Experiment Center of China Three Gorges University (certificate no.
SYXK2017-0061). The animal experiments were approved by the Animal
Care and Use Department of China Three Gorges University. All rats
were housed in a controlled temperature (25±2°C) with relative
humidity 60±5% and 12:12-h light/dark cycle. All rats were housed
in a specific pathogen-free barrier environment with free access to
food and water.
The rats were randomly divided into the following
five groups (n=15/group): i) Sham operation group (sham group); ii)
saline injection and ischemic and reperfusion (I/R) operation group
(I/R); iii) myocardial I/R with Ad-RP105 group (Ad-RP105); iv)
myocardial I/R with Ad-RP105-small interfering RNA (siRNA) group
(Ad-RP105-siRNA); and v) myocardial I/R with Ad-enhanced green
fluorescence protein (EGFP)-siRNA group (Ad-G-siRNA).
Construction of adenoviral
vectors
The adenovirus vectors expressing EGFP-RP105- siRNA
(5′-UAAUAGCUCUGGCAAUUUGGC-3′; Shanghai GeneChem Co., Ltd.),
EGFP-RP105 (full-length cDNA of RP-105) or EGFP-siRNA
(5′-TAGTCGCATACGGAACATTCG-3′; Shanghai GeneChem Co., Ltd.) were
constructed by inserting the sequence into the PL-SIN lentiviral
vector (Plasmid #21316; Addgene, USA). Then, 5 µg of plasmids were
transfected into 293 cells to amplify the adenovirus. The
adenoviruses were further purified with the Adeno-X TM Virus
Purification kit (BD Biosciences Clontech) after transfection for
48 h at romm temperature according to the manufacturer's
instructions.
MIRI model
The establishment of the MIRI model was performed as
previously described (22).
Animals were anesthetized by intraperitoneally injecting 40 mg/kg
sodium pentobarbital. The subcutaneous tissue was cut and separated
along the upper part of the breastbone. The trachea was fully
exposed for insertion; a venous indwelling 24G needle was used to
ventilate the animal via endotracheal intubation. The animal
ventilator was set at the rate of 80 breaths/min and the
inspiratory/expiratory ratio was set as 2:1. The chest was exposed
by a left thoracotomy between the fourth and fifth ribs. In total,
100 µl of saline, 30 µl of lentivirus expressing Ad-RP105-EGFP,
Ad-G-siRNA or Ad-RP105-siRNA was injected at the apex of four
separate positions in the heart of each group. After 3 days, the
rats were again subjected to anesthesia, ventilation and
thoracotomy as described above. The heart was exposed once again,
and the left anterior descending coronary artery (LAD) was ligated
using 6-0 silk suture with medical latex tubing to ensure full
occlusion of LAD. Myocardial ischemia was recorded by serum
detection. All animals were underwent 30 min of myocardial ischemia
followed by 3 h of reperfusion as described previously (23–25).
Rats in the sham group were subjected to the same operation but
without blockage of LAD. After I/R surgery, 200 µl of blood was
collected from the inferior vena cava of the rats. The euthanasia
of rats was performed via cervical dislocation after anesthesia.
Humane endpoints set for the euthanasia of rats at the end of the
experiment included behavioral changes such as labored breathing,
not eating or drinking, half-closed eyes and reduced exploration.
The death of the animals was verified by checking the breathing
rate, heartbeat and pupillary reflexes. The health and behaviors of
rats were monitored every day during the experiments. Myocardial
tissues were collected for subsequent experiments.
Immunofluorescence assay
After I/R for 3 h, myocardium was immediately fixed
with 4% paraformaldehyde at room temperature for 2 h, embedded in
paraffin and sliced (4 µm). Slices were dewaxed and then underwent
antigen fixing. Slices were blocked with 1% BSA (Sigma-Aldrich;
Merck KGaA) for 30 min at room temperature (RT) and incubated with
anti-RP105 antibody (cat. no. Sc-13592; 1:1,000; Santa Cruz
Biotechnology, Inc.) overnight at 4°C. Then, the slices were
incubated with goat anti-Rat IgG (H+L) cross-adsorbed secondary
antibody (1:5,000; cat. no. A-11006; Thermo Fisher Scientific,
Inc.) for 60 min at RT and subsequently stained with DAPI at room
temperature for 10 min in the dark. The signal was measured using a
fluorescence microscope (magnification, ×100; cat. no. BX51;
Olympus Corporation).
Western blotting
Cardiac tissues were lysed using RIPA lysis buffer
(Beyotime Institute of Biotechnology). The protein concentration
was determined using a bicinchoninic acid protein assay kit
(Beyotime Institute of Biotechnology). Then, 20 ug of protein was
separated via 12% SDS-PAGE and transferred onto nitrocellulose
membranes. The membranes were first blocked with 5% non-fat milk
for 1 h at RT, and then incubated with specific primary antibodies
overnight at 4°C, including anti-TLR4 (1:1,000; cat. no. ab13556;
Abcam), anti-TLR2 (1:2,000; cat. no. sc-10739; Santa Cruz
Biotechnology, Inc.), anti-RP105 (1:1,000; cat. no. Sc-13592; Santa
Cruz Biotechnology, Inc.), anti-Bcl-2 (1:1,000; cat. no. ab196495;
Abcam), anti-cleaved caspase-3 (1:1,000; cat. no. ab2302; Abcam),
anti-MYD88 (1:1,000; cat. no. ab2064; Abcam) and anti-NF-κB
antibody (1:1,000; cat. no. Ab32536; Abcam). Membranes were then
incubated with horseradish peroxidase-conjugated secondary
antibodies (1:5,000; cat. nos. Ab97046 and Ab97057; Abcam) at RT
for 1 h. The bands were exposed with Pierce ECL western blotting
substrate (cat. no. 32106; Thermo Fisher Scientific, Inc). The
expression of GAPDH (1:3,000; cat. no. Ab181602; Abcam) was
detected to normalize the signals. The expression of proteins was
qualified using ImageJ software (1.47; National Institutes of
Health)
Evaluation of myocardial infarct
size
The myocardial infarct area was measured with the
triphenyltrazolium chloride (TTC) staining. After being frozen at
−80°C for 5 min, the hearts were sliced into five sections
(thickness, 2 mm) and incubated with 1.5% TTC at 37°C for 15 min.
Normal myocardial tissues stained red and the infarction region
stained white with TTC. The myocardial infarct size was calculated
using Image-Pro Plus 5.0 software (Media Cybernetics, Inc.).
Hematoxylin and eosin (H&E)
staining
Samples of cardiac tissue were fixed for 24 h at RT
with 4% buffered formalin, embedded in paraffin and then cut into
slices (thickness, 5 mm). Afterwards, the heart tissues were
stained with H&E for 5 min at room temperature and imaged with
a light microscope (magnification, ×200).
Indexes of blood
The blood of rats treated with 1 ml 10% KCl was
collected and centrifuged at 1,500 × g for 10 min at 4°C. The serum
was obtained and the levels of creatine kinase (CK), MB isoenzyme
of CK (CK-MB), and lactate dehydrogenase (LDH) was determined with
lactate dehydrogenase kit (cat. no. 2400837; Beijing Kemeidongya
Biotechnology Co., Ltd.) according to the manufacturer's
protocols.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the myocardial samples
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). RNA was reverse transcribed into cDNA using the
High-Capacity RNA-to cDNA kit (cat. no. 4387406; Thermo Fisher
Scientific, Inc.). RT-qPCR was conducted with an SYBR Green Master
Mix kit (Thermo Fisher Scientific, Inc.) on an ABI Prism 7500
system (Applied Biosystems; Thermo Fisher Scientific, Inc.). The
PCR conditions were set as follows: 2 min at 50°C, 95°C for 10 min,
then 40 cycles of 95°C for 30 sec and 60°C for 30 sec. The
expression of RP105 was normalized to that of GAPDH using the
2−ΔΔCq method (26).
The primers used were as follows: RP105, forward
5′-GAGCCTGAACCTGAGCTACA-3′, reverse 5′-GGAGGCAGTGGGAGAGATTT-3′;
GAPDH, forward 5′-ACAGCAACAGGGTGGTGGAC-3′, reverse
5′-TTTGAGGGTGCAGCGAACTT-3′.
Co-immunoprecipitation
Tissues were added into homogenizing microtubes
(OMNI International, Inc.) containing 1 ml home-made NP-40 lysis
buffer [In 1l of NP-40 lysis buffer: 30 ml of 5 M NaCl, 100 ml of
10% NP-40, 50 ml of 1 M Tris (pH 8.0) and 820 ml of H2O.
Stored at 4°C] and immediately homogenized using the
Precellys®24 Homogenizer (Bertin Instruments) at 4,000 ×
g for 1 min at 4°C. The homogenized mix was centrifuged at 10,000 ×
g for 15 min at 4°C and the clarified supernatant was collected.
The supernatant was pre-cleared by incubating with protein A beads
for 1 h at 4°C. Afterwards, lysate was incubated with anti-MD2
antibody (2 µg; cat. no. sc-80183; Santa Cruz Biotechnology, Inc.)
at 4°C overnight. IgG antibody (1:200; cat. no. sc-2025; Santa Cruz
Biotechnology, Inc.) was used as the negative control. Then,
Protein A agarose beads was added and incubated for 4 h at 4°C,
followed by washing twice with PBS on ice. The binding between MD2
and TLR2/TLR4 was detected by western blotting with anti-MD-2 and
anti-TLR2/4 antibodies (anti-TLR4: 1:1,000; cat. no. ab13556;
Abcam; and anti-TLR2; 1:2,000; cat. no. sc-10739; Santa Cruz
Biotechnology, Inc.).
Statistical analysis
Data are presented as mean ± SD. Statistical
significance was analyzed using one-way ANOVA followed by Tukey's
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Overexpression or knockdown of RP105
in rat cardiac muscle tissue
To explore the anti-inflammatory and
cardioprotective roles of RP105 in MIRI, the adenoviral vectors
expressing Ad-RP105-siRNA or Ad-EGFP-RP105 were injected into four
individual sites at the apex of each rat heart. The transfection
efficiency was identified by detecting the expression level of EGFP
in the Ad-EGFP-RP105 and Ad-EGFP-RP105-siRNA groups (Fig. 1A). Compared with the Ad-G-siRNA
group, both the mRNA and protein expression levels of RP105 in the
Ad-RP105-siRNA group were significantly decreased (Fig. 1B and C). Transfection with
Ad-EGFP-RP105 significantly increased the expression levels of
RP105 in the myocardial tissue compared with the I/R group.
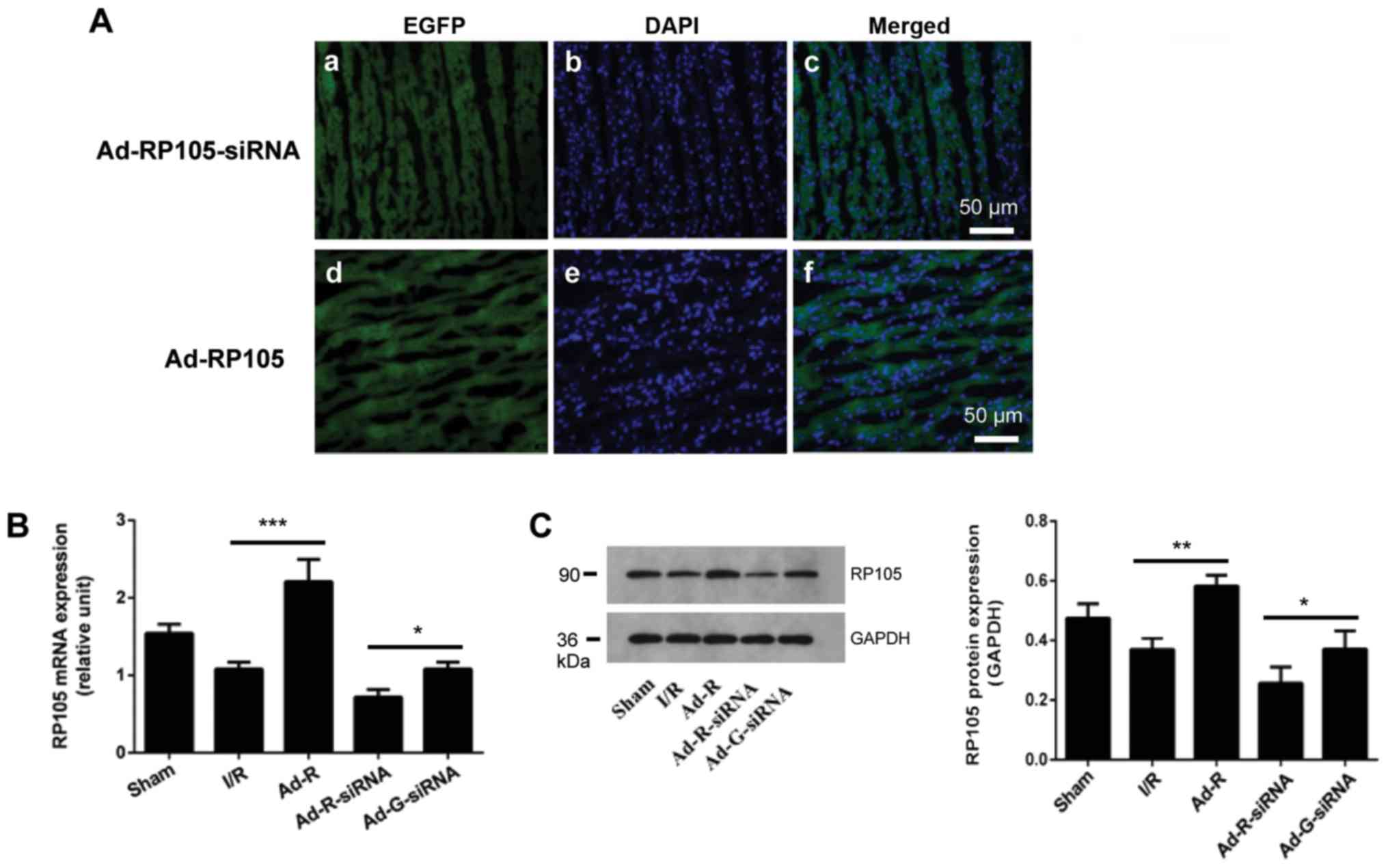 | Figure 1.Overexpression or knockdown of RP105
in rat cardiac muscle tissue. (A) Expression levels of EGFP, with
the indicated adenovirus gene injection for RP105 overexpression
and RP105-siRNA, and DAPI-labeled nuclei density of cardiomyocytes.
Scale bar, 50 µm. (B) mRNA expression of RP105 in rat cardiac
muscle tissues was detected by reverse transcription-quantitative
PCR after transfection with the indicated vector. (C) Protein
expression levels of RP105 in rat cardiac muscle tissue.
*P<0.05, **P<0.01, ***P<0.001. Ad, adenovirus; RP105,
radioprotective 105 kDa protein; siRNA, small interfering RNA;
EGFP, enhanced green fluorescent protein; sham, control sham group;
I/R, ischemic and reperfusion; Ad-R, Ad-RP105 group; Ad-R-siRNA,
Ad-RP105-siRNA group; Ad-G-siRNA, Ad-EGFP-siRNA group. |
Knockdown of RP105 promotes the
activity of serum myocardial enzymes during MIRI
The concentrations of serum myocardial enzymes
CK-MB, LDH and CK were increased after ischemia for 30 min followed
by reperfusion for 3 h (I/R group vs. sham group; Fig. 2A-C). Additionally, knockdown of
RP105 significantly increased the concentration of CK-MB, LDH and
CK in myocardium compared with the Ad-G-siRNA group (Fig. 2A-C). In contrast, overexpression of
RP105 significantly decreased the levels of CK-MB, LDH and CK in
the rat myocardium (Ad-R vs. I/R group; Fig. 2A-C). The present results suggested
that knockdown of RP105 enhanced the activity of myocardial enzymes
during MIRI in rats.
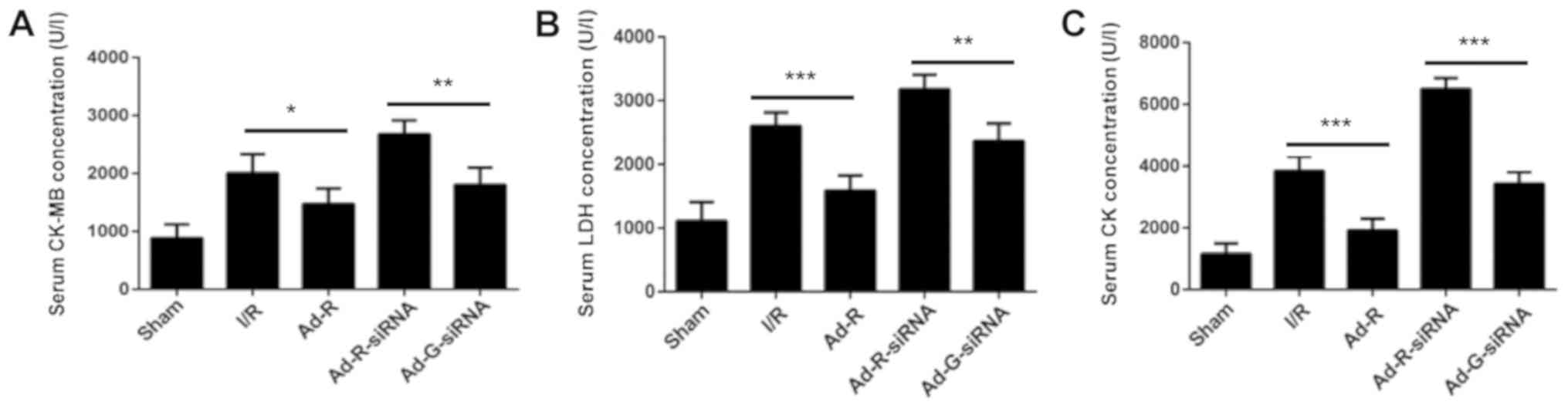 | Figure 2.Knockdown of RP105 promotes the
activity of serum myocardial enzymes during MIRI. Levels of serum
(A) CK-MB, (B) LDH and (C) CK in sham, I/R, Ad-R, Ad-R-siRNA and
Ad-G-siRNA groups. *P<0.05, **P<0.01, ***P<0.001. Ad,
adenovirus; RP105, radioprotective 105 kDa protein; siRNA, small
interfering RNA; EGFP, enhanced green fluorescent protein; CK,
creatine kinase; CK-MB, MB isoenzyme of CK; LDH, lactate
dehydrogenase; sham, control sham group; I/R, ischemic and
reperfusion; Ad-R, Ad-RP105 group; Ad-R-siRNA, Ad-RP105-siRNA
group; Ad-G-siRNA, Ad-EGFP-siRNA group. |
Silencing of RP105 increases
myocardial infarction during MIRI
To investigate whether the knockdown of RP105
aggravated myocardial damage during MIRI, the myocardial infarction
of each rat was evaluated by TTC staining. The myocardial
infarction area was significantly increased in the RP105 knockdown
group compared with the Ad-G-siRNA group (Fig. 3A and B). The present results
indicated that overexpression of RP105 suppressed myocardial
infarction in rats. Histopathological alterations in ischemic
myocardia followed by reperfusion were identified via H&E
staining (Fig. 3C). Only a few
myocardial infarcts were identified in the control group (Fig. 3A and B). However, obvious
myocardial degeneration and necrosis, myocardial fibers disorder,
interstitial edema and infiltration of inflammatory cells were
identified in the other experimental groups. Notably, these
phenotypes were more significant in RP105 knockdown group compared
with Ad-G-siRNA group (Fig. 3A and
B). Mild myocardial fiber disorder and swelling were observed
in RP105 overexpression group (Fig.
3C). The present results indicated the protective role of RP105
against myocardial infarction during MIRI.
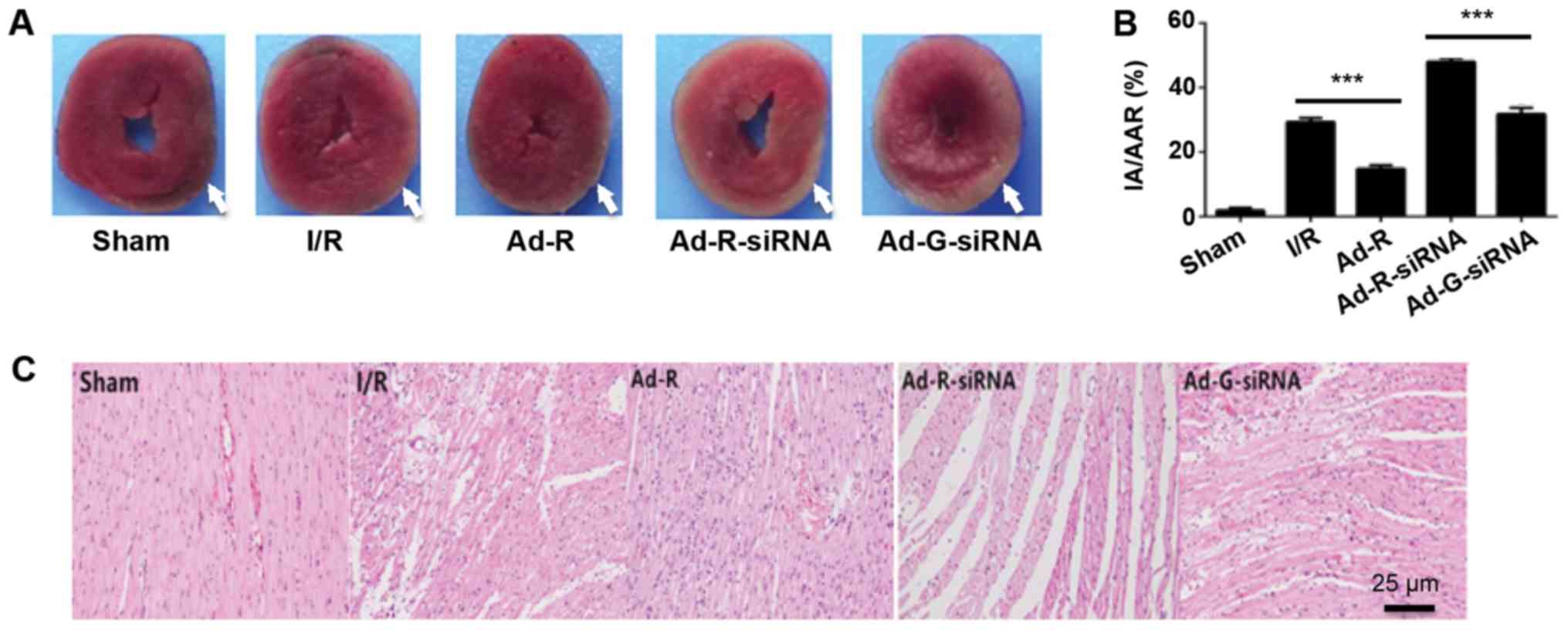 | Figure 3.Silencing of RP105 increases
myocardial infarction during MIRI. (A) Area of myocardial
infarction in each group was imaged and analyzed using Image-Pro
Plus 5. The injury areas were highlighted with white arrows. (B)
Statistical analysis of the myocardial infarction of each group.
(C) Hematoxylin and eosin staining of the myocardium tissue slices
in each group. Scale bar, 25 µm. ***P<0.001. Ad, adenovirus;
RP105, radioprotective 105 kDa protein; siRNA, small interfering
RNA; EGFP, enhanced green fluorescent protein; sham, control sham
group; I/R, ischemic and reperfusion; Ad-R, Ad-RP105 group;
Ad-R-siRNA, Ad-RP105-siRNA group; Ad-G-siRNA, Ad-EGFP-siRNA group;
IA, infarct area; AAR, area at risk. |
Knockdown of RP105 increases the
expression levels of TLR2 and TLR4 in MIRI
To investigate the involvement of the TLR2/4
signaling pathway in MIRI, myocardial tissues of rats were
collected after myocardial ischemia for 30 min and reperfusion for
3 h. The expression levels of TLR2 and TLR4 in rat myocardium were
detected via western blotting. The present results suggested that
compared with the Ad-G-siRNA group, the expression levels of TLR2
and TLR4 were significantly increased in the RP105 knockdown group
(Fig. 4A-C). However, both the
expression levels of TLR2 and TLR4 in the RP105 overexpression
group were significantly decreased compared with the I/R group
(Fig. 4A-C). Overexpression of
RP105 decreased the interaction between MD2 and TLR2/4, while
knockdown of RP105 increased the interaction of MD2 and TLR2/4
(Fig. 4C). Similarly, compared
with control groups, after knockdown of RP105, the expression
levels of inflammatory factors interleukin (IL-6) and tumor
necrosis factor (TNF)-α were significantly increased, but they were
decreased following overexpression of RP105 (Fig. 4D-F). The present results suggested
that RP105 may negatively regulate the TLR2/4 signaling pathway in
MIRI.
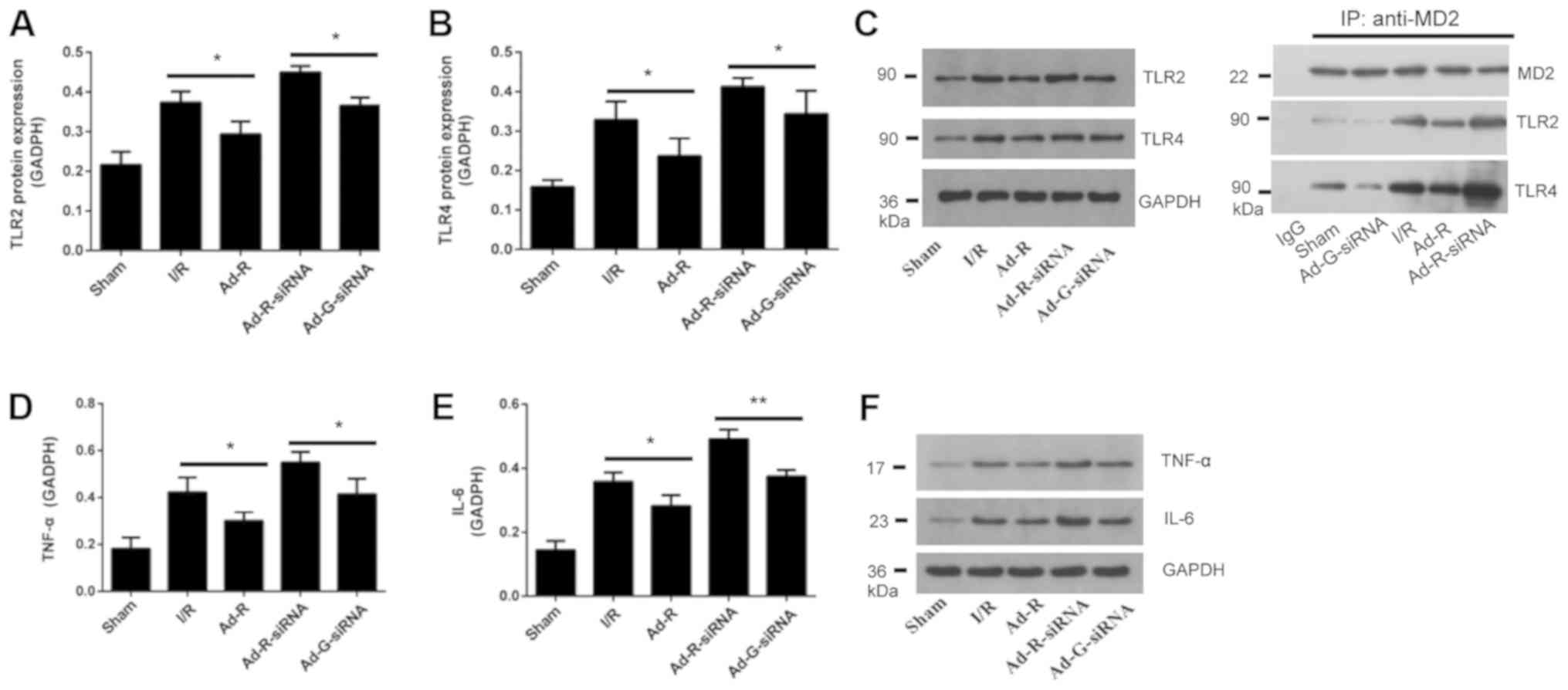 | Figure 4.Knockdown of RP105 increases the
expression levels of TLR2 and TLR4 in MIRI. Protein expression
levels of serum (A) TLR2 and (B) TLR4 in each group. (C) Western
blotting for the expression levels of TLR2 and TLR4 in each group.
The interactions of MD2 with TLR2 and TLR4 following overexpression
or knockdown of RP105 were investigated. Expression level of (D)
TNF-α and (E) IL-6 in each group. (F) Western blotting bands of
TNF-α and IL-6. *P<0.05, **P<0.01. Ad, adenovirus; RP105,
radioprotective 105 kDa protein; siRNA, small interfering RNA;
EGFP, enhanced green fluorescent protein; sham, control sham group;
I/R, ischemic and reperfusion; Ad-R, Ad-RP105 group; Ad-R-siRNA,
Ad-RP105-siRNA group; Ad-G-siRNA, Ad-EGFP-siRNA group; TLR2/4,
Toll-like receptor 2/4; TNF-α, tumor necrosis factor; IL-6,
interleukin 6; MD2, myeloid differentiation protein 2; IP,
immunoprecipitation. |
Silencing of RP105 increases
myocardial apoptosis
To detect whether knockdown of RP105 was associated
with apoptosis of myocardial cells, the expression levels of
apoptosis-related markers were investigated. The present results
suggested that the protein expression level of Bcl-2 in myocardial
tissue with overexpressed RP105 was significantly higher compared
with the I/R group, while knockdown of RP105 decreased the
expression level of Bcl-2 (Fig. 5A and
C). In addition, the protein expression level of cleaved
caspase-3 in RP105 overexpressed group was significantly decreased
compared with the control group (Fig.
5B and C). The expression level of cleaved caspase-3 with
RP-105 knockdown was significantly increased.
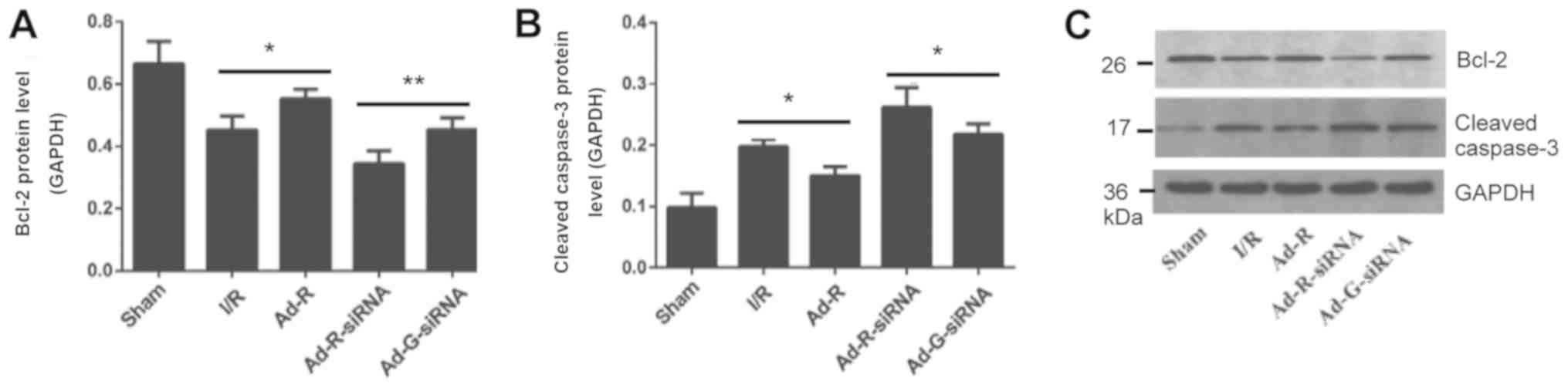 | Figure 5.Silencing of RP105 increases
myocardial apoptosis. Protein expression levels of serum (A) Bcl-2
and (B) cleaved caspase-3 in each group. (C) Western blotting bands
of Bcl-2 and cleaved caspase-3. *P<0.05, **P<0.01. Ad,
adenovirus; RP105, radioprotective 105 kDa protein; siRNA, small
interfering RNA; EGFP, enhanced green fluorescent protein; sham,
control sham group; I/R, ischemic and reperfusion; Ad-R, Ad-RP105
group; Ad-R-siRNA, Ad-RP105-siRNA group; Ad-G-siRNA, Ad-EGFP-siRNA
group. |
Knockdown of RP105 may regulate the
MyD88 and NF-κB signaling pathways
The expression levels of MyD88 and NF-κB in rat
myocardium following myocardial ischemia for 30 min and reperfusion
for 3 h were investigated. The protein expression levels of
inflammatory factors MyD88 and NF-κB were significantly increased
after knockdown of RP-105 compared with the control group (Fig. 6A-C). The protein expression levels
of MyD88 and NF-κB in the presence of RP-105 were significantly
reduced compared with the control groups. There was no significant
difference between the I/R group and the Ad-G-siRNA group (Fig. 6A-C).
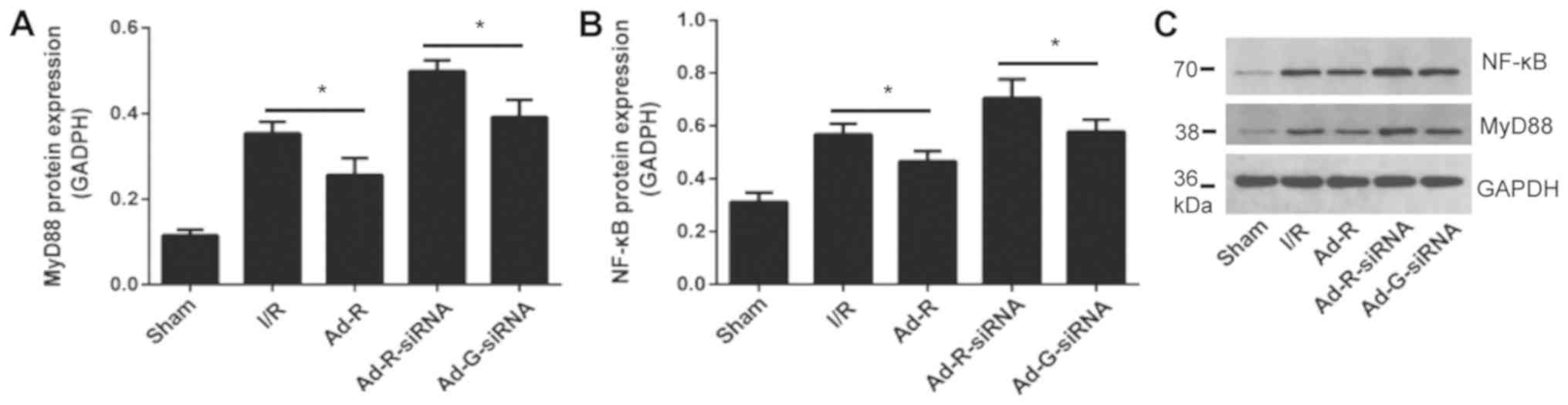 | Figure 6.Knockdown of RP105 regulates the
MyD88 signaling pathway. Protein expression levels of serum (A)
MyD88 and (B) NF-κB in each group. (C) Western blotting bands of
MyD88 and NF-κB in each group. *P<0.05. Ad, adenovirus; RP105,
radioprotective 105 kDa protein; siRNA, small interfering RNA;
EGFP, enhanced green fluorescent protein; sham, control sham group;
I/R, ischemic and reperfusion; Ad-R, Ad-RP105 group; Ad-R-siRNA,
Ad-RP105-siRNA group; Ad-G-siRNA, Ad-EGFP-siRNA group; MyD88,
myeloid differentiation factor 88. |
Discussion
Previous studies have shown that the prevention and
treatment of MIRI depends on the following features: i) Simply
targeting the protective gene or harmful gene; ii) interfering with
the expression of a single gene to play a comprehensive role in
multiple pathogenic processes; or iii) modulating the expression of
target genes via non-coding RNAs (27,28).
Although these strategies provide novel ways to alleviate MIRI, the
clinical effect is still unsatisfactory (22,24,29–31).
This may be due to the following reasons: i) MIRI results from the
imbalance of both protective genes and harmful genes, and therefore
unilateral intervention has limited impact; ii) both upstream and
downstream pathways regulate MIRI, meaning targeting a single
factor cannot fully control the pathological process; and iii)
non-coding RNAs are involved in the progression of MIRI (32–34).
Previous studies have shown that serine/threonine
kinase receptor-associated proteins regulated cell proliferation
and apoptosis, and mediated the immune response to pathogens via
the TLR2/4 signaling pathway (35–37).
Our previous studies showed that RP105 plays a negative role in the
TLR4 signaling pathway (38,39).
TLR4 recognizes, and is activated by, bacterial lipopolysaccharide
(LPS) via the adaptor protein MD2, which induces the receptor
heterodimer LPS/MD2/TLR4 on the cell membrane (40–42).
The formation of the heterodimer initiates the intracellular
TLR4/MyD88/NF-κB signaling cascade. The recruitment of MyD88
promotes the recruitment of secondary messengers and enzymes to
form protein complexes, such as p38, c-Jun N-terminal kinase and
extracellular-signal-regulated kinases 1/2 (43). The following result is the
activation of NF-κB and the transcription of target genes,
including TNF-α, IL-1β and IL-18 (25). Additionally, with the activation of
TLR4, TRIF-related adaptor protein promotes conformational changes
in TRIF, and recruits TNF receptor associated factor 3 (TRAF3) and
TRAF6, eventually leading to the nuclear translocation of NF-κB and
AP-1 (44). In the present study,
silencing of RP-105 triggered apoptotic and inflammatory-related
pathways in rats, suggesting a negative role for RP-105 in the
inflammatory response. The present results suggested that RP105 may
be utilized as an inhibitor of the TLR4 inflammatory signaling
pathway to treat MIRI.
Numerous signaling pathways are involved in the
remodeling of the cardiovasculature (45,46).
Vascular and inflammatory cells-derived matrix metalloproteinases
and cysteine proteases play key roles in cardiovascular remodeling
and atherosclerotic plaque growth in human and animal models
(47–50). TLR2 has been found to bind more
ligands than other members of the TLR family (51). An increase in TLR2 ligands
significantly activates the PI3K/Akt pathway and protects
myocardium from ischemic injury, which indicates the
cardioprotective role of TLR2 in ischemic injury (52). RP105 is associated with the TLR2
signaling pathway in mediating inflammatory responses (53). A previous study showed that
Staphylococcus aureus activates RP105 in macrophages to
induce the accumulation of TLR2 (54). The increased TLR2 by inflammatory
cytokines via MyD88 leads to infection resistance in rats (54). After Mycobacterium
tuberculosis infection, RP105 interacts with TLR2 and modulates
the inflammatory response (55).
TLR2 forms a complex with TLR1 and TLR6, which identify bacterial
lipoproteins by recruiting the TIR domain-containing adaptor
protein and MyD88 (56). RP105/MD1
interacts with TLR2 to activate NF-κB and MAPK in macrophages,
which promote the release of inflammatory factors (53). However, excessive inhibition of
TLR2 may result in unfavorable remodeling in the cardiac tissue
(57). Therefore, inflammation may
be a double-edged sword, and a balance between regulating the
infarct size and the healing of damaged myocardium needs to be
investigated (58).
Acting as a homologue of TLR, RP105 promotes the
response of TLR2 ligand receptor by forming a heterologous complex
and negatively regulates TLR4 signaling in macrophages (53). RP105 is a possible factor in the
immune response (53); however,
whether RP105 interferes with TLR2/4 and affects the progression of
MIRI is not fully known. RP105 was reported as a critical regulator
of TLR4 to alleviate MIRI by regulating the activation of the
TLR4/Myd88 or TLR4/TRIF/IRF3 signaling pathway (17,38).
Moreover, an equilibrium state of the RP105-TLR2/4 signaling
pathway is maintained under normal cardiomyocytes, whereas MIRI
causes the imbalance of the RP105-TLR2/4 signaling pathway
(59). The present results
suggested the involvement of RP105 in the myocardium, which
indicated that TLR2 and TLR4 were key upstream regulators of ligand
receptor signaling in modulating the process of MIRI.
The main limitation of this study is that the RP105
knockdown experiments were performed only in rats, rather than a
higher primate species. However, using primate models is difficult
for functional studies of RP105 in MIRI due to the lack of
available drugs targeting RP105.
Various diseases caused by genetic mutations such as
hypertrophic cardiomyopathy and restrictive cardiomyopathy still
lack effective treatment strategies. The present results suggested
RP105 targeted both TLR2 and TLR4, which indicated RP105 may be a
promising candidate for the prevention and treatment of MIRI.
Acknowledgements
Not applicable.
Funding
The present study was supported by The National
Natural Science Foundation of China (grant nos. 81402568, 81670333
and 81470387) and Hubei Province's Outstanding Medical Academic
Leader Program and Technology Support Program of Hubei Province
(grant no. 2015BKA340).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WLH and JuY designed the study. WLH performed the
majority of experiments. JiY performed the H&E staining and
CO-IP assays. CH analyzed the data. JuY wrote the manuscript. All
authors contributed to the revision of the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The animal experiments were approved by the Animal
Care and Use Department of China Three Gorges University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Dalen JE, Alpert JS, Goldberg RJ and
Weinstein RS: The epidemic of the 20(th) century: Coronary heart
disease. Am J Med. 127:807–812. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Aggarwal B and Menon V: Recent advances in
treatment of acute coronary syndromes. F1000Prime Rep. 5:562013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Alexander W: American academy of
dermatology and american college of cardiology. P T. 39:370–374.
2014.PubMed/NCBI
|
|
4
|
Levitsky S: Protecting the myocardial cell
during coronary revascularization. The William W. L. Glenn lecture.
Circulation. 114 (Suppl 1):I339–I343. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fang Y and Hu J: Toll-like receptor and
its roles in myocardial ischemic/reperfusion injury. Med Sci Monit.
17:RA100–RA109. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Akira S, Uematsu S and Takeuchi O:
Pathogen recognition and innate immunity. Cell. 124:783–801. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Luo M, Yan D, Sun Q, Tao J, Xu L, Sun H
and Zhao H: Ginsenoside Rg1 attenuates cardiomyocyte apoptosis and
inflammation via the TLR4/NF-kB/NLRP3 pathway. J Cell Biochem. Nov
11–2019.(Epub ahead of print).
|
|
8
|
Akbarshahi H, Axelsson JB, Said K,
Malmström A, Fischer H and Andersson R: TLR4 dependent heparan
sulphate-induced pancreatic inflammatory response is IRF3-mediated.
J Transl Med. 9:2192011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen PG, Guan YJ, Zha GM, Jiao XQ, Zhu HS,
Zhang CY, Wang YY and Li HP: Swine IRF3/IRF7 attenuates
inflammatory responses through TLR4 signaling pathway. Oncotarget.
8:61958–61968. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang Y, Chen L, Tian Z, Shen X, Wang X, Wu
H, Zou J and Liang J: CRISPR-Cas9 mediated gene knockout in human
coronary artery endothelial cells reveals a pro-inflammatory role
of TLR2. Cell Biol Int. 42:187–193. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ohto U, Miyake K and Shimizu T: Crystal
structures of mouse and human RP105/MD-1 complexes reveal unique
dimer organization of the toll-like receptor family. J Mol Biol.
413:815–825. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Miyake K, Yamashita Y, Hitoshi Y, Takatsu
K and Kimoto M: Murine B cell proliferation and protection from
apoptosis with an antibody against a 105-kD molecule:
Unresponsiveness of X-linked immunodeficient B cells. J Exp Med.
180:1217–1224. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Selimovic D, Hassan M, Haikel Y and Hengge
UR: Taxol-induced mitochondrial stress in melanoma cells is
mediated by activation of c-Jun N-terminal kinase (JNK) and p38
pathways via uncoupling protein 2. Cell Signal. 20:311–322. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Karper JC, Ewing MM, de Vries MR, de Jager
SC, Peters EA, de Boer HC, van Zonneveld AJ, Kuiper J, Huizinga EG,
Brondijk TH, et al: TLR accessory molecule RP105 (CD180) is
involved in post-interventional vascular remodeling and soluble
RP105 modulates neointima formation. PLoS One. 8:e679232013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wezel A, van der Velden D, Maassen JM,
Lagraauw HM, de Vries MR, Karper JC, Kuiper J, Bot I and Quax PH:
RP105 deficiency attenuates early atherosclerosis via decreased
monocyte influx in a CCR2 dependent manner. Atherosclerosis.
238:132–139. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang H, Wang H, Ju Z, Ragab AA, Lundbäck
P, Long W, Valdes-Ferrer SI, He M, Pribis JP, Li J, et al: MD-2 is
required for disulfide HMGB1-dependent TLR4 signaling. J Exp Med.
212:5–14. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li X, Yang J, Yang J, Dong W, Li S, Wu H
and Li L: RP105 protects against myocardial ischemia-reperfusion
injury via suppressing TLR4 signaling pathways in rat model. Exp
Mol Pathol. 100:281–286. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang J, Yang C, Yang J, Ding J, Li X, Yu
Q, Guo X, Fan Z and Wang H: RP105 alleviates myocardial ischemia
reperfusion injury via inhibiting TLR4/TRIF signaling pathways. Int
J Mol Med. 41:3287–3295. 2018.PubMed/NCBI
|
|
19
|
Liu QS, Cheng ZW, Xiong JG, Cheng S, He XF
and Li XC: Erythropoietin pretreatment exerts anti-inflammatory
effects in hepatic ischemia/reperfusion-injured rats via
suppression of the TLR2/NF-kappaB pathway. Transplant Proc.
47:283–289. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pope MR and Fleming SD: TLR2 modulates
antibodies required for intestinal ischemia/reperfusion-induced
damage and inflammation. J Immunol. 194:1190–1198. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ulbrich F, Lerach T, Biermann J, Kaufmann
KB, Lagreze WA, Buerkle H, Loop T and Goebel U: Argon mediates
protection by interleukin-8 suppression via a
TLR2/TLR4/STAT3/NF-kappaB pathway in a model of apoptosis in
neuroblastoma cells in vitro and following ischemia-reperfusion
injury in rat retina in vivo. J Neurochem. 138:859–873. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang J, Chen L, Yang J, Ding J, Li S, Wu
H, Zhang J, Fan Z, Dong W and Li X: MicroRNA-22 targeting CBP
protects against myocardial ischemia-reperfusion injury through
anti-apoptosis in rats. Mol Biol Rep. 41:555–561. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang HB and Yang J, Ding JW, Chen LH, Li
S, Liu XW, Yang CJ, Fan ZX and Yang J: RNAi-mediated
down-regulation of CD47 protects against
ischemia/reperfusion-induced myocardial damage via activation of
eNOS in a rat model. Cell Physiol Biochem. 40:1163–1174. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang B, Zhong S, Zheng F, Zhang Y, Gao F,
Chen Y, Lu B, Xu H and Shi G: N-n-butyl haloperidol iodide protects
cardiomyocytes against hypoxia/reoxygenation injury by inhibiting
autophagy. Oncotarget. 6:24709–24721. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang X, Du Q, Yang Y, Wang J, Dou S, Liu
C and Duan J: The protective effect of Luteolin on myocardial
ischemia/reperfusion (I/R) injury through TLR4/NF-kappaB/NLRP3
inflammasome pathway. Biomed Pharmacother. 91:1042–1052. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu NB, Wu M, Chen C, Fujino M, Huang JS,
Zhu P and Li XK: Novel molecular targets participating in
myocardial ischemia-reperfusion injury and cardioprotection.
Cardiol Res Pract. 2019:69351472019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Davidson SM, Ferdinandy P, Andreadou I,
Bøtker HE, Heusch G, Ibáñez B, Ovize M, Schulz R, Yellon DM,
Hausenloy DJ, et al: Multitarget strategies to reduce myocardial
ischemia/reperfusion injury: JACC review topic of the week. J Am
Coll Cardiol. 73:89–99. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Quiat D and Olson EN: MicroRNAs in
cardiovascular disease: From pathogenesis to prevention and
treatment. J Clin Invest. 123:11–18. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang JX, Zhang XJ, Li Q, Wang K, Wang Y,
Jiao JQ, Feng C, Teng S, Zhou LY, Gong Y, et al: MicroRNA-103/107
regulate programmed necrosis and myocardial ischemia/reperfusion
injury through targeting FADD. Circ Res. 117:352–363. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang CJ, Yang J, Yang J and Fan ZX:
Radioprotective 105kDa protein (RP105): Is a critical therapeutic
target for alleviating ischemia reperfusion induced myocardial
damage via TLR4 signaling pathway. Int J Cardiol. 222:1069–1070.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Eltzschig HK and Eckle T: Ischemia and
reperfusion-from mechanism to translation. Nat Med. 17:1391–1401.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Joshi S, Wei J and Bishopric NH: A cardiac
myocyte-restricted Lin28/let-7 regulatory axis promotes
hypoxia-mediated apoptosis by inducing the AKT signaling suppressor
PIK3IP1. Biochim Biophys Acta. 1862:240–251. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sun Y, Ye L, Jiang C, Jiang J, Hong H and
Qiu L: Over-expression of HSPA12B protects mice against myocardium
ischemic/reperfusion injury through a PPARgamma-dependent
PI3K/Akt/eNOS pathway. Am J Transl Res. 7:2724–2737.
2015.PubMed/NCBI
|
|
35
|
Huh HD, Ra EA, Lee TA, Kang S, Park A, Lee
E, Choi JL, Jang E, Lee JE, Lee S and Park B: STRAP acts as a
scaffolding protein in controlling the TLR2/4 signaling pathway.
Sci Rep. 6:388492016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Takeda K and Akira S: TLR signaling
pathways. Semin Immunol. 16:3–9. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dunne A, Carpenter S, Brikos C, Gray P,
Strelow A, Wesche H, Morrice N and O'Neill LA: IRAK1 and IRAK4
promote phosphorylation, ubiquitination, and degradation of MyD88
adaptor-like (Mal). J Biol Chem. 291:248022016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Guo X, Jiang H, Yang J, Chen J, Yang J,
Ding JW, Li S, Wu H and Ding HS: Radioprotective 105 kDa protein
attenuates ischemia/reperfusion-induced myocardial apoptosis and
autophagy by inhibiting the activation of the TLR4/NF-kappaB
signaling pathway in rats. Int J Mol Med. 38:885–893. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yang J, Guo X, Yang J, Ding JW, Li S, Yang
R, Fan ZX and Yang CJ: RP105 protects against apoptosis in
ischemia/reperfusion-induced myocardial damage in rats by
suppressing TLR4-mediated signaling pathways. Cell Physiol Biochem.
36:2137–2148. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nagai Y, Akashi S, Nagafuku M, Ogata M,
Iwakura Y, Akira S, Kitamura T, Kosugi A, Kimoto M and Miyake K:
Essential role of MD-2 in LPS responsiveness and TLR4 distribution.
Nat Immunol. 3:667–672. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
41
|
Brandl K, Glück T, Hartmann P, Salzberger
B and Falk W.: A designed TLR4/MD-2 complex to capture LPS. J
Endotoxin Res. 11:197–206. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Roh E, Lee HS, Kwak JA, Hong JT, Nam SY,
Jung SH, Lee JY, Kim ND, Han SB and Kim Y: MD-2 as the target of
nonlipid chalcone in the inhibition of endotoxin LPS-induced TLR4
activity. J Infect Dis. 203:1012–1020. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lee SM, Hutchinson M and Saint DA: The
role of Toll-like receptor 4 (TLR4) in cardiac
ischaemic-reperfusion injury, cardioprotection and preconditioning.
Clin Exp Pharmacol Physiol. 43:864–871. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Piao W, Ru LW, Piepenbrink KH, Sundberg
EJ, Vogel SN and Toshchakov VY: Recruitment of TLR adapter TRIF to
TLR4 signaling complex is mediated by the second helical region of
TRIF TIR domain. Proc Natl Acad Sci USA. 110:19036–19041. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ayme-Dietrich E, Aubertin-Kirch G,
Maroteaux L and Monassier L: Cardiovascular remodeling and the
peripheral serotonergic system. Arch Cardiovasc Dis. 110:51–59.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ayme-Dietrich E, Marzak H, Lawson R, Mokni
W, Wendling O, Combe R, Becker J, El Fertak L, Champy MF, Matz R,
et al: Contribution of serotonin to cardiac remodeling associated
with hypertensive diastolic ventricular dysfunction in rats. J
Hypertens. 33:2310–2321. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lei Y, Yang G, Hu L, Piao L, Inoue A,
Jiang H, Sasaki T, Zhao G, Yisireyili M, Yu C, et al: Increased
dipeptidyl peptidase-4 accelerates diet-related vascular aging and
atherosclerosis in ApoE-deficient mice under chronic stress. Int J
Cardiol. 243:413–420. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yang G, Lei Y, Inoue A, Piao L, Hu L,
Jiang H, Sasaki T, Wu H, Xu W, Yu C, et al: Exenatide mitigated
diet-induced vascular aging and atherosclerotic plaque growth in
ApoE-deficient mice under chronic stress. Atherosclerosis.
264:1–10. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cheng XW, Huang Z, Kuzuya M, Okumura K and
Murohara T: Cysteine protease cathepsins in atherosclerosis-based
vascular disease and its complications. Hypertension. 58:978–986.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Cheng XW, Shi GP, Kuzuya M, Sasaki T,
Okumura K and Murohara T: Role for cysteine protease cathepsins in
heart disease: Focus on biology and mechanisms with clinical
implication. Circulation. 125:1551–1562. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Selmi C: Autoimmunity in 2016. Clin Rev
Allergy Immunol. 53:126–139. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ha T, Hu Y, Liu L, Lu C, McMullen JR,
Kelley J, Kao RL, Williams DL, Gao X and Li C: TLR2 ligands induce
cardioprotection against ischaemia/reperfusion injury through a
PI3K/Akt-dependent mechanism. Cardiovasc Res. 87:694–703. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Liu B, Zhang N, Liu Z, Fu Y, Feng S, Wang
S, Cao Y, Li D, Liang D, Li F, et al: RP105 involved in activation
of mouse macrophages via TLR2 and TLR4 signaling. Mol Cell Biochem.
378:183–193. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Liu B, Fu Y, Feng S, Zhang X, Liu Z, Cao
Y, Li D, Liang D, Li F, Zhang N and Yang Z: Involvement of RP105
and toll-like receptors in the activation of mouse peritoneal
macrophages by Staphylococcus aureus. Scand J Immunol. 78:8–16.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Blumenthal A, Kobayashi T, Pierini LM,
Banaei N, Ernst JD, Miyake K and Ehrt S: RP105 facilitates
macrophage activation by Mycobacterium tuberculosis lipoproteins.
Cell Host Microbe. 5:35–46. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Frazao JB, Errante PR and Condino-Neto A:
Toll-like receptors' pathway disturbances are associated with
increased susceptibility to infections in humans. Arch Immunol Ther
Exp (Warsz). 61:427–443. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Mersmann J, Habeck K, Latsch K, Zimmermann
R, Jacoby C, Fischer JW, Hartmann C, Schrader J, Kirschning CJ and
Zacharowski K: Left ventricular dilation in toll-like receptor 2
deficient mice after myocardial ischemia/reperfusion through
defective scar formation. Basic Res Cardiol. 106:89–98. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Lepper PM and Bals R: On the edge:
Targeting Toll-like receptor 2 in ischemia/reperfusion injury. Circ
Cardiovasc Interv. 5:146–149. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Divanovic S, Trompette A, Petiniot LK,
Allen JL, Flick LM, Belkaid Y, Madan R, Haky JJ and Karp CL:
Regulation of TLR4 signaling and the host interface with pathogens
and danger: The role of RP105. J Leukoc Biol. 82:265–271. 2007.
View Article : Google Scholar : PubMed/NCBI
|














